

С.В.Рожнов, Г.А.Заварзин
Аннотация
Содержание
- 1 Аннотация
- 2 Введение
- 3 Рифы в глобальном балансе
- 4 Рифовая система – это геоморфологический барьер, седиментационный и химико-биологический фильтры
- 5 Архитектура рифа
- 6 Трофические система современных коралловых рифов
- 7 Трофическая система палеозойских рифов
- 8 Трофическая система строматолитового рифа
- 9 Появление иловых холмов
- 10 Появление каркасных рифов
- 11 Седиментация, связанная с так называемыми консервативными ионами: Cl-, Na+, SO2-4, Mg2+, Ca2+
- 12 Рифовые системы: колыбель или музей биоразнообразия?
- 13 Геоморфология конвергентных карбонатных платформ, строматолитов и рифов в масштабе эволюции биосферы
- 14 Список литературы
Рифовые системы являются геоморфологическим барьером, седиментационными и химико-биологическими фильтрами в направленном от суши к океану потоке влияния гео-биосферных процессов. Геоморфологический барьер рифовой системы может быть выражен в разном масштабе биогенного рельефа. Прямое воздействие биоты создает карбонатный геохимический барьер со шлейфом последствий для осадконакопления и, в более широких масштабах, для формирования осадочной оболочки. Топографически продуценты и консументы рифа могут быть разнесены на большое расстояние или быть настолько сближены, что представляют собой единое целое.
Трофическую систему современного кораллового рифа можно представить в виде двойной петли. По внутренней петле симбиотические водоросли поставляют органику кораллам, а кораллы водорослям биогены. Внешняя петля обеспечивает развитие биоразнообразия в коралловом рифе и увеличивает продуктивность кораллов.
Введение
Под рифами нередко понимают любые биогенные постройки, выраженные в топографическом рельефе (Wood, 1999). Выраженность рифов в рельефе подразумевает прямое воздействие биоты на геоморфологию. Топография, как и рифы, может быть разного масштаба. Но в целом рифы тесно связаны с карбонатными платформами, под которыми понимается обширное карбонатное тело с более или менее горизонтальной верхней поверхностью и обрывистыми окраинами. Карбонатные платформы располагаются обычно в пределах шельфа и классифицируются по морфологическим, тектоническим и генетическим признакам.
Многие рифовые системы нередко приурочены к бровке шельфовой карбонатной платформы, маркируя перегиб в сторону океанических глубин. Рифы и карбонатные платформы обязаны своим развитием, прежде всего, биоте, поставляющей карбонатный материал и образующей сложную гео-биологическую систему с окружающей средой. На развитие рифов и карбонатных платформ прямо или опосредованно влияют многие факторы, нередко образующие положительные и отрицательные обратные связи с рифовой системой. Воздействие этих факторов может быть разной продолжительности и разного масштаба. Факторы, действующие в масштабах эволюции биосферы, оказывают более значительное влияние на развитие рифовых систем, чем простое суммирование и усреднение коротких по продолжительности и ограниченных по площади факторов.
Рифы приурочены к определенной ландшафтной и климатической зоне и представляют наиболее наглядный пример прямого воздействия биоты на геоморфологический рельеф обитаемой поверхности Земли. Они образуют важный пограничный биоценоз на границе суши и моря (pис. 1).
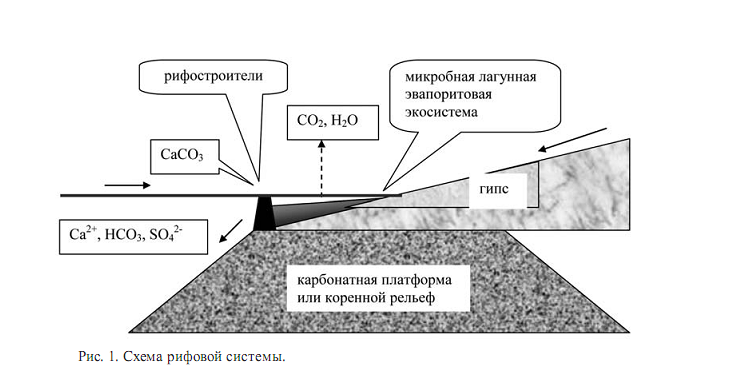
Прямое воздействие биоты создает карбонатный геохимический барьер со шлейфом последствий для осадконакопления и, в более широких масштабах, для формирования осадочной оболочки. В зарифовом пространстве развиваются геохимические и биологические процессы лагунного типа, свойственные амфибиальным ландшафтам. Перед фронтом рифа существует пелагическая система открытого моря. Контрастная обстановка создает условия для быстрого протекания геохимических процессов в масштабах мегабиологии. Опосредованное воздействие биоты может превосходить прямые процессы образования скелетных структур.
Влияние различных факторов на развитие рифовых систем и, шире, карбонатных платформ, активно изучается многими исследователями в мире. В нашей стране особое место в изучении кораллов и рифовых фаций принадлежит академику Б.С. Соколову. В 1951–1955 гг. вышла его фундаментальная монография «Табуляты палеозоя Европейской части СССР» – серия из шести книг, в которой впервые в мировой литературе на огромном материале рассмотрены общие вопросы систематики, эволюции, филогении, стратиграфического и географического распространения книдарий (табулят, гелиолитид, хететид) с охватом всей палеозойской истории, от возникновения и до вымирания. В семидесятые годы под его руководством была создана уникальная по своим масштабам научная программа сравнительного изучения современных и ископаемых рифовых экосистем, которая связала воедино имеющие первоочередное экономическое значение палеонтологические исследования кораллов и изучение рифогенных систем современного Мирового океана. Работами академика А.Ю. Розанова по археоциатам и стратиграфии кембрия Сибири были заложены основы понимания развития и функционирования кембрийских рифовых систем. Многие факты по стратиграфическому и географическому распространению рифовых систем и влияющим на их развитие факторов (тектоника, климат, седиментация, течения, соленость, биоразнообразие, эвстатика) собраны в единую базу данных, например в рамках проекта “Carbonate Analogs Through Time”, закончившегося десять лет назад (Lukasik, Simo, 2008). Значительно меньше обращают внимания на другой процесс – влияние рифовых систем на развитие биосферы. Этот аспект развития рифовых систем требует специального изучения, которое предполагает выявление положительных и отрицательных обратных связей в развитии рифовой системы, вписанной в географическое, геологическое и биологическое окружение.
Рифы в глобальном балансе
В глобальном балансе рифовые системы играют роль стока СО2 с образованием известняковых карбонатных толщ, но одновременно служат источником так называемого «кораллового потока СО2», возвращающего половину растворенной углекислоты в атмосферу. Только в этом смысле, как играющие большую роль в цикле углекислого газа, они могут называться легкими планеты, как выделяющие углекислый газ. Но захоронение углерода в виде карбонатов как конечного результата стока углекислого газа не связано с выделением кислорода и его глобальным балансом. Поэтому сравнение связывания углекислого газа в рифах и захоронения органики в болотных отложениях торфа (Жирков, 2010) некорректно. По глобальному масштабу процесса связывания СО2 в осадочные породы рифовые системы конкурируют с образованием карбонатного ила планктонными организмами и отложением скелетных образований многоклеточными организмами, кокколитофоридами и фораминиферами вне рифовой системы.
Обстановка накопления карбонатов жестко связана с климатом и происходит в условиях избыточного испарения и удаления СО2 вследствие падения растворимости углекислоты на прогреваемом мелководье. Привнос растворенной углекислоты в географическую зону образования рифов происходит с холодной водой вследствие глобального конвейера морских течений. Огромное количество захороненного карбоната в рифах и карбонатных платформах (в фанерозое не менее 50 % всех осадочных образований) не могло не быть мощным регулятором углекислого газа в атмосфере.
Наибольшее количество карбоната кальция образуется в тропиках, то есть при достаточно высокой температуре воды и атмосферы, что связано с нарушением карбонатного равновесия в теплых водах в связи с уменьшением растворимости углекислоты. Выделение в атмосферу углекислого газа приводит к парниковому эффекту и потеплению, а его связывание биогенным карбонатом должно было приводить к уменьшению содержания углекислого газа в атмосфере, уменьшению парникового эффекта и буферизации климатических изменений. Особенно важна буферная роль рифов в додевонский период, когда наземный растительный покров был недостаточно развит.
Образование рифовых систем на перепаде глубин указывает на возможную важность апвеллинга. В то же время, волновое движение вызывает опускание воды по фронту рифа. В разные геологические эпохи при различном расположении материков и разном климате роль апвелингов и даунвелингов в распространении рифов сильно менялась.
Рифовая система – это геоморфологический барьер, седиментационный и химико-биологический фильтры
Эволюция биосферы происходит преимущественно в двух географических сферах: в океанах и на суше, взаимодействие которых происходит через морскую и континентальную прибрежную полосу, и воздействует на третью сферу – атмосферу. Эта поверхностная область планеты получила название географической оболочки в смысле, приданном ей акад. А.А. Григорьевым в 1932 г. (Григорьев, 1932, 1937). Биосфера располагается в еe пределах. Рифовая система, или шире, карбонатная платформа является геоморфологическим барьером и седиментационным и химико-биологическим фильтром в направленном от суши к океану потоке влияния гео-биосферных процессов. Рифовые системы, формируя донный рельеф, являются фильтром для взвеси, поступающей с береговой суши.
Поступление терригенного материала, его объем, минералогический и гранулометрический состав зависят от рельефа, климата и состава выветриваемых на суше пород. Взвесь может улавливаться рифовой системой и осаждаться в ней как на щетке, формируя строматолиты и иловые холмы, а может осаждаться в лагуне между рифом и берегом, чередуясь с эпизодами эвапоритовой седиментации.
Рифовая система отражает наступление суши на море с поясом рифов, располагающимся на границе в качестве волнолома. Зарифовое пространство служит ловушкой для хемогенной садки эвапоритов морского происхождения и здесь развиваются характерные для амфибиальных талассофильных ландшафтов микробные процессы. Особо следует обратить внимание на доминирующий в лагунах цикл серы.
Архитектура рифа
Рифогенное сообщество – сложная экосистема, в которой каркасообразователи создают локальные условия для взаимодействующих между собой организмов. Структура создается организмами-строителями, эдификаторами. Образование рифов защищает берег от размывания при трансгрессии моря, создавая условия для сохранения захороненных слоев. Геоморфологический барьер рифовой системы может быть выражен в разном масштабе биогенного рельефа. На море и на суше осуществляются принципы единой архитектуры от биогенного нанорельефа болотной кочки с депрессией-мочажиной до строматолитовых биогермов и от них к рифам и рифогенным структурам, образованным разными организмами, от археоциат, колониальных губок, кишечнополостных, мшанок до одиночных брахиопод (рихтгофении), двустворчатых моллюсков (рудисты) и даже иглокожих (эдриоастероидея Cyathocystis). Основой единства архитектуры при различии эдификаторов-строителей является возникновение обратных положительных и отрицательных связей между развитием организмов и оптимумом условий на месте их поселения при росте потомства на отмерших слоях родительских поколений, в одном случае органических остатков (кочка), в другом – литифицированных (строматолиты), в третьем случае на скелетных структурах (большинство рифостроителей). В случае кочки, образующейся в исключительно спокойной гидродинамической обстановке болота или луговины, причиной может служить тяга к свету растений-эдификаторов, часто мхов и не обязательно сосудистых растений. Для строматолитовых биогермов в виде подушек может действовать аналогичная тяга к свету и преимущество в росте для расположенных ближе к самой высокой точке поверхности группировок цианобактерий (рис.2).

Случайная неравномерность исходного нанорельефа при первом заселении может сразу давать начало возникновению обратных связей между вселенцами и формирующимся биорельефом поверхности дна. Поселенцы на слегка возвышающемся микроучастке дна развиваются быстрее из-за немного лучших условий по сравнению с соседними слегка заглубленными поверхностями. Отмирающие родительские слои быстрее приподнимают живых вселенцев над окружающей поверхностью. Новые поколения развиваются еще быстрее. Возникает обратная положительная связь. Но при достижении определенной высоты над уровнем окружающей поверхности деструктивные силы (гидродинамика в море, ветер на суше) делают конструкцию неустойчивой, и положительная обратная связь переходит в отрицательную. Образование мочажин между кочками и промежутков между подушками строматолитов связано не столько с усиленной деструкцией, сколько со слабой скоростью роста, подавляемого, например, скоплением ила в промежутках между подушками или затенением. В этих депрессиях сразу же возникает обратная отрицательная связь: чем быстрее в понижениях растет сообщество, тем сильнее действуют на него деструктивные факторы. Поэтому ни мочажины, ни промежутки между подушками строматолитов не являются результатом деструкции единого плоского рельефа, а возникают при усиленном росте соседних участков.
Литогенетическая зональность рифа основана на выделении участков биогенного роста, деструкции, транспорта и осадконакопления, как биогенного, так и кластического (Преображенский, 1986). Наибольший биогенный рост наблюдается на поверхности рифового плато благодаря высокой скорости роста здесь известковых водорослей. Вместе с тем, эта зона наиболее активного роста приурочена к зоне волнолома, где происходит наиболее активное механическое разрушение биогенного материала. Процессы накопления и деструкции сбалансированы в этой зоне активного волнового воздействия, благодаря чему поверхность плато все время расположена на одной и той же метровой глубине.
Грубообломочный материал, образующийся при постоянном волновом разрушении рифового плато, осаждается в районе системы баттресс («контрфорсов»). В этой зоне происходит также значительный биогенный рост, повышающий общий уровень платформы баттресс, так как здесь наблюдается максимальная скорость роста кораллов и массовое приживание обломков кораллов, сорванных штормами с плато. Прижившиеся обломки цементируют отлагающийся среди них органогенно-грубообломочный плохо сортированный материал, укрепляя риф. Наиболее мелкий и медленно оседающий материал спускается по склонам рифа, образуя у его подножья рифовый шлейф.
В тыловой части рифа и в пририфовой части лагуны происходит активная донная эрозия благодаря приливно-отливным и штормовым течениям. В лагуне накапливается преимущественно мелкообломочный материал, отложения которого имеют отчетливую косую слоистость. Такое распределение литодинамического процесса по зонам приводит к суммарному перемещению поверхности рифа, направление которого зависит, прежде всего, от соотношения скоростей роста рифа и эвстатического погружения.
В геологическом времени судьба рифа определяется тектоникой и эвстатическими колебаниями уровня моря. При сбалансированном опускании рифа он растет вверх, а при опускании уровня воды его поверхность размывается и риф растет фронтально. Риф и связанная с ним карбонатная платформа могут достигать большого объема и веса за геологически короткое время. Это может способствовать опусканию рифа и находящегося под ним геологической структуры под действием силы тяжести. Рифовые барьерные экосистемы располагаются на пассивном крае кратонов как коренной геологической структуры. Они отличаются от островных рифовых образований на возвышенностях под поверхностью моря с концентрическим ростом биоты в сторону фронта.
У строматолитов существует противопоставление плоских слоистых образований отмелого берега и столбчатых строматолитов (биогермов) мористого фронта. Если поверхностные продукционные слои рифогенной структуры представлены более или менее хрупкими отдельностями, то массивное тело рифа возникает в результате микробной трансформации, ведущей к цементации. Осуществляется тот же принцип, что в дереве: отмершие части выполняют механическую функцию поддержания макроструктуры. Столбчатые строматолиты имеют разные размеры от сантиметровых столбиков до многометровых биогермов (рис. 2). Классификация форм столбчатых строматолитов (конофитоны и др.) первоначально связывалась с различием цианобактерий-строителей и потому могла иметь значение для стратиграфии. Теперь это связывают преимущественно с гидродинамическими условиями среды обитания.
Необходимо различать биологически активную поверхность рифогенного сообщества, которое действует в темпе биологического времени, и образование геологической структуры, на которую действуют факторы многолетнего изменения климата, тектоники и эвстатики. Это хорошо видно на примере современных коралловых рифов: большая масса скелета живых кораллов покрыта тончайшей пленкой микронной толщины мягкого тела. При малой массе относительно скелета эта пленка имеет огромную поверхность, позволяющую активно взаимодействовать животному с внешней средой. Отмирающие скелеты животных составляют основную часть рифа как элемента рельефа.
Трофические система современных коралловых рифов
С биологической точки зрения рифы представляют автономное сообщество с собственной первичной продукцией, как за счет водорослей, так и за счет фототрофных симбионтов животных-эдификаторов. Рифы обычно омываются водами, крайне бедными биогенами. Поэтому без существенного притока биогенов экосистема рифа должна быть в высокой степени замкнутой. В целом сообщество рифа зависит от освещения и поэтому возникает и развивается на дневной поверхности. Высокая продуктивность рифовой экосистемы служит источником для Сорг.
В современном рифе выделяются две автотрофные зоны (внешняя, покрытая герматипными кораллами и внутренняя в виде водорослевого гребня на рифовой платформе) и одна гетеротрофная, расположенная в лагуне и отделенная от автотрофных зоной волнолома (Преображенский, 1986). Растворенный и взвешенный органический материал, поступающий из автотрофных зон, волнами забрасывается в лагуну, где заселяется бактериальными агрегатами и осаждается на дно. Этими агрегатами питаются многочисленные планктонофаги. Это и приводит к парадоксу обилия планктонофагов в рифовых лагунах при крайней бедности зоо- и фитопланктона. Экосистема кораллового рифа с энергетической позиции описана Одумом (Odum, 1971).
Баланс продукции и деструкции с попыткой определить значительный вклад микробной компоненты в Сорг -цикл сделан Ю.И. Сорокиным (1990). Их данные можно представить в обобщенном виде «сдвоенной рифовой петли» (рис.3).
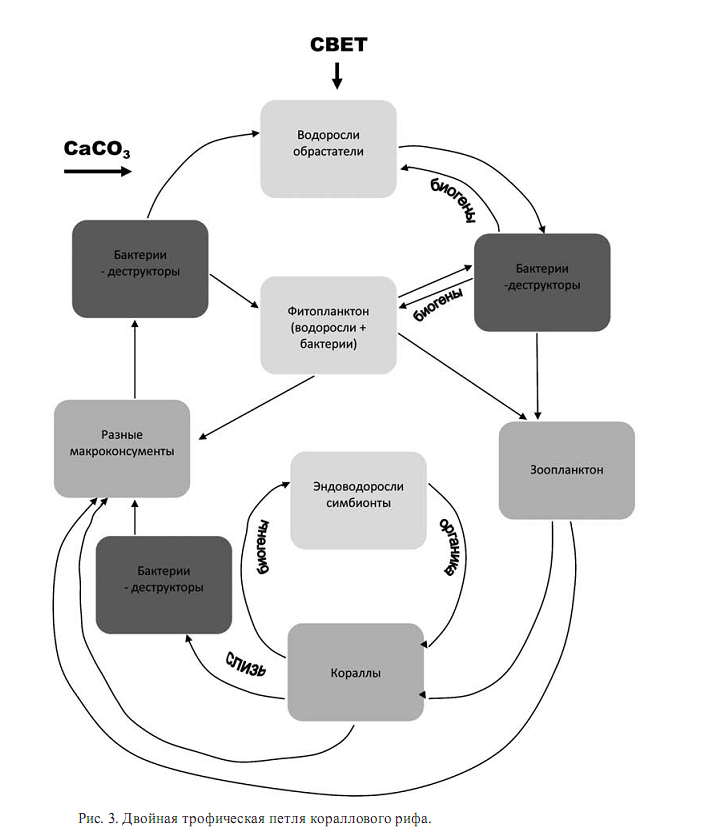
Рифовая система в своей основе всегда должна иметь продуцентов, которые либо непосредственно являются эдификаторами рифа, либо снабжают консументов-эдификаторов рифа необходимым потоком органического вещества. Топографически продуценты и консументы рифа могут быть разнесены на большое расстояние (глубоководные рифы с притоком органики из фотической зоны или в результате апвелинга) или быть настолько сближены, что представляют собой единое целое (кораллы с симбиотическими водорослями). При значительном притоке органики к рифу должен быть соответствующий баланс ее оттока и/или захоронения, потому что расширение рифовой системы ограничено геоморфологическими, тектоническими и климатическими факторами. Баланс этих перемещений органики важен для оценки глобального количества высвобождающегося кислорода в рамках более крупной экосистемы и распределения этого количества в пространстве и во времени.
Многочисленные специальные исследования показали, что уровень первичной продукции коралловых рифов очень высокий, а отношение первичной продукции к дыханию равно единице. Это означает, что такое рифовое сообщество производит продукции за год ровно столько, сколько потребляет. Конечно, сами кораллы не могут производить первичную продукцию.
Ее производят водоросли: симбиотические эндозойные в герматипных кораллах, известковые водоросли на рифовом плато и водоросли-обрастатели на отмерших скелетах организмов. Продукция симбиотических водорослей поступает непосредственно в ткани хозяина и потери при транспортировке биогенов от продуцентов к консументам оказываются минимальными. Поэтому, несмотря на крайне низкое содержание, например, фосфора в окружающей среде, высокая продуктивность системы поддерживается благодаря многократному его использованию без выхода его за пределы системы. Из этого следует незаменимая роль эндозойных водорослей в функционировании современной системы кораллового рифа. Кораллы выделяют огромное количество слизи. Эта слизь является питательной средой для бактерий, бурно развивающихся на этой слизи. Бактерии, в свою очередь, являются пищевым ресурсом для зоопланктона в коралловом рифе и макроконсументов, например, рыб. Зоопланктон становится добычей коралловых полипов, являясь, видимо, дополнительной к получаемой от симбионтов пище, а не основной. Таким образом, современная система кораллового рифа не нуждается в значительном притоке биогенов и первичной продукции извне. Но в этом случае, она не может являться источником органического вещества, уходящего в другие экосистемы или захороняющегося. Трофическую систему современного кораллового рифа можно представить в виде двойной петли. Внутренняя петля трофической структуры кораллового рифа является наиболее надежной. По этой петле симбиотические водоросли поставляют органику кораллам, а кораллы водорослям биогены.
Как показывают проведенные опыты, внутренняя петля может функционировать автономно и не может быть разорвана. Внешняя петля обеспечивает развитие биоразнообразия в коралловом рифе и увеличивает продуктивность кораллов. Такая схема энергетического баланса характерна для современных и, видимо, мезо-кайнозойских герматипных коралловых рифов. Но для мшанковых рифов и большинства палеозойских рифов такая схема может не работать, так как в них не доказано существование симбионтных эндозойных водорослей. Для таких рифов необходимо строить трофическую пирамиду, включающую как поступление аллохтонного органического вещества для фильтраторов, то есть превышение над концентрацией органических частиц клеток/мл, так и автохтонную продукцию. Автохтонная продукция может обеспечить существование рифогенного сообщества либо за счет бентосного фототрофного сообщества, прежде всего, известковых водорослей, красных и зеленых, водорослей-обрастателей, и за счет симбиотических водорослей, если удается доказать их существование.
При преобладании автохтонной продукции риф развивается как автономная экосистема с циклом органического углерода и накоплением минеральных компонентов при высокой локальной продуктивности «оазиса» в противоположность бедному «голубому» океану и бедной пустынной суше. При преобладании аллохтонного поступления органического вещества мы должны искать пути его поступления. Поступление его с суши возможно только после девона, когда первичная продуктивность суши стала достаточно высокой.
Но прямое поступление взвеси на риф должно приводить к его гибели из-за загрязнения и опреснения. Поэтому аллохтонное органическое вещество, если оно необходимо для развития рифа, должно попадать в его систему со стороны моря. Или, в особых случаях, со стороны особо большой лагуны. Поступающий с суши материал может быть транзитным в глубоководных рифах. При этом органический материал отфильтровывается, а обломочный смывается течением вниз по склону. Если рассматривать риф и лагуну как целостную трофическую систему, то при отсутствии поступления органики как с суши, так и с моря, эта система будет полностью замкнутой. При этом органика заносится в лагуну с автотрофной системы рифа и питает ее гетеротрофную систему. Освобождающиеся при разложении органики в лагуне биогены, возвращаясь в продукционную часть рифа, компенсируют потерю биогенов в автотрофной системе. Дополнительные биогены попадают в этом случае с суши (фосфор) и из атмосферы через продуцирование сине-зелеными водорослями (азот). Особенно характерна такая экосистема была для раннего палеозоя, когда продуцирование органики на суше было незначительным. В целом, палеозойская рифовая система заметно отличалась от мезо-кайнозойской из-за вероятного отсутствия в палеозое герматипных кораллов, скорость роста которых на порядок выше агерматипных кораллов.
Трофическая система палеозойских рифов
У палеозойских коралловых рифов, возможно, внутренней трофической петли не существовало из-за отсутствия эндозойных водорослей симбионтов. Об этом может свидетельствовать небольшое распространение значительных массивов именно коралловых рифов, что могло быть связано с малой скоростью их роста, обусловленной их агерматипностью. Не существовала такая петля и в мшанковых рифах, как древних, так и современных, так как у мшанок неизвестны эндозойные водоросли-симбионты.
В этом случае оставалась только внешняя трофическая петля, которая могла быть разорвана уходом части органики за пределы рифовой системы, например захоронением органики или уносом ее течениями. Для поддержания равновесия в такой рифовой системе отток органики должен был обеспечиваться ее дополнительным притоком извне для питания консументов-эдификаторов или притоком биогенов в фотическую часть рифа для жизнедеятельности продуцентов, обеспечивающих консументов-эдификаторов питанием и для развития непосредственно продуцентов-эдификаторов рифа. Непосредственное поступление на риф как «живой» органики, так и взвешенных «мертвых» органических частиц, в додевонский период могло происходить только со стороны моря, так как в то время поток органики с суши, до ее взрывной колонизации растениями, был незначителен. После девона поток органики с суши значительно увеличился, но все равно непосредственно с берега биогены и органическая взвесь на риф попадать не могли, так как риф в этом случае погибал от загрязнения и от опреснения. Большую роль в трофической системе рифа играли лагуны. Морскими волнами, перекатывающимися через риф и разрушающими его волноприбойную зону, органика с рифа сносилась в лагуну, откуда после прохождения лагунной трофической цепочки и окончательной деструкции бактериями, биогены обратными течениями заносились на риф, поддерживая его продуцирующую составляющую. Поэтому оптимальное место для развития рифов является перегиб рампа или иной карбонатной платформы на некотором удалении от берега.
Трофическая система строматолитового рифа
Трофическая система строматолитового рифа была иной. Продуценты и консументы здесь всегда находились в непосредственном контакте. Отток органики был небольшой. Поэтому небольшого стабильного притока биогенов им было достаточно. Основной приток биогенов в первую очередь фосфора, шел, видимо, с суши вместе с плоскостным твердым стоком пологого рельефа. Но существует проблема оттока органики. Содержание органики в ископаемых строматолитах всегда минимальное. Поэтому возникает вопрос о ее судьбе после отмирания глубоких слоев строматолита.
Можно себе представить, что она вымывалась в последующем диагенезе осадков. Но в этом случае должны быть остатки, пятна такой органики хотя бы в некоторых местах строматолитовых массивов в отдельных местонахождениях. Но такие находки неизвестны. Нам остается только предположить, что органика быстро разлагалась и биогены использовались поверхностными слоями строматолитового сообщества. Если такое предположение верно, то трофическая система строматолитов могла быть тоже в значительной степени замкнутой. Строматолитовые постройки распространялись по всему мелководью. Поэтому они создавали амфибиальный ландшафт, игравший в докембрии важную роль в биотическом взаимодействии суша-море. Редкие эвапоритовые отложения в докембрии были связаны преимущественно с небиогенной изоляцией лагун.
Появление иловых холмов
Для эпиконтинентальных палеозойских морей, с их большими мелководными пространствами, удаленными как от берега, так и от значительных глубин океанического склона, характерны иловые холмы, возникающие в результате заиления биогенных построек собственными отходами.
Рифовые постройки животного происхождения строятся преимущественно фильтрующими организмами. Поэтому основная проблема возможности существования таких построек состоит в том как не стать занесенными илом, не заилиться. Значительная часть илистого материала может оседать в биогерме непосредственно из течений. Тем не менее, основная часть ила попадает на биогерм или колонию фильтрующих организмов через поток фильтруемой взвеси, из которой органическая часть в значительной степени усваивается организмом, а минеральная выбрасывается им наружу в виде фекальных пеллет, оседающих вокруг организма. В этом основная причина относительной недолговечности устричных и, особенно, мидиевых банок. Небольшая скорость вертикального роста устричных колоний, нарастающих друг на друга цементированием, и отсутствие посмертного скрепления мидий, прикрепляющихся биссусом, при мощной фильтрующей системе приводит к периодическому заиливанию их поселений, гибели и перемещению поселений на другое место.
В противоположность этим моллюскам, другие ископаемые моллюски, меловые рудисты, были способны создавать крупные рифовые постройки. Проблема защиты от заиливания может решаться несколькими путями: поселением в местах, обедненных взвешенными минеральными частицами, как терригенными, так и карбонатными; поселением на перегибе рельефа с течениями или волнением воды, вымывающими ил из биогерма; скоростью роста вверх, превышающей скорость накопления ила. В последнем случае, при относительно быстром осадконакоплении образуются иловые холмы, не очень сильно возвышающиеся над окружающей поверхностью дна в момент формирования, но в геологическом разрезе выглядящие как высокие рифы. Такие иловые холмы, или мадмаунды, характерны для платформенных эпиконтинентальных морей, где они могли вырастать на плоском или слабо наклонном рампе. Скорость роста современных рифов с герматипными кораллами почти на порядок выше глубоководных агерматипных. Поэтому можно предположить, что многие палеозойские рифы, эдификаторами которых служили агерматипные кораллы, в действительности были слабо возвышающимися над дном иловыми холмами.
Иловые холмы отличались от настоящих каркасных рифов нередко слабой механической связью между скелетными организмами, особенно посмертной, и медленным ростом. Из-за этого они не могли противостоять сильным волновым движениям воды, но хорошо улавливали мелкодисперсный материал, который заносил промежутки между организмами. Этот ил при преобладающем карбонатном составе и химическом составе поровых вод мог цементироваться вскоре после отложения и создавать уплотненное основание этих рифовых построек. Поэтому, играя первоначально роль улавливающих ил щеток, эти постройки на зрелой стадии могли создавать барьер, изолирующий лагуну от открытого моря. Сочетание биогермов из фильтрующих организмов с водорослевыми постройками, способными, образовать крепкий волнолом, способствовал созданию такого барьера.
Зарифовая лагуна в этом случае могла занимать обширные пространства, распространяясь на сотни километров от рифа в сторону берега. При жарком климате, это приводило к отложению больших объемов эвапоритов, как например, в кембрии на Сибирской платформе. Объем эвапоритов в это время был настолько большим, что мог существенно менять общую соленость океана. Иловые холмы появились в раннем кембрии, где они строились преимущественно археоциатами.
В более молодых отложениях они нередко строились более типичными губками. При этом губковые поселения, накапливая вокруг себя ил, сами могли разрушаться после смерти, обогащая ил спикулами. Нередко иловые холмы, образованные поселениями губок, покрывались микробиальными микритовыми известняками, на отмерших и затвердевших частях которых развивалась богатая и разнообразная бентосная фауна.
Одним из примеров этого могут служить микритовые кольцевые структуры в среднем ордовике Ленинградской области (рис. 4). В центре этих структур и непосредственно под микритовыми известняками располагаются глины. Благодаря следам от растворенных спикул в глинах можно предположить, что глиняные холмы формировались в результате поселения губок, благодаря интенсивной фильтрации которых осаждался ил (Федоров, 1996).
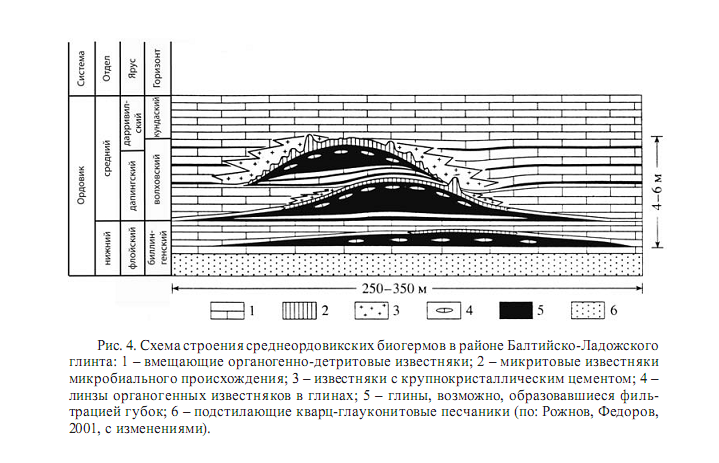
Получавшиеся таким образом холмы осваивали микробиальные маты, постепенно покрывавшие и бронировавшие иловый холм от подножия к центру быстро затвердевавшим микритовым известняком. Другим интересным примером иловых холмов могут служить верхнеордовикские биогермы близ пос. Вазалемма в Эстонии (рис. 5).

Полоса этих биогермов протягивается на 35 км с запада на восток при ширине от полкилометра до километра. Биогермы были построены водорослями, мшанками и иглокожими. Водоросли образовывали систему волноломных биогермов, между которыми образовывались менее крепкие биогермы, каркас которых строился мшанками и эдриоастероидеями (иглокожие) Cyathocystis (рис. 6).
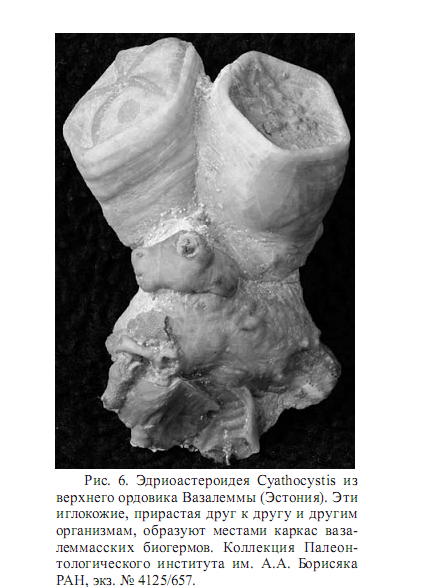
Каркас мшанковых и иглокожных рифов заполнялся карбонатным детритом, синседиметологическая цементация которого приводила местами к укреплению биогерма. Основным поставщиком детрита были стебельчатые иглокожие (ромбиферные цистоидеи) Hemicosmites, которые селились по краям биогермов и поставляли огромное количество карбонатного материала. Благодаря этому вокруг этой рифовой зоны образовались обширные слои цистоидного известняка значительной мощности (Мянниль, 1960; Рыымусокс, 1970). Иловые холмы с конца позднего девона до конца мезозоя сосуществуют с каркасными рифами, но в кайнозое они редки.
Появление каркасных рифов
Настоящие каркасные рифы появляются в истории Земли не раньше позднего девона, когда развиваются ветвистые колониальные четырехлучевые кораллы – ругозы. Это приводит к появлению модульной организации собственно рифов. Каждая колония многоклеточных животных, например, мшанок или кораллов, имеет модульную организацию, позволяющую ей принимать наиболее выгодную в данных условиях форму. Но эта модульная организация обособлена в каждой колонии, слагающей риф. При способности колоний успешно селиться на старых и отмирающих частях друг друга возникает модульная организация рифа. Наиболее успешно строительство каркасного рифа могло происходить при способности кораллов срастаться друг с другом, что могло обеспечиваться поселением личинок на отмерших частях старых колоний и высокой скоростью роста кораллов.
Если в рифообразующих кораллах отсутствовали эндозойные водоросли-симбионты, то скорость роста скелета кораллов была, видимо, небольшой. Но невысокая скорость роста могла компенсироваться их объемным ветвлением, что быстро увеличивало объем рифа. Если при этом промежутки между кораллитами быстро заполнялись осадком при последующей быстрой его цементации, то такие рифы могли достигать значительных размеров. Модульная организация каркасного рифа не обязательно связана с колониальными организмами. Крупные формы брахиопод (рихтгофении) и двустворчатых моллюсков (рудисты в мелу), нараставшие друг на друга с успехом могли строить каркасный риф по модульному принципу.
Седиментация, связанная с так называемыми консервативными ионами: Cl-, Na+, SO2-4, Mg2+, Ca2+
Основу рифовой системы, как и карбонатной платформы в целом, составляет седиментация, связанная с так называемыми консервативными ионами: Cl-, Na+, SO2-4, Mg2+, Ca2+. Пропорции этих элементов постоянны в открытом океане. Это связано с тем, что химические реакции, в которых они участвуют, протекают медленнее, чем их перемешивание течениями и волновым движением вод. В прибрежных районах пропорции и концентрации этих элементов могут меняться из-за притока пресных вод и эвапоризации.
Принципиальным вопросом, не нашедшим пока удовлетворительного объяснения, является корреляция строматолитовых построек с преимущественно быстрым внеклеточным отложением доломитов, сохраняющим ламинарную структуру биопленок циано-бактериального сообщества, в то время как рифовые структуры построены из кальцита и арагонита метазойными скелетными организмами и известковыми водорослями. Отложение карбонатов происходит в геохимических условиях самих по себе благоприятствующих осадке минеральных форм и достаточно четко связанных с климатическими условиями. Бесскелетная микробиота развивалась как эндоэвапоритовая группировка при садке карбонатов, отвечая за текстуру осадков и защищаясь от выедания. Та же линия наблюдается у известковых водорослей.
В рифах биогенным путем осаждается огромное количество карбоната кальция в виде кальцита и арагонита. Годовая продукция производимого современными коралловыми рифами карбонатного материала оценивается примерно в 2.5 млрд. тонн (Сорокин, 1990). Строматолиты и карбонатная платформа могут отличаться от каркасных рифов существенным участием доломита, в то время как внутренние компоненты скелета всегда представлены кальциевыми карбонатами. Соотношение концентрации кальций/магний и углекислого газа в морской воде имеет большое значение в синседиментационных диагенетических процессах, так как, по мнению ряда исследователей, влияет на растворимость арагонита и цементирование карбонатного ила осаждением кальцита из поровых вод. Этот процесс имеет важное значение для иловых холмов, особенно в палеозое. Поэтому для построения рифа большое значение имели животные-рифолюбы, способные вырабатывать большое количество карбонатного детрита, который мог быстро цементироваться кальцитом, осаждавшимся из поровых вод.
Среди таких животных наиболее важное значение в палеозое имели прикрепленные иглокожие (рис. 7). В ордовике это были преимущественно цистоидеи и эокриноидеи, а позже это были морские лилии и бластоидеи. Скелет иглокожих обычно состоит из большого количества монокристаллических пористых табличек, на которые он чаще всего распадается после смерти. В краевой части рифа плотность поселения иглокожих могла быть очень высокой. Не менее высокой была их карбонатная продуктивность. Таблички и их осколки могли занимать очень большой объем, захороняться в рифовых нишах и у основания рифа, обычно с мористой стороны. Кальцитовый состав табличек скелета иглокожих способствовал их синседиментационной консолидации осаждения при осаждении кальцита из поровых вод. В отличие от современных условий, когда в мелководных морях осаждается из поровых вод арагонит или, реже высокомагнезиальный кальцит, в течение фанерозоя из поровых вод в некоторые периоды осаждался преимущественно кальцит. В фанерозое происходило чередование так называемых кальцитовых морей с преимущественным осаждением кальцитовых оолитов и кальцитового цемента из поровых вод в осадке и арагонитовых морей с преимущественным осаждением арагонитовых оолитов и арагонитового цемента из поровых вод.
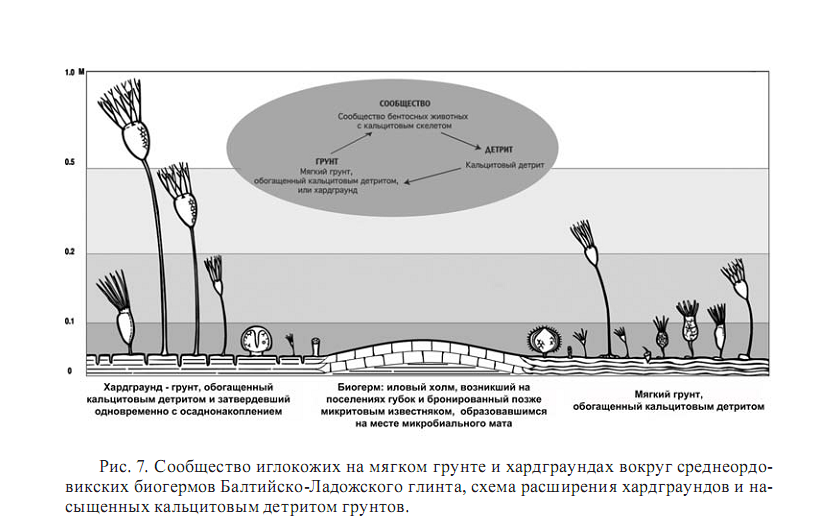
Это различие в характере осаждения карбоната кальция связывают с глобальным изменением соотношения кальция и магния в морской воде и растворенного углекислого газа. В кальцитовых морях кальцитовый детрит в осадке хорошо цементировался кальцитовым цементом, осаждавшимся из поровых вод. Поэтому в рифах и биогермах развитие прикрепленных иглокожих приводило к укреплению всей постройки при синседиментационной консолидации остатков их скелета в самом рифе и в окружающих его фациях в эпохи кальцитовых морей. В мезозое и кайнозое морские лилии стали на рифах относительно редки в отличие от свободноживущих иглокожих морских ежей и морских звезд. Роль этих иглокожих принципиально изменилась – они стали играть в рифе в основном деструктивную роль, объедая полипы и водоросли и рассверливая тело рифа.
Рифовые системы: колыбель или музей биоразнообразия?
Изменение таксономического состава биологического разнообразия прямым образом влияет на формирование рифовых систем. Этим обусловлена смена каркасостроителей рифа и рифолюбов с течением геологического времени. Но и сами рифы влияют на развитие биоразнообразия. По современным подсчетам более 60 % таксонов морских беспозвоночных в фанерозое впервые появились в мелководных карбонатных тропических фациях, а около 40 % именно в рифовых фациях. Это свидетельствует о важности рифовых местообитаний как колыбели для многих таксонов. Но в действительности географические закономерности происхождения надвидовых таксонов могут оказаться значительно сложней, и рифовые системы могут являться не столько колыбелью, центром происхождения и расселения таксонов, сколько музеем или насосом, втягивающим в себя разнообразные таксоны из различных умеренно-тепловодных местообитаний. По всей вероятности, для одних таксонов они могут оказаться колыбелью, а для других рефугием. В рифах разных геологических эпох и разных континентов соотношение «колыбельных» и «музейных» таксонов было различным.
Например, сравнение фауны холодноводных среднеордовикских (волховских) и тропических позднеордовикских (вазалемаских) рифов Балтийского эпиконтинентального бассейна показывает, что фауна иглокожих в биогермах формировалась из представителей окружающих биогермы биотопов мягкого и твердого грунтов (рис. 7). Но среди биогермов их численное соотношение было иным, чем в окружающем пространстве. Большинство родов иглокожих, живших на холодноводных биогермах волховского горизонта, исчезло к концу кундаского горизонта, ненадолго пережив существование самих биогермов. Исключение представляет морская лилия Hoplocrinus. Два рода иглокожих, появившиеся в балтийском бассейне сначала на мягких грунтах в холодных водах (род Cyathocystis) волховского моря и в несколько более теплых водах кундаского моря (род Hemicosmites) продолжали жить в Балтийском бассейне и в вазалемасских тропических биогермах кейлаского моря резко достигли огромной численности. При этом индивидуальные размеры их представителей увеличились в несколько раз. Увеличение разнообразия, индивидуальных размеров, численности и кальцитовой продуктивности иглокожих являются характерными отличиями тропических кейласких рифов от холодноводных волховских (Rozhnov, 2010).
Геоморфология конвергентных карбонатных платформ, строматолитов и рифов в масштабе эволюции биосферы
В масштабе времени в миллиарды лет важна геоморфологическая конвергенция карбонатных платформ, строматолитов, и рифов. В геологической истории они играли сходную геоморфологическую и геохимическибалансовую функцию. Биогенные консолидированные структуры служат бронирующим образованием, защищающим берег от размывания. В этом смысле можно говорить о «геоморфологическом естественном отборе», исключающим из геологической истории геоморфологические образования, не удовлетворяющие требованиям для собственного сохранения. Говорить о филогенетической «эволюции» одного сообщества из другого не приходится. Здесь есть только последовательность во времени. Разные организмы приспосабливаются к сходным условиям и выживают в них. И строматолиты, и рифы служат примером сукцессии в конкретном месте обитания биоты, обусловленном накоплением мертвого продукта. Географическая обстановка служит определяющим фактором для геохимических условий, в которых разные по происхождению биологические сообщества удовлетворяют и физическим (гидродинамика), и геохимическим условиям. При формировании рифов и аналогичных барьерных структур для биолога существенна геобиофизическая характеристика среды обитания, как на макроуровне, так и на микроуровне в смысле обитания в пористом пространстве рифов. Иерархия требований к формированию рифов начинается с географического положения, для современных условий соответствующих низким широтам в климатическом поясе ; далее к требованию современного геохимического состава океана с 10 мМ кальция, концентрация которого могла быть иной в строматолитовую эпоху; далее к коренному рельефу дна, на котором возникает постройка рифа; далее к преобладающему направлению нагона морской воды, из которой извлекается кальций для построения рифа; далее к суточному режиму освещения и затемнения и, в особенности, двукратному изменению уровня в приливно-отливной зоне. Каркасные рифы насчитывают менее 500 Ма из общей истории прибрежных карбонатных образований более 3 млрд. лет. Рифы, создавая особый рельеф на границе суши и океана, нередко продолжали выполнять геоморфологическую роль, оказываясь в результате эпейрогенических движений, на суше вдали от моря. Препарируясь денудационными процессами, они создавали на земной поверхности своеобразный рельеф из наземных гор и холмов, повторяющих рифовый рельеф когда-то существовавшего здесь древнего бассейна. Погребенные рифы равнинных областей создали своеобразный внутренний геоморфологический скелет и оказывают сильнейшее воздействие на циркуляцию подземных вод. Ископаемые рифы в этом отношении отличны от бронирующих рифов экотонов.
Список литературы
Григорьев А.А. 1932. Материалы к физической географии северо-восточной части Кольского полуострова. Труды СОПС АН СССР, сер. «Кольская», вып. 4, Л.
Григорьев А.А. 1937. О некоторых основных физико-географических закономерностях. Изв. АН СССР, сер. геогр. и геофиз., № 4.
Жирков И.А. 2010. Жизнь на дне. Био-география и био-экология бентоса. М.: Товарищество научных изданий КМК. 453 с.
Заварзин Г.А. 2003. Лекции по природоведческой микробиологии. М.: Наука. 348 с.
Мянниль Р.М. 1960. Стратиграфия оандуского («вазалеммаского») горизонта. Труды Геологического института АН ЭССР. № 5. С. 89–115.
Преображенский Б.В. 1986. Современные рифы. М.: Наука. 244 с.
Рожнов С.В., Федоров П.В. 2001. Новый род криптокринитид (Eocrinoidea, Echinodermata) из прибиогермных отложений волховского яруса (верхний арениг, Ордовик) Ленинградской области // Палеонтол. журн. № 6. С. 42–49.
Рыымусокс А. 1970. Стратиграфия вируской и харьюской серий (Ордовик) северной Эстонии. Таллин: Валгус. 346 с.
Сорокин Ю.И. 1990. Экосистемы коралловых рифов. М.: Наука. 503 с.
Федоров П.В. 1996. Значение процесса осаждения терригенной взвеси бентосными биофильтраторами для образования раннеордовикских органогенных построек северо-запада Русской платформы // Вестник СПбГУ. Сер. 7. Геология, география. Вып. 2 (№ 14). С. 143–151.
Lukasik J., Simo J.A.T. 2008. Controls on development of Phanerozoic carbonate platforms and reefs – introduction and synthesis // Lukasik J. and J.A.T. Simo (eds.).
Controls on carbonate platforms and reef development. SEPM (Society for Sedimentary Geology) Special Publication 89. Tulsa, O�lahoma, 2008. Pp. 5–12.
Odum E.P. 1971. Fundamentals of ecology. Third Edition. Philadelphia, London, Toronto: W.B. Saunders Company (русский перевод: Одум. Ю. 1975. Основы экологии. М.: Мир, 1975. 740 c.).
Rozhnov S.V. 2010. Sibling echinoderm taxa on isolated Ordovician continents: Problem of center of origin // Bulletin of Geosciences. N 85 (4). Pp. 671–678.
Wood R. 1999. Reef Evolution. Oxford, New York: Oxford University Press. xi + 414 pp.
Рифогенные формации и рифы в эволюции биосферы. Отв. ред. С.В. Рожнов. Серия «Гео-биологические процессы в прошлом». Москва, ПИН РАН, 2011. 228 с.








