
Н.Н.Мешкова, Е.Ю.Федорович
Замечание и незамечание новизны особями разных рангов
«Разнообразное и продолжительное обследование новой территории, периодически возобновляющееся ее переобследование позволяет синантропным грызунам — домовой мыши, серой крысе хорошо знать окружающую обстановку и реагировать на малейшие изменения в ней. Как показали исследования,, эти животные замечают не только появление, но и исчезновение, переустановку предметов и даже замену их на сходные (Мешкова и др., 1992; Федорович, Мешкова, 1992). После замечания изменения зверьки сразу же прерывают «повседневную» деятельность (например, кормежку, патрулирование территории), а вся активность как бы «стягивается» к местам изменений, обследование и переобследование которых возобновляются несколько раз.
Эксперименты с изменением знакомой зверькам среды позволили выявить закономерности развертывания исследовательского поведения в этих условиях, а также проследить особенности построения и функционирования образа среды у животных, его зависимость от характеристик особи, ее мотивационного состояния, особенностей жизнедеятельности (Федорович, Мешкова, 1992; Федорович, в печ.).
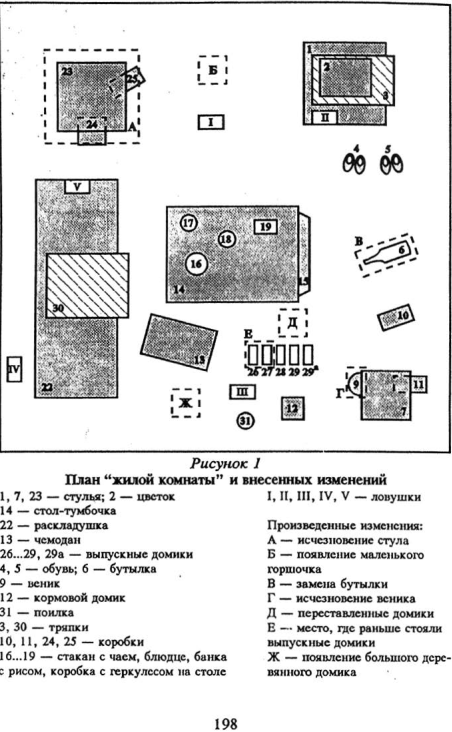
В выгородке 4×4 м воссоздавалась «жилая комната», имитирующая жилище человека. В помещение одновременно выпускали группу незнакомых между собой животных. Через 4 — 5 дней «комната» полностью функционально ими осваивалась, отношения в группе стабилизировались; в дальнейшем работа велась только с группами, в которых формировались четкие иерархические отношения с деспотическим доминированием одного из самцов.
На шестой день после выпуска животных в обстановке «комнаты» производились одновременно 7 изменений следующих типов: исчезновение, замена, перестановка, появление предметов. Основная часть материала излагается по дипломной работе Е. Ю. Федорович, выполненной на факультете психологии МГУ. Поведение серых крыс в сходных условиях изучалось студенткой биологического факультета МГУ А. В. Белоусовой.
Мы предположили, что ранг животного в группе ставит перед особью специфические задачи жизнедеятельности, определяя его образ жизни, систему отношений с различными компонентами среды. Это и обусловливает в значительной мере особенности функционирования и развития у каждого зверька регулирующего его поведение психического отражения, образа среды.
Для установления влияния ранга зверька на вероятность возникновения и характер протекания ориентировочно-исследовательской деятельности в каждой группе мы анализировали поведение двух самцов — доминанта и активного подчиненного (субдоминанта). Эти две категории зверьков были выбраны по следующей причине: при выраженном отношении «доминирования — подчинения» между ними эти особи имели приблизительно равную двигательную активность и при этом перемещались по всей территории. Иерархическое положение среди самок, за редким исключением, было выражено неявно; у малоактивных подчиненных — «изгоев» — активность ограничивалась небольшими участками помещения и сводилась в основном к быстрым перебежкам от убежища к местам кормежек, поэтому в большинстве мест появления изменений они не бывали.
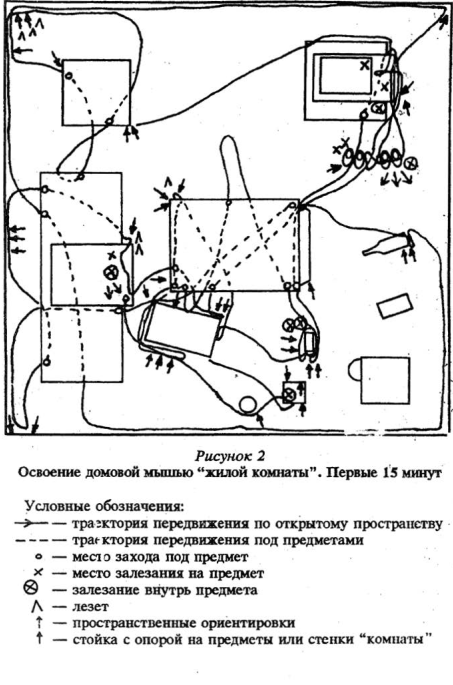
В результате проведенных наблюдений было выявлено, что доминанты и активные подчиненные отличались по целому ряду количественных и качественных поведенческих показателей по отношению к изменениям.
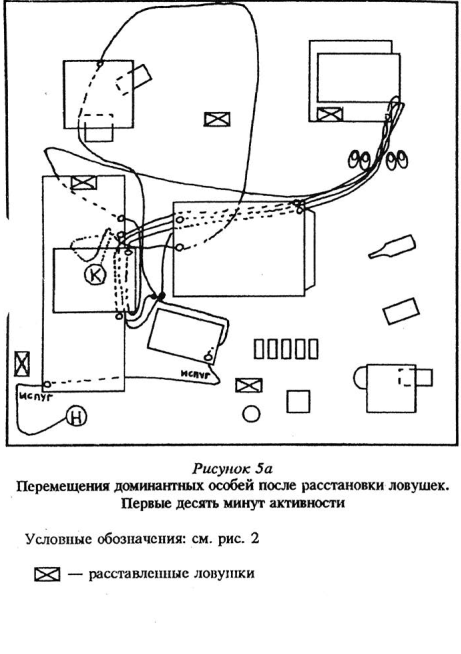
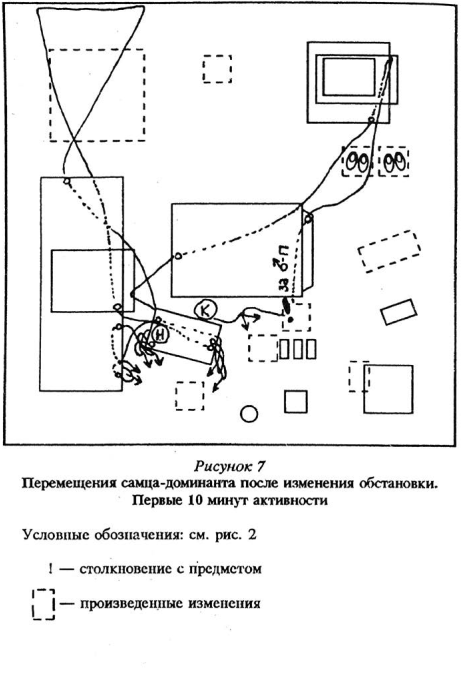
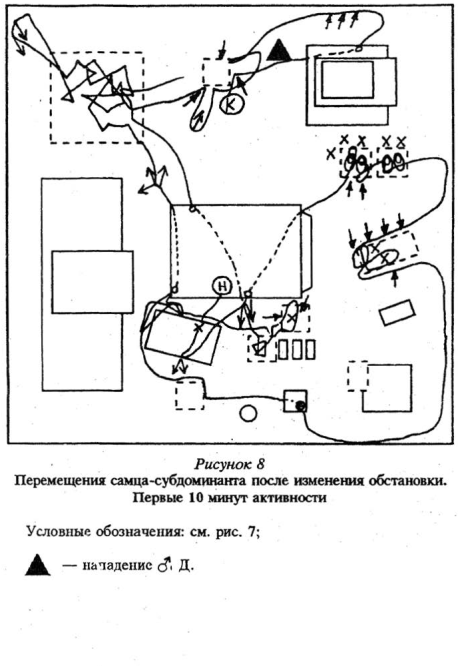
Доминантные особи, возобновляя активность в измененной среде, обнаруживали изменения через больший промежуток времени, чем субдоминантные, при этом лишь на 2 — 5-й раз нахождения рядом с местами, где они произошли. В целом при первом подходе доминантные самцы заметили 19% изменений, а активные подчиненные — субдоминанты — 75%. См. рисунки 5 — 8 Приложения, там же план «жилой комнаты».
Следует отметить, что чаще всего с первого раза замечались те изменения, которые появились на открытом месте в наиболее посещаемых животными местах, рядом с центрами активности и маршрутами передвижений. Однако однозначной зависимости моментального замечания изменения от места его появления мы не выявили.
Случаи незамечания «новизны» мы объединили, проанализировав ситуации нахождения зверьков рядом с нею, в 4 группы.
1. Доминантные особи не замечают появления изменений, преследуя, разыскивая подчиненных зверьков или самок. Вот несколько примеров. Самец-доминант два раза пробегал рядом с появившимся домиком на звук прыжков подчиненного, но заметил его, только наткнувшись на этот предмет в ходе погони. Доминант из другой группы «охотился» за подчиненным, который как раз в это время ел приманку с только что поставленной ловушки (плоской давилки Геро). Будучи нацелен на объект своей «охоты», доминант погнал подчиненного прямо в ловушку. И только в следующий раз, находясь в этом же районе, доминирующий самец заметил появление ловушки и обследовал ее. Этот же доминант прошел, почти касаясь боком другой ловушки, идя по следу подчиненного, но заметил и начал обследовать ее, только возвращаясь обратно. Много раз мы наблюдали, как доминанты, пробежав в ходе погони за каким-либо членом группы по всей «комнате» и, соответственно, мимо большинства изменений, никак не реагируя на них, » спокойном состоянии замечали их, проявляя неофобию.
Так, один из доминантов даже напал на зверька, сидящего на новом цветочном горшочке, не проявив по отношению к новому предмету неофобии, хотя до этого несколько раз «не решался» подойти к нему. Нередко доминанты замечали появившийся или переставленный предмет только в результате непосредственного столкновения с ним опять же при погонях или в результате нападения на других членов группы, обследующих в этот момент эти предметы. В таких случаях преследование прекращалось, и доминантный самец или испуганно отскакивал от «неожиданно возникшего» предмета, или приступал к его осторожному обследованию.
Иногда доминантный самец в возбужденном состоянии принимал новый предмет за искомого подчиненного и атаковал его. Так, самец-доминант три раза подряд налетал на небольшой цветочный горшочек и кусал его. То же наблюдали и у серых крыс — доминантный самец, после выяснения отношений с крысами из соседней группировки подошел в угрожающей позе к новой бутылке и пытался ее атаковать.
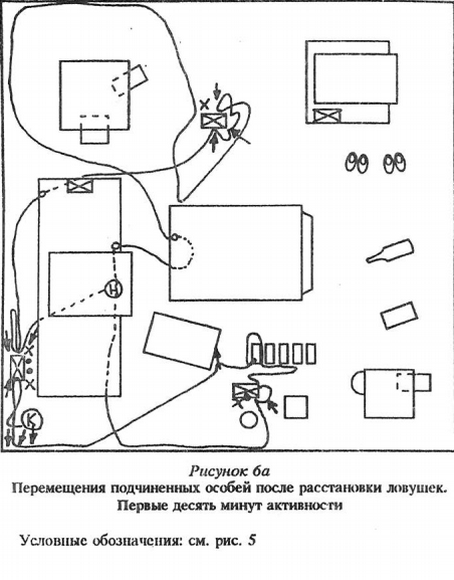
2. Подчиненные мыши пробегали мимо произведенных изменений, спасаясь от преследования. Целый рад примеров дает нам поведение малоактивных подчиненных, «изгоев». Эти зверьки, перебегая по коротким маршрутам от убежища к еде и обратно, не реагировали, как правило, на появившиеся изменения. Один из таких зверьков, имея убежище в одном из выпускных домиков, не «заметил» исчезновения других, стоявших рядом, хотя выбегал из своего убежища 5 раз за время наблюдений.
3. Мыши (чаще подчиненные) пробегали мимо изменений, целенаправленно перемещаясь из убежищ к кормовым точкам после сна или при активизации доминанта — в убежища.
4. И доминанты, и подчиненные зверьки не замечали одни изменения, обследуя другие.
Так, один из самцов прошел в первый раз мимо нового домика, отвлекшись на обнюхивание следов, оставленных наблюдателем. Мы видели, как обследование зверьком одного, важного для выполнения его жизненных функций, изменения, долго «не давало возможности» заметить и/или обследовать другое, расположенное рядом. Так, трое подчиненных мышей из разных групп продемонстрировали одну и ту же закономерность: каждый из них проходил в десяти сантиметрах от появившегося цветочного горшочка (в других ситуациях он замечался с 1 метра), ориентируясь в сторону места, где раньше находился стул, на котором у этих особей было убежище.
Возвращаясь обратно, эти мыши замирали перед «неожиданно» возникшим перед ними горшочком, отбегали в сторону, после чего подходили и обследовали его. В нескольких группах замечание доминантами появления нового домика рядом с их убежищем (чемоданом) на все три часа наших наблюдений отвлекло этих зверьков от обследования других изменений. Попытки подобраться к этому предмету с разных сторон (домик вызвал сильную неофобию) перемежались с погонями и поиском подчиненных по всей «комнате». Однако доминанты вновь и вновь возвратились к попыткам начать обследование этого предмета, никак не реагируя на появление других изменений. Один из доминантных зверьков нашел и несколько раз обследовал прокопанную дикими мышами нору под сетчатым полом «комнаты». Выход этой норы располагался между местом, где находился убранный нами стул, и поставленным горшочком. На эти изменения доминант внимания не обратил и их не обследовал.
А. В. Белоусовой описан случай, когда крыса, подбираясь к сильно пугавшей ее верши, прошла по стоявшей на ее пути новой ловушке, пс заметив ее. Впоследствии ловушка сильно пугала эту самку.
Таким образом, мы видели, что изменения могут замечаться или не замечаться с первого раза в зависимости от мотивационного состояния животного, направленности его деятельности в данный момент. Прежде всего замечаются изменения наиболее значимые для каждой особи в этот самый момент. Н.Ю.Войтонис (1949) рассматривал подобное незамечание животным даже биологически значимых изменений в среде как проявление установки, мобилизацию организма на какую-то одну, актуальную в данный момент деятельность.
Хотя доминантные особи и перемещались по всему объему «жилой комнаты», в целом они заметили не все произведенные изменения — в среднем 4,0 из 7 возможных; разброс — от 1 до б. В то время как подчиненные зверьки заметили и обследовали в среднем 6,4 из 7 возможных изменений, разброс 3 — 7. Чаще всего доминантные особи замечали и проявляли наиболее выраженную неофобию и исследовательское поведение по отношению к появившимся впервые или на новом месте предметам; исчезновение, а также замену предметов, если эти изменения не были связаны с их жизнедеятельностью (например, в переставленном домике могло находиться убежище), они не замечали, либо обследовали крайне мало (1 — 3 подхода). Субдоминантные особи замечали изменения всех типов, кроме исчезновения небольших предметов, если только они не были связаны опять же с их жизнедеятельностью.
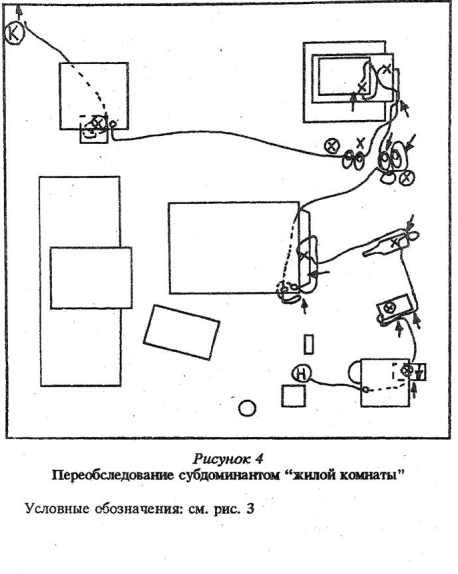
Достаточно неожиданным для нас было увидеть, насколько тщательно и разнообразно интересно обследуется этими зверьками место, с которого убирался какой-либо предмет. Вот конкретный пример. Из комнаты выносили стул. Мышь, пробегая от этого места на расстоянии примерно полутора-двух метров, неожиданно останавливаясь и развернувшись головой в сторону убранного предмета, замирала. Затем следовала серия ориентировочных стоек из разных мест, при этом мышь крутилась, перебегая от укрытия к укрытию по большому диаметру вокруг. Потом с большей или меньшей настороженностью зверек подходил непосредственно к участку пола, над которым стоял стул, и проходил его, делая частые мелкие зигзаги — нюхая пол и меняя направление движения через 2 — 3 шажка. Иногда зверек останавливался и делал серию пространственных ориентировочных стоек в разных направлениях. Иногда — отбегал в сторону, ориентировался там, лотом снова подходил к измененному месту, проходя его мелкими зигзагами (см.рис. 8 Приложения).
Еще один пример. Исследователи убирали веник, стоящий прислоненным к ножке одного из стульев. Чаще всего исчезновение этого предмета мыши замечали с сиденья стула, над которым возвышалась ручка веника. Мыши крутились на краю сиденья, заглядывая вниз, перемежая это ориентировочными стойками, отбегали на другой конец сиденья, ориентировались оттуда в сторону, где ранее возвышался веник. Затем зверьки поднимались и спускались несколько раз по ножке стула, к которой был прислонен веник и, в заключение, обходили, делая мелкие зигзаги, тот участок на полу, на который непосредственно опирался веник.
Когда передвигали часть выпускных домиков, зверьки обследовали как сами передвинутые домики, так и то место, с которых они убраны.
В целом для активных подчиненных зверьков типичными были случаи замечания изменений издалека, а также целенаправленные подходы к ним, как правило, без выраженных признаков иеофобии. Обследование «новизны» — продолжительное (в среднем 10 — 12 секунд, в отдельных случаях до трех, и даже шести минут (!) непрерывно) , цепочки действий по обследованию при первом подходе состояли из 4 — 10 действий. Как мы уже говорили, доминантные зверьки дольше не замечали произведенные изменения, часть обнаружений изменений у них была связана либо со случайным столкновением с появившимся или переставленным предметом, либо с подражанием другим особям — доминант первый раз подходил к появившемуся предмету, предварительно пронаблюдав за другими зверьками или зверьком, его обследовавшими. В 95% случаев первый подход доминантов к изменениям следовал уже после того, как рядом побывали и обследовали его другие мыши.
Яркой отличительной особенностью поведения этой категории зверьков на начальном этапе ознакомления с произошедшими изменениями была сильная неофобия по отношению к ним. Отдельные особи в течение всех трех часов наблюдений так и «не решались» подойти хотя бы к одному из поставленных нами новых предметов. Однажды мы наблюдали проявление иеофобии по отношению даже к пустому месту, с которого были убраны выпускные домики. Примечательно, что нсофобия по отношению к произведенным изменениям проявлялась не только до первого обследования их. Часто у доминантных особей мы замечали чередование незамечания изменения при пробегании мимо (при слежке или погоне за подчиненным или самкой в эструсе) с проявлением по отношению к нему же, при другом подходе, неофобии. Один из доминирующих самцов проявлял сильно выраженную неофобию по отношению к переставленным домикам, так и «не решившись» подойти к ним, отскакивая всякий раз к ближайшему укрытию. Когда же он следил, пробегая от предмета к предмету, за одним из подчиненных самцов, спрятался за этими выпускными домиками без каких-либо признаков боязни. Это не помешало доминанту настороженно отнестись к ним при последующем возобновлении их обследования.
Проявление сильно выраженной неофобии к новым предметам у доминантных самцов серых крыс, в отличие от других членов группы, в сходных экспериментальных условиях описано и А.В.Белоусовой. Частично вследствие иеофобии, первые контакты с «новизной» у доминантных зверьков были фрагментарными (1 — 3 секунды), цепочка действий по их обследованию состояла из 1 — 2 элементов, ориентировочно-исследовательская деятельность при последующих подходах разворачивалась крайне медленно. Более подробно явление неофобии будет описано ниже.
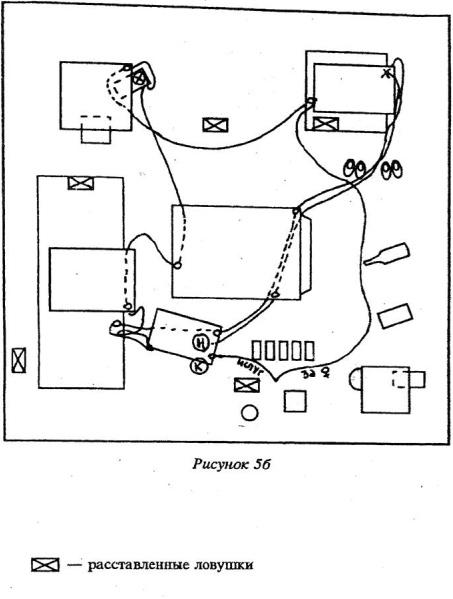
То, что субдоминантные и доминантные особи замечали различное количество изменений и, кроме того, активные подчиненные, в отличие от доминантов, замечали и обследовали преимущественно все типы изменений, можно объяснить различной представленностью, дифференцированностью элементов окружающей среды в психическом образе этих зверьков. Косвенно это подтверждается и особенностями характера передвижений зверьков обеих этих категорий до дня изменений, в стабильной, знакомой обстановке.
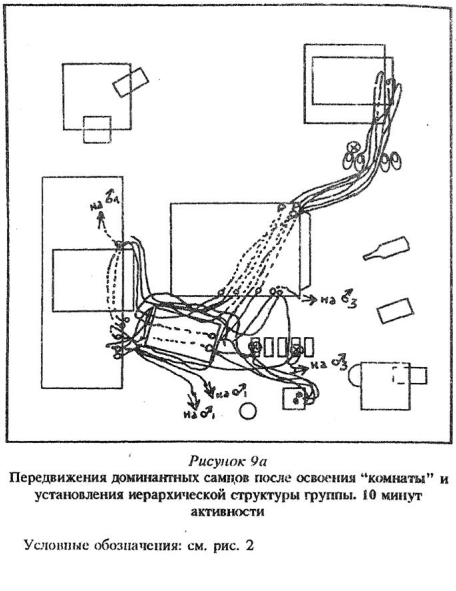
В целом передвижения доминантных мышей отличались меньшей вариативностью, при спокойной обстановке в помещении они бегали по одним и тем же маршрутам, соединявшим важные для них участки, убежища, места кормежек и питья. Передвижения но таким маршрутам были стремительны, при этом мыши достаточно стереотипно обегали предметы с одной и той же стороны, уходили в убежища и выходили из них в одних и тех же местах и одним и тем же способом. С набеганных маршрутов доминантные особи сходили в основном в двух случаях: во время погони или слежки за какой-нибудь мышью, а также при патрулировании территории, причем доминант обходил по очереди те места, где раньше обнаруживал искомую особь, или шел по следам убежавшей мыши.
Так как расположение предметов не менялось во всех группах, для доминантов из разных групп в целом были характерны похожие маршруты передвижений (рис. 9 Приложения).
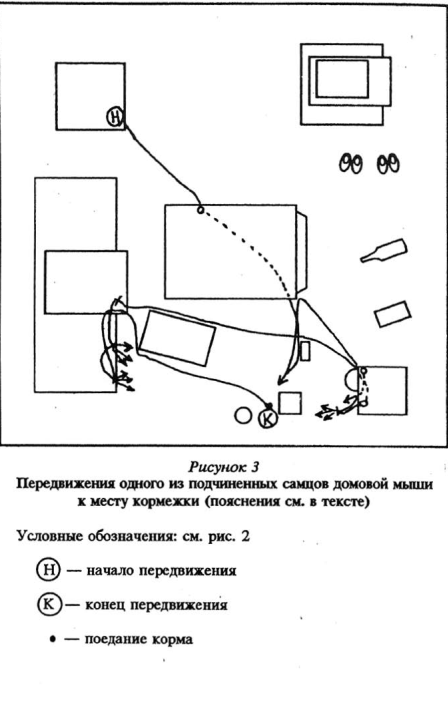
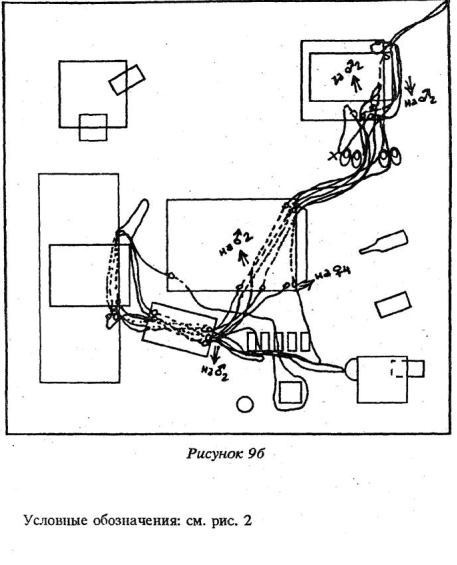
Для активных подчиненных зверьков, субдоминантой, было характерно следующее: после выхода из убежища и подходов к пище и воде они, как правило, обходили территорию. Траектории подчиненных особей не складывались в «тропы», как у доминантов, а располагались «диффузно» (рис.10 Приложения). У подчиненных также были короткие набеганные маршруты, проходя которыми они передвигались стереотипно. Подчиненные зверьки пользовались ими, в противоположность доминантам, чаще всего когда ситуация в помещении становилась напряженной: эти маршруты связывали места кормежки и убежища.
Кроме того, именно у субдоминантиых особей, как было уже описано выше, мы наблюдали интереснейшую форму поведения — переобследование знакомой, хорошо известной территории — эти зверьки обходили комнату с «привязкой» своего движения к практически всем наземным предметам, находящимся в помещении, переобследуя при этом и сами предметы: влезали внутрь, сверху, обходили по периметру, обнюхивали. Можно предположить, что подобное переобследование приводило к более детальному ознакомлению с предметной обстановкой «жилой комнаты», а также способствовало в дальнейшем замечанию этими зверьками любых изменений окружающей среды.
Вернемся к поведению зверьков в измененной нами «жилой комнате». Благодаря активному, разнообразному и продолжительному обследованию появившихся изменений, субдоминантные особи быстро корректировали свое поведение, «втягивая» обследованные изменения обстановки в сферу своей жизнедеятельности: например, своевременно обегая их новые или переставленные предметы при погонях, используя их как новые убежища, наблюдательные пункты, первыми начинали кормиться с расставленных ловушек (ненастороженных в наших экспериментах).
Другая же категория — доминанты на протяжении довольно длительного времени не учитывали произошедших изменений в своей жизнедеятельности. Так, в ходе погонь за другими членами группы они нередко сталкивались с появившимися или переставленными предметами. Например, один из доминантов сшибал выпускной домик, передвинутый на новое место, три раза подряд в ходе одной и той же погони. Мы наблюдали также, как после неоднократного обследования какого-либо изменения доминанты, подбегая к нему с другой стороны, «пугались» его (повторно проявляли неофобию).
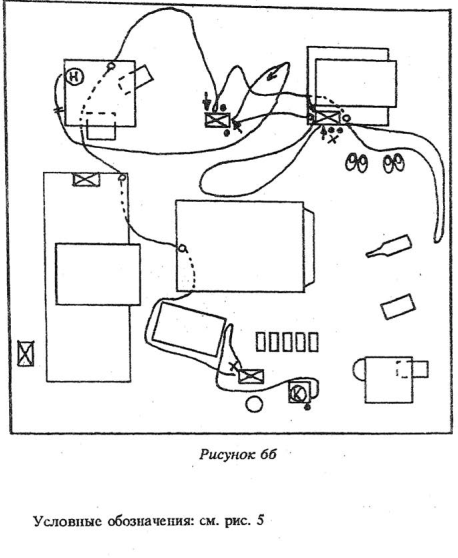
Одним из важных факторов, определяющим особенности ориентировочно-исследовательской деятельности зверьков в измененной среде, явился характер использования места, на котором произошло изменение, его значимостью для конкретной особи. Изменения, производившиеся в центре зоны активности данного зверька, например рядом с его убежищем, замечались (через больший или меньший отрезок времени) в 100% случаев как активными подчиненными, так и доминантами. Например, переставленные на место друг друга две пары обуви замечались и обследовались только теми доминантами, чье убежище находилось рядом — в тряпке, свисающей со стула, хотя все доминанты бывали в этом районе в течение нашего наблюдения неоднократно. Исчезновение веника замечали многие подчиненные зверьки, однако продолжительно и неоднократно это место обследовали только тс особи, которые имели убежище на стуле, к которому этот веник был прислонен,— они обходили зигзагами участок пола, на который опирался веник, пролезали вверх-вниз по ножке стула, ориентировались с места сиденья, над которым возвышалась ручка веника. Вообще произведение изменений рядом с убежищами или местом кормежки вызывало как наиболее сильную неофобию, так и наиболее сильно выраженную исследовательскую реакцию.
Таким образом, как показали вышеприведенные данные наблюдений, развертывание ориентировочно-исследовательской деятельности в условиях, близких к естественным, не происходит автоматически в ответ на любые изменения в знакомом животному окружении. Характер поведенческих ответов зверьков в ситуациях «новизны» во многом определяется не столько формальными признаками самих изменений (характер изменений, их размер и т. п.), сколько особенностями жизнедеятельности самою зверька (в нашем случае — общим контекстом деятельности в момент нахождения животного рядом с изменением, его потребностно-мотивационным состоянием, характером использования территории, на которой произошло изменение, значимостью произошедшего изменения для животного).
Некоторые из приведенных выше наблюдений позволяют сделать заключение об активном характере отражения зверьками окружающей среды, проявляющемся в данном случае прежде всего в избирательном, «пристрастном» характере восприятия изменений.
Активный характер психического отражения выражался прежде всего в различной степени готовности особей воспринимать изменения окружающей среды, что обусловливалось, по нашему мнению, значимостью самого факта появления изменений для животного. Большая готовность подчиненных зверьков к восприятию и учету изменений проявлялась в следующем: активные подчиненные особи, для которых значимым являлся поиск любых новых возможностей приспособления к существующим условиям через установление новых связей со средой (прежде всего поиск новых источников пищи и убежищ), замечали большинство изменений с первого раза, как правило, издалека, после чего целенаправленно подходили к нему. (Некоторые мыши, забираясь на высокий предмет — тумбочку, ориентировались сверху, после чего слезали и ниш прямо к какому-либо изменению, например на место убранного стула.)
Доминанты, положение которых в группе зависело от поддержания существующей иерархической структуры в гpyппе и значительная часть активности которых сводилась к поиску, преследованию подчиненных, долго не замечали произошедшие изменения, в целом видели меньше изменений, долго не учитывали их в своей жизнедеятельности.
Нам кажется также, что тот факт, что для подчиненных в целом не было свойственно проявление неофобии по отношению к изменениям, еще раз подчеркивает повышенную готовность этих зверьков воспринимать внесенные изменения. Единичные случаи проявления иеофобии у подчиненных относились к ситуациям неожиданного для животного замечания предмета, в том месте, где, предполагалось, его нет. Потенциальную готовность подчиненных к восприятию изменений мы можем увидеть и в описанных выше случаях регулярно повторяющихся нереобследованиях комнаты предметов, находящихся в ней.
[Думаю, именно здесь кроется известное этологам превосходство «ума» и «гибкости» подчинённых особей над «силой» доминантов, которое, на мой взгляд – биологическая предпосылка успеха свойственной людям борьбы с угнетением. Прим.публикатора]
Группа случаев, когда обследование одного, более значимого изменения «не давало» животному заметить другие, рядом расположенные, а также случаи, когда доминант, преследуя или отслеживая подчиненных, не замечает «новизны» в окружающей обстановке, еще раз подтверждает активный характер регулирующих поведение психических процессов.
Представленный материал, как нам кажется, убедительно показывает, что животные не просто осуществляют выбор поступающих стимульных воздействий, последовательно отфильтровывая их по «регистрам» — новизне, интенсивности, неопределенности значимости (Соколов, 1960; Гращенко, Латаш, 1965; Barry, 1990), но сами активно вычерпывают из окружающей среды нужную информацию, нужный стимул (Леонтьев, 1979; Смирнов, 1985; Вилюнас, 1986; Latash, 1990). Причем «тот стимул, который ищется, уже заранее имеет интерпретацию, значение…» (Смирнов, 1985, С.6). То, какая информация, какие объекты, их свойства и связи будут восприняты особью, то, каким содержанием наполнится психический образ, определяется самим животным, всем контекстом его деятельности, его образом жизни.
Неофобия
Обсуждая поведение синантропных грызунов в ситуациях появления или перестановки предметов в знакомой животным обстановке, необходимо отдельно остановиться на форме поведения, проявляющейся в избегании животным какой-либо «новизны» — неофобии. Наглядно это проявляется следующим образом. Так, серые крысы при клеточном содержании подолу избегают подходить к новой кормушке или старой, но повешенной в новом месте. В естественных условиях серые крысы перестают пользоваться тропой на сутки и более, если возле нее появляется новый предмет или меняется положение старых (Baniett, 1963).
Подробное описание проявления домовыми мышами неофобии к новым, измененным элементам среды дано в предыдущей главе. По мнению известного ученого в области поведения сииантронных грызунов С.Барнетта, неофобия — это защитный механизм, выработавшийся у грызунов, у которых избегание новых предметов (а ими часто оказываются ловушки и отравленные приманки) обеспечивает выживание в условиях непрекращающейся борьбы человека с ними.
Однако, как показывает целый рад исследований, феномен неофобии не такой простой, каким может показаться на первый взгляд, и не сводится к таким определениям как
«небольшие изменения окружающей обстановки вызывают исследовательское поведение, а значительные — избегание» (Хайнд, 1975),
или «появление нового предмета на освоенной территории вызывает у синантропных крыс и мышей его избегание» (Barnett, 1963).
Оказывается, что неофобия при одних и тех же изменениях в обстановке может проявляться в большей или меньшей степени, вообще отсутствовать или проявиться лишь у части животных. На то, будет ли животное избегать произошедшие изменения и как долго, влияет целый ряд факторов, и не последнюю роль здесь играют индивидуальные особенности зверьков, их предыдущий опыт, положение в структуре группы, их актуальное потребностно-мотивационное состояние и т. п.
Разберемся более подробно, что же обусловливает тип поведенческого ответа синантропных грызунов в ситуациях подобного рода.
1. Объем пространства и его сложность. Чем оно больше и сложнее, тем менее выражена исофобия в ситуации изменения обстановки (Мешкова, 1989).
2. Продолжительность пребывания зверька в освоенном им пространстве (Мешкова, 1985, 1989), знание им территории (Мешкова, 1990; Misslin, Ropartz, 1981). Если животные обнаруживали новый предмет в хорошо освоенной ими камере или вольере, то они, как правило, избегали приближаться к предмету довольно длительное время. Если же помещение недостаточно знакомое, предмет не вызывал боязни и зверьки практически сразу же приступали к его обследованию.
3. Стабильность обстановки. Неофобия будет тем сильнее, чем меньше изменялась обстановка перед этим (Мешкова, 1989; Федорович, Мешкова, 1992).
В естественных условиях эту закономерность описал Бойс (Boicc, 1971). Серые крысы, которые живут на свалках, где окружающая обстановка постоянно меняется, не обладают столь характерной для их сородичей, обитающих в домах, настороженностью по отношению к ловушкам и легко в них ловятся.
4. Характер использования животными того места, где появился незнакомый предмет или произошла перестановка (Лялин, 1974; Мешкова, 1985, 1990; Мешкова и др., 1992; Barnett, 1963). Наиболее сильную и продолжительную неофобию вызывает тот предмет или их перестановка (и даже исчезновение), которые появились вблизи убежища, норы или тропы или маршрута передвижений зверьков, т. с. непосредственно рядом с зоной активности данной особи.
5. Проявление неофобии зависит от ранга особи, от его групповой роли. Как подробно показано выше, была выявлена следующая закономерность. У домовых мышей из групп с деспотическим доминированием одного из самцов наиболее выражена неофобия у доминантных особей, наименее — у субдоминантов, активных подчиненных (Мешкова и др., 1983; Федорович, Мешкова, 1992).
Сходные данные получены А. В. Белоусовой для серой крысы. Наряду с этим имеются наблюдения, свидетельствующие, что первыми к новой кормушке или приманке подходят доминантные самцы серых крыс (Galеf, 1983), после чего уже и другие крысы начинают пользоваться новым источником корма.
6. Проявление неофобии по отношению к изменениям в окружающей знакомой обстановке зависит от мотивационного состояния зверька в момент нахождения рядом с «новизной» (Федорович, Мешкова, 1992) (более подробно см. раздел 5.1.2.).
Несомненно, проявление неофобии в ситуации появления или перестановки предметов обстановки показывает, что тот или иной зверек заметил произошедшее изменение, вследствие чего прерывается предыдущая деятельность животного. Однако какой поведенческий ответ последует после того, зависит от совокупности приведенных выше факторов.
Исходя из представлений о функционировании психического отражения (Леонтьев, 1983; Смирнов, 1985; Вилюнас, 1986), можно предположить, что степень проявления неофобии, ее длительность, сила будут тем более выражены, чем более значительным будет рассогласование между сложившимся психическим образом среды с настоящим чувственным отражением. Это зависит, как мы видели, от:
1. знания животным окружающей обстановки, степени ее изученности (п.п. 1, 2);
2. от значимости произошедшего изменения для животного (п. 4), а также
3. от того, какая часть психического образа актуализирована в данный момент (п.п. 5, 6). Опережающий характер функционирования психического образа наглядно выступает, если проанализировать природу проявления неофобии в одних ситуациях и ее практически полное отсутствие в других или у определенной категории особей (п.п. 4, 5, б). Неофобия проявляется в момент и как следствие рассогласования «ожидаемого, прогнозируемого будущего» с реальным чувственным опытом. Подтверждением этому служит, например, та закономерность, что чем более стабильна обстановка,’ тем наиболее сильно выражена неофобия у животных в случае ее изменения. В ходе наших экспериментов, если мы вносили изменения в «обстановку жилой комнаты» в течение пяти дней подряд, неофобия у зверьков практически не проявлялась, начиная с третьих суток.
Заслуживает внимания тот факт, что наименьшую неофобию, боязнь произошедших изменений, проявили именно субдоминантиые особи домовых мышей; после короткого замирания они переходили к их обследованию. Как мы писали выше, именно у этих животных мы наблюдали периодически возобновляющееся «переобследование территории», в ходе которого животные последовательно обходили, повторно обследовали широкий ряд предметов «жилой комнаты». Создавалось впечатление, что зверек обходит хорошо знакомую ему территорию в поисках изменений, произошедших в его отсутствие. В дальнейшем именно эта категория зверьков в ситуации появления «новизны», как правило, не избегала ее, а буквально «притягивалась» к местам изменений и наиболее тщательно, длительно и разнообразно их обследовала».
Н.Н.Мешкова, Е.Ю.Федорович. Ориентировочно-исследовательская деятельность, подражание и игра как психологические механизмы адаптации высших позвоночных к урбанизированной среде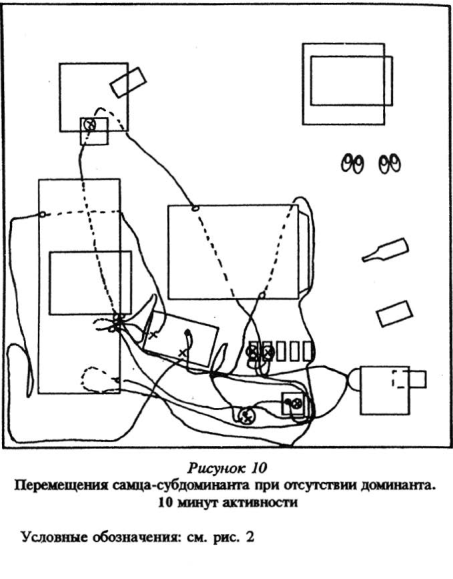 . Москва «Аргус» 1996. С.78-86
. Москва «Аргус» 1996. С.78-86
Рекомендуем прочесть
 Два механизма «копирования поведения»
Два механизма «копирования поведения» Часть и целое
Часть и целое Синдром социальности: сравнительное исследование поведения песчанок
Синдром социальности: сравнительное исследование поведения песчанок Социальный контроль поведения млекопитающих: ревизия концепции доминирования
Социальный контроль поведения млекопитающих: ревизия концепции доминирования Борьба с угнетением: биологический бэкграунд
Борьба с угнетением: биологический бэкграунд Как иерархия в группе влияет на её эффективность?
Как иерархия в группе влияет на её эффективность? Эмоции, их распознавание и дискриминация женщин
Эмоции, их распознавание и дискриминация женщин «Маккиавеллиевский интеллект» и социальное познание у приматов
«Маккиавеллиевский интеллект» и социальное познание у приматов
