
«Запасной ум» в психике животного
Содержание
- 1 «Запасной ум» в психике животного
- 2 «Левое и правое крыло» дарвинизма
- 3 «Потенциальная социальность» позвоночных
- 4 Желудёвый дятел: пластичность социальных отношений сообразно местным условиям при постоянстве «типа»
- 5 От моногамных пар к полигинадриии с помощниками — прочие виды
- 6 Важность дедуктивной реконструкции «типа» социальной организации
- 7 Каковы брачные отношения пёстрых дятлов — «моногамия с исключениями» или полигинандрия?
- 8 …и тот же вопрос применительно к певчим птицам
Давным-давно, в «Эволюции и психике» А.Н.Северцов предположил, что «верхняя планка» интеллекта животных устанавливается эволюцией «с сильным запасом», и животные (как минимум птицы с млекопитающими) сильно умнее, чем обычно мы видим в их собственной среде обитания и чем нужно обычно для выживания в ней.
«Когда животное приспособилось к наступившим изменениям и установилось некоторое новое состояние равновесия, так сказать, некоторая рутина жизни, то эта интенсивная изобретательность, игравшая большую роль в период сильного изменения условий, не требуется, и животное ее может обычно не проявлять, но способность к ней, так сказать, некоторый «запасной ум» в психике животного сохраняется и при случае, т. е. при наступлении нового изменения условий, может проявиться.
Таким образом, мы приходим к заключению (я отношусь к этому предположению только как к гипотезе), что высшие позвоночные животные (птицы и млекопитающие) в общем умнее, чем эта кажется при наблюдении их в обычных условиях их жизни. Мне кажется, что опыты дрессировки диких животных (особенно таких, от которых по условиям их существования трудно ждать высокой психики, как, например, тюлени или морские львы) вполне подтверждают эту гипотезу о «запасном уме» млекопитающих. Может быть, эта гипотеза могла бы оказаться полезной при суждении об исключительных проявлениях ума животных, которые мы имеем у лошадей Краля, собак проф. Циглера и т. д. Вдаваться в разбор этих вопросов, требующих специально психологического разбора (от которого я в этой статье сознательно уклоняюсь), я не буду».
«Левое и правое крыло» дарвинизма
Исследования интеллекта животных блестяще подтвердили это предположение. Не только ворОны-вОроны и человекообразные обезьяны, но и ящерицы с костистыми рыбами по исследованным аспектам «ума» обычно оказываются «сильно умнее», чем требуется для выживания в их естественной среде обитания. Скажем, грачи в условия эксперимента спонтанно используют орудия (камни, прутики), производят выбор наилучшего орудия для данной задачи, в т.ч. вместе с ворОнами решают протоорудийную задачу с нитями. В природе всего перечисленного они не делают (в отличие, скажем, от новокаледонских галок, да и ворОн), ибо не требуется для добывания корма в естественной среде — они не собирают его, а зондируют в почве наугад, и разнообразие кормовых объектов существенно ниже[1].
Почему так получается – отдельная тема. Я думаю, потому, что верны теории «левого, а не правого крыла дарвинизма», и эволюция биологической формы (начиная с видообразования) определяется тем, что филогенетическая косность целого важней адаптивной гибкости частей. См. подробнее «О дарвиновской эстетике».
В этом аспекте эволюция движется «по Шмальгаузену, а не Майру с Вилсоном», в сторону увеличения автономизации от внешней среды. Важным (скорей всего самым важным) фактором этой автономизации выступает социальность, развитие которой связано с созданием некоторого «гарантированного избытка» ресурсов и восприятием событий и изменений во внешней среде не напрямую, но опосредовано вызванными ими «откликами» в сдвигах социальных связей, взаимодействий и пр. событий внутри социума. Одновременно (как минимум у птиц и млекопитающих) прогресс социальной организации «движет вперёд и вверх» развитие индивидуальности, рассудочной деятельности и интеллекта особей, составляющих данный социум.
«Потенциальная социальность» позвоночных
Вполне изоморфную избыточность этологи обнаружили в «конструкциях» социальных систем разных видов птиц и млекопитающих. Они неизменно оказывались много сложнее, чем кажется исходя из паттерна отношений «среднего» поселения вида в обычном местообитании. Как только среда обеспечит заведомый избыток какого-то из ресурсов (чаще всего корма и/или безопасности), так вид немедленно демонстрирует существенно более сложные отношения, крупные группы и разнотипные взаимодействия, чем обычно. И это всегда неожиданность для специалистов.
Сложность структуры отношений в социальных системах, как и во всех прочих, оценивается разнообразием типов связей между разными категориями особей, так и числом индивидов, соединённых данными связями в общий «круг« компаньонов (Kumpan К.Лоренца). Большая сложность «конструкции» системы-социум проявляется в связывании большего числа особей более разнотипными отношениями одновременно, так что задействуются более сложные формы регулирования их активности, смены форм поведения и дифференциации ролей. Но не всегда более совершенные, почему усложнение часто сопровождается дезорганизацией.
Соответственно, «обычно наблюдаемые» отношения представляют собой не типичную, но упрощённую и вырожденную форму более сложной «конструкции« отношений. Её и нужно считать типической для данного вида; «обычная» репрезентирует «тип» искажённо или не отражает его вообще.
Желудёвый дятел: пластичность социальных отношений сообразно местным условиям при постоянстве «типа»
Так, на значительной части ареала в Центральной Америке желудёвый дятел Melanepres formicivorus живёт моногамными парами, круглый год охраняющими каждая свою территорию. Питаются они в основном насекомыми и фруктами, желудями — в существенно меньшей степени, и не запасают их. Действительно, здесь отсутствуют дубы с обильным плодоношением, обеспечивающие птиц кормом на всю зиму (а где присутствуют — скажем, в Белизе — они запасают жёлуди и живут только группами, одиночные пары не встречены). Поэтому молодняк расселяется, занимая собственные территории, им смысла нет оставаться на территориях взрослых в качестве «помощников». Преобладание моногамных пар (80% населения) наблюдается в штате Аризона (США), уже по другой причине. Здесь дятлы перелётны и присоединение молодых к взрослым не облегчает зимовки.

Самец в Калифорнии

Самка в Аризоне
Напротив, в Калифорнии (отчасти в Нью-Мехико) дятлы оседлы. Местный урожай желудей достаточен для прокормления на территориях не только пар, но и групп из нескольких особей (в первом случае — обычно, во втором — время от времени). В связи с чем развивается запасание желудей осенью, настолько интенсивное, что группа может кормиться всю зиму и ещё останется. В стволах отдельных деревьев дятлы выдалбливают тысячи отверстий, в каждое из которых загоняется жёлудь, сохраняющийся таким образом всю зиму. Такие нашпигованные желудями дубы называют «амбарами»; члены коммунальных групп спокойно едят их всю зиму (Sci. Amer. 1.08.1984, V.251, №2).

При высыхании жёлуди могут выпасть из лунки, и члены группы их время от времени перекладывают в более узкие отверстия. Запасы определяют и зимнее выживание, и весеннее размножение, поэтому деятельность группы по сбору, транспортировке, размещению желудей, их перепрятыванию, охране «амбаров» от попыток других групп и других видов разграбить их. Размножающиеся особи во всех этих «общих работах» участвуют интенсивней и больше времени, чем помощники. Однако же скорость запасания зависит от возраста, не от репродуктивного статуса, зависимости от пола нет вообще. Больше участвуя в «общих работах», помощники много больше времени тратят на вылазки по соседним территориям; самки вносят в заботу о гнезде и птенцах больший вклад, чем самцы, однако последние интенсивней чистят гнездо и только они насиживают ночью (Источник).
Так, бедный урожай желудей 1983 г. заметно повлиял на численность и социальную организацию калифорнийских дятлов. 71% социальных групп (из 34) распавшись, оставили территории. Судьба большинства из них неизвестна, лишь единицы возможно вернулись на данные территории весной. Из оставшихся на зиму на своих территориях птиц к февралю сохранились 52%. наибольшие потери фиксируются среди молодых помощников (75%), нежели среди размножающихся особей (20% — самцы, 32% — самки). Взрослые помощники по смертности промежуточны. Голод зимой существенно обострил агрессивность, и подчинённые особи часто исчезали из групп после стычек с доминантами.
Сходным образом одиночные зимовочные территории больших пёстрых дятлов включают в себя избыточный запас шишек, так что в апреле, при переходе к питанию насекомыми, часть их остаётся несъеденной. А если избытка нет, дятлы не занимают постоянные территории в этой местности или при истощении шишки — покидают заблаговременно в середине зимы.
При жизни отдельными парами гнездовая и зимовочная территории чаще всего разделены. Птицы зимуют на одиночных участках, при образовании пары участки самца и самки объединяются, после чего пара обычно ищет место для гнезда в другом месте, и достаточно далеко. Напротив, «коммуны» круглогодично территориальны. Охраняют и рекламируют «своё» пространство они интенсивней и строже, чем пары.
Иными словами, если бы соответствующие виды дубов вымерли до начала орнитологических исследований, возможность образования «коммун» — более сложно устроенных, многочисленных, с более интенсивными взаимодействиями — так и осталась бы чисто потенциальной.
Вот на этом заведомом избытке корма вид демонстрирует на первый взгляд совершенно иные и сильно более сложные варианты социальных отношений. Значительная часть территорий занята не парами, но полигинно-полиандрическими «коммунами» из нескольких размножающихся самцов и самок, каждая из которых более или менее свободно спаривается с прочими особями противоположного пола. В местах с регулярно высокой урожайностью наряду с парами (около 22%занятых территорий) встречаются группы из 3-х (23%), 4-х (17%), 5 (13%) и более (до 15 особей, как исключение фиксируются «коммуны» из 44-х самцов и 3-х самок).

Моногамная пара; Oak Creek Каньон, Аризона
Охрана (с сопровождением) партнёра, за которым ухаживают, появляется лишь при возможности «соблазнения» чужаками, от «своих» — того и другого практически нет вообще. Хотя в каждый отдельный момент временные пары выделяются вполне отчётливо, однако при значительной социальной связанности вступающих в отношения самцов и самок сколько-нибудь значительной преграды адюльтерам не воздвигается, они очень часты.
В группе существует система иерархии, сквозная и раздельная у разных полов, самцы доминируют над самцами, самки над самками, доминирующие особ и обычно старше и/или раньше поселились тут. При охране территории самцы группы атакуют самцов захватчиков, самки — самок. Поскольку последние преобладают, самки тратят на территориальные конфликты значительно больше времени, чем на заботу о потомстве.
В «коммунах» также присутствуют «помощники», обычно более молодые (до 10). Они сами не размножаются, находятся в подчинённом состоянии, взрослые их атакуют при попытках ухаживания за размножающимися птицами. Они в полной мере участвуют в охране территории, окрикивании хищников и выкармливании птенцов. К насиживанию их не допускают; этим занимаются размножающиеся особи, несущиеся в общее гнездо. Конкуренция между ними за право насиживать ведёт к социальной дезорганизации, повышающей гибель потомства. См. Koenig W. D., Stacey P.B., 1990. Acorn woodpeckers: Group living and food storage under contrasting ecological conditions// Cooperative Breeding in Birds: Long-term Studies of Ecology and Behavior (P. B. Stacey and W. D. Koenig, editors). Cambridge University Press, Cambridge. P.415-453.
«…мирный ход событий уступaет место нерaзберихе в сaмое ответственное время — когдa сaмки нaчинaют нестись в дупло, выдолбленное совместными усилиями всех членов коммуны. Дело в том, что сaмкa, готовaя отложить в гнездо свое первое яйцо, не терпит, чтобы в этот момент здесь нaходился кaкой-либо посторонний предмет, кaковым онa считaет и яйцо, отложенное ее нaпaрницей по группе. Поведение сaмки в подобной ситуaции бескомпромиссно: онa попросту выбрaсывaет из гнездa все те яйцa, которые уже успелa снести сaмкa, нaчaвшaя клaдку первой.
Сaмкa желудевого дятлa несет по одному яйцу рaз в сутки и зaвершaет отклaдку зaконченной порции яиц в течение 5-6 дней. Нетрудно видеть, что сaмкa рискует лишиться всей своей клaдки, если из-зa своей поспешности онa нaчнет нестись нa неделю рaньше своей нaпaрницы. Еще большие неприятности ожидaют обитaтельниц коммунaльной квaртиры, когдa их не две, a три: сaмкa, приступaющaя к яйцеклaдке последней, уничтожaет все бесценное содержимое гнездa, принaдлежaщее двум другим ее товaркaм. Коль скоро сaмки не отличaют отложенных ими сaмими яиц от чужих и действуют по велению слепого aвтомaтизмa, иногдa случaется тaк, что мaмaшa ошибaется в оценке ситуaции и выбрaсывaет из гнездa свое собственное яйцо. Впрочем, тaкое происходит нечaсто, ибо, кaк только сaмкa, нaчaвшaя клaдку последней, снеслa свое первое яйцо, онa прекрaщaет рaзбой и уже не трогaет яиц других сaмок, продолжaющих пополнять содержимое гнездa одновременно с ней.
Америкaнский орнитолог Р. Мaмми и его коллеги в стaтье под броским нaзвaнием «Гнездовaя конкуренция у желудевых дятлов: сестры уничтожaют яйцa друг другa» приводят следующие любопытные результaты своих исследовaний. Изучив ход рaзмножения в 14 гнездaх, кудa неслись по две сaмки, и в одном, принaдлежaщем трем мaмaшaм, ученые зaрегистрировaли 48 случaев уничтожения яиц сaмкaми и 3 — сaмцaми. В 35 эпизодaх сaмки удaляли из гнездa яйцa своих нaпaрниц и в 9 — свои собственные.
Нередко случaется тaк, что сaмкa, вынося из дуплa яйцо, не бросaет его, a поедaет, не препятствуя при этом и другим членaм группы присоединиться к трaпезе. И, что особенно любопытно, в четырех подобных ситуaциях сaмa мaмaшa, чье яйцо в дaнный момент подвергaлось нaдругaтельству, присоединялaсь к грaбителю, смaкуя вместе с ним вкусное содержимое скорлупы. В общей сложности в этих 15 гнездaх зa счет столь экстрaвaгaнтной мaнеры поведения их хозяев погиблa почти третья чaсть от общего числa яиц, отложенных сaмкaми-хозяйкaми».
Е.Н.Панов. Бегство от одиночества.

Типичный состав «коммун»
В Нью-Мехико урожай желудей достаточно регулярен, но нее везде значителен. В противоположность Калифорнии, не на всех территориях дятлы могут создать крупные запасы (>3000 дупел-«хранилищ» в одном ли нескольких стволах-«амбарах»). На части территорий урожай желудей позволяет создать только средние («амбаров» нет, 1-3 тыс.дупел рассеяны оп территории группы) или малые запасы (<1000 дупел-хранилищ, также рассеянных). В отличие от Аризоны, здесь низкий урожай не вызывает перелётности, не препятствует формированию коммунальных групп.
Однако на территориях малых и средних запасов значительно ниже вероятность обеспечения кормов на зиму (0.59; 0.23 и 0.09 соответственно). Если запасы кончаются, группа зимой покидает территорию; чем раньше это произойдёт, тем больше вероятность невозвращения на гнездование там весной. Территории больших запасов заняты каждое лето, а похуже часть сезонов пустуют – средние в 8%, малые в 54%. Успех размножения нa разных типах территорий падает пропорционально обеспеченности кормом, также как среднее число размножающихся особей в группах и вероятность самцам-первогодкам остаться помощниками на родной территории (2.33-1.76-0.96 слётков; 3.2-2.6-2.2 особей; 27%-15%-2% соответственно.
Самки в этой популяции помощниками не становятся; 54% их гнездится на соседних территориях, остальные исчезают. В отличие от Калифорнии, большая часть особей в группах пришлые: 56%, 80% и 87% на лучших, средних и пессимальных участках. Из 11 помощников-самцов 7 (64%) позже гнездились на родных территориях, 2 (18%) — на соседних и 2 исчезли. Таким образом, в Нью-Мехико среда ненасыщенная, в отличие от Калифорнии.
Объединение особей в промискуитетные группы (вместо моногамных пар) и тем более готовность молодых стать помощником на какой-либо территории вместо поиска мест для самостоятельного размножения вызвано собственными преимуществами жизни в группе, а отнюдь не отсутствием «вакансий». Среди них – снижение риска гибели и увеличение репродуктивного успеха в сравнении с поиском мест в чужой группе или моногамными парами. Действительно, в Калифорнии помощники есть лишь у 50% групп.
Те же выгоды группового образа жизни действуют и в горных лесах Колумбии, где подвид M.f.flavigula не зависит от желудей, не запасает их. Дятлы предпочитали древостой с преобладанием дуба Quercus humboldti, хотя обитают в районах, где дуб отсутствует, и на пастбищах с отдельно стоящими деревьями. Андские дятлы питаются преимущественно соком дубов, их серёжками, нектаром Spirotheca и Ochronia lagopus (Bombacaceae), насекомыми и только пот ом – желудями и др. плодами. Разнообразие и достаточное обилие корм позволяет поддерживать группы без запасания желудей. Как и везде, все члены группы участвуют в насиживании, долблении дупла и выкармливании птенцов.
В Калифорнии успешность размножения 83%, когда группы сохраняются до весны, 20% — при несохранении. Некоторые (крупные) «амбары« имеют >30000 лунок, средние — 10000, в других популяциях — существенно меньше. В Аризоне из-за редкости и низкой продуктивности дубов дятлы не создают запасов и не долбят лунок, а прячут жёлуди в естественные трещины. Запасами этого не назвать: жёлуди кончались уже осенью, и птицы бросали территории, мигрируя в Мексику. Весной они гнездились отдельными парами на территориях, не связанных с прошлогодними. Помощников здесь нет, молодняк исчезает с одной территории вместе со взрослыми. В Калифорнии он остаётся как минимум до весны, если дольше, то превращается в помощников. На родной территории молодняк размножается лишь в случае гибели взрослых противоположного пола.
Групповой образ жизни, как везде, неотделим от социальной дезорганизации, пропорциональной числу особей одного пола и интенсивности взаимодействий между ними. Здесь это уничтожение яиц и птенцов дятлами в общем гнезде, часто случающееся при совместном размножении двух самок — сестёр.
От моногамных пар к полигинадриии с помощниками — прочие виды
Аналогичное усложнение социальных систем (от моногамных пар (1) через полиандрические группировки (2) к полигинно-полиандрическим «коммунам» с «помощниками» (3)) — отмечено у лесных завирушек оседлого английского подвида Prunella modularis occidentalis при круглогодичном обитании в ботсадах и других «зелёных островах» города, также при изобилии корма и отсутствии хищников[2]. Частота (1) и (2) примерно одинакова, (3) встречается несколько реже: в последних 2-х случаях одна или же две-четыре самки обитают на совместном участке 2-3 самцов.

Лесная завирушка
Те же формы социальной организации фиксируются у лесных завирушек в других частях ареала, хотя и несколько реже.
«Почти полный контроль гнёзд в 2005-2007 годах [в заповеднике Кивач] показал, что кроме моногамных пар у завирушек были довольно широко распространены брачные группировки, состоявшие из одной самки и двух самцов. В 2005 г. из 2 проконтролированных гнёзд одно принадлежало паре, а другое – группе из двух самцов и самки. В 2006 г. зарегистрировано 6 пар и 1 «трио», имевшее 2 успешных гнезда. Особенно много гнёзд с «помощниками» оказалось в 2007 г. В этом сезоне 5 найденных гнёзд принадлежало группам из самки и двух самцов. Возле шестого гнезда наблюдали даже трёх самцов, хотя постоянно держались поблизости лишь два из них. Ещё в двух случаях можно было предполагать наличие самца-«помощника», во всяком случае, пение второго самца отмечали в непосредственной близости гнезда. Таким образом, в 2007 году брачным трио принадлежало большинство (от 6 до 8) из 11 гнёзд, а в сумме за 3 года – около половины – 9-11 из 21 гнезда, находившегося под наблюдением. Кроме того, 6 полиандричных группировок мы зарегистрировали в «Киваче» в 1982-2001 годах.
Кладку у завирушки насиживает только самка (Яковлева 1983; Tomek 1988), в выкармливании птенцов принимает участие самец. У моногамных птиц в гнёздах с 8-10-сут птенцами самец и самка приносили корм в гнездо примерно с равной частотой (Tomek 1988, наши данные). В «трио» поведение и степень участия самцов в выкармливании птенцов значительно варьировали. Из 12 групп, за которыми проводилось наблюдение, оба самца принимали участие в выкармливании лишь в 6 случаях. Однако, как правило, один из самцов, а иногда и оба прилетали к гнезду с кормом значительно реже самки.
Как установлено в Великобритании, у лесной завирушки один из защищающих совместную территорию самец (α) доминирует над другим (β) (Birkhead 1981), причём β-самец принимает участие в выкармливании только в том случае, если до этого спаривался с самкой, т.е. мог быть отцом птенцов (Davies 1983; Honotte 1989). Нам удалось зарегистрировать конфликты между самцами, которые выявили доминирующих особей, у 5 гнезд. Однако однозначной зависимости степени участия самца в выкармливании птенцов от степени его доминирования в группе не наблюдалось. У α-самца, по крайней мере в мае-июне, часто значительную часть времени занимало сопровождение кормящей птенцов самки и ухаживание, и его участие в выкармливании было очень незначительным (1 гнездо), или не наблюдалось вовсе, и птенцов кормили лишь самка и β-самец (2 гнезда). Тем не менее, следует заметить, что эти наблюдения проводились незадолго до вылета птенцов первого выводка, и вскоре самка могла приступить к откладке яиц в новом гнезде. Возможно, что в этот период самец охраняет её более тщательно, чтобы предотвратить спаривание с β-самцом, тогда как в другое время уделяет большее внимание выкармливанию птенцов.
По нашим наблюдениям, некоторые самки начинали вторую кладку уже через 4-6 дней после вылета первого выводка, когда возраст птенцов составлял всего от 14 до 16 дней. Это подтверждают наблюдения за двумя гнездами, принадлежавшими одной и той же группе, состоявшей из двух самцов и одной самки. В первом цикле размножения в то время, когда птенцам было 10-11 сут, их кормила преимущественно самка (82% прилётов с кормом) и в незначительной степени – β-самец (18% прилётов, n = 111). Самец-доминант сопровождал приносившую корм самку, ухаживал за ней, гонял другого самца, однако близко к гнезду обычно не подлетал.
Подросших птенцов второго выводка α-самец кормил даже чаще, чем самка (54% прилётов с кормом к 4-сут птенцам, n = 28 и 74% – к 9-суточным, n = 54). «Помощник» же продолжал принимать в выкармливании незначительное участие и незадолго до вылета птенцов (22 июля), видимо, оставил гнездо. В 2 гнездах из 5, наоборот, β-самец в кормлении птенцов почти или совсем не участвовал. Он лишь изредка появлялся вблизи гнезда, а в одной из групп пытался ухаживать за самкой в тех случаях, если она прилетала к гнезду одна.
Значительно реже пар и полиандричных трио мы регистрировали бигамных самцов. Нам известно всего 2 случая, когда самец выкармливал птенцов в двух гнёздах, сроки размножения в которых перекрывались. В одном таком случае (в Приладожье) гнёзда одного и того же самца располагались в 550 м друг от друга, причём между ними были ещё 2 гнезда лесной завирушки, размножение в которых проходило в эти же сроки. Во втором случае два гнезда самца находились на смежных участках и дистанция между гнёздами не превышала 150 м. Разница в сроках вылупления птенцов в гнёздах одного и того же самца составляла 9 дней в первом случае и 11 дней во втором.
Ещё одну из зарегистрированных нами репродуктивных групп, возможно, следует отнести к полигинандричным группировкам. Самец из пары, пойманной возле одного из гнёзд, через 4 дня был отловлен в составе полиандричного трио (разница в возрасте птенцов составляла всего 5 дней, расстояние между гнёздами – около 200 м), а позднее – и у второго выводка в составе этой же группы.
В Великобритании, где образование репродуктивных групп у завирушек наиболее обычно, около половины самок размножается в составе группировок, состоящих из самки и 2-3 самцов. Почти столь же часто встречается моногамия, реже – полигиния и полигинандрия (Birkhead 1981; Davies 1983, 1985; Davis, Lundberg 1984; и др.). Широкому распространению полиандрии способствует преобладание самцов в популяции. Оно возникает из-за того, что в зимних группировках лесных завирушек, которые там оседлы, самцы доминируют над самками, что приводит к более высокой смертности последних (Birkhead 1981). В Карелии, несмотря на отличие условий существования (завирушки улетают на зиму, а гнездятся в естественных ландшафтах со значительно более низкой плотностью), были обнаружены те же формы брачных отношений, причём кроме моногамии также преобладала полиандрия. То, что полиандрия не была обнаружена в Приладожье (Яковлева 1983), объясняется, видимо, использовавшимися методиками. Отлов родителей у гнёзд тогда производился бойком. При этом отлавливались лишь активно кормившие птенцов взрослые птицы. Регулярные же наблюдения у гнёзд за мечеными особями, которые могли бы выявить «помощников», не проводились. Вполне вероятно, что и в «Киваче» применявшиеся методы не могли выявить все случаи полиандрии.
О наличии второго самца мы судили главным образом на основании наблюдений у гнёзд с подросшими птенцами, при этом иногда его присутствие проявлялось лишь в токовании у гнезда и попытках ухаживать за самкой. Поскольку со второй половины июля откладка яиц прекращается и токование угасает, обнаружить таких особей очень трудно. Возможно, что может оставлять гнездо и самец, участвовавший в выкармливании птенцов. Начиная с 19 июля у части гнездящихся самцов мы регистрировали начальные стадии линьки.
Как показали исследования, выполненные на других видах, у многих воробьиных птиц Карелии при позднем гнездовании самцы, начиная линьку, часто переставали кормить птенцов (Зимин 1988).
Гнезда с «помощниками» обнаружены у лесной завирушки и в других частях ареала, хотя и несколько реже. Так, в Финляндии на 38 пар было 3 полиандричных группы (Tuomenpuro 1989), в Польше помощник отмечен лишь у одного из 12 обследованных гнёзд (Tomek 1988).
Высокую встречаемость коммунальных ячеек у завирушки в Великобритании иногда связывают с высокой плотностью там её населения (Tomek 1988). Плотность населения завирушки в Великобритании достигает 5 пар/га (Vogel, Tuomenpuro 1997), а величина гнездового участка составляет лишь 0.1-0.6 га (Snow, Snow 1982).
В Северной и Восточной Европе плотность населения этого вида существенно ниже, а величина гнездовых участков – выше. В Польше пара занимает участок около 2.5-3 га (Tomek 1988), в Финляндии – 1.54 га (Tuomenpuro 1989). В Карелии плотность населения P. modularis также значительно ниже, чем в Великобритании, а величина гнездовых участков (от 1.5 до 3 га) сходна со значениями, полученными в Финляндии и Польше. Однако, как показывают наши данные, и в условиях низкой плотности гнездового населения полиандрия в отдельные годы может быть вполне обычной. Мы наблюдали гнёзда с помощниками как в биотопах с высокой плотностью завирушек (берег Суны с густым подростом ели), так и на территориях с низкой плотностью гнездования этого вида (сосновые боры), на участках, которые занимались не ежегодно. Из-за небольшого объёма материала за отдельные годы нельзя говорить о большей распространённости полиандричных групп в Карелии, чем в других местах на континенте, т.к. соотношение пар и полиандричных групп может варьировать по годам. Во всяком случае, на Британских островах выявлена значительная межгодовая изменчивость структуры репродуктивных ячеек, что объясняют повышенной смертностью самок в суровые зимы (Birkhead 1981).
Яковлева М.В., 2008. О формах брачных отношений у лесной завирушки//Русский орнитологический журнал. Т.17. Экспресс-выпуск 408. С.452-458.
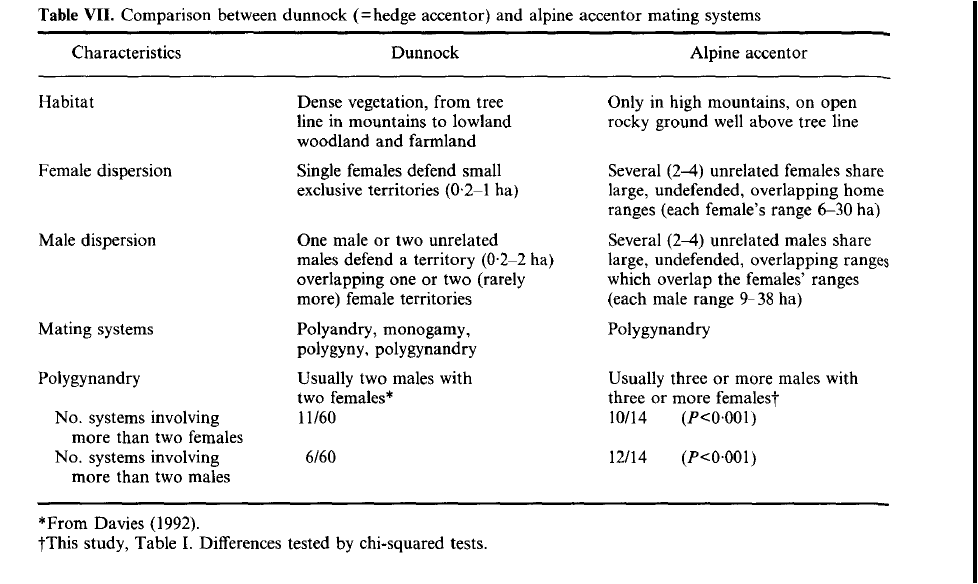
Позднее такое же усложнение «конструкции» социума было найдено у альпийской Laiscopus collaris, бледной и других видов завирушек[3]. См. сравнительную таблицу их социальных систем: в более суровых условиях высокогорья второй вид демонстрирует большую «коммунальность» и меньшее разнообразие брачных связей (только полигинандрия без её вырожденных производных, как у лесной зарушки), при полностью редуцированной территориальности.
 Подобных примеров множество не только у птиц, но и у млекопитающих — скажем, у тех видов полёвок, что исходно считались моногамными, на деле реализуется всё разнообразие брачных отношений, производное от полигиандрии-полиандрии (сосновая) или полигинандрии-полигинии (прерийная полёвка, полёвки китайская и Брандта).
Подобных примеров множество не только у птиц, но и у млекопитающих — скажем, у тех видов полёвок, что исходно считались моногамными, на деле реализуется всё разнообразие брачных отношений, производное от полигиандрии-полиандрии (сосновая) или полигинандрии-полигинии (прерийная полёвка, полёвки китайская и Брандта).

Альпийская завирушка
Мы видим, что прогресс в изучении социальной организации позвоночных заставил этологов вслед за выявлением «теневой стороны социальности» ввести сходное понятие «потенциальная часть социальной организации вида». Как непроявленная в природе часть интеллекта включает более сложные способности и/или более тонкие «чувства», чем присутствующая везде и всегда, так «скрытая» часть социальной организации включает «конструктивно наиболее сложные» отношения. В преобладающих условиях обитания они неустойчивы и/или требуют большей избыточности ресурсов для своего поддержания (притом что социальное поведение вообще – «предмет роскоши», развёртывающийся лишь при условии некоторого заведомого избытка ресурсов, иначе реализуется «экономный» вариант распределения территорий, образования пар и пр., без отношений и социальной коммуникации).
Соответственно, наиболее обычные, повсеместно встречающиеся формы социальной организации соответствующего вида представляют собой не самостоятельный феномен, но «обломки» или отдельные аспекты более сложной системы, существующей скорей идеально (тут просится на ум слово «архетип») и восстанавливаемой в хотя столь же «идеальной» процедуры реконструкции, для которой ползучего эмпиризма недостаточно, он ведёт к ошибкам. И эта более сложная «конструкция», демонстрирующая социальную систему во всей полноте, целиком воплощается достаточно редко, лишь при особых условиях или только у некоторых видов (вроде желудёвого дятла, сравни его социальную систему с «моногамией с исключениями» у пёстрых дятлов, разбираемых ниже). У большинства видов, которым она в принципе присуща, её надо реконструировать по редким уклонениям от наиболее частого состояния (однако не случайным, а системного характера).
Важность дедуктивной реконструкции «типа» социальной организации
Наши примеры показывают, что «потенциальная социальность» перечисленных видов восходит к одному и тому же плану строения — полигинно-полиандрической системе, с помощниками или без[4]. Однако он не всеобщий — из анализа В.В.Иваницким эволюционных факторов, вызывающих переход певчих птиц от клаустрофильного гнездования с равным или преимущественным участием самца в гнездостроении и выкармливании птенцов, к открытым чашеобразным гнёздам, строящимся только самкой, понятно, что «коммуны» и «полигинандрия» в сверхблагоприятных условиях характерны только для первой группы видов и исчезают во второй. Что мы и видим в действительности.
Важно подчеркнуть, что у меньшинства «несоциальных» видов тех же таксонов напрочь отсутствуют потенции развития более сложных социальных систем, чем фиксируется наблюдателем в «среднем случае». Даже там, где местные условия создают значительный избыток пищи или же в ряде местообитаний плотность повышена, особи не усложняют структуру отношений, а дифференцируют активность так, чтобы меньше встречаться. См. примеры с тамарисковой и полуденной песчанкой в противоположность краснохвостой и монгольской. «Избыточная социальность» фиксируется лишь у социальных видов (прошу извинить за тавтологию), т.е. социально связанных и социально зависимых во взаимодействиях друг с другом (в т.ч. существенно более зависимых от сигналов сообщества, будь то долгосрочные элементы сигнального поля или текущие демонстрации компаньонов по взаимодействию, чем от внутренних побуждений).
Что отсюда следует? Как минимум, индуктивный метод в реконструкции социальных систем недостаточен, как максимум – он приводит к ошибкам, даже когда не сводится к «ползучему эмпиризму». Масса «обычных случаев» давит немногочисленные «уклонения». Однако последние — не простые статистические единицы, а как бы «прожектор», в свете которого видим (полностью или частично) ту самую наиболее сложную конструкцию социальной системы вида, чьим производным будут все «массовые« и «средние» случаи.
Т.е. искомый результат нашего анализа — «конструкция» социальной системы вида, всё равно, взятая в описании или в начертании. Она представляет она собой идеальное, «типическую» структуру связей, соединяющих «типичные» позиции в системе, и в «средних случаях» проявляющаяся лишь частично или не всегда. Чем сложнее конструкция «типа», тем (при прочих равных) реже встречаются ситуации, в которых он «вырисовывается» полностью, и исключительней условия, в которых это бывает. Поэтому, не рассчитывая на полную реализацию «типа», приходится восстанавливать его «по кусочкам».
Каковы брачные отношения пёстрых дятлов — «моногамия с исключениями» или полигинандрия?
Скажем, пёстрые дятлы (рода Dendrocopos и Picoides), в отличие от желудёвого не запасают корм. Здесь доминирующий вариант социальности — одиночная территориальность во внегнездовое время, и парная — в гнездовое. Моногамные пары возникают объединением одиночных участков самца и самки: в «среднестатистическом» варианте объединённый участок пары (гнездовой) самцы защищают от самцов, а самки от самок. Хотя при сильной угрозе захвата со стороны соседей или вселенцев возможна взаимная координация усилий между партнёрами, и члены пары сигнализируют друг другу о степени угрозы, требует она этой координации или нет. См. данные по пушистопёрому дятлу Picoides pubescens.
Данная схема преобладает настолько, что многие авторы, следуя «ползучему эмпиризму», не только утверждают отсутствие иных территориальных и брачных отношений, но и выдумывают «убедительное» социобиологическое объяснение, почему иное невозможно. Скажем, Клаус Михалек и Ганс Винклер сперва не нашли случаев внебрачной копуляции у больших пёстрых дятлов и средних дятлов в значительной выборке гнездящихся пар (как и отклонений от моногамии у их членов). После чего предположили что, поскольку у этого вида (как и других пёстрых дятлов с доминированием самки, см. ниже) дупло строит и о кладке/птенцах заботится преимущественно самец, при таком крупном родительском вкладе, мол, невыгодно трахаться на стороне. Дальше довольно-таки быстро выяснилось, что это не так[5]; но даже тогда, в 1990-е годы, было видно, что из такой модели невозможно понять, как возникает социальная система краснохохлого D.borealis – одного из пёстрых, но однотипная таковой желудёвого, с той только разницей, что самка в «коммуне» одна (на мой взгляд, он — также как южноамериканские формы ligniarius и mixtus — ближе к восточноазиатским острокрылым дятлам, чем к другим североамериканским пёстрым).

Краснохохлый дятел, головы самца и самки
Он также живёт коммунальными группами (до 6 особей), но с одной самкой, с которой спаривается преимущественно доминирующий самец — но иногда также и бета-особи. Сплочённость группы минимальна во время насиживания, выкармливания молодых и в конце лета, когда птицы из её состава часто отсоединяются, чтобы долбить дупла. Пространство защищают все члены группы, включая подросших птенцов. Последние часто остаются на родной территории (самцы сильно чаще чем самки) и в статусе «помощников» выкармливают следующие выводки. Переход в репродуктивный статус связан с занятием «вакансий» размножающихся, освободившихся в собственной или соседней группах (то и другое примерно с равной частотой). Размножающиеся самцы обычно пожизненно привязаны к своей территории, самки подвижней — 17% из них ежегодно переселяются в одну из соседних групп. Причём в 34% случаев они оставляют прежнего самца, и в 59% им удаётся размножиться в новой группе. За 1979-1989 гг. не отмечено появления групп на новых территориях, только преемственная смена особей в существующих (Источник).
Краснохохлые дятлы заботятся о птенцах, более родственных им, хуже, чем неродственных, притом что присутствие помощников сильней всего влияет на выживание потомства. Величина и качество территории группы — много слабее. Одна и та же группа в разные годы может иметь или не иметь помощников. Возраст самок в группе положительно коррелировал с успешностью размножения, если помощников не было, и отрицательно — если были. Причём в группах со старой самкой помощники были потомством обоих родителей, с молодой — только самца. В группах из >2-х птиц несколько самцов спариваются с единственной самкой (источник).
Сегодня мы знаем, что соответствующие данные были неполны, а объяснения вроде приведённого выше просто не соответствуют фактам. У всех видов пёстрых дятлов, кто специально исследовался в этом плане, обнаружены системные отклонения от парной территориальности с моногамией, причём закономерно следующие из системы брачного доминирования в паре – у одних видов доминирует самец, у других – самка, что и структурирует «разделение труда» между партнёрами на разных этапах репродукции.

Виды с доминированием самок эпизодически «уклоняются» в сторону полиандрии или её «облегчённого варианта» — сдвоенного гнездования, когда самка спаривается с двумя самцами и несётся в 2 разных гнезда, одну кладку насиживает она сама, другую — её самец (тогда как часть яиц оплодотворена «чужим»). См. соответствующие данные для большого пёстрого дятла (1-2).
Это происходит в собственном поселении, на «лицевой» стороне социальности вида, но также присутствует «теневая», представленная дальними рейдами резидентов за пределы участка. Здесь выделяются подчинённые самцы, хуже защищающие общую территорию пары (их доминирующие самки всего чаще гоняют). Они постоянно совершают дальние выходы (1-5 км) за пределы участков и в других поселениях пробуют занять территорию и сближаться с самками, особенно зимовавшими вдали от них и прибывшими в пятна лучших гнездовых биотопов (где зимовка почти невозможна) позже всех, в начале мая. Эта политерриториальность установлена точно; вероятно, такие сближения иногда ведут и к реальному спариванию, т.е. полигинии.
Иными словами, в регулярных «уклонениях« обоих групп видов дятлов с альтернативной организацией брачного доминирования «прорисовывается» та самая полигинно-полиандрическая система отношений, которая в полной мере реализована у желудёвого и краснохохлого дятлов. Только здесь она как бы «размазана» во времени и пространстве; больше того, на гнёздах у всех видов дятлов, в том числе обоих описываемых здесь групп эпизодически присутствуют и помощники.
По всей видимости, полигинно-полиандрические группы с помощниками представляют собой архетип социальной организации всех дятлов (по крайней мере исследованных в этом отношении, см. данные по золотому дятлу Colaptes auratus). У всех видов дятлов, пёстрых уж точно, есть потенция развития в эту сторону, просто не реализующаяся в обычных условиях их обитания, где низкая плотность населения и/или межгодовая неустойчивость популяционных группировок реализуют почти исключительно упрощённый вариант.
Известное исключение составляют виды с наибольшим развитием доминирования самок (большой пёстрый с сирийским). До образования пары, в период взаимного поиска и «тестирования« потенциальных партнёров самцы интенсивно конкурируют за внимание самок; если их больше одного, самка вполне равнодушно ждёт выявления «лучшего» в ходе довольно-таки долгих конфликтов и с ним уже начинает сближаться, не стесняясь отвешивать тумаки, если он уменьшает дистанцию быстрей, чем интенсифицирует демонстрирование. Там самец, «натравливаемый» самкой на потенциальных захватчиков, после образования пары чем дальше тем больше берёт на себя охрану всего пространства участка, а территориализм самок редуцируется. То и другое обычно случается через неделю-две после объединения участков и перемещения пары, подыскивающей места для гнезда, из токовой зоны территории — куда самец со своего участка летал ухаживать ещё до образования пары в зону постройки гнезда, обычно на территории самца, куда самец переводит самку.
Другое же — хоть и менее выраженное — исключение представляют собой виды с доминированием самцов — белоспинный, трёхпалый и в меньшей степени малый пёстрый дятлы. В период поиска партнёров самки здесь конкурируют за внимание самцов, достаточно равнодушно ждущих разрешения их конфликтов. Однако же эти виды, в противоположность предшествующей категории — В-, а не А-стратеги по классификации В.В.Иваницкого (1981). Успешное развитие брачной (и территориальной) коммуникации здесь ведёт к урежению соответствующих взаимодействий, снижению «эмоциональности« участников, рост эффективности коммуникации сопрягается с падением интенсивности и напряжённости взаимодействий между соответствующими категориями компаньонов, а не их интенсификацией при росте возбуждённости, как у А-стратегов.

Токующий малый пёстрый дятел. Фото В.В.Солодушкина.
Поэтому образующиеся пары рассредотачиваются тем быстрей, чем успешней идёт токование, что быстро прекращает территориальную агрессию даже у малого дятла, где она хорошо выражена. У трёхпалого с белоспинным участки практически неохраняемые, охраняется лишь индивидуальная дистанция — самками в брачный период предсказуемо яростнее, чем самцами, и то лишь в начале ухаживания.

Самец белоспинного дятла. Фото В.В.Солодушкина
По этой причине у видов с доминированием самцов ждёшь «уклонений» от моногамии и парной территориальности, противоположных таковым видов с доминированием самок. Действительно, у самого территориального из них — малого дятла — наблюдается территориальная полигиния, размножение самцов с двумя самками. То же самое, но пореже, видим у белоспинного дятла. «Теневая сторона» социальности состоит в том, что пары, ведомые подчинённой самкой (брачные связи здесь сильно прочней, чем у видов предыдущей категории, поэтому выходы совершаются не в одиночку) совершают исследовательские выходы.
Особенно это выражено у трёхпалого дятла, меньше — у белоспинного, всего хуже — у малого[6]. При ухудшении условий обитания на прежней территории это ведёт к реальному переселению; другой, существенно более редкий вариант — самка из «выходящей» пара сближается с одиночным самцом из соседнего поселения. Единично это ведёт к спариванию и действительной полиандрии; особенно если переселения не произошло, и пара по завершении выходов возвращается к себе на участок[7].

Самка трёхпалого дятла с кормом. Фото В.В.Солодушкина
Однако в большинстве случаев «собственный« самец эффективно защищает самку от нового (на время формируется трио, где первый доминирует над вторым, не давая сближаться с ухаживаниями). По-видимому, шансы «второго» самца тем ниже, он тем раньше «отстанет« от пары, чем дальше продвинулась коммуникация в ней, в том числе чем старше «первый« самец и самка. Пока это предположение, основанное лишь на отдельных наблюдениях и требующее проверки.

Самец большого пёстрого дятла кормит птенца. Фото В.В.Солодушкина
Вышеописанная «конструкция» брачных отношений у пёстрых дятлов не вполне верно называют «моногамией с уклонениями»; ошибка состоит в том, что акцентируется «моногамия», как будто бы именно она — тип отношений для данной группы. На деле ж системный характер уклонений указывает на иной тип (ту самую полигинандрию), просто оставшийся потенциальным в связи со слишком напряжённым существованием и/или низкой плотностью. Что лучше всего видно у малых пёстрых дятлов, где выраженность доминирования самца и самки в паре сравнима[8], и сильно варьирует между разными парами. Здесь 8.5% самок демонстрируют политерриториальную полиандрию, и одновременно 2.9% самцов – политерриториальную полигинию, хотя, как у всех видов с доминированием самки, верность партнёру высока (ежегодно распадается ~3.4%) и пары сохраняются несколько лет.
Оба типа политерриториальных уклонений от моногамии – полиандрия и полигиния – встречаются при отклонении соотношения полов от 1:1 и именно у тех особей противоположного пола, которые заняли наилучшие территории/находятся в лучшем физическом состоянии. Однако у малого дятла также фиксируются «коммуны» с двумя самками, выкармливающими птенцов в одном гнезде, и помощниками.
Самки в полиандрических трио выкармливают на 39% больше чем моногамные; хотя они распределяют усилия между разными гнёздами более-менее поровну, каждому достаётся меньше, что компенсируется пропорциональной интенсификацией родительской заботы со стороны самца. Полигинические самцы большую часть заботы тратят на первое гнездо и выкармливают меньше потомства, чем моногамные (в других районах столько же).
…и тот же вопрос применительно к певчим птицам
Сходные «уклонения» от моногамии изучены в поселениях мухоловки-пеструшки в Томске, в самом городе и в ближних пригородах, существующих в условиях повышенной плотности (1-2-3). В обоих преобладали моногамные пары (87,2% и 61,6% соответственно). Оставшиеся демонстрировали полигинандрическую систему отношений: самцы одновременно спаривались с несколькими самками (в т.ч. благодаря политерриториальности), а самки последовательно — с несколькими самцами. Одновременно самцы могут выкармливать чужих птенцов в собственном гнезде или играть роль «помощников» у соседних пар. Самки могут а) принимать кладки от другой самки на стадии инкубации; б) подкладывать яйца в гнёзда тем самцам, с которыми копулировали; в) завершать прерванные кладки, подкладывая яйца к другим парам мухоловок-пеструшек.
Фактически тут мы видим «коллективное гнездо», вроде существующего в коммуне желудёвых дятлов, только «разорванное» во времени и пространстве. Это даёт независимое подтверждение верности определения начальной стадии в эволюции социальной организации певчих птиц у В.В.Иваницкого.
Поэтому реконструкция социальной системы вида требует много большего, чем обобщение всех непосредственно наблюдаемых случаев. Нужен гипотетико-дедуктивный метод, когда разнотипным местным реализациям, как при складывании паззла из кусочков, восстанавливается в первом приближении то самое идеальное (формы отношений), которое вслед за Ю.М.Плюсниным зовём архетипом.


Дальше оно используется как гипотеза, в большей или меньшей степени объясняющая социальную динамику во всех многочисленных местных реализациях (отдельных группировках). По обнаруживающимся невязкам исправляется идеальное (формы отношений, которые используем как объяснительный принцип, рабочую гипотезу, принимаемую или отвергаемую исходя из степени соответствия эмпирическим данным), и вновь сопоставляется с данными и пр. Через несколько таких итераций данный метод «нащупает» общую схему «конструкции» социальной системы вида; так происходит восхождение от наблюдаемых взаимодействий между особями к структуре отношений, существенной для вида в целом. В большинстве случаев она будет сложней наблюдаемых частных реализаций, оставляя место для «потенциальной социальности».
Вышеописанные усложнения в таком случае оказываются «окошком», через которое непосредственно наблюдается часть этого самого идеального, облегчая путь гипотетико-дедуктивному методу.
Примечания
[1] Важный момент здесь — бОльшая социальность грача в сравнении, например, с галками. Им, как и ворОнам с ворОнами, свойственно «утешение жертвы» после конфликтов. У галок оно отсутствует, а у грачей играет важную роль в репарации социальных связей, «нарушенных» чьей-то агрессией. Это ещё раз подтверждает важную закономерность о причинно-следственной связи между развитием социальности вида и когнитивным развитием составляющих его особей.
[2]Они, как и специализированные конкуренты, существенно отстают от своих жертв в освоении урболандшафтов. Что выступает одним из «плюсов» последнего, толкающим к урбанизации «дикие» виды птиц и млекопитающих. См. «Город как заказник-2»
[3] Davies N. B., Hartley I. R., Hatchwell B. J., Desrochers A., Skeer J., Nebel D. 1995. The polygynandrous mating system of the alpine accentor Prunella collaris. I. Ecological causes and reproductive conflicts// Animal Behaviour. V.49. P.769-788; Hartley I. R., Davies N. B., Hatchwell B. J., Desrochers A., Nebel D., Burke T. 1995. The polygynandrous mating system of the alpine accentor Prunella collaris. II. Multiple paternity and parental effort// Animal Behaviour. V.49. P.789-803.
[4]Важно отметить, что данный тип отношений эволюционно наиболее древний для птиц: он присущ дневненёбным (страусовым, тинами) и сохраняется достаточно долго как плезиоморфия в наиболее древних ветвях более продвинутых групп, вроде трёхпёрсток из журавле-, якан и цветных бекасов из ржанкообразных и пр. В исходном варианте самец присоединяется к группе самок и формирует гарем; те несутся в общее гнездо, а самец заботится о потомстве. Самки могут затем переходить к другим самцам, как у нанду и ряда видов тинаму (Crypturellus boucardi, Nothoprocta cineraceus), а могут оставаться с самцом, осуществляя тот или иной вклад в инкубацию, как у африканского страуса и тинаму Nothocercus bonapartei. Второе вероятней при территориальности гаремных самцов. В другом случае самки не объединяются в группе и держатся поодиночке. После спаривания с привлекшими их самцами они переходят к другим самцам, а те привлекают новых самок (тинаму Crypturellus variegatus) или остаются с самцом у гнезда (эму, казуары). Забота о потомстве и здесь преимущественно лежит на последнем.
Т.е. исходная система размножения характеризуется одновременной полигинией самцов и последовательной полиандрией самок, часто с реверсией половых и родительских ролей. Она в полном виде сохранилась у современных тинаму, эму и казуаров (хотя здесь токуют самцы). Упрощением этой системы получаем 1) гаремную полигинию как у нанду и африканского страуса (с факультативной полиандрией); во втором случае самец делит родительские обязанности с «главной самкой гарема»; 2) моногамию (с эпизодической полиандрией) как у киви и ряда видов тинаму. При этом у тинаму, казуаров, киви и моа половой диморфизм в размерах и окраске обратный, у африканского страуса и нанду — прямой, у эму — нейтральный. См. Панов Е.Н. Социальные системы палеогнат как исходное состояние для класса птиц в целом; Коблик Е.А. Эволюция и диверсификация палеогнат: вероятный сценарий// Древненёбные птицы (очерки филогении, систематики, морфологии и хозяйственного использования). М.: Т-во научных изданий КМК, 2010. С.21, 49-50.
Хотя у наиболее молодых групп куликов (песочники), эта система вторично возникла из территориальной моногамии. У дятлов, о которых речь ниже, это скорей всего сохранившийся примитивный признак, как и у предковой для них группы афроазиатских бородастиков (американские бородастики ближе не к дятлам, а к туканам).
[5] Напротив, у среднего дятла Dendrocopos medius до сих пор не нашли каких – либо отклонений от моногамии, что хорошо согласуется со вторичной редукцией брачного доминирования в парах этого вида, отсутствием зимней территориальности и очень поздней консолидацией пар этого вида – лишь во время гнездостроения. До этого самцы и самки, не связанные территориальными запретами, обследуют значительное пространство, активно сближаются между собой, формируют временные пары, и до начала выдалбливания дупла меняют по нескольку партнёров. См. Бутьев В.Т., Фридман В.С., 2005. Средний пёстрый дятел// Птицы России и сопредельных регионов. Совобразные-дятлообразные. М.: Т-во научных изданий КМК. С.371-383
[6]У него же относительное участие полов во второй половине выкармливания птенцов ближе всего к таковому у видов с доминированием самок («надёжные отцы» и «приспосабливающиеся или дезертирующие матери»). В более ранний период выкармливания и инкубации участие самок примерно соответствует таковому самцов или несколько больше; дупло строит преимущественно самец и он же насиживает ночью. См. 1-2-3.
[7] См. Бутьев В.Т., Фридман В.С. Белоспинный дятел. Трёхпалый дятел// Птицы России и сопредельных регионов. Совообразные-Дятлообразные. М.: Т-во научных изданий КМК. С.383-397, 423-433.
[8]в т.ч. самцы за самок конкурируют с той же интенсивностью, что самки за самцов, в отличие от большого пёстрого дятла, с одной стороны, и белоспинного – с другой.
Рекомендуем прочесть
 Как Европа открыла для себя Московскую Русь, а потом закрыла
Как Европа открыла для себя Московскую Русь, а потом закрыла COVID-19 вскрывает правду
COVID-19 вскрывает правду «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве «Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)
«Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)- Социальная сегрегация в городах
 «Жесты», вόроны и шалашники
«Жесты», вόроны и шалашники- Банковское дело и эволюция целей банковского регулирования
 Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей
