
В последние годы в популяциях всех позвоночных обнаруживаются значительные межиндивидуальные различия в поведении (и психофизиологических особенностях, детерминирующих это поведение), которые настолько множественны, разноплановы, и являются настолько устойчивой характеристикой индивида, что этологи стали говорить об «индивидуальности» (personality). Соответственно, в плане реализации видоспецифического поведения популяция не однородна, а полиморфна, особи реализуют множество поведенческих типов (behavioural types), выбор которых характеризует индивидуальность, а набор – популяцию.
Введение
Какие переменные структурируют разнообразие поведенческих типов, я писал на примере синиц, грызунов, муравьёв и рыб; важно, что в работах С.Будаева было показано, что у всех классов позвоночных от рыб до человека, разнообразие психофизиологии особей, поддерживающее непосредственно наблюдаемую мозаику поведенческих типов, может описываться одной и той же двумерной шкалой, впервые предложенной психологами для человека, где по одной оси «интравертность-экстравертность», по другой — «нейротизм и тревожность». Естественно, соответствующее тестирование у рыб и рептилий требует иной постановки опыта, чем у нас; так как животных не спросишь, их надо ставить в проблемную ситуацию, специфически важную именно для данного вида, и дальше смотреть на индивидуальную реакцию на проблему – будет ли зверь пугаться, тормозить, либо отважно бросится разрешать ситуацию. См. ниже описание тестирования рыбок при определении поведенческих типов.
В популяциях разных видов позвоночных множественность поведенческих типов описана по таким параметрам, как смелость (готовность рисковать в незнакомой ситуации и/или сближаться с потенциальной опасностью, чтобы прояснить ситуацию), агрессивность, уровень неофобии и уровень исследовательской активности. Понятно, что это лишь отдельные измерения, по которым конструируется поведенческий тип, у животных имеющий смысл «индивидуальности» у человека. Животная personality образуется комбинацией «координат» по нескольким параметрам, 2-3-м как минимум, что хорошо видно у рыб и синиц, так что в каждом малом сообществе, за которым наблюдает этолог, включающем порядка 10-20 индивидов, каждый может своим поведенческим типом отличаться от всех остальных.
Неожиданное открытие столь высокой «персональности» особей в популяции (тогда как отцы-основатели этологии полагали, что реализация видового инстинкта у всех особей единообразна, они могут отличаться лишь в параметрах, связанных с обучением и рассудочной деятельностью – «как люди») ставит вопрос об эволюционном смысле и адаптивном значении данного полиморфизма. Если исследование достаточно тщательно, практически каждому из элементов набора поведенческих типов удаётся найти свой «адаптивный максимум» — ту область характеристик экологической и социальной среды, при которых его обладатели имеют явное преимущество над остальными, и у разных поведенческих типов эти области чаще всего взаимно-дополнительных друг к другу, как в случае с альтернативными стратегиями, носители которых сортируются внутри популяции.
То есть биологический смысл этого полиморфизма отыскать достаточно просто; сложней выбрать модель факторов эволюции, конституирующих данный полиморфизм и устойчиво воспроизводящих его в популяциях вопреки разного рода внешним и внутренним «шумам». В числе «шумов», преодоление которых – необходимое условие всякой популяционной устойчивости а, значит и жизнеспособности вида, входят 3 вида неопределённостей: демографическая, стохастическая и средовая. Первая порождается случайными событиями, связанными с выживанием и воспроизводством особей, вторая связана с природными катастрофами, вроде наводнений, пожаров, засух и пр., время от времени непредсказуемым образом разрушающих часть местообитаний вида с непредсказуемой периодичностью. Третья возникает за счёт случайных (по крайней мере, непредсказуемых для вида на основе его адаптаций) перемен погоды, обилия пищи и воздействия со стороны популяций, конкурентов, хищников, паразитов и пр. [1].
Вспомнив противопоставление «левого и правого крыла дарвинизма», теории стабилизирующего отбора (+ продолжающей его ЭТЭ) и СТЭ, мыслимы 2 альтернативных объяснения относительно механизмов, поддерживающих и воспроизводящих полиморфизм поведенческих типов в популяциях. Первое (с «правого фланга») – «гены управляют поведением, подставляя его под отбор»: разные поведенческие типы связаны с разными генными комплексами, селективная отшлифовка их есть фактически отбор генов, устанавливающий нужные соотношения между генами, определяющими разные варианты поведенческих типов.
Второй, с «левого фланга»: каждый из поведенческих типов селектируется на максимальную устойчивость воспроизводства «в своей среде», в тех условиях, когда он нужен и выгоден, и по ходу этой селекции всё больше эмансипируется от разнообразия генного пула популяции и от генотипа конкретной особи, превращается в эквифинальную стадию. В этом случае в онтогенезе выраженность поведенческих типов нарастает постепенно, и когда последние обретут полную законченность и максимально дифференцируются друг от друга, какая либо зависимость их от генотипа не обнаруживается. Здесь отбор стабилизирует соответствующие эквифинальные стадии, а изменения на уровне генов связаны с соответствующими элементарными эволюционными явлениями не прямо, а косвенно (п.4).
При верности этого объяснения гены «не управляют поведением», а лишь создают для него «сырьё и комплектующие»: как именно «собирается» дефинитивное поведение из этих «запчастей», предоставленных биологией и генетикой индивида, определяется состоянием среды, в первую очередь социальной ситуацией в сообществе, куда входит особь, т.е. формообразующее влияние на разнообразие поведенческих типов оказывают не атомарные, а реляционные характеристики особи, связанные с её положением в системе более высокого уровня, в данном случае в сообществе и популяции.
Исследование карпозубой рыбки
Недавно нашёл интересную работу M.Edenbrow & D.P.Croft (2011), выполненную на карпозубой рыбке Kryptolebias marmoratus (жравшей гуппей вот в этом исследовании), результаты которой решительно поддерживают 2-ю точку зрения. Было показано, что индивидуальность и характер рыб не зависят от генов, а последние «не управляют поведением», как минимум когда оно полностью сформировано на эквифинальной стадии. Даже если разные генотипы отличаются по возрасту первого размножения и репродуктивному успеху, эти различия не были связаны с типами поведения как до, так и после полового созревания.
Надо сказать, что оная рыбка – идеальный объект для данной работы. Живёт она в в мангровых болотах Северной и Южной Америки, легко переносит пересыхание водоёма, прячась в сколько-нибудь влажных местах и поглощая кислород всей поверхностью кожи. При этом она – самооплодотворяющийся гермафродит, причём склонный к самооплодотворению. Периодически в популяции появляются самцы, за счёт которых идёт перемешивание генов, но это происходит достаточно редко, и население одного болота на протяжении нескольких поколений представлено сериями клонов, никак не обменивающимися генетическим материалом. Соответственно, от этих клонов легко взять отводки, и изучить в лаборатории их онтогенез + становление разнообразия поведенческих типов по ходу последнего, зависит оно или не зависит от генотипа. Так и сделали; прослеживали 120 особей 20 генотипов.
Разнообразие поведенческих типов описывали в двумерной шкале координат, как некое сочетание разной степени смелости и склонности к исследовательскому поведению у рыб, поставленных в проблемную ситуацию. Поведенческий тип измеряли для 5-ти разных точек онтогенеза, с 30-ти дневным интервалом, начиная со 2-го дня после вылупления. В день вылупления случайным образом определяли порядок тестирования каждого индивида, и этот порядок выдерживался в течение всего исследования. Сначала рыбу тестировали на склонность к исследованию, а потом на смелость (для каждого из возрастов). Опыты записывались на видеокамеру. Для отслеживания движений рыбы использовали программу Ethovision. Вода в сосудах менялась перед каждым опытом. Чтобы ограничить структурную новизну в тестах на смелость, из сосуда для содержания рыб делали «арену» для определения смелости, удаляя гравий и добавляя удаляемые структурные компоненты (сетку и белый гравиевый диск) за 24 часа до тестирования смелости.
Тестирование исследовательской активности производились с использованием нового белого лабиринта, который увеличивали в размерах по мере роста рыбы. Чтобы поддерживать постоянство структуры лабиринта, генерируя в то же время новизну, меняли место входа (зону акклиматизации) для каждой последующей серии измерений. Поле было поделено на 24 равных зоны (исключая вход для акклиматизации). Рыбы помещались в тестовый сосуд через вход (где была редкая подложка из гравия), который был окружён белым непрозрачным барьером. Через 5 мин акклиматизации барьер снимался. Поведение регистрировалось для периода в 10 мин, во время которых фиксировали расстояние, на которое перемещалась рыба (мм), среднюю скорость (мм/c), продолжительность передвижений (с) и число посещённых зон. В конце каждого испытания индивиды возвращались в макет для тестирования смелости, устроенный в сосуде для содержания как минимум за 3 ч до анализа смелости.
Макет для анализа смелости делился на две секции по длинной стороне. Отгороженная зона была сконструирована так, что позволяла опускать в неё грузы (чтобы симулировать приближение хищника с воздуха). После введения рыбы на «арену» для измерения смелости выдерживался 5 мин период акклиматизации, после чего груз, подвешенный в 13 см над отгороженной зоной, вводился на «арену» с погружением в воду, имитируя рассечение поверхности воды хищником с воздуха. После это фиксировалось время, проходящее до первого движения (передвижение более чем на 1 корпус), дистанция перемещения (мм), и средняя скорость (мм/с) за пятиминутный период наблюдения. В конце опыта индивиды возвращались в свой сосуд, содержащий гравий. См.схему экспериментальной установки: а – арена для оценки развития исследовательского поведения, b – арена для оценки уровня смелости/агрессивности.
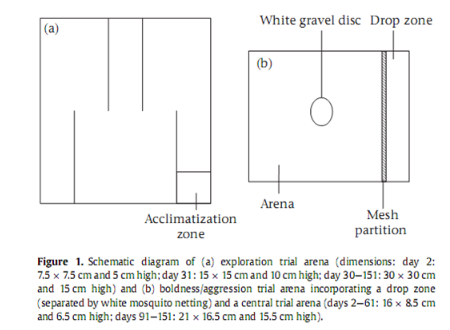
Схема
Далее проводили анализ главных компонент, чтобы понять, насколько разнообразие поведенческих характеристик, измеренных в данных экспериментах, может описываться в 2-мерном пространстве компонент, интерпретируемых как «смелость» и «исследовательская активность». Оказалось, что да, может, и данные 2 компоненты описывают значительный %% дисперсии наблюдаемого поведения, причём они равно применимы ко всей выборке (табл.1-2, стр.3-4).
Что получилось?
Прежде всего, как и положено эквифинальной стадии, выраженность всех типов поведения у всех 20 генотипов возрастает во время раннего онтогенеза и достигает асимптоты примерно к половому созреванию (см. онтогенетические траектории развития «смелости» и «исследовательской способности» на популяционном уровне, а, и уровне отдельных генотипов, b). В начале периода развития все генотипы показывают достаточно высокий уровень пластичности поведения (и разные генотипы значимо различаются по уровню пластичности между собой), но с 61-го дня поведенческие типы «кристаллизуются» — появляется сильная значимая положительная корреляция между исследовательской активностью и смелостью.
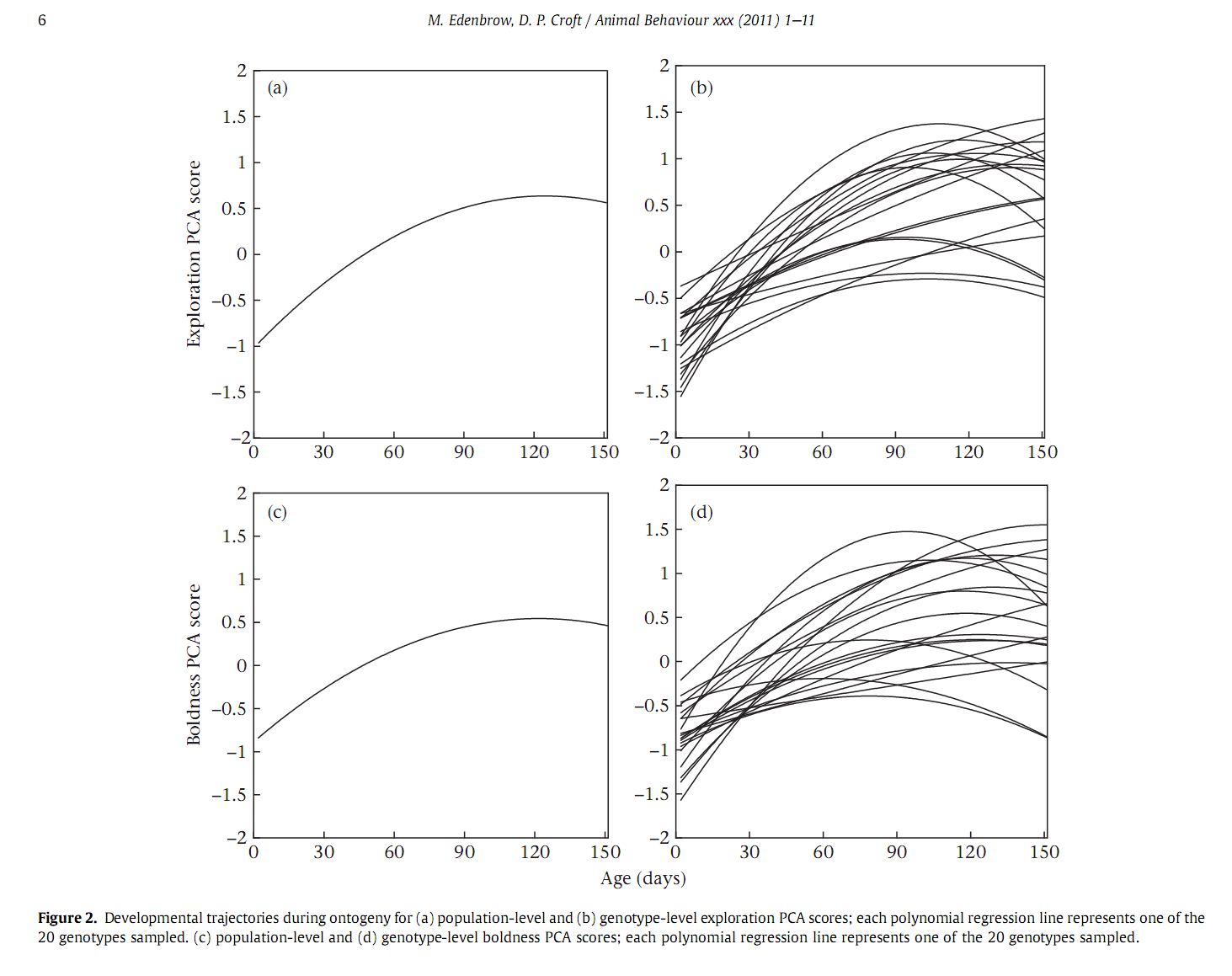
Самое важное: для всех генотипов повторяемость оценок смелости и исследовательской активности была практически нулевой или отрицательной за счёт высокой внутрииндивидуальной изменчивости по сравнению с изменчивостью между индивидами. БОльшая часть направленной изменчивости объясняется возрастом. Смелость и исследовательская активность возрастают в раннем развитии и выходят на плато при созревании. В особенности это касается исследовательской активности, которая резко возрастает на ранних стадиях и выходит на плато примерно на 110 день развития. Смелость возрастала медленнее, но выходила на плато примерно к тому же возрасту в 110 дней. Генотип объяснял только 1% вариаций как для смелости, так и для исследовательской активности. Однако генотип очень сильно (на 68%) влиял на траекторию развития исследовательской активности, существенно больше, чем параметры индивидуального развития (28%). Ещё более существенным было влияние генотипа на траекторию развития смелости (92%) с минимальным влиянием параметров индивидуального развития (5%). Наблюдалось некоторое влияние генотипа на скорость развития этих качеств в раннем онтогенезе, на время достижения плато и на последующее уменьшение смелости и исследовательской активности после созревания рыбки.
Наблюдали слабую незначимую корреляцию между средней для генотипа исследовательской активностью и средней смелостью со 2-го по 31-й день. При этом коэффициенты корреляции отличались для 2-го и 31-го дня незначительно. Для 31-го и 61-го дня коэффициенты корреляции отличались значительно, что может говорить об изменении корреляционной структуры между этими возрастами. Для 61-го и последующих дней наблюдалась сильная достоверная положительная корреляция, причём коэффициенты корреляции не отличались достоверно для 61-го, 91-го и 151-го дня. Скорость роста зависит от возраста и практически не зависит ни от генотипа, ни от взаимодействия генотип/возраст. Не зависит от генотипа также и траектория роста. Быстрее всего рост происходит на первых стадиях развития, замедляясь после 61-го дня.
Иными словами, полностью сформированный тип поведения от генотипа не зависит, хотя до некоторой степени зависит «путь», по которому формируется эта эквифинальная стадия. Что же зависит? От генотипа зависит возраст первой яйцекладки и общее число отложенных яиц, таким образом, генотип влиял на репродуктивную историю. Экспериментальные генотипы отличались от основного более поздним возрастом первой яйцекладки и меньшим общим числом отложенных яиц. Вспомнив различие «структуры» и «наполнения» в поведении, можно сказать, что от генотипа зависит не первое, но второе, и особенно «скороспелость» и «продуктивность» рыбки. Авторы называют эти характеристики «жизненной стратегией», которую надо отличать от «поведенческого типа». И в отношении психики «наполнение» у всех позвоночных, от нас до рыб, примерно одно и то же, заданное той самой двумерной системой координат, описывающей разнообразие темпераментов. А вот что различается – это сложность «структуры» психики, которая в разных филогенетических ветвях оказывается функцией сложности социальной организации, опосредованной прогрессивным развитием мозга.
Не было связи между средней характерной для генотипа скоростью роста и средними исследовательской активностью и смелостью. Средние смелость и исследовательская активность в 91 день (пока рыбы ещё не приступили к репродукции, которая начинается в среднем в 125 дней) не коррелировали со средним возрастом первой яйцекладки. Средняя смелость после созревания тоже не коррелировала со средним возрастом первой яйцекладки или общим числом отложенных яиц. Однако наблюдалась значимая корреляция между средней исследовательской активностью после созревания и возрастом первой яйцекладки, корреляция же с общим числом отложенных яиц была недостоверной.
Технически удобно считать, что индивидуальное поведение животного складывается под влиянием его личной истории: грубо говоря, в каком возрасте было «первое свидание», где оно происходило, как себя вели участники, сколько потомства от этого свидания осталось, какие были последствия, меняющие поведение в следующий отрезок жизни и пр. Если индивидуальная история подчиняется генетическим сигналам, которые определяют, когда организм созревает, когда ему пора размножаться и как себя вести, то клоны с идентичными генами должны приобрести схожие черты в поведении, если нет, то нет. Работа M.Edenbrow & D.P.Croft показывает, что «продукт поведения» до некоторой степени определяется генетическими сигналами (и созданной ими биологией организма), а вот форма реализуемого поведения, в той части которая определяет индивидуальность особи («животный» аналог наших личностных особенностей) – нет.
Или иначе, разнообразие поведенческих типов практически ортогонально разнообразию жизненных стратегий особей, определяющих их темпы роста и продуктивность, и в значительной степени зависящих от генотипа. Поведенческий тип – содержательная характеристика реализуемой особью онтогенетической траектории, относящаяся к биологической форме, он от генотипа не зависит и, видимо, «собирается» в зависимости от условий среды в месте роста рыбки, включая плотность скоплений и следующий отсюда групповой эффект.
Заключение
Отсюда можно сделать вывод общего характера — если «поведение управляет генами», и достаточно эффективно (примерно также как рыночная конъюнктура предъявляет больший или меньший спрос на продукцию тех или иных фабрик), то обратное неверно. Гены (вместе с длинным и сильно независимым от них аппаратом осуществления признака) не «управляют» поведением, а предоставляют для него «сырьё и комплектующие», примерно также, как охотники оставляют в избушке соль, спички, растопку и сухари для следующего вновь пришедшего. Как уже он использует этот ресурс — зависит от его опыта и ситуации: иными словами, «точка сборки» дефинитивного поведения из «комплектующих», представленных генетикой и биологией индивида, определяется состоянием надындивидуальной системы – популяции и сообщества, в которую индивид включается как малая и заменимая часть.
Частным проявлением этой асимметрии является управляющее воздействие «социального» на «биологическое» (даже у животных, а у человека особенно). Поскольку наш мозг, по словам Иоахима Бауэра, «делает из психологии биологию«, в каждом социальном явлении есть биологический стержень, но лишь в исполнительском звене, а не в управляющей части и не в пусковых механизмах. Надстройка «социального» над «биологическим» как раз и проявляется в том, что первое «перехватывает управление» и подчиняет биологические по сути реакции, вроде сексуальных, родительских, агрессивных и прочих, своим, чисто социальным стимулам и ситуациям, отстраняя от руля чисто биологические влечения. Как это видно в примере с брокерами, детьми и так далее.
В частности, я оспаривал предложенный А.В.Марковым концепт взаимодействия генов и поведения именно потому, что там опущено главное — социальные механизмы управления поведением (и через него – генами). В частности, присущие человеку способы распространения мнений и установления единомыслия в обществе, которые принято обозначать как «уступчивость» и «конформизм», вырабатывались именно так, чтобы эффективно действовать на людей с любой биологией и темпераментом. То же верно и для других механизмов социальной зависимости и социального влияния, у каждого вида, а тем более нашего, они развиваются так, чтобы быть максимально эмансипированными от мотивационной подосновы поступков и, соответственно, от биолого-генетических особенностей акторов.
И возвращаясь от человека к животным: особи вместе с их признаками в эволюционном процессе представляют собой не самоценные сущности (субъекты изменения), а всего лишь способ организовать статистические испытания, позволяющие отобрать лучшие поведенческие типы, жизненные стратегии и пр., в рамках того типа «столкновений» между индивидами, который следует из пространственно-этологической структуры популяции, а она видоспецифична. Отдельные особи имеют значение лишь как «лётчики-испытатели» разных поведенческих типов или жизненных стратегий; в процессе эволюции шлифуются и совершенствуются именно они, в том числе и по способности устойчиво реализовываться на максимально широком разнообразии биологии и генетики индивидов. То, что предлагал Шмальгаузен для пренатального онтогенеза, говоря о его стабилизации и рационализации, действует для его продолжения на постнатальной стадии, по крайней мере в том аспекте, что связан с психикой и поведением.
[1] Щипанов Н.А., 2000. Некоторые аспекты популяционной устойчивости мелких млекопитающих // Зоологический журнал. Т.120. №1. С.73-87. В.К.
Рекомендуем прочесть
 Истинный «маммонист» или как херр Федер боролся за «национальный социализм»
Истинный «маммонист» или как херр Федер боролся за «национальный социализм» Дело о жабьих лапках: воскрешение научной репутации Пауля Каммерера
Дело о жабьих лапках: воскрешение научной репутации Пауля Каммерера Об агрессии
Об агрессии Почему насилие над женщинами в Индии сходит с рук преступникам?
Почему насилие над женщинами в Индии сходит с рук преступникам? Есть ценностей незыблемая ска́ла…
Есть ценностей незыблемая ска́ла… Немец
Немец Социальный профиль членов и руководителей НСДАП
Социальный профиль членов и руководителей НСДАП РПЦ претендует в 2014 году на получение 1 млрд 158 млн рублей из федерального бюджета
РПЦ претендует в 2014 году на получение 1 млрд 158 млн рублей из федерального бюджета
