

Рис. 1. Бабочка Euphydryas editha, ее исходное кормовое растение коллинсия мелкоцветковая (Collinsia parviflora) и новое растение, на которое бабочки опрометчиво перешли, — подорожник ланцетолистный (Plantago lanceolata). Фото с сайтов butterfliesandmoths.org и wikimedia.org
Резюме. Относительность адаптаций всего ярче видна в т.н. «ловушках», в которые, как рыбу в вершу, вид заводит специализация. Экологическая ловушка наблюдается в случае, когда вид, приспосабливаясь к каким-либо биотопам, способам сбора корма и пр., стал от них зависеть настолько жёстко, что при изменении других элементов ландшафта (т.ч. эти, значимые для вида, оказываются в необычном для себя окружении) не может скорректировать эти связи и «расплачивается» повышенной гибелью гнёзд, выводков, взрослых особей и т.д. именно в характерных биотопах, где всё это д.б. снижено. См. пример с ласточковым попугаем Lathamus discolor и Schläpfer M., Ruge M., Sherman P., 2002. Ecological and evolutionary traps // Trends. Ecol. and Evol. Vol.17. P.474-480.
Так, «с точки зрения вида» от периферии города к центру растут кормность местообитаний и одновременно неспецифическое беспокойство, риск гибели особей, риск разорения гнезда и т.п. эффекты «экологической ловушки», которую представляет собой город для всех «диких» видов. Периферия города более безопасна, но менее обеспечена ресурсами (Jokimäki, Huhta, 2000).
Экологическая ловушка возникает, когда вследствие изменений среды непредсказуемо меняется значение источников информации об окружающей среде, используемых живыми организмами. Обычные реакции организмов на данные стимулы тогда уже не ведут к ожидаемым результатам, так что особи избирают плохой биотоп, основываясь на тех самых ключевых стимулах, которые раньше информировали о высоком качестве. Экологические ловушки – часть более широкого феномена эволюционных ловушек, включающих любое рассогласование между стимулами, обычно используемыми видом при принятии поведенческих или жизненных решений, и результатами, ассоциируемыми с «верной» реакцией на эти стимулы (Schlűpfer et al., 2003).
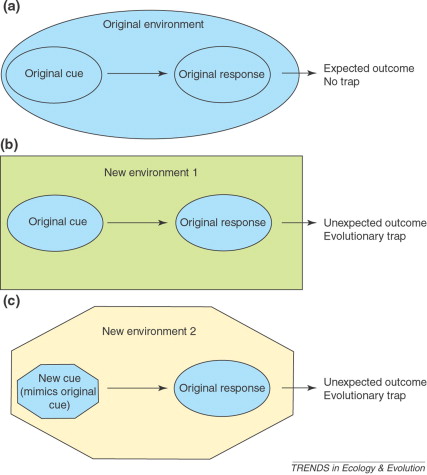
Разные способы попадания в «ловушку». Из: Schläpfer et al., 2002
Следование видовым экологическим предпочтениям и иным видовым стереотипам в городах почти всегда значит попасть «в экологическую ловушку» (Schlűpfer et al., 2003). Урбанизация «диких» видов, лабилизируя их, вид из «ловушек» выводит, почему представляет собой адаптацию широкого значения. Вообще выход возможен двумя путями — за счёт роста пластичности и за счёт естественного отбора; в первом случае сдвиг средних значений важного в этом смысле признака сопровождается ростом вариансы, во втором — уменьшении. Один и тот же вид, скажем, большие синицы Parus major, в разных районах используют оба.
Следовательно, «ловушка» — это пример, когда выработанная специализация неожиданно для вида «подводит» вследствие наступивших средовых изменений, что ставит его в рискованное положение, а то и губит. В неё «зайти» можно что на эволюционных, что на экологических масштабах времени; между теми и другими «ловушками» нет существенной разницы, тем более что недавно показано одновременное осуществление процессов приспособления и микроэволюции: они идут одинаково быстро, приспосабливаясь друг к другу, участники экологических взаимодействий (конкуренты, хищник и жертва и пр.) одновременно эволюционируют и наоборот, одно без другого не бывает.
Первую «ловушку», с бабочкой Euphydryas editha, многие видели, а вот вторая, с малым черноголовым дубоносом Eophona migratoria, явно осталась незамеченной. А она не менее показательна: вид в последние 100-130 лет успешно освоил «антропогенную саванну» на Дальнем Востоке, поднял численность, расширил ареал… но его подкосил начавшийся 30 лет в том же ландшафте назад быстрый рост численности врановых птиц, особенно сороки. Гнёзда дубоноса перед ней практически беззащитны — и прежний ареал вида распался на изолированные и сокращающиеся фрагменты. В обоих случаях вид не «оказался» в ловушке от собственной неизменности при изменениях среды вокруг а наоборот, сам «зашёл туда», быстро и эффективно приспособившись к случившимся новообразованиям… увы, преходящим.
Слишком быстрая адаптация к новым условиям завела бабочек в «эволюционную ловушку»
Многолетние наблюдения за популяцией бабочек на уединенной горной лужайке в штате Невада позволили описать новый тип «эволюционной ловушки»: в нее может попасть популяция диких животных, адаптируясь к последствиям хозяйственной деятельности человека. Выпас скота привел к распространению на лужайке подорожника ланцетолистного, на котором выживаемость гусениц оказалась выше, чем на исходном кормовом растении — коллинсии мелкоцветковой. Под действием отбора бабочки за несколько лет полностью перешли с коллинсии на подорожник, не подозревая, что оказались в «эволюционной ловушке». Ловушка захлопнулась, когда в 2005 году умер хозяин ранчо и на лужайке перестали пасти скот. Разросшиеся травы затенили подорожник, и теплолюбивые бабочки вымерли. При этом по краям лужайки оставалось много незатененной коллинсии, где бабочки могли бы жить и дальше, если бы не отказались полностью от своего старого кормового растения. Исследование показало, что способность животных быстро приспосабливаться к антропогенным изменениям среды может завести их в тупик, поскольку человек меняет среду еще быстрее, чем меняются самые быстро эволюционирующие животные.
Изменения среды, вызываемые деятельностью человека, ставят перед живыми существами трудные эволюционные задачи. Нередко в новых условиях старые адаптации, выработанные за миллионы лет эволюции, оказываются губительными. В таких случаях говорят об «экологических ловушках» или «эволюционных ловушках» <…>. Например, после того как в Австралию для борьбы с вредителями завезли ядовитых жаб Bufo marinus (см.: Ядовитые жабы оккупируют Австралию, «Элементы», 20.02.2006), под угрозой вымирания оказались местные популяции варанов: этим хищникам все их древние охотничьи инстинкты подсказывали, что новые жабы — превосходная добыча.
Другой пример — подёнки, которые находят подходящие для откладки яиц водоемы по поляризованному отраженному свету и иногда принимают за воду асфальт, который поляризует отраженный свет примерно так же (C. J. Jolly et al., 2016. The impacts of a toxic invasive prey species (the cane toad, Rhinella marina) on a vulnerable predator (the lace monitor, Varanus varius)). Считается, что если попавшая в такую ловушку популяция не вымрет полностью, то со временем она приспособится к изменившимся условиям, либо научившись не использовать неподходящий ресурс, либо выработав устойчивость к его вредоносным эффектам.
Новая статья специалистов по эволюционной экологии Майкла Сингера (Michael C. Singer) и Камиллы Пармезан (Camille Parmesan), опубликованная в журнале Nature, показывает, что животные могут оказаться в гибельной ловушке не только из-за того, что они недостаточно быстро подстраиваются к меняющимся условиям, но и наоборот, из-за слишком быстрой и успешной адаптации.
Сингер и его коллеги более 30 лет наблюдали за изолированной популяцией бабочки-шашечницы Euphydryas editha (Edith’s checkerspot butterfly) на окруженном лесом лугу в горах Невады. Владелец луга, фермер по имени Гарри Шнайдер (Harry Schneider), пас на нем коров.
Расселительная способность у бабочек E. editha очень низкая. Обычно они не улетают от места своего рождения дальше, чем на пару километров. Расстояние до ближайшей соседней популяции — около 40 км. Поэтому бабочки с лужайки Шнайдера жили практически в полной изоляции. Размножаются они один раз в год. Молодые гусеницы живут большими группами и оплетают стебли растений паутиной, что облегчает их поиск и учет.
Исходным кормовым растением для гусениц E. editha является коллинсия мелкоцветковая (Collinsia parviflora). Выпас скота привел к распространению на лугу Шнайдера (как и во многих других местах на западе США) завезенного из Старого Света подорожника ланцетолистного (Plantago lanceolata). Бабочки начали иногда откладывать яйца на новое растение, и вдруг оказалось, что оно даже лучше подходит для развития гусениц, чем коллинсия.
Дело в том, что коллинсия — однолетнее растение, которое начинает стареть и вянуть раньше, чем большинство гусениц успевает завершить свое развитие. Поэтому многие гусеницы погибают от голода, так и не окуклившись. Подорожник, напротив, растение многолетнее. Он не так быстро вянет, и, хотя гусеницы на подорожнике растут медленнее, чем на колинсии, их выживаемость в итоге оказывается намного выше.
Ранее авторы детально изучили взаимоотношения между гусеницами и растениями, пытаясь понять, почему бабочки не адаптировались к быстрому увяданию своего кормового растения. Ведь они, казалось бы, могли это сделать, просто ускорив собственное развитие, то есть окукливаясь раньше. Ученые пришли к выводу, что причина в «эволюционном компромиссе» (см. Trade-off) между выживаемостью гусениц и плодовитостью взрослых бабочек. Если гусеница будет окукливаться раньше, она не успеет набрать достаточную массу. Вышедшая из куколки бабочка получится худосочной и отложит меньше яиц. По-видимому, для E. editha высокая плодовитость оказалась важнее, чем выживаемость личинок. Молодые самки этого вида настолько массивны, что даже летают с трудом, пока не отложат первые порции яиц.
Поэтому появление нового кормового растения, на котором гусеницы могут развиваться дольше, не рискуя погибнуть голодной смертью, стало для бабочек настоящим подарком. Они же не знали, что этот подарок у них скоро отберут. Начиная с 1980-х годов Сингер и его коллеги год за годом наблюдали, как бабочки на лугу Шнайдера стремительно (по эволюционным меркам) переходят с коллинсии на подорожник.
Исходно бабочки E. editha предпочитали откладывать яйца, естественно, на коллинсию. Эксперименты с бабочками из «наивных» популяций (живущих там, где нет подорожника) показали, что при наличии выбора 80% самок уверенно выбирают коллинсию, а остальные 20% не проявляют избирательности и откладывают яйца на колинсию и подорожник с равной вероятностью. Такова, по-видимому, исходная ситуация с изменчивостью по предпочтениям, характерная для бабочек, никогда не видевших подорожника.
В 1982 году на лугу Шнайдера были зарегистрированы первые самки, предпочитающие подорожник коллинсии. Эволюционный переход на новое растение начался. К 1990 году доля самок, предпочитающих подорожник, достигла 50%. Было показано, что предпочтения определяются генетически (наследуемость признака равна 0,9). Кроме того, потомство самок, предпочитающих подорожник, стало расти на подорожнике быстрее, чем гусеницы, матери которых предпочитали коллинсию. Это должно было дополнительно повысить репродуктивный успех бабочек, выбирающих подорожник, и тем самым ускорить эволюционный переход.
Эксперименты, проведенные в 2005 и 2007 годах, показали, что теперь уже 100% самок на лугу Шнайдера предпочитают подорожник. Старое кормовое растение было полностью забыто. Все гусеницы развивались теперь на подорожнике.
Еще в 1993 году Сингер с коллегами предсказали, что наблюдаемый ими эволюционный процесс может завести бабочек в смертельную ловушку. Быстро и успешно адаптируясь к антропогенным изменениям среды, насекомые рискуют попасть в зависимость от продолжения людьми тех же практик (в данном случае — от продолжения выпаса скота). Это серьезный риск, потому что, как бы быстро ни эволюционировали насекомые, поведение человека может меняться еще быстрее (M. C. Singer et al., 1993. Rapid human-induced evolution of insect–host associations).
В новой статье авторы с мрачным торжеством сообщают, что их предсказание сбылось. В конце 2005 года Гарри Шнайдер умер, ранчо было продано, выпас скота прекратился, на лугу разрослись буйные травы, и весь подорожник оказался в тени. Эволюционная ловушка захлопнулась.
Средняя дневная температура на листьях подорожника, куда бабочки откладывали яйца, понизилась на 7°C, а освещенных участков открытой почвы практически не осталось. Это оказалось губительным для теплолюбивых гусениц. Они уже не могли, как раньше, нежиться на солнышке, лежа на земле рядом со своим растением. В 2007 году их видели отчаянно ползающими в густой траве в тщетных попытках найти солнечный пятачок. При этом по краям лужайки, где не было густой травы, оставалось много хорошо освещенной коллинсии. Но бабочек это не спасло, потому что они больше не использовали свое старое кормовое растение.
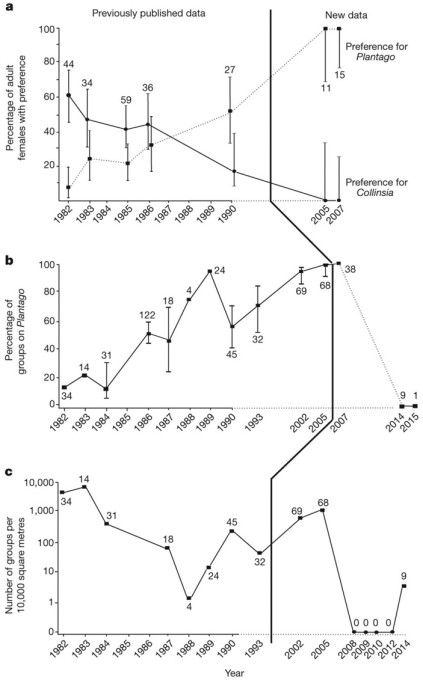
Рис. 2. Переход бабочек на новое кормовое растение и последующее вымирание популяции. По горизонтальной оси — годы. a — процент самок, предпочитающих подорожник (пунктир) и коллинсию (сплошная линия). Эти линии не являются зеркальными отражениями друг друга, потому что были еще и неразборчивые самки, откладывавшие яйца на оба растения. b — процент групп гусениц, встреченных на подорожнике. c — общее число групп гусениц на 10 000 кв м. Рисунок из обсуждаемой статьи в Nature
В 2008 году, несмотря на тщательные поиски, на лугу не удалось найти ни одной гусеницы, ни одной кладки и ни одной взрослой бабочки. Такой же результат дали осмотры в 2009, 2010 и 2012 годах. Популяция лужайки Шнайдера вымерла.
Конец у истории, впрочем, оказался не столь трагичным. Пышное разнотравье быстро истощило удобренную коровами почву и сошло на нет уже в 2008 году. Подорожник опять оказался на солнышке. Теперь бабочки, предпочитающие подорожник, снова могли бы здесь жить. Но им неоткуда было взяться.
В 2014 году авторы снова осматривали лужайку, чтобы еще раз подтвердить факт вымирания. Каково же было их удивление, когда они обнаружили целых 9 скоплений гусениц и характерные паутинки: бабочки E. editha вернулись!
Ученые подчеркивают, что совершенно этого не ожидали, потому что, как уже говорилось, ближайшая соседняя популяция находится почти в 40 км, а миграционные способности E. editha оставляют желать лучшего. Но, видимо, исследователи их все-таки недооценили. То, что это были не чудом уцелевшие и пять лет скрывавшиеся местные бабочки, а пришлые, следовало из того, что все гусеницы и паутинки были найдены на коллинсии, а не на подорожнике. Местная же популяция, как мы помним, за несколько лет до вымирания полностью перешла на подорожник.
Авторы предполагают, что повторное заселение лужайки Шнайдера было связано с резким ростом численности соседней популяции E. editha (той самой, что почти в 40 км). Этот рост, в свою очередь, был вызван пожаром 2012 года, после которого на выгоревшей земле коллинсия стала лучше расти и не так быстро вянуть.
Теперь, по-видимому, всё готово к тому, чтобы эволюционная драма на лужайке Шнайдера повторилась. Бабочки могут снова перейти на подорожник, а затем снова вымереть, если с подорожником что-то пойдет не так.
Исследование продемонстрировало новый, ранее не описанный вариант «эволюционной ловушки». Оказалось, что причиной вымирания диких животных может стать не только слишком медленный, но и чересчур энергичный эволюционный ответ на антропогенные изменения среды. Впрочем, авторы признают, что в случае с бабочками E. editha данный механизм, по-видимому, опасен для локальных популяций, но не вида в целом. Популяции живут в разных условиях, и, если одна из них попадет в «эволюционную ловушку» и вымрет, ее место со временем займут мигранты из уцелевших популяций.
Источник elementy.ru
Взлёт и падение популяции малого черноголового дубоноса Eophona migratoria в Уссурийском крае на протяжении XX столетия
А.А.Назаренко, О.П.Вальчук, С.Г.Сурмач
Практически исчез малый черноголовый дубонос Eophona migratoria. Как и должно было быть, падение численности этого «характерного вида культурного ландшафта Приморья» (Воробьёв 1954; Горчаковская 1954) долгое время оставалось незамеченным. И только тогда, когда пролёт, особенно осенний, на протяжении последних нескольких лет перестал фиксироваться во Владивостоке и на стационаре по кольцеванию птиц под Находкой, стало очевидным, что с этим видом происходит что-то неладное. Наши опасения неожиданно нашли подтверждения из районов зимовок в юго-восточном Китае: «Этот ранее широко распространённый зимующий вид значительно сократил свою численность в последние годы» (Carey et al. 1999, p. 87).
Анализируя информацию о малом черноголовом дубоносе, мы столкнулись с первым парадоксом: как оказалось, этот совсем недавно обычнейший вид юга российского Дальнего Востока не всегда был таковым. Более того, ни Р.Маак (1861) в 1859 году, ни Н.М.Пржевальский (1870) в 1867-1869 годах вообще не обнаружили его в районах их работ в долине Уссури и на западном, лесостепном, побережье озера Ханка. А малый черноголовый дубонос – очень заметная птица и во время гнездования, и на пролёте, и пропустить его, если он есть, просто невозможно. Всё это сделало необходимым тщательным образом проследить хронологию встреч с этим видом на протяжении последних 130 лет с тем, чтобы оценить долговременную динамику его популяции и темпы расселения.
 Малый черноголовый дубонос был введён в состав фауны России по экземплярам, добытым Б.Дыбовским и В.Годлевским в 1875 г., и по сборам профессиональных коллекторов братьев Ф. и Г.Дёрриес (Dörries) в 1877-1884 гг. на острове Аскольд, под Владивостоком и в бухте Сидими. По оценке В.Тачановского (Taczanowski 1891-1893, p. 625), в тот период это был редкий вид крайнего юга Уссурийского края.
Малый черноголовый дубонос был введён в состав фауны России по экземплярам, добытым Б.Дыбовским и В.Годлевским в 1875 г., и по сборам профессиональных коллекторов братьев Ф. и Г.Дёрриес (Dörries) в 1877-1884 гг. на острове Аскольд, под Владивостоком и в бухте Сидими. По оценке В.Тачановского (Taczanowski 1891-1893, p. 625), в тот период это был редкий вид крайнего юга Уссурийского края.
* Назаренко А.А., Вальчук О.П., Сурмач С.Г. 2001. Взлёт и падение популяции малого
черноголового дубоноса (Eophona migratoria) в Уссурийском крае на протяжении XX столетия: обращение к коллегам // Инвентаризация, мониторинг и охрана ключевых орнитологических территорий России 3: 134-140.
Впервые было указано и местообитание малого черноголового дубоноса: дубовые редколесья на пологих склонах холмов. Экземпляр от 24 мая 1884 г. из Сидими послужил типом подвида migratoria (Eophona melanura migratoria Hartert, 1903). Ни у села Казакевичево близ устья Уссури в 1874 г., ни в устье Бикина, по работам братьев Дёрриес в 1882 и 1887 гг., этот вид не был отмечен, даже на пролёте.
В 1908-1910 годах на западном, лесостепном, побережье Ханки были осуществлены сборы птиц для музея Общества изучения Амурского края (Черский 1915а). Собирался обширный серийный материал. Тем не менее, было добыто всего 3 экз. малого черноголового дубоноса: самец от 24 мая 1908 и самец и самка от 20 и 23 мая 1909. Этот же автор (Черский 1915б) в течение весны, лета и осени 1911 г. провёл обширные сборы и наблюдения над птицами в долине р. Одарка – местность в 22 км к востоку от современного г. Спасск-Дальний. Не было встречено ни одного малого черноголового дубоноса, даже на пролёте.
Наличие в сборах 17 индикаторных видов, в том числе козодоя Caprimulgus indicus, иволги Oriolus chinensis (полного экологического аналога E. migratoria), голубой сороки Cyanopica cyanus, сибирского жулана Lanius cristatus и других, однозначно указывало на то, что в этой местности уже существовала необходимая для обитания малого черноголового дубоноса экологическая обстановка. Совершенно очевидно, что в тот период малый черноголовый дубонос был крайне малочислен, если вообще обитал, в районе Спасска – западное побережье озера Ханка.
Не был E. migratoria найден в 1909 году на восточном побережье Приморья в низовьях р.Тетюхе (ныне р. Рудная), хотя наличие индикаторных видов (в частности, короткокрылой камышевки Horeites diphone) в сборах из этого места (Черский 1915а) свидетельствовало о
наличии потенциально пригодной экологической среды. К середине 1920-х годов ситуация уже заметно изменилась. В июне 1926 г. Г.Х.Иогансен (1927) нашёл малого черноголового дубоноса в качестве уже обычного вида в той же местности, где в 1911 г. работал А.И.Черский (1915б). Л.М.Шульпин, судя по его обширным сборам1926-1928 годов, обнаружил этот вид на гнездовании только в районе Краскино–Посьет (крайний юго-запад Приморья): 8 экз. в Зоологическом институте РАН от 7-11 июня 1927. У озера Ханка птицы были встречены, видимо, только как пролётные: экземпляры от 21 мая 1927 и 19 и 20 мая 1928 с южного побережья озера, где местность была явно непригодной для обитания вида. Экскурсия 21-25 июля 1926 по западному побережью Ханки и кратковременная поездка 3-6 июня 1928 в район современного пос. Пограничный на крайнем западе Приморья (Шульпин 1936), оказались безрезультатными: сборов E. migratoria из этих мест нет. Наверняка малый черноголовый дубонос здесь уже обитал, но был крайне малочислен. Очевидно, оставался он малочисленным и в бассейне реки Сучан (ныне Партизанская), откуда в сборах Л.М.Шульпина имеется лишь 1 экз. – самец от 6 июня 1926, хотя это был первый год его работы в Приморье, и коллектировал он в этом районе длительное время.
Остаётся отметить, что Л.М.Шульпин не нашёл малого черноголового дубоноса на восточном побережье Приморья ни в июле 1927 г. в низовьях Маргаритовки (тогда р. Пхусун), ни в июне 1928 г. в низовьях Рудной (Тетюхе). Равным образом Г.Х.Иогансен (1927) не встретил E. migratoria в июле 1926 г. у сёл Яковлевка и Чугуевка, это внутренние районы Южного Приморья.
Существование слабого пролёта у озера Ханка свидетельствовало о том, что птицы должны были гнездиться где-то севернее. Это стало очевидным в 1938-1939 годах, когда Е.П.Спангенберг (1965) обнаружил E. migratoria в качестве малочисленного вида в низовьях Имана (ныне Большая Уссурка). Исследования Л.О.Белопольского (1950) и К.А.Воробьёва (1954) позволили несколько уточнить динамику расселения E. migratoria в юго-восточном и восточном Приморье. В 1943-1945 гг. в Судзухинском (ныне Лазовском) заповеднике вид отмечался только на пролёте (Белопольский 1950). Летом 1946 г. его здесь не обнаружил и К.А.Воробьёв.
В 1960-1963 годах малый черноголовый дубонос стал уже обычной гнездящейся птицей заповедника (Литвиненко, Шибаев 1971). Летом 1948 г. К.А.Воробьёв (1954) не нашёл малого черноголового дубоноса в Сихотэ-Алинском заповеднике. По крайней мере, с 1950 года E.migratoria уже обитал у Арсеньева, в 30 км к югу от с. Яковлевка (см. выше). В начале июня 1958 г. была оценена его численность: 4.9 пар/кв.км в приречных галерейных лесах (данные А.А.Назаренко).
Вероятно, численность популяции большого черноголового дубоноса и её территориальная экспансия в Уссурийском крае достигли своего апогея в середине-конце 1960-х. На восточном побережье птицы достигли широты Сихотэ-Алинского заповедника (Елсуков 1982). Именно в эти годы было установлено обитание E. migratoria в долине Уссури к северу от р. Иман: в низовьях Бикина, в междуречье Бикина и Хора, в Большехехцирском заповеднике под Хабаровском и даже к северо-востоку от него – по долине Амура (Neufeldt, Vietinghoff-Scheel 1982).
В смежном районе, на юге Еврейской АО, в 1964 г. вид был найден в качестве вполне обычного (Смогоржевский 1965). По данным Г.С.Кисленко (1969), плотность популяции у села Венюково (долина Уссури, «междуречье» Бикина и Хора) в 1964 г. составляла 20.3 пары/кв.км в широколиственно-дубовых перелесках на склонах и 4.9 пары/км2 вприречных галерейных лесах. На крайнем западе Приморья, в долине среднего течения Комиссаровки (ранее р. Синтухе) в 1964 году мы нашли этот вид вполне обычным. Расширилась и экологическая ниша популяции: дубоносы, помимо редколесий на склонах и долинных ленточных лесов [второе было впервые отмечено в 1926 г. Иогансеном (1927)], освоили культурный ландшафт (Воробьёв 1954); более того, они стали гнездиться в черте населённых пунктов (Кисленко 1969; Назаренко 1971; Панов 1973).
В те годы это была обычная птица не только лесистых пригородов Владивостока (В.А.Нечаев, устн. сообщ.), но и городских парков (Назаров, Кызыханова 1986). В мае, в период миграций, на островах залива Петра Великого наблюдались пролётные стаи до 150 птиц (Лабзюк и др. 1971). С середины августа выводки и небольшие общества малых черноголовых дубоносов повсеместно появлялись на огородах, где кормились поспевающими семенами подсолнечника. Это было очень характерно (Литвиненко, Шибаев 1971; наши наблюдения).
Исчезновение малого черноголового дубоноса произошло совершенно незаметно. Где-то с начала 1980-х годов птицы перестали встречаться летом у г. Арсеньева, после 1986 г. – в долине Комиссаровки на крайнем западе Приморья (наши данные). В конце мая-начале июня 1990 мы не встретили ни одной птицы в типичных условиях у пос. Бойцово – несколько южнее мест, где в 1964 г. работал и проводил учёты птиц Г.С.Кисленко (1969). Ещё южнее, к западу от Лучегорска, специальные поиски E. migratoria 16-20 июня 1997 оказались тщетными, хотя были обследованы типичные места: дубовые редколесья на склонах (в т.ч. у обширной просеки вдоль пограничной«системы») и приречные леса по Уссури. Поиски E. migratoria 16-18 июня 2001 в окрестностях с. Венюково – «классическом» месте работы Г.С.Кисленко в 1963-1964 гг. (см. выше), не принесли результатов. В 1991-1996 годах эти дубоносы ни разу не отмечались у пос. Малышево, к северо-востоку от Хабаровска, хотя ранее здесь изредка встречались.
Отсутствие вида обнаружилось и на кратковременных экскурсиях у пос. Бабстово (юг Еврейской АО), точно там, где в 1964 году работал Л.А.Смогоржевский (1965). Наконец, в 1993-2000 годах малые черноголовые дубоносы ни разу не были встречены у Биробиджана (запад Еврейской АО), где мы имели многолетний стационар. Ещё далее к западу, в среднеамурской части ареала E. migratoria (Neufeldt, Vietinghoff-Scheel 1982), в 1993 г. птицы не наблюдались по нижней Бурее близ пос. Ново-Бурейский, а в 1994 г. – у с. Иннокентьевка на берегу Амура.
Депрессию популяции малого черноголового дубоноса мы связываем с событием, подробно освещённом в многочисленных публикациях: с мощным ростом популяции врановых птиц, имевшем место в бассейне Амура и в Уссурийском крае на протяжении последних 30 лет. Особенно энергично экспансирует сорока Pica pica, заселившая ныне практически все освоенные человеком районы. Отсутствует она пока только на восточном побережье Приморья к северу от пос. Пластун (Волошина и др. 1999). Факты разорения гнёзд E.migratoria сорокой известны (Тарасов, Глущенко 1995). Численность сороки в настоящее время такова, что, например, во Владивостоке её гнёзда располагаются в 100-150 м друг от друга. К несчастью, среда обитания малого черноголового дубоноса и крупных врановых птиц практически идентична.
Трудно строго обрисовать современное кружево ареала E. migratoria. Скорее всего, оно раздроблено на изолированные локальные поселения. Так, ныне это редкий вид самых низовьев Бикина (Михайлов и др. 1998). Изредка встречается в низовьях Большой Уссурки и её притока Малиновки (данные С.Г.Сурмача). Редкий вид прибрежной полосы у Сихотэ-Алинского заповедника (Волошина и др. 1999). Малочислен у Находки (данные В.Б.Манаева). В июне 1999 г. две пары дубоносов встречены в конце мая-начале июня 2001 на окраине Борисовского плато, крайний юго-запад Приморья (А.Б.Курдюков устн. сообщение). Специальные поиски малого черноголового дубоноса в июне 1999-2001 гг. в ряде мест на крайнем западе Приморья имели следующий результат: 17 июня 1999 был встречен самец к западу от пос. Пограничный (данные А.А.Назаренко).
 Замечательно, что почти все эти встречи приходятся на дубовые редколесья за пределами речных долин и на порядочном удалении от населённых пунктов. Очевидно, малый черноголовый дубонос вновь вернулся к своей «исконной» экологической среде. Впрочем, в этом нет ничего загадочного: просто в подобных местах численность врановых и в частности сороки остаётся пока, к счастью, невысокой. Долговременный тренд популяции E. migratoria и динамика его расселения показаны на рисунке. Мы не берёмся предсказывать судьбу малого черноголового дубоноса на российском Дальнем Востоке. В Уссурийском крае обитает лишь её часть. Наша задача – привлечь внимание к этому виду всех, кто работает в бассейне Амура. Наши негативные результаты на западе Еврейской АО и в низовьях Буреи и данные В.И.Поторочи (2000), как кажется, дают основания для беспокойства и в отношении среднеамурской популяции.
Замечательно, что почти все эти встречи приходятся на дубовые редколесья за пределами речных долин и на порядочном удалении от населённых пунктов. Очевидно, малый черноголовый дубонос вновь вернулся к своей «исконной» экологической среде. Впрочем, в этом нет ничего загадочного: просто в подобных местах численность врановых и в частности сороки остаётся пока, к счастью, невысокой. Долговременный тренд популяции E. migratoria и динамика его расселения показаны на рисунке. Мы не берёмся предсказывать судьбу малого черноголового дубоноса на российском Дальнем Востоке. В Уссурийском крае обитает лишь её часть. Наша задача – привлечь внимание к этому виду всех, кто работает в бассейне Амура. Наши негативные результаты на западе Еврейской АО и в низовьях Буреи и данные В.И.Поторочи (2000), как кажется, дают основания для беспокойства и в отношении среднеамурской популяции.
Малый черноголовый дубонос – бесспорный кандидат для включения в Красную книгу России. Пока же он внесён в подготовленную Красную книгу Приморского края. Не думаем, что экспансия малого черноголового дубоноса в начале-середине XX века носила спонтанный характер. Скорее всего, она была спровоцирована изменениями природных и «ложноприродных» ландшафтов, связанными с освоением Россией бассейна Амура и Уссурийского края. Но это был лишь финальный этап истории, начавшейся много ранее. Малого черноголового дубоноса, как и целый ряд других видов (в т.ч. и почти всех врановых), мы относим к фаунистической категории, чья новейшая история и динамика ареалов были предопределены возникновением и экспансией земледельческой цивилизации на востоке Азии (Назаренко 1999). Разнообразные вторичные леса, редколесья и древесно-кустарниковые заросли (антропогенная «саванна»), луга и пастбища, суходольные и рисовые поля, наконец, сам культурный ландшафт – это те антропогенно обусловленные экологические (и географические) среды, что стали возникать ещё в раннем-среднем голоцене на месте бескрайних первозданных лесов. Эти среды дали шанс для их освоения громадному числу видов с соответствующим мощным перераспределением их ареалов на обширной террито-рии, охватывающей субтропики и тропики Южной Азии, умеренные
районы бассейна Амура и районы Внутренней Азии и восточную периферию материка (Назаренко 1999).
Виды этой категории – это наиболее динамичная компонента современных региональных фаун. Пример малого черноголового дубоноса не является уникальным, целый ряд видов появился либо широко расселился по Уссурийскому краю именно в течение XX столетия. В их числе индийская кукушка Cuculus micropterus, райская мухоловка Terpsiphone paradise, китайская белая трясогузка Motacilla alba leucopsis, серый скворец Sturnus cineraceus, сорока, грач Corvus frugilegus, белая лазоревка Parus cyanus, толстоклювая Phragmaticola aedon, дроздовидная Acrocephalus arundinaceus и пестроголовая Acrocephalus bistrigiceps камышевки; из аборигенных видов – обыкновенная белоглазка Zosterops erythropleura. А популяция ястребиного Сарыча Butastur indicus претерпела точно такой же «зигзаг удачи», что и популяция малого черноголового дубоноса. Не всем им равно повезло, но большинство – процветает.
Данный феномен требует специального анализа и непредвзятой оценки в контексте современной актуальной коллизии: хозяйственная деятельность человека – региональное биоразнообразие. Наша позиция состоит в том, что любое биологическое разнообразие самоценно, вне зависимости от того, создал ли его Господь Бог или же грешный человек, тем более, что принципиально невозможно узнать, на чём остановился Первый, а где его эстафету подхватил второй (Назаренко 1990, с. 97-98).
Авторы признательны В.М.Лоскоту и С.С.Москвитину за их содействие в получении ценных данных их коллекционных сборов давних лет и старой литературы. Мы благодарны В.Н.Чернобаевой за техническое оформление рукописи и С.К.Холину за подготовку и редактирование графического файла рисунка.
Литература
1. Белопольский Л.О. 1950. Птицы Судзухинского заповедника (воробьиные и ракшеобразные) // Памяти академика П.П.Сушкина. М.; Л.: 360-406.
2. Волошина И.В., Елсуков С.В., Вдовин А.Н. 1999. Кадастр позвоночных животных Сихотэ-Алинского заповедника и Северного Приморья. Владивосток: 1-91.
3. Воробьёв К.А. 1954. Птицы Уссурийского края. М.: 1-360.
4. Горчаковская Н.Н. 1954. Род восточноазиатские дубоносы Eophona Gould, 1851 // Птицы Советского Союза. М., 5: 171-177.
5. Елсуков С.В. 1982. Птицы // Растительный и животный мир Сихотэ-Алинскогозаповедника. М.: 195-217.
6. Иогансен Г.Х. 1927. Материалы по орнитофауне Южно-Уссурийского края // Uragus 4, 3: 19-29.
7. Кисленко Г.С. 1969. Птицы некоторых ландшафтов нижнего течения Уссури //Учён. зап. Моск. обл. пед. ин-та им. Н.К.Крупской 224: 49-74.
8. Лабзюк В.И., Назаров Ю.Н., Нечаев В.А. 1971. Птицы островов северо-западной части залива Петра Великого // Орнитологические исследования на юге Дальнего Востока. Владивосток: 52-78.
9. Литвиненко Н.М., Шибаев Ю.В. 1971. К орнитофауне Судзухинского заповедника и долины реки Судзухе // Экология и фауна птиц юга Дальнего Востока. Владивосток: 127-186.
10. Михайлов К.Е., Шибнев Ю.Б., Коблик Е.А. 1998. Гнездящиеся птицы бассейна Бикина (Аннотированный список видов) // Рус. орнитол. журн. 7 (46): 3-19.
11. Назаренко А.А. 1971. Краткий обзор птиц заповедника «Кедровая Падь» // Орнитологические исследования на юге Дальнего Востока. Владивосток: 12-51.
12. Назаренко А.А. 1990. Орнитофаунистический обмен между Южной и Северной Азией на восточной периферии континента: последний ледниково-межледниковый цикл // Журн. общ. биол. 51, 1: 89-106.
13. Назаренко А.А. 1999. Хозяйственная деятельность как фактор роста биоразнообразия фаун и сообществ птиц на восточной окраине Азии: приглашение к дискуссии // Вестн. ДВО РАН 1: 22-30.
14. Назаренко А.А. 2000. Ястребиный сарыч Butastur indicus (Gm., 1788) // Красная книга Российской Федерации: Животные. М.: 430-431.
15. Назаров Ю.Н., Казыханова М.Г. 1986. Летняя авифауна Владивостока // Изучение птиц СССР, их охрана и рациональное использование. Л., 2: 99-100 [2-е
изд. Назаров Ю.Н., Казыханова М.Г. 2006. Летняя авифауна Владивостока //Рус. орнитол. журн. 15 (316): 390-391].
16. Панов Е.Н. 1973. Птицы Южного Приморья. Новосибирск: 1-376.
17. Потороча В.Н. 2000. О весеннем пролёте птиц на северо-востоке Амурской области// Современные проблемы орнитологии Сибири и Центральной Азии. Улан-Удэ: 134-137.
18. Пржевальский Н.М. 1870. Путешествие в Уссурийском крае в 1867-1869 гг. СПб.: 1-297.
19. Смогоржевский Л.А. 1965. Птицы южной части Еврейской АО // Новости орнитологии. Алма-Ата: 354-355.
20. Спангенберг Е.П. 1965. Птицы бассейна реки Иман // Сб. тр. Зоол. музея Моск. ун-та 9: 98-202.
21. Тарасов А.А., Глущенко Ю.Н. 1995. Врановые Приханкайской низменности //Проблемы сохранения вводно-болотных угодий международного значения: озеро Ханка. Спасск-Дальний: 57-68.
22. Черский А.И. 1915а. Орнитологическая коллекция музея Общества изучения Амурского края во Владивостоке // Зап. Общ-ва изуч. Амур. края 14: 143-276.
23. Черский А.И. 1915б. Орнитологические сборы с 8 марта по 20 октября 1911 года вдолине верхнего течения речки Одарки, близ д. Ново-Владимировки Иманского уезда Приморской области // Зап. Общ-ва изуч. Амур. края 14: 79-141.
24. Шульпин Л.М. 1936. Промысловые, охотничьи и хищные птицы Приморья. Владивосток: 1-436.
25. Carey G.J., Kennerley P.R., Cheung H.F. et al. 1999. Systematic list // Hong Kong Bird report 1997: 15-92.
26. Neufeldt I.A., Vietinghoff-Scheel E. 1982. Eophona migratoria // Atlas der Verbreitung palaearktischer Vögel. 10 Lief. Berlin.
Рус. орнитол. журн. 2006. Том 15. Экспресс-выпуск № 316
Источник cyberleninka.ru
Рекомендуем прочесть
 Пазухи свода собора святого Марка и парадигма Панглосса: критика адаптационистской программы
Пазухи свода собора святого Марка и парадигма Панглосса: критика адаптационистской программы Групповые адаптации без группового отбора?
Групповые адаптации без группового отбора? Птицы-печники против «общих мест» эволюционной экологии
Птицы-печники против «общих мест» эволюционной экологии Паразитические манипуляции фенотипом хозяина: эффекты во внутренней и внешней среде
Паразитические манипуляции фенотипом хозяина: эффекты во внутренней и внешней среде Так кто всё-таки сэкономил на мозгах?
Так кто всё-таки сэкономил на мозгах? Птицы и муравьи не мешают друг другу есть гусениц
Птицы и муравьи не мешают друг другу есть гусениц Страсти по сочинскому самшиту: как защитить реликтовый лес
Страсти по сочинскому самшиту: как защитить реликтовый лес Перестройка в экологии: от описания видимого к пониманию скрытого
Перестройка в экологии: от описания видимого к пониманию скрытого
