
М.А. Еремина, Н.Е. Грунтенко
Нейроэндокринная стресс-реакция является эффективным способом защиты организмов от неблагоприятных воздействий различной природы. Эта реакция универсальна и возникает в ответ на стимулы, непривычные для жизни и обитания не только данного вида организмов, но и отдельной популяции. В обзоре рассматривается последовательная смена теоретических концепций, подходов и методов исследования в этой научной области: от становления концепции стресса Г. Селье до наших дней.
В 1982 г. Г. Селье определил стресс как совокупность стереотипных филогенетических запрограммированных реакций организма, которые вызываются любыми сильными, сверхсильными, экстремальными воздействиями и сопровождаются перестройкой адаптивных сил организма. Агент, вызывающий стресс, был назван стрессором.
В динамике комплекса неспецифических защитно-приспособительных реакций в ответ на стрессовое воздействие, направленных на создание устойчивости (резистентности) организма к любому фактору, закономерно прослеживаются три стадии («триада Селье»):
1) реакция тревоги,
2) стадия резистентности,
3) стадия истощения.
Длительность и выраженность каждой из них могут варьировать в зависимости от природы и силы стрессорного агента, вида животного и физиологического состояния организма. Отсутствие гипоталамо-гипофизарно-адренокортикальной системы у насекомых считалось доказательством невозможности развития у них стресс-реакции по типу теплокровных животных. Тем не менее с начала 1980-х годов получено достаточно доказательств развития реакции стресса у насекомых, что подчеркивает консервативность стресс-реакции у млекопитающих и насекомых.
Cходство в нейрохимических и физиологических изменениях у беспозвоночных и позвоночных в ответ на стрессорное воздействие свидетельствует о том, что реакция на стрессор – это совокупность древних, сохраненных в эволюции механизмов. Насекомые предоставляют уникальные возможности для экспериментов, которые могут помочь понять основные механизмы стрессовых реакций. У личинок насекомых механизм стресс-реакции изучен детально. В этом веке основные усилия исследователей направлены на изучение механизмов стресс-реакции у имаго насекомых и генетического контроля отдельных ее звеньев. Изучение стресс-реакции у насекомых важно и в теоретическом плане – оно демонстрирует конвергентность путей эволюции адаптивных преобразований у таких далеко отстоящих друг от друга таксонов, как насекомые и млекопитающие, и в практическом – закономерности механизма этой реакции можно использовать при моделировании наследственных или приобретенных заболеваний человека, разработке методов разведения хозяйственно ценных насекомых и способов борьбы с насекомыми-вредителями.
***
Термин «стресс» переводится с английского языка как «напряжение». Физиологические аспекты стресса впервые были освещены У. Кенноном (Cannon, 1932) в классических работах по универсальной реакции «бороться или бежать». Кроме того, он ввел в физиологию понятие «гомеостаз», считая, что постоянство внутренней среды организма поддерживается благодаря симпатической нервной системе и гормонам. Чем сложнее организм, тем тоньше, многочисленнее и разнообразнее система противовесов в нем. Она обеспечивается формированием как постоянных, так и временных связей, устанавливающих соотношения между мельчайшими элементами внешнего мира и тончайшими реакциями организма (Cannon, 1932).
Множество операций, проведенных в школе У. Кэннона над животными, доказало, что внешне наблюдаемые изменения в поведении, которые можно назвать эмоциональными, порождаются глубинными сдвигами в нейрогуморальных процессах. Эти сдвиги готовят организм к критическим ситуациям, требующим повышенной траты энергии, снятия усталости, предотвращения кровопотери и т. п. В одном из докладов о своих открытиях Кеннон сообщил, что благодаря выбрасываемому в кровь при сильных эмоциях адреналину в числе других его «мобилизационных» эффектов происходит увеличение в крови уровня сахара, поступающего к мышцам (Cannon, 1932).
А.А. Виру (1981) указывает на то, что в отдельных работах отечественных ученых (А.Д. Сперанского, Д.Н. Насонова, М.Я. Александрова, А. Цанка) четко прослеживаются верные направления в раскрытии механизмов неспецифической адаптации. Так, еще в 1934 г. Д.Н. Насонов в своих исследованиях показал, что содержимое живой клетки (протоплазма) поразительно однотипно реагирует на любые изменения окружающей среды. Этот стереотип ответов клетки был назван Насоновым «паранекрозом». Так фактически был открыт стресс, но не на организменном, а на клеточном уровне (Апчел, Цыган, 1999).
Классическое учение о стрессе и адаптационном синдроме создано Г. Селье (1979). Он писал, что впервые «наткнулся» на идею стресса и общего адаптационного синдрома в 1925 г. 4 июля 1936 г. в журнале «Nature» было опубликовано его письмо в редакцию «Синдром, вызываемый различными повреждающими агентами», в котором приводились данные о стандартных реакциях организма на действие различных болезнетворных агентов (Апчел, Цыган, 1999). Это сообщение положило начало учению о стрессе, хотя в отдельных, более ранних, научных трудах В. Вундта (1980) и В. Джемса (1905) было упоминание о проблеме стресса с психологической точки зрения.
Г. Селье длительное время избегал употребления термина «стресс», поскольку тот широко использовался для обозначения нервно-психического напряжения. Только в 1946 г. Селье начал систематически использовать термин «стресс» для общего адаптационного напряжения. Он сформулировал концепцию стресса, при этом рассматривал физиологический стресс как ответ на любые предъявленные к организму требования и считал, что с какой бы трудностью не столкнулся организм, с ней можно справиться двумя типами реакций: активной, или борьбы, и пассивной, в виде бегства от трудностей или готовности терпеть их. Полная свобода от стресса, как считал Г. Селье, означала бы смерть (Китаев-Смык, 1983). В 1982 г. Селье определил стресс как совокупность стереотипных филогенетических запрограммированных реакций организма, которые вызываются любыми сильными, сверхсильными, экстремальными воздействиями и сопровождаются перестройкой адаптивных сил организма (Селье, 1987).
В общебиологическом плане, по мнению Ф.З. Меерсона (1981), стресс-реакция сформировалась в процессе эволюции как необходимое неспецифическое звено более сложного целостного механизма адаптации. Стрессор – всякое достаточное сильное (не обязательно экстремальное) воздействие: тепло, холод, эмоциональное воздействие, болевые раздражения, голодание, интоксикация и т. д.
В динамике комплекса неспецифических защитно-приспособительных реакций в ответ на стрессовое воздействие, направленных на создание устойчивости (резистентности) организма к любому фактору, закономерно прослеживаются три стадии («триада Селье»): 1) реакция тревоги, 2) стадия резистентности, 3) стадия истощения. Длительность и выраженность каждой стадии могут варьировать в зависимости от природы и силы стрессорного агента, вида животного и физиологического состояния организма (Селье, 1979).
Первая стадия синдрома (реакция тревоги) характеризует остро протекающую, активную мобилизацию адаптационных процессов в организме в ответ на всякое смещение гомеостаза при стрессе (на первичный шок). В это время устойчивость организма к воздействиям быстро возрастает. Во второй стадии (резистентности) устанавливается повышенная сопротивляемость к стрессору, которая носит общий, неспецифический характер. Например, если стресс вызывается холодом, то на стадии резистентности выявляется повышенная устойчивость не только к холоду, но и к действию повышенной температуры, рентгеновских лучей, токсинов и т. д. В случаях, когда стресс слишком сильный или длительный, защитно-приспособительные механизмы организма могут истощаться и общий адаптационный синдром переходит в третью стадию (стадия истощения), характеризующуюся снижением резистентности организма к данному стрессору и другим видам стрессорных воздействий. Эта стадия называется также вторичным шоком (Селье, 1979).
По признанию Г. Селье, он заимствовал очень многое у И.П. Павлова. То, что Павлов трактовал с точки зрения нервной системы, он перевел на язык и термины гуморальной системы. Г. Селье не считал стресс вредным, а рассматривал его как реакцию, помогающую организму выжить. Он назвал отрицательный стресс дистрессом, а положительный стресс – эустрессом (Селье, 1979).
В своем учении о стрессе и адаптационном синдроме Селье основное внимание обращал на роль гормональных изменений, не анализируя участие нервной системы в механизме формирования стресса. Последнее подверглось критике в отечественной литературе. Причем неоднократно подчеркивалось, что стресс-реакция развивается как цепь событий, опосредованных гипоталамо-гипофизарноадренокортикальной системой (Косицкий, Смирнов, 1970; Горизонтов, 1973; Погодаев, 1976).
Нейроэндокринная стресс-реакция насекомых
Содержание
Отсутствие гипоталамо-гипофизарно-адренокортикальной системы у насекомых считалось доказательством невозможности развития у них стресс-реакции по типу теплокровных животных (Косицкий, Смирнов, 1970). Тем не менее с начала 1980-х годов получено доcтаточно доказательств развития реакции стресса у насекомых (Rauschenbach et al., 1980, 1983a, b, 1987, 2014; Cymborowski et al., 1982; Chernysh, 1991; Harris, Woodring, 1992; Mobius, Penzlin, 1993; Hirashima et al., 2000; Pszczolkowski, Chiang, 2000; Gruntenko et al., 2004, 2010; Neckameyer, Weinstein, 2005; Tauchman et al., 2007; Gruntenko, Rauschenbach, 2008).
Эволюцией создано две «стратегии выживания» насекомых в неблагоприятных условиях внешней среды: состояние диапаузы и состояние задержки метаморфоза у недиапаузирующих насекомых. И тот и другой тип приспособления позволяют насекомым пережить неблагоприятные условия. Гормональную основу обоих типов составляет нейроэндокринная стресс-реакция, обладающая двумя основными чертами – неспецифичностью по отношению к воздействию (возникает при действии экстремальных факторов различной природы) и адаптивностью. Но именно такие характеристики свойственны реакции, обозначенной термином «стресс» у млекопитающих (Rauschenbach et al., 1987).
В стресс-реакцию насекомых вовлечены различные гормоны, в частности биогенные амины (дофамин (ДА), октопамин (ОА), серотонин), экдистероиды, ювенильный гормон (ЮГ), адипокинетический гормон (АКГ) (Kodrík et al., 2015) и инсулин (Грунтенко, 2008; Johnson, White, 2009; Gruntenko, Rauschenbach, 2017).
Эндокринные органы синтезируют, накапливают и выделяют в гемолимфу гормоны, регулирующие разнообразные физиологические процессы. Основные эндокринные органы насекомых охватывают нейросекреторные клетки (НСК) (место синтеза биогенных аминов), ретроцеребральный комплекс, состоящий из двух пар желез, называемых кардиальными (corpora cardiaca) (место синтеза АКГ) и прилежащими телами (corpora allata, СА) (место синтеза ЮГ), перисимпатические органы и проторакальные железы (место синтеза 20-гидроксиэкдизона (20Э)) (Тыщенко, 1986).
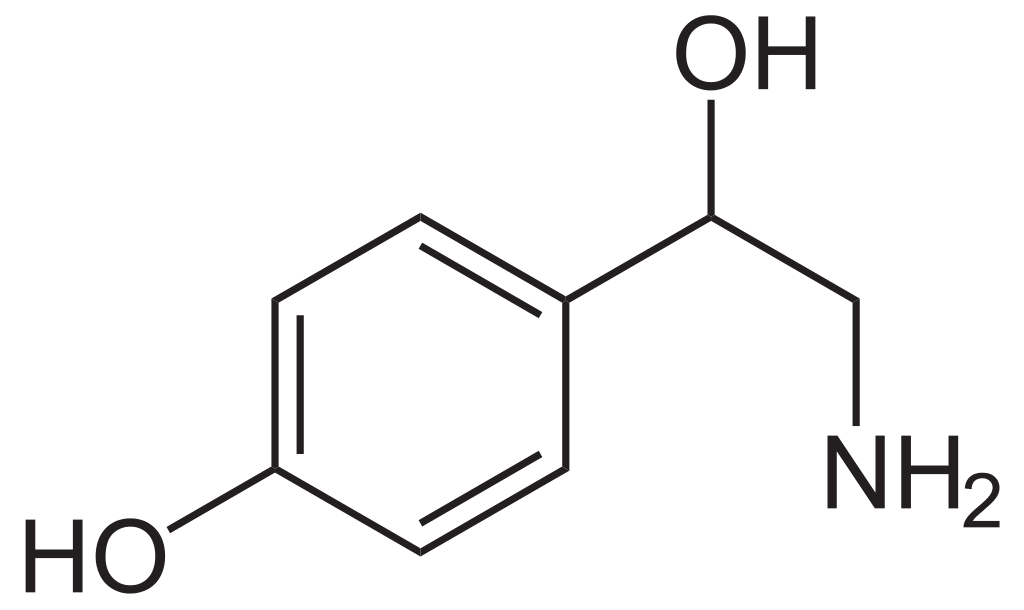
Октопамин
Биогенные амины у насекомых выполняют функции нейромедиаторов, нейромодуляторов, оказывающих локальное влияние на близлежащие клетки в центральной нервной системе, способствующее изменению эффекта нейромедиаторов и нейрогормонов, переносимых током гемолимфы и действующих на больших расстояниях (Evans, 1985; Janković-Hladni, 1991; Johnson, White, 2009).
В ходе личиночного развития насекомого ЮГ определяет характер линьки, инициированной 20Э, и препятствует преждевременному метаморфозу: при высоком титре ЮГ происходит личиночная линька, при его резком падении – метаморфоз. У имаго ЮГ и 20Э играют роль гонадотропинов: определяют половое поведение самок, контролируют развитие яичников и вителлогенез, стимулируют синтез феромонов, а также участвуют в регуляции личиночной и имагинальной диапаузы (Грунтенко, 2008). АКГ стимулирует распад резервных триглицеридов в жировом теле, освобождая диглицеридную транспортную форму жиров, поступающих из трофоцитов в гемолимфу (Тыщенко, 1986).
Еще одним организатором стрессовых реакций как у позвоночных, так и у насекомых является сигнал инсулинового пути (Holzenberger et al., 2003; Jonson, White, 2009).
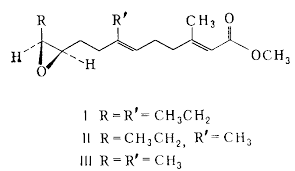
Ювенильный гормон
Проведены многочисленные исследования по изучению влияния высоких температур, низкого качества корма, инсектицидов и других стрессоров на НСК мозга и подглоточного ганглия (Ivanović et al., 1975, 1979, 1985, 1989; Janković-Hladni et al., 1983; Leković et al., 2001;
Perić-Mataruga et al., 2001). Различные стрессорные воздействия разной интенсивности вызывают определенные изменения, что, в свою очередь, влияет на синтез и секрецию нейросекреторного материала (Janković-Hladni et al., 1992). В ряде работ показано, что ответ на уровне НСК у разных видов насекомых зависит от интенсивности стресс-фактора и продолжительности воздействия (Ivanović et al., 1975; Mrdaković et al., 2003).
Сигнал стрессора, полученный экстерорецепторами насекомого, передается через сенсорные нервные пути к мозгу. Первая стадия стресс-реакции в большой степени регулируется нейрогормонами и биогенными аминами (Davenport, Evans, 1984; Грунтенко, 2008). Под их влиянием в течение нескольких минут в жировом теле мобилизуются запасные вещества. На определенной стадии развития стресс-реакции к этой группе нейрогормонов присоединяются экдистероиды. Вторая стадия развития стресс-реакции проходит в полной зависимости от экдистероидов, синтез которых, в свою очередь, регулируется экдизиотропными нейрогормонами (экдизиотропинами) и ювенильным гормоном (Грунтенко, 2008).
Таким образом, как у млекопитающих, так и у насекомых в неблагоприятных условиях внешней среды развивается неспецифическая стандартная гормональная реакция, имеющая адаптивный характер. У первых это реакция напряжения, увеличивающая энергетические возможности организма для обеспечения адаптации, у вторых – реакция угнетения гормональной системы, приводящая к торможению развития, что также обеспечивает адаптацию к неблагоприятным воздействиям. Следовательно, существо реакции, ее биологическое значение одинаково в обоих случаях (Rauschenbach et al., 1987; Раушенбах, 1990).
Исследования М. Vigas (1983) и К.V. Sudakov (1983) на млекопитающих свидетельствуют о том, что неспецифическая по механизму реакция стресса обладает определенной специфичностью по степени ее выраженности:
а) стрессогенность воздействия зависит от его силы (существует порог); б) степень реакции зависит от вида животного (один и тот же стрессор может вызывать стресс у одних организмов и быть подпороговым у других). Сходство между рассматриваемой нейроэндокринной реакцией насекомых и стрессом млекопитающих существует и вэтом плане:
а) наличие порога при воздействии одного типа показано для Rhodnius prolixus (развитие личинок при 32 °С не сказывается на превращении в имаго, а при 35 °С имагинальная линька задерживается (Wigglesworth, 1952) и для Drosophila virilis (плотность в 40 личинок не влияет на метаморфоз, а плотность в 80 личинок задерживает егонаступление (Раушенбах и др., 1983);
Кровососущий клоп Rhodnius prolixus
б) зависимость от вида насекомого следует из данных о том, что у D. virilis температура 32 °С вызывает развитие стрессорной реакции (Rauschenbach et al., 1987), а для R. prolixus она является подпороговой (Wigglesworth, 1952, 1955).
Более того, продемонстрировано, что один и тот же фактор может быть, а может и не быть стрессорным для вида, в зависимости от того, на какой стадии развития он действует. Так, например, не всегда высокая плотность неблагоприятна для развития насекомых. Исходя из биологии вида, стрессорной может оказаться и низкая плотность популяции. Это положение хорошо иллюстрируется работой (Singh, Pandey, 1980). Сравнивая развитие личинок первого и третьего возраста Diacrisia oblique в условиях низкой и высокой плотности, авторы обнаружили, что самый длительный срок развития и самая высокая смертность характерны для культур личинок первого возраста с низкой плотностью и третьего возраста – с высокой. Таким образом, для первых стрессорными являются условия развития с низкой плотностью, тогда как для последних – с высокой. Другим примером могут служить исследования (Ivanovic et al., 1985), продемонстрировавшие, что температура 8 °С, стрессорная для личинок Morimus funereus, собранных в июне (вызывает ингибирование НСК, секретирующих проторакотропный гормон, ПТТГ), не вызывает стресса у личинок, собранных в ноябре.

Кровососущий триатомовый клоп Rhodnius prolixus
В то же время температура 23 °С является стрессорной для личинок, собранных в ноябре, и не приводит к стрессу июньских личинок. Когда стрессорное воздействие существенно выше порогового, оно может вызвать у насекомых обратную реакцию – ускорение метаморфоза. Так, в работе С.И. Черныша (1983) продемонстрировано, что погружение личинок Calliphora eryphrocephala в 40 % раствор формальдегида на 180 мин ускоряло формирование пупариума, тогда как погружение их на 90 мин вызывало замедление развития.
Ускоренный метаморфоз отмечали при электрошоке личинок Bombyx mori (Черныш, 1983). Преждевременный метаморфоз наблюдается при повреждении кутикулы личинок при откладке яиц паразитирующей самкой (Lawrence, 1988). Следовательно, этот эффект имеет место только в случае многократного прокалывания кутикулы, при однократном прокалывании он отсутствует. Вместе с тем при умеренном повреждении кутикулы (пороговом) наблюдается стандартная стрессорная реакция – задержка метаморфоза, ингибирование секреции ПТТГ, задержка секреции экдизона, увеличение содержания ЮГ и снижение ЮГ-эстеразной активности (O’Kasha, 1968; McCaleb, Kumaran, 1980; Bogus et al., 1986; Rauschenbach et al., 1987). Важно подчеркнуть, что структура стрессорной реакции несколько различается у насекомых со строго фиксированным числом личиночных возрастов и подвергающихся сверхчисленным линькам (Раушенбах, 1990).
Гормональный статус насекомых со сверхчисленными линьками в неблагоприятных условиях среды последовательно изучали Б. Цимборовский и М.И. Богус (Cymborowski, Bogus, 1976; Bogus, Cymborowski, 1981). Б. Цимборовский предложил схему стрессорной реакции, возникающей у личинок Galleria mellonella в условиях холодового стресса (Cymborowski, 1988). Автор предположил, что информация о температурном статусе тела личинок G. mellonella передается в мозг через вентральную нервную хорду, вызывая задержку в переключении с личиночной на куколочную программу. Позже исследователями была обнаружена целая группа регуляторных белков, выделяемых из нервной ткани и контролирующих синтез ЮГ посредством СА, – аллатотропины и аллатостатины, стимулирующие и ингибирующие модуляторы, которые достигают СА либо посредством прямой нейросекреторной иннервации, либо через гемолимфу (Tobe, Stay, 1985; Cymborowski, 1988; Richard et al., 1990; Kramer et al., 1991; Stay et al., 1996; Bendena et al., 1999).
Таким образом, показано, что изменения в содержании гормонов при развитии насекомых в неблагоприятных условиях взаимосвязаны и что в этих условиях у них возникает неспецифическая гормональная реакция, подобная стрессу млекопитающих (Rauschenbach et al., 1987; Cymborowski, 1988).

Совка Diacrisia oblique
Современное состояние исследований нейроэндокринной стресс-реакции насекомых
Насекомые представляют уникальные возможности для экспериментов, которые могут помочь определить основные механизмы стрессовых реакций. Кроме того, прикладные аспекты контроля численности как вредных, так и полезных насекомых требуют более глубокого понимания механизмов действия стрессоров на организм (Johnson,White, 2009).
У личинок насекомых механизм стресс-реакции изучен детально. Показано, что именно повышение содержания ЮГ при стрессе вызывает состояние диапаузы или задержку метаморфоза у недиапаузирующих насекомых, позволяя им таким образом переждать неблагоприятные условия (Rauschenbach et al., 1987; Cymborowski, 1988). Кроме того, у личинок насекомых наблюдается повышение содержания биогенных аминов (Hirashima, Eto, 1993; Hirashima et al., 1994).
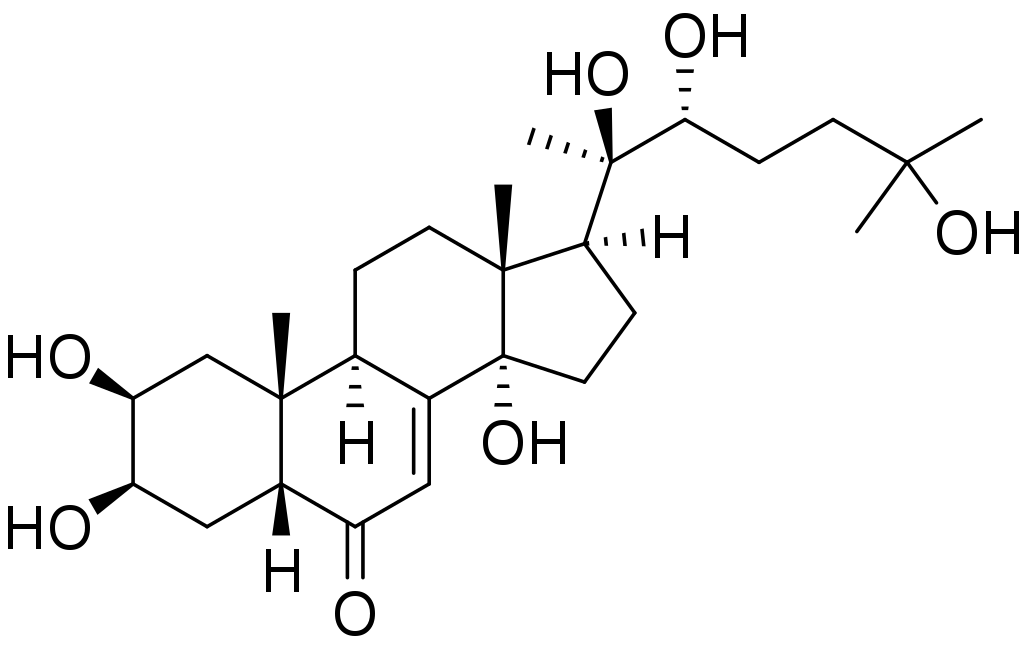
20Э
У имаго насекомых стресс-реакция направлена на попытку избежать вредного воздействия или же адаптироваться к неблагоприятным условиям, если первое невозможно (Раушенбах, 1997). В настоящее время основные усилия исследователей направлены на изучение механизмов стресс-реакции у имаго насекомых и генетического контроля отдельных ее звеньев (Ченцова и др., 2007; Грунтенко, 2008).
В работах И.Ю. Раушенбах с коллегами (Rauschenbach et al., 1987, 2014; Раушенбах, 1990, 1997; Раушенбах и др., 2000) и Н.Е. Грунтенко с коллегами (Gruntenko et al., 2000, 2003a, b, 2004, 2005a, b, 2010, 2012) исследован механизм стресс-реакции в процессе онтогенеза у D. virilis и D. melanogaster, определены основные маркерные характеристики стресс-реакции у личинки и имаго дрозофилы. Выяснено, что ни один из стресс-связанных гормонов дрозофилы не является пусковым звеном в развитии стресс-реакции. Для D. virilis показан моногенный контроль ответа на стресс всех гормональных систем (Раушенбах, 1997). У D. melanogaster выявлен комплекс генов, мутации в которых приводят к изменению интенсивности ответа на стрессор всех стресс-связанных гормонов (ДА, ОА, ЮГ и 20Э) и ферментов их метаболизма (Грунтенко, 2008).
В ряде работ (Беньковская и др., 2000, 2004, 2005, 2008; Гайфуллина и др., 2004; Салтыкова и др., 2005; Соколянская и др., 2005; Беньковская, Соколянская, 2008) обобщены результаты многолетних исследований, посвященных выяснению роли стресс-реакции в формировании онтогенетических адаптаций на примерах комнатной мухи Musca domestica, колорадского жука Leptinotarsa decemlineata и медоносной пчелы Apis mellifera. Показаны универсальный фазовый характер стресс-реакции насекомых и сложная структура стадии реакции тревоги, согласующаяся с фундаментальной схемой Г. Селье (1979) и подтверждающая общность защитной реакции в отдаленных таксонах животного мира. Установлены роль стресса и место стресс-реакции в микроэволюционных преобразованиях популяций насекомых как механизма реализации адаптивного потенциала особи и популяции.
Одновременно продемонстрирована роль стресс-реакции как преадаптивного стимула, повышающего адаптационные возможности особи и расширяющего границы адаптивного потенциала популяций насекомых.
Модель Н. Ивен с коллегами описывает общую реакцию стресса у медоносных пчел (Even et al., 2012). Эта работа обеспечивает основу для понимания того, как A. mellifera может реагировать на стрессоры, а также направлена на стимулирование исследований в данной области. Сравнение путей реагирования на стресс у позвоночных и медоносных пчел демонстрирует гомологичную структуру реакции в этих двух группах животных, включая регуляцию возбуждения и стимуляции когнитивных функций в головном мозге при помощи катехоламинов в сочетании с нейрогормональными сигналами, стимулирующими мобилизацию энергии на периферии. Тем не менее степень, до которой пути стрессоустойчивости эволюционно сохраняются, остается неясной. Исследователи также выделили аспекты стрессовой реакции, которые, по-видимому, специфичны для медоносных пчел в результате их особой социальной организации.

Огнёвка пчелиная Galleria mellonella
В работе Е.А. Чертковой (2015) впервые показано повышение уровня ДА у капустной совки Mamestra brassicae и вощинной огневки G. mellonella при воздействии стресс-факторов абиотической природы (температура, механические повреждения). Также выявлено, что к увеличению уровня ДА приводит заражение личинок G. mellonella бактериями Bacillus thuringiensis, а колорадского жука L. decemlineata и M. brassicae – энтомопатогенными грибами Metarhizium robertsii и Beauveria bassiana. Кроме того, обнаружено, что степень повышения уровня ДА зависит от вирулентных свойств энтомопатогенного микроорганизма.
Уровень ДА в гемолимфе личинок L. decemlineata повышается также при инфицировании энтомопатогенными бактериями Bacillus thuringiensis, и это повышение носит дозозависимый характер. По мнению автора, результаты этих исследований могут быть использованы для дальнейшего изучения стресс-реакции насекомых, лежащей в основе их резистентности к химическим и микробиологическим инсектицидам, и разработок методических подходов к ее преодолению (Черткова, 2015).
Множество работ посвящено изучению реакции окислительного стресса, ее механизму и способам защиты организма от него (Ahmad, 1995; Bi, Felton, 1995; Meng et al., 2009; Lalouette et al., 2011; Velki et al., 2011; Zhao, Haddad, 2011; Krishnan, Kodrík, 2012; Bednářová et al., 2013). У насекомых реакции антиокислительного стресса, по-видимому, регулируются преимущественно АКГ. Очевидное участие АКГ в контроле защиты против окислительного стресса является типичным примером эндокринной регуляции такого ответа. Механизмы действия АКГ при окислительном стрессе, понимание которых необходимо для детального выяснения роли АКГ в стресс-реакции, до сих пор мало изучены.
Тем не менее интенсивные исследования, преимущественно на модели D. melanogaster, могут принести интересные данные в ближайшем будущем. В настоящее время легко доступны мутанты D. melanogaster, созданные с помощью методов генной инженерии, перепроизводством или дефицитом гормонов стресса, подавлением экспрессии рецепторов этих гормонов. Они представляют собой мощный инструмент для изучения ответа на стресс (Kodrík et al., 2015).
Несколько нейродегенеративных заболеваний, включая болезнь Паркинсона и болезнь Альцгеймера, связаны с развитием окислительного стресса и интенсивно изучаются. Большая часть данных получена на позвоночных животных, однако ряд работ за последние несколько лет показал, что насекомые представляют собой важную модель для исследования окислительного стресса. Кроме того, результаты, полученные на насекомых, очень часто универсальны и могут способствовать выяснению важнейших механизмов окислительного стресса у животных, в том числе и у людей (Kodrík et al., 2015).
Одним из направлений исследований С.А. Адамо (Adamo, 2012, 2014, 2017a, b; Adamo et al., 2013, 2017) является изучение стресс-зависимой иммунной функции с использованием моделей насекомых. Показано, что у насекомых медиаторы стрессового ответа (т. е. гормоны стресса) меняют иммунный статус насекомого, в том числе частично перераспределяя молекулярные ресурсы в пользу двигательной активности, интенсификация которой необходима для борьбы или бегства. Например, белки переноса липидов участвуют как в стрессе, так и в иммунных реакциях, что приводит к снижению устойчивости к болезням, когда эти белки переключаются на стресс-ответ. Октопамин необходим также как для развития стресс-реакции, так и для активации иммунного ответа (фагоцитоза) (Adamo, 2014, 2017a).
Однако С.А. Адамо полагает, что при стрессе у насекомых происходит, скорее, реконфигурация иммунной системы, обеспечивающая снижение затрат на иммунный ответ, чем прямой обмен ресурсами между этими двумя защитными системами организма (Adamo, 2017a). Так, стресс-реакция индуцирует провоспалительное состояние, которое, по мнению автора, усиливает ранний иммунный ответ.
Cходство в нейрохимических и физиологических изменениях у беспозвоночных и позвоночных в ответ на стрессорное воздействие показывает, что реакция на стрессор – это совокупность древних, сохраненных в эволюции механизмов. Изучение стресс-реакции у насекомых важно как в теоретическом плане – определяется конвергентность путей эволюции адаптивных преобразований у таких далеко отстоящих друг от друга таксонов, как насекомые и млекопитающие, так и в практическом – закономерности механизма этой реакции можно использовать при моделировании наследственных или приобретенных заболеваний человека, разработке методов разведения хозяйственно ценных насекомых и способов борьбы с насекомыми-вредителями.
Список литературы
Апчел В.Я., Цыган В.Н. Стресс и стрессоустойчивость человека. CПб.: ВМА, 1999.
Беньковская Г.В., Леонтьева Т.Л., Удалов М.Б. Резистентность колорадского жука к инсектицидам на Южном Урале. Агрохимия. 2008;8:55-59.
Беньковская Г.В., Николенко А.Г., Салтыкова Е.С., Ишмуратова Н.М., Харисов Р.Я., Ишмуратов Г.Ю. Адаптогенное действие препарата биосил на медоносную пчелу и комнатную муху. Агрохимия. 2005;3:74-78.
Беньковская Г.В., Соколянская М.П. Чувствительность к тепловому стрессу имаго комнатной мухи лабораторных линий, селектированных инсектицидами и абиотическими факторами. Агрохимия. 2008;3:52-56.
Беньковская Г.В., Поскряков А.В., Николенко А.Г. Влияние метопрена на переносимость теплового шока в онтогенезе колорадского жука. Агрохимия. 2000;12:58-61.
Беньковская Г.В., Удалов М.Б., Поскряков А.В., Николенко А.Г. Феногенетический полиморфизм колорадского жука Leptinotarsa decemlineata Say и его чувствительность к инсектицидам на территории Башкирии. Агрохимия. 2004;12:23-28.
Виру А.А. Гормональные механизмы адаптации и тренировки. Л.: Наука, 1981.
Вундт В. Основы физиологической психологии. Чувства и аффекты. Л., 1980;55(3).
Гайфуллина Л.Р., Салтыкова Е.С., Беньковская Г.В., Николенко А.Г. Иммунные реакции личинок и имаго колорадского жука (Leptinotarsa decemlineata Say) при применении препарата биологической защиты картофеля. Агрохимия. 2004;9:1-7.
Горизонтов П.Д. Резистентность и поражение. Вопросы общей патологии. Патологическая физиология экстремальных состояний. М.: Медицина, 1973.
Грунтенко Н.Е. Стресс и размножение насекомых: гормональный контроль. Новосибирск; Москва: ТО научн. изданий КМК, 2008.
Джемс В. Психология. СПб., 1905.
Китаев-Смык Л.А. Психология стресса. М.: Наука, 1983.
Косицкий Г.И., Смирнов В.М. Нервная система и «стресс» (о принципе доминанты в патологии). М.: Наука, 1970.
Меерсон Ф.З. Адаптация, стресс и профилактика. М.: Наука, 1981.
Погодаев К.И. К биологическим основам «стресса» и «адаптационного синдрома». Актуальные проблемы стресса. Кишинев: Штиинца, 1976.
Раушенбах И.Ю. Нейроэндокринная регуляция развития насекомых в условиях стресса: Генетико-физиологические аспекты. Новосибирск: Наука, 1990.
Раушенбах И.Ю. Стресс-реакция насекомых: механизм, генетический контроль, роль в адаптации. Генетика. 1997;33:1110-1118.
Раушенбах И.Ю., Лукашина Н.С., Корочкин Л.И. Влияние высокой плотности культуры на выживаемость, гормональный статус и активность ЮГ-эстеразы Drosophila virilis. Генетико-эндокринная регуляция развития Drosophila в экстремальных условиях среды. Сообщ. III. Генетика. 1983;19:1439-1445.
Раушенбах И.Ю., Суханова М.Ж., Хирашима А., Суцугу Е., Куано Е. Роль системы экдистероидов в регуляции размножения Drosophila в стрессирующих условиях среды. Докл. РАН. 2000; 375:568-570.
Салтыкова Е.С., Беньковская Г.В., Гайфуллина Л.Р., Новицкая О.П., Поскряков А.В., Николенко А.Г. Реакция отдельных физиологических барьеров при бактериальной инфекции у различных рас медоносной пчелы Apis mellifera. Журн. эволюц. биохимии и физиологии. 2005;41(3):254-258.
Селье Г. Стресс без дистресса. М.: Прогресс, 1979.
Селье Г. От мечты к открытию. М.: Прогресс, 1987.
Соколянская М.П., Беньковская Г.В., Николенко А.Г. Динамика формирования резистентности личинок комнатной мухи к стресс-факторам различной природы. Агрохимия. 2005;9: 70-75.
Тыщенко В.П. Физиология насекомых. Уч. пособие для студентов ун-тов, обучающихся по спец. «Биология». М.: Высш. шк., 1986.
Ченцова Н.А., Алексеев А.А., Грунтенко Н.Е., Раушенбах И.Ю. Влияние октопамина на активность экдизон-монооксигеназы у Drosophila. Докл. АН. 2007;415:559-561.
Черныш С.И. Реакция нейроэндокринной системы на повреждающие воздействия. Тр. Всесоюз. энтомол. о-ва. 1983;64:118-127.
Черткова Е.А. Изменение уровня дофамина в гемолимфе личинок капустной совки Mamestra brassicae (Lepidoptera: Noctuidae) и колорадского жука Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae) при различных патогенезах. Новые знания о паразитах. Матер. V Межрегион. конф. «Паразитологические исследования в Сибири и на Дальнем Востоке». Новосибирск, 2015;131-132.
Adamo S.A. The effects of the stress response on immune function in invertebrates: An evolutionary perspective on an ancient connection. Horm. Behav. 2012;62:324-330.
Adamo S.A. The stress response and immune system share, borrow, and reconfigure their physiological network elements: evidence from the insects. Horm. Behav. 2017a:88:25-30. DOI 10.1016/j.yhbeh.2016.10.003.
Adamo S.A. Stress responses sculpt the insect immune system, allowing for better defense in an ever – changing world. Dev. Comp. Immunol. 2017b;66:24-32. DOI 10.1016/j.dci.2016.06.005.
Adamo S.A. The effects of stress hormones on immune function may be vital for the adaptive reconfiguration of the immune system during fight-or-flight behavior. Integr. Comp. Biol. 2014;54(3):419-426. DOI 10.1093/icb/icu005.
Adamo S.A., Easy R., Kovalko I., MacDonald J., McKeen A., Swanburg T., Turnbull K.F., Reeve C. Predator stress-induced immunosuppression: trade-off, immune redistribution or immune reconfiguration? J. Exp. Biol. 2017;220:868-875. DOI 10.1242/jeb.153320.
Adamo S.A., Kovalko I., Mosher B. The behavioural effects of predator-induced stress responses in the cricket (Gryllus texensis): the upside of the stress response. J. Exp. Biol. 2013;216:4608-4614.
Ahmad S. Oxidative stress from environmental pollutants. Arch. Insect Biochem. Physiol. 1995;29:135-157. DOI 10.1002/arch.940290205.
Bednářová A., Krishnan N., Cheng I.-C., Večeřa J., Lee H.-J., Kodrík D. Adipokinetic hormone counteracts oxidative stress elicited in insects by hydrogen peroxide: in vivo and in vitro study. Physiol. Entomol. 2013;38:54-62. DOI 10.1111/phen.12008.
Bendena W.G., Donly B.C., Tobe S.S. Allatostatins: A growing family of neuropeptides with structural and functional diversity. Ann. N. Y. Acad. Sci. 1999;897:311-329.
Bi J.L., Felton G.W. Foliar oxidative stress and insect herbivory: Primary compounds, secondary metabolites, and reactive oxygen species as components of induced resistance. J. Chem. Ecol. 1995;21:1511-1530. DOI 10.1007/BF02035149.
Bogus M.I., Cymborowski B. Chilled Galleria mellonella larvae: Mechanism of supernumerary moulting. Physiol. Entomol. 1981;6: 343-348.
Bogus M.I., Wisniewski J.R., Cymborowski B. Effect of injury to the neuroendocrine system of last instar larvae of Galleria mellonella. J. Insect Physiol. 1986;32:1011-1018.
Cannon W.B. The Wisdom of the Body. N. Y.: W.W. Norton, 1932.
Chernysh S.I. Neuroendocrine system in insect stress. Eds. J. Ivanovic, M. Jankovic-Hladni. Hormones and Metabolism in Insect Stress. Boca Raton: CRC Press, 1991;69-98.
Cymborowski B. Effect of cooling stress on endocrine events in Galleria mellonella. Endocrinological frontiers in physiological insect ecology. Wroclaw. 1988;1:203-212.
Cymborowski B., Bogus M.I. Juvenilizing effect of cooling on Galleria mellonella. J. Insect Physiol. 1976;22:669-672.
Cymborowski B., Bogus M., Beckage N.E., Williams C.M., Riddiford L.M. Juvenile hormone titrres and metabolism during starvation-induced supernumerary larval moulting of the tabacco hornworm, Manduca sexta L. J. Insect Physiol. 1982;28:129-135.
Davenpont A.K., Evans P.D. Stress-indeced changed in octopamine levels of insect hemolymph. Insect Biochem. 1984;14:135-150.
Evans P.D. Octopamine. Eds. G.A. Kerkut, L.I. Gilbert. Comprehensive Insect Physiology, Biochemistry and Pharmacology. Oxford: Pergamon Press, 1985;11:499-538.
Even N., Devaud J.M., Barron A.B. General stress responses in the honey bee. Insects. 2012;3:1271-1298. DOI 10.3390/insects3041271.
Gruntenko N.E., Bogomolova E.V., Adonyeva N.V., Karpova E.K., Menshanov P.N., Alekseev A.A., Romanova I.V., Li S., Rauschenbach I.Y. Decrease in juvenile hormone level as a result of genetic ablation of the corpus allatum cells affects the synthesis and metabolism of stress related hormones in Drosophila. J. Insect Physiol.
2012;58:49-55. Gruntenko N.E., Bownes M., Terashima J., Suchanova M.Zh., Rauschenbach I.Yu. Heat stress affects oogenesis differently in wild type Drosophila virilis and a mutant with altered juvenile hormone and 20-hydroxyecdysone levels. Insect Mol. Biol. 2003a;12:393-404.
Gruntenko N.E., Chentsova N.A., Andreenkova E.V., Bownes M., Segal D., Adonyeva N.V., Rauschenbach I.Yu. Stress response in a juvenile hormone deficient Drosophila melanogaster mutant apterous 56f . Insect Mol. Biol. 2003b;12:353-363.
Gruntenko N.E., Chentsova N.A., Andreenkova E.V., Karpova E.K., Glazko G.V., Monastirioti M., Rauschenbach I.Yu. The effect of mutations altering biogenic amine metabolism in Drosophila on viability and the response to heat stress. Arch. Insect Biochem. Physiol. 2004;55:55-67.
Gruntenko N.E., Karpova E.K., Adonyeva N.V., Chentsova N.A., Faddeeva N.V., Alekseev A.A., Rauschenbach I.Yu. Juvenile hormone, 20-hydroxyecdisone and dopamine interaction in Drosophila virilis reproduction under normal and nutritional stress conditions. J. Insect Physiol. 2005a;51:417-425.
Gruntenko N.E., Karpova E.K., Alekseev A.A., Chentsova N.A., Saprykina Z.V., Bownes M., Rauschenbach I.Yu. Effects of dopamine on juvenile hormone metabolism and fitness in Drosophila virilis. J. Insect Physiol. 2005b;51:959-968.
Gruntenko N.E., Rauschenbach I.Yu. Interplay of juvenile hormone, 20-hydroxyecdisone and biogenic amines under normal and stress conditions and its effect on reproduction. J. Insect. Physiol. 2008; 54:902-908.
Gruntenko N.E., Rauschenbach I.Yu. The role of insulin signalling in the endocrine stress response in Drosophila melanogaster: A minireview. Gen. Comp. Endocrinol. 2017. DOI 10.1016/j.ygcen.2017. 05.019.
Gruntenko N.E., Wen D., Karpova E.K., Adonyeva N.V., Liu Y., He Q., Faddeeva N.V., Fomin A.S., Li S., Rauschenbach I.Yu. Altered juvenile hormone metabolism, reproduction and stress response in Drosophila adults with genetic ablation of the corpus allatum cells. Insect Biochem. Mol. Biol. 2010;40: 891-897.
Gruntenko N.E., Wilson T.G., Monastirioti M., Rauschenbach I.Yu. Stress-reactivity and juvenile hormone degradation in Drosophila melanogaster strains having stress-related mutations. Insect Biochem. Mol. Biol. 2000;30:775-783.
Harris J.W., Woodring J. Effects of stress age, season and source colony on the level of octopamine, dopamine and serotonin in the honey bee (Apis melifera L.) brain. J. Insect Physiol. 1992;38:29-35.
Hirashima A., Eto M. Chemical-induced changed in the biogenic amine levels of Periplaneta americana L. Pestic. Biochem. Physiol. 1993;46:131-140.
Hirashima A., Nagano T., Eto M. Effect of various insecticides on the larval growth and biogenic amine levels of Tribolium castaneum Herbst. Comp. Biochem. Physiol. 1994;107C:393-398.
Hirashima A., Rauschenbach I.Yu., Sukhanova M.Jh. Ecdisteroids in stress responsive and nonresponsive Drosophila virilis lines under stress conditions. Biosci. Biotech. Biochem. 2000;64:2657-2662.
Holzenberger M., Dupont J., Ducos B., Leneuve P., Géloën A., Even P.C., Cervera P., Le Bouc Y. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature. 2003;42: 182-187.
Ivanović J., Janković-Hladni M., Milanović M.P. Possible role of neurosecretory cells, Type A in response of Morimus funereus larvae to the effect of temperature. J. Therm. Biol. 1975;1:53-57.
Ivanović J., Janković-Hladni M., Stanić V., Milanović M.P., Nenadović V. Possible role of neurohormones in the process of acclimatization and acclimation in Morimus funereus larvae (Insecta). I. Changes in the neuroendocrine system and target organs (midgut, hemolymph) during the annual cycle. Comp. Biochem. Physiol. 1979;80A:107-112.
Ivanović J., Janković-Hladni M., Stanić V., Kalafatć D. Differences in the sensitivity of protocerebral neurosecretory cells arising from the effect of different factors in Morimus funereus larvae. Comp. Biochem. Physiol. 1985;80A:107-113.
Ivanović J., Janković-Hladni M., Stanić V., Nenadović V., Frušić M. The role of neurosecretion and metabolism in development of an oligophagous feeding habit in Morimus funereus larvae (Col., Cerambycidae). Comp. Biochem. Physiol. 1989;74:189-197.
Janković-Hladni M. Hormones and metabolism in insect stress (Historical survey). Eds. J. Ivanovic, M.I. Jankovic-Hladni. Hormones and Metabolism in Insect Stress. Boca Raton: CRS Press, 1991; 5-27.
Janković-Hladni M., Chen C.A., Ivanović J., Djordjević S., Stanić V., Perić Mataruga V., Frušić M. Effects of diet and temperature on Morimus funereus larvae hemolymph cation concentrations. Arch. Insect Biochem. Physiol. 1992;20:205-214.
Janković-Hladni M., Ivanović J., Nenadović V., Stanić V. The selective response of the protocerebral neurosecretory cells of the Cerambyx cerdo larvae to the effect of different factors. Comp. Biochem. Physiol. 1983;74A:131-136.
Johnson E.C., White M.P. Stressed-Out Insects: Hormonal Actions and Behavioral Modifications. Eds. D.W. Pfaff, A.P. Arnold, S.E. Fahrbach, A.M. Etgen, R.T. Rubin. Hormones, Brain and Behavior. San Diego: Academic Press, 2009;1069-1096.
Kodrík D., Bednářová A., Zemanová M., Krishnan N. Hormonal regulation of response to oxidative stress in insects – an update. Int. J. Mol. Sci. 2015;16:25788-25816.
Kramer S.J., Toschi A., Miller C.A., Kataoka H., Quistad G.B., Li J.P., Carney R.L., Schooley D.A. Identification of an allastatin from the tabacco hornworm Manduca sexta. Proc. Natl. Acad. Sci. USA. 1991;88:9458-9462.
Krishnan N., Kodrík D. Endocrine control of oxidative stress in insects. Eds T. Farooqui, A.A. Farooqui. Oxidative Stress in Vertebrates and Invertebrates: Molecular Aspects of Cell Signaling. New Jersey: Wiley-Blackwell, 2012;261-270.
Lalouette L., Williams C.M., Hervant F., Sinclair B.J., Renault D. Metabolic rate and oxidative stress in insects exposed to low temperature thermal fluctuations. Comp. Biochem. Physiol. 2011;158:229-234. DOI 10.1016/j.cbpa.2010.11.007.
Lawrence P.O. Hormonal interactions between parasitoids and hosts: adaptation to stress? Endocrinological Frontiers in Physiological Insect Ecology. Wroclaw: Tech. Univ. Press, 1988;423-435.
Leković S., Lazarević J., Nenadović V., Ivanović J. The effect of heat stress on the activity of A1 and A2 neurosecretory neurons of Morimus funereus (Coleoptera: Cerambycidae) larvae. Eur. J. Entomol. 2001;98:13-18.
McCaleb D.C., Kumaran A.K. Control of juvenile hormone esterase activity in Galleria mellonella larvae. J. Insect Physiol. 1980;26: 171-177.
Meng J.-Y., Zhang C.-Y., Zhu F., Wang X.-P., Lei C.-L. Ultraviolet light-induced oxidative stress: Effects on antioxidant response of Helicoverpa armigera adults. J. Insect Physiol. 2009;55:588-592. DOI 10.1016/j.jinsphys.2009.03.003.
Mobius P., Penzlin H. Stress-induced release of octopamine in the American cockroach Periplaneta americana L. Acta Biol. Hung. 1993;44:45-50.
Mrdaković M., Ilijin L., Vlahović M., Janković-Tomanić M., Perić Mataruga V., Lazarević J., Nenadović V. The effects of different constant temperatures on the activity of corpora allata in Morimus funereus (Coleoptera: Cerambycidae) larvae. Arch. Biol. Sci. 2003; 55:21-22.
Neckameyer W.S., Weinstein J.S. Stress affects dopaminergic signaling pathways in Drosophila melanogaster. Stress. 2005;8:117-131.
O’Kasha A.Y.K. Effect of sub-lethal high temperature on insect, Rhodnius prolixus (Stal). I. Induction of delayed moutling and defects. J. Exp. Biol. 1968;48:455-463.
Perić-Mataruga V., Lazarević J., Nenadović V. A possible role for the dorsolateral protocerebral neurosecretory neurons in the trophic adaptations of Lymantria dispar (Lepidoptera : Lymantriidae). Eur. J. Entomol. 2001;98:257-264.
Pszczolkowski M.A., Chiang A.S. Effects of chilling stress on allatal growth and juvenile hormone synthesis in the cockroach, Diploptera punctate. J. Insect Physiol. 2000;46:923-931.
Rauschenbach I.Y., Budker V.G., Korochkin L.I. Pupal esterase of D. virilis splits juvenile hormone. Dros. Inf. Serv. 1983a;59:104.
Rauschenbach I.Yu., Karpova E.K., Adonyeva N.V., Andreenkova O.V., Faddeeva N.V., Burdina E.V., Alekseev A.A., Menshanov P.N., Gruntenko N.Е. Disruption of insulin signalling affects the neuroendocrine stress reaction in Drosophila females. J. Exp. Biol. 2014;217:1-9.
Rauschenbach I.Y., Lukashina N.S., Korochkin L.I. Role of pupal esterase in the regulation of the D. virilis stosks differing in response to high temperature. Dev. Genet. 1980;1:295-310.
Rauschenbach I.Y., Lukashina N.S., Korochkin L.I. Genetic of esterase in Drosophila. VII. The genetic control of the activity level of the JH-esterase and heat-resistance in Drosophila virilis under high temperature. Biochem. Genet. 1983b;21:253-266.
Rauschenbach I.Y., Lukashina N.S., Maksimovsky L.F., Korochkin L.I. Stress-like reaction of Drosophila to advers environmental factors. J. Comp. Physiol. B. 1987;157:519-531.
Richard D.S., Applebaum S.W., Gilbert L.I. Allatostatic reulation of juvenile hormone production in vitro by the ring gland of Drosophila melanogaster. Mol. Cell. Endocrinol. 1990;68:153-161.
Singh H., Pandey P.N. Experimental assessment of effects of larval crowding on development and reproduction in Diacrisia obliqua Walker (Lep., Arctiidae). Biochem. Exp. Biol. 1980;16:157-164.
Stay B., Fairbairn S., Yu C.G. Role of allaststins in the regulation of juvenile hormone synthesis. Arch. Insect Biochem. Physiol. 1996;32: 287-297.
Sudakov K.V. Specific mechanisms of emotional stress. Third Symp. on Catecholamines and other Neurotransmitters in Stress, Smolenica Castle, Czchoslovakia. 1983;87.
Tauchman S.J., Lorch J.M., Orth A.P., GoodmanW.G. Effects of stress on the hemolymph juvenile hormone binding protein titres of Manduca sexta. Insect Biochem. Mol. Biol. 2007;37:847-854.
Tobe S.S., Stay B. Structure and regulation of the corpus allatum. Adv. Insect Physiol. 1985;18:305-432.
Velki M., Kodrík D., Večeřa J., Hackenberger B.K., Socha R. Oxidative stress elicited by insecticides: A role for the adipokinetic hormone. Gen. Comp. Endocrinol. 2011;172:77-84. DOI 10.1016/j.ygcen.2010.12.009.
Vigas M. Problem of definition of stress stimulus and specificity of stress response. Third Symp. on Catecholamines and other Neurotransmitters in Stress, Smolenica Castle, Czchoslovakia, 1983;94.
Wigglesworth V.B. Hormone balance and the control of metamorphosis in Rhodnius prolixus (Hemiptera). J. Exp. Biol. 1952;29:620-631.
Wigglesworth V.B. High temperature and arrested growth in Rhodnius: Quantitative requirements for ecdysone. J. Exp. Biol. 1955;32: 649-655.
Zhao H.W., Haddad G.G. Review: Hypoxic and oxidative stress resistance in Drosophila melanogaster. Placenta. 2011;32:S104-S108. DOI 10.1016/j.placenta.2010.11.017.
Источник Вавиловский журнал генетики и селекции. 2017. Т.21. №7. 825-832
Рекомендуем прочесть
 Паразитические манипуляции фенотипом хозяина: эффекты во внутренней и внешней среде
Паразитические манипуляции фенотипом хозяина: эффекты во внутренней и внешней среде Моллюски помогли ученым оценить масштабы нынешнего сокращения биоразнообразия
Моллюски помогли ученым оценить масштабы нынешнего сокращения биоразнообразия Антиподы: другая Земля
Антиподы: другая Земля Природный процесс генной модификации
Природный процесс генной модификации «Город как заказник»: (пока?) упускаемая возможность
«Город как заказник»: (пока?) упускаемая возможность Второе предупреждение ученых человечеству
Второе предупреждение ученых человечеству Хоботковые бабочки возникли до цветковых растений
Хоботковые бабочки возникли до цветковых растений Происхождение богомолов
Происхождение богомолов
