

Резюме. В продолжение темы стадий деградации дикой природы под антропогенным воздействием приведены данные о первой из них — плейстоценовом перепромысле. Оказывается, на всех континентах вымирание мегафауны обусловлено истреблением человеком, иногда вкупе с преобразованием ландшафтов огнём, а не климатом и другими естественными причинами. Также цитируется удивительно красивое доказательство перепромысла через анализ сегодняшней конкуренции овцебыка с северным оленем на изолированных арктических островах.
Позднечетвертичное вымирание крупных млекопитающих не имело аналогов в предшествующие эпохи
Содержание
- 1 Позднечетвертичное вымирание крупных млекопитающих не имело аналогов в предшествующие эпохи
- 2 Истребление древними охотниками крупных животных Австралии привело к необратимым изменениям природных экосистем
- 3 Вымирание крупных хищников в Восточной Африке лучше объясняется эволюцией гоминид, чем климатом
- 4 Овцебык против северного оленя: и древние охотники
Александр Марков
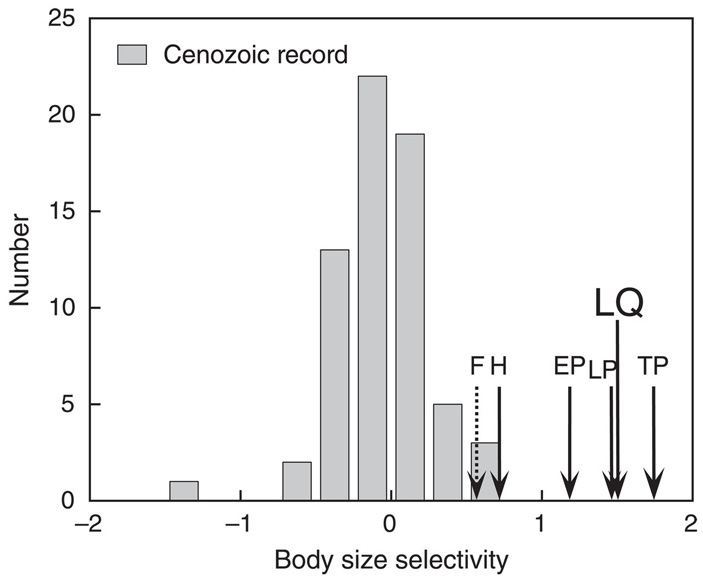
Рис. 1. Зависимость вероятности вымирания млекопитающих от массы тела в разные эпохи кайнозоя. Значения на горизонтальной оси показывают направленность и величину изменения вероятности вымирания вида с ростом массы тела (средний прирост натурального логарифма вероятности вымирания с увеличением массы тела в 10 раз). Положительные значения соответствуют преимущественному вымиранию крупных животных, отрицательные — мелких.
Высота столбиков отражает количество интервалов кайнозойской эры (длительностью по 1 млн лет каждый), когда наблюдался данный уровень избирательности. Стрелками показаны уровни, характерные для позднечетвертичного времени: LP — Late Pleistocene (125–70 тысяч лет назад), EP — End Pleistocene (70–20 тысяч лет назад), TP — Terminal Pleistocene (20–10 тысяч лет назад), H — Holocene (10–0 тысяч лет назад), LQ — Late Quaternary, среднее значение за всю позднечетвертичную эпоху (125–0 тысяч лет назад); F — Future, прогноз на ближайшие 200 лет (основан на предположении, что вымрут все виды, ныне находящиеся под угрозой вымирания). Рисунок из обсуждаемой статьи в Science
Анализ палеонтологических данных по кайнозойским млекопитающим показал, что характер вымирания видов резко изменился на всех континентах в позднечетвертичное время одновременно с расселением первобытных охотников. В течение всего кайнозоя крупные и мелкие млекопитающие вымирали примерно с одинаковой скоростью. Однако с появлением средне- и верхнепалеолитических охотников вымирание повсюду становилось резко избирательным: начинали исчезать в первую очередь крупные животные. Данная тенденция, результатом которой стало глобальное измельчание фауны млекопитающих, продолжается по сей день. Полученные результаты подкрепляют точку зрения о ведущей роли человека в позднечетвертичном вымирании мегафауны.
Споры о том, что было главной причиной позднечетвертичного вымирания крупных млекопитающих — человек или климат — продолжаются уже не первое десятилетие. «Элементы» не раз рассказывали о ходе этой бурной дискуссии (см. подборку ссылок в конце новости Главной причиной позднечетвертичного вымирания все-таки были люди, а не климат, «Элементы», 09.06.2014).
Важным аргументом против «человеческой» версии считается тот факт, что крупные животные вообще более уязвимы, поскольку их популяции имеют низкую численность (см.: Крупным млекопитающим может грозить вымирание из-за накопления вредных мутаций, «Элементы», 28.08.2007; Вероятность вымирания в позднем плейстоцене разных видов млекопитающих зависела от их размера, «Элементы», 07.12.2009). Конечно, первобытные охотники могли предпочитать (и действительно предпочитали) крупную добычу*. Однако и другие катаклизмы, в том числе климатические, по идее, тоже должны приводить к избирательному вымиранию крупных животных.
В таком случае мы должны ожидать, что крупные виды млекопитающих вымирали чаще, чем мелкие, не только в конце четвертичного периода, но и в другие эпохи. В том числе — при переменах климата, которые в кайнозое происходили многократно.
Палеонтологи уже накопили достаточно данных, чтобы не принимать это утверждение (при всем его правдоподобии) за аксиому. Его можно проверить, что и попытались сделать ученые из четырех американских университетов, чья статья опубликована в свежем выпуске журнала Science.
Авторы проанализировали данные по пространственно-временному распространению всех видов нелетающих наземных кайнозойских млекопитающих, по которым есть достаточно точные датировки первых и последних находок (2786 видов). Для разных временных интервалов оценивалась скорость вымирания видов в зависимости от массы тела. Кайнозой поделили на отрезки длительностью в 1 млн лет, и для каждого отрезка рассчитали степень избирательности вымирания в зависимости от размера животных.
Данные по позднечетвертичным (от 125 тысяч лет назад до настоящего времени) и современным видам анализировались отдельно. Всего был учтен 3301 вид, причем если вид существует сегодня, но не известен в ископаемом состоянии, то считалось, что он уже существовал 125 тысяч лет назад. Данные разделили на четыре части, соответствующие разным этапам экспансии средне- и верхнепалеолитических охотников (рис. 1):
1) «Поздний плейстоцен» (Late Pleistocene, 125–70 тысяч лет назад). В это время в Европе процветали неандертальцы, в Азии — вероятно, денисовцы, а группы сапиенсов начали периодически выходить за пределы Африки;
2) «Конец плейстоцена» (End Pleistocene, 70–20 тысяч лет назад). На этот период приходится быстрое расселение сапиенсов по Евразии, а также заселение Австралии;
3) «Терминальный плейстоцен» (Terminal Pleistocene, 20–10 тысяч лет назад). В это время сапиенсы заселили Америку;
4) Голоцен (10–0 тысяч лет назад).
Общий масштаб позднечетвертичного вымирания отражен в таблице (рис. 2).
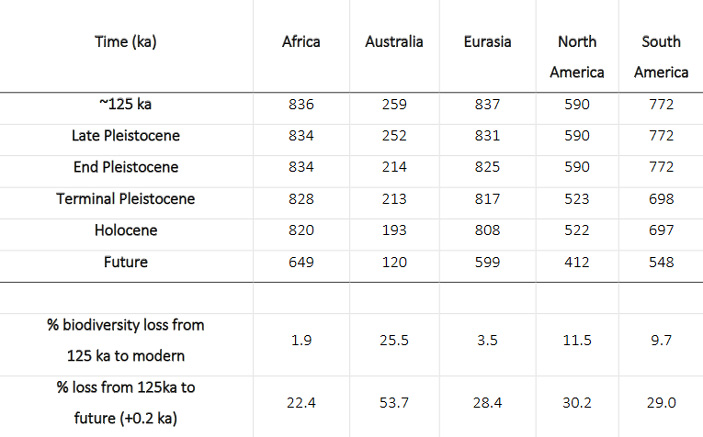
Рис. 2. Изменение числа видов нелетающих наземных млекопитающих на разных континентах в течение позднего плейстоцена. Нижние две строки показывают, на сколько процентов снизилось видовое разнообразие с начала позднего плейстоцена по настоящее время, а также ожидаемые потери в будущем, если вымрут все «краснокнижные» виды. Таблица из дополнительных материалов к обсуждаемой статье в Science
Оказалось, что в течение почти всей кайнозойской эры вымирание млекопитающих было неизбирательным по размеру (на рис. 1 это выражается в том, что самые высокие столбики расположены недалеко от нулевой отметки). Вымирали с равной вероятностью как мелкие, так и крупные виды. Около 29 млн лет назад, в олигоцене, был даже эпизод избирательного вымирания мелких млекопитающих, предположительно связанный с уменьшением площади лесов и распространением травяных биомов (степей и саванн).
Принципиально иная ситуация сложилась в позднечетвертичное время (стрелки на рис. 1). Уже в начале позднего плейстоцена, 125–70 тысяч лет назад, вымирание стало резко избирательным. Начиная с этого времени вымирали в основном крупные животные. Данная тенденция сохранялась и в дальнейшем — вплоть до настоящего времени. Правда, в голоцене избирательность немного сгладилась. Скорее всего, сглаживание связано с тем, что с развитием сельского хозяйства люди стали истреблять фауну другими способами. Охота отступила на второй план, а на первый вышло уничтожение местообитаний, от которого страдают не только крупные, но и мелкие животные.
Главный вывод состоит в том, что резко повышенная вероятность вымирания крупных животных, характерная для позднечетвертичного времени, вовсе не является универсальной закономерностью. В течение большей части кайнозоя такой избирательности не было. Это уникальная особенность именно позднечетвертичного вымирания, когда к издревле существовавшим причинам вымирания добавился новый суперхищник, овладевший эффективными методами охоты на крупных животных.
Выяснилось также, что резкие климатические изменения сами по себе не приводят к избирательному вымиранию крупной фауны. Сопоставив избирательность с палеоклиматическими данными, авторы не обнаружили связи между ними. Крупные и мелкие млекопитающие примерно одинаково реагировали на климатические изменения вплоть до позднего плейстоцена.
Избирательность позднечетвертичного вымирания выглядит уникальной не только на фоне предшествующей динамики разнообразия кайнозойских млекопитающих. Преимущественное вымирание крупных животных отмечается во время некоторых кризисов (см.: После массового вымирания на рубеже девона и карбона позвоночные измельчали, «Элементы», 16.11.2015). Однако столь сильного «перекоса» в сторону вымирания крупных видов, как у млекопитающих в позднем плейстоцене и голоцене, не было во время предшествующих массовых вымираний ни у других позвоночных, ни у беспозвоночных.
Результаты в целом хорошо согласуются с идеей о решающей роли людей в позднечетвертичном вымирании. Этот вывод становится еще очевиднее при детальном рассмотрении полученных данных. На рис. 3 показана динамика средней и максимальной массы тела млекопитающих в течение кайнозоя, в том числе отдельно по континентам. Видно, что вплоть до плейстоцена млекопитающие в среднем становились всё крупнее. Росла как средняя, так и максимальная масса тела существующих видов. Однако в позднем плейстоцене тенденция сменилась на противоположную.
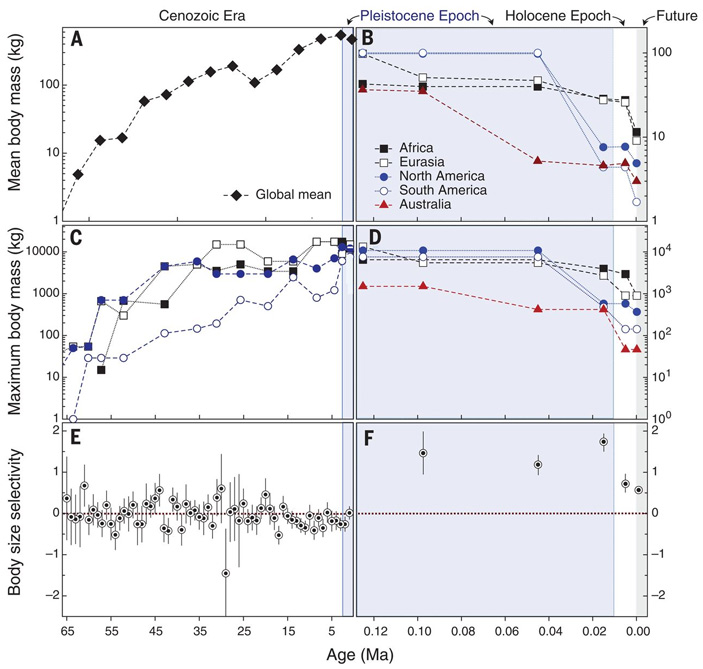
Рис. 3. Изменение размеров млекопитающих в течение кайнозоя. В каждой паре графиков слева показан весь кайнозой (от 66 млн лет назад до настоящего времени), справа — поздний плейстоцен и голоцен (от 125 000 лет до ближайшего будущего). По горизонтальной оси — время в млн лет назад. Первая пара графиков (A, B) — средняя масса тела, вторая (C, D) — максимальная масса тела, третья (E, F) — избирательность вымирания по размеру (как на рис. 1). Изображение из обсуждаемой статьи в Science
За последние 125 000 лет фауна млекопитающих резко измельчала, причем динамика этого процесса довольно точно отражает историю расселения первобытных охотников. Так, в Евразии размер млекопитающих пошел на убыль уже 125–70 тысяч лет назад (расцвет среднепалеолитической культуры неандертальцев, первые волны миграции сапиенсов). В Австралии катастрофическое измельчание млекопитающих приходится на период от 70 до 20 тысяч лет назад, а первые люди появились там около 65 тысяч лет назад. Любопытно, что в этот период Северная и Южная Америка, куда люди еще не добрались, ненадолго стали подлинными заповедниками мегафауны: обитавшие там звери были крупнее, чем где бы то ни было на планете. Резкое измельчание американских млекопитающих произошло позже — в «терминальном плейстоцене» (20–10 тысяч лет назад), когда туда прорвались люди из Берингии.
Что касается Африки, то на этом континенте позднечетвертичное вымирание мегафауны было выражено слабее всего. Это традиционно объясняют тем, что в Африке крупные звери коэволюционировали с гоминидами миллионы лет. Поэтому у них была возможность постепенно адаптироваться к уловкам плотоядных двуногих обезьян. Кроме того, не исключено, что в результате этой коэволюции размерное распределение африканских млекопитающих сместилось в сторону измельчания намного раньше, чем на остальных континентах. Как показано на рисунке 3, B, африканские виды млекопитающих по своему среднему размеру заметно уступали евразийским и американским (и были сравнимы с австралийскими) уже в начале позднего плейстоцена. Здесь нужно учитывать, что одним из факторов, способствующих эволюции крупной фауны, считается площадь континента. Этим объясняют, в частности, относительно более мелкие размеры представителей вымершей австралийской мегафауны по сравнению с плейстоценовыми млекопитающими Евразии и Америки. Тот факт, что уже 125 000 лет назад африканская фауна в среднем была не намного крупнее австралийской, авторы предположительно связывают с влиянием древних гоминид.
Ученые попытались также сделать прогноз на будущее, основанный на допущении, что вымрут все «краснокнижные» звери. Это приведет к еще большему измельчанию млекопитающих, а также к дальнейшему росту доли грызунов и снижению доли копытных в экосистемах.
Таким образом, полученные результаты свидетельствуют в пользу того, что деятельность гоминид сильно влияла на фауну млекопитающих и структуру сообществ еще в доисторические времена. Антропогенное вымирание видов, происходящее в наши дни, является не более чем продолжением давней тенденции. Авторы прозрачно намекают, что если дело и дальше так пойдет, то через пару веков фауна диких млекопитащих станет в основном фауной грызунов, а самым крупным наземным животным будет домашняя корова.
P. S. В исследовании есть один скользкий момент, связанный с неполнотой палеонтологической летописи, и довольно странно, что он не обсуждается ни в статье, ни в дополнительных материалах к ней. При анализе позднечетвертичного разнообразия учитывались данные по современным видам. Принималось, что все современные виды существовали уже в позднем плейстоцене, даже если по ним нет ископаемых находок. С одной стороны, это наверняка справедливо для большинства видов, потому что средняя продолжительность существования видов у млекопитающих составляет 1–2 миллиона лет, и если вид существует сегодня, то с большой вероятностью он существовал и 125 000 лет назад. С другой стороны, мелкие кости хуже сохраняются в ископаемом состоянии, чем крупные. Иными словами, не исключено, что палеонтологическая летопись крупных животных в принципе полнее. Поэтому такая процедура должна была искусственно завысить долю мелкого зверья в позднечетвертичное время по сравнению с остальными эпохами кайнозоя, для которых использовались только палеонтологические данные. И даже не просто мелкого, а мелкого и не вымершего. Это могло привести к завышению уровня избирательности позднечетвертичного вымирания. Возможно, есть основания полагать, что данный эффект слаб и не мог повлиять на выводы, но авторам, на мой взгляд, следовало бы сказать об этом прямо.
Источник: Felisa A. Smith, Rosemary E. Elliott Smith, S. Kathleen Lyons, Jonathan L. Payne. Body size downgrading of mammals over the late Quaternary // Science. 2018. V. 360. P. 310–313. DOI: 10.1126/science.aao5987.
Источник Элементы.ру
Истребление древними охотниками крупных животных Австралии привело к необратимым изменениям природных экосистем
Алексей Гиляров
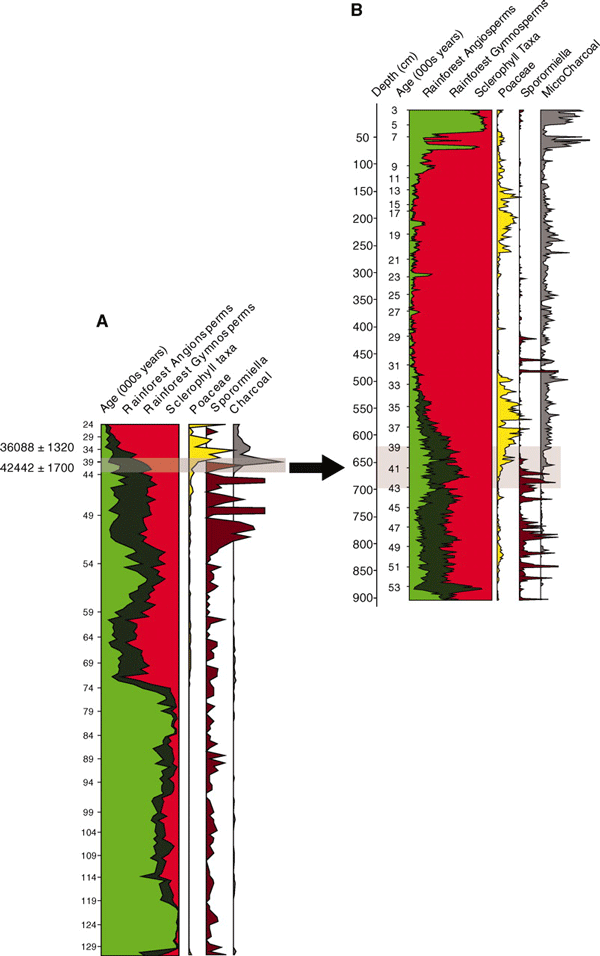
Результаты анализа первого (A) и второго (B) кернов (колонок) озерных отложений из «кратера Линча» за 127 тыс. лет. Серым цветом показан участок отложений, где резко уменьшилось содержание спор Sporormiella и впервые появились частички древесного угля. Видимо в это время и произошло истребление охотниками крупных травоядных животных. Шкала времени (тысячи лет) — слева от каждого графика. Глубина залегания — слева от графика B. Разными цветами показано относительное обилие спор и пыльцы (слева направо): покрытосеменных дождевого леса (выделено зеленым); голосеменных дождевого леса (выделено черным); склерофильной растительности (выделено красным); злаков, или мятликовых (выделено желтым); спор Sporormiella (выделено темно-красным); древесного угля (выделено серым). Изображение из обсуждаемой статьи в Science
Спор о том, что явилось причиной массового вымирания крупных животных в среднем и позднем плейстоцене (40–12 тыс. лет назад) — изменение климата или истребление древними охотниками, — в случае Австралии, видимо, можно считать закрытым. Крупные травоядные сумчатые были истреблены вскоре после того, как люди добрались до этого континента, примерно 40 тыс. лет назад. Детальное изучение древних озерных отложений (образовавшихся в период с 130 до 3 тыс. лет назад) на северо-востоке Австралии показало, что крупные животные исчезли очень быстро, причем никаких изменений климата и характера растительности этому не предшествовало. А вот после их вымирания были отмечены значительные изменения растительного покрова и всего ландшафта в целом. Изменения климата, наблюдавшиеся как задолго до исчезновения крупных животных, так и после него, не могли быть причиной их вымирания.
Массовое вымирание так называемой мегафауны — крупных млекопитающих (как плацентарных, так и сумчатых), крупных нелетающих птиц и некоторых рептилий, наблюдавшееся практически повсеместно в среднем и позднем плейстоцене, необычайно интенсивно обсуждается в научной литературе. Хотя гипотез, объясняющих это явление, довольно много (для Северной Америки их предложено аж 13!), основных две. Первая — изменение климата и вызванное им сокращение площади ландшафтов, пригодных для обитания тех или иных животных (например, тундростепей в случае мамонтов; см.: Мамонты вымерли потому, что им негде стало жить, «Элементы», 05.04.2008). Вторая — прямое истребление животных древними охотниками (во многих случаях вымирание действительно отмечалось именно тогда, когда до данного района добирались расселяющиеся племена Homo sapiens). Есть и компромиссное объяснение: изменение климата привело к резкому сокращению ареалов и численности крупных животных, а человек уже добил их.

Самое крупное существовавшее на Земле сумчатое — дипротодон (Diprotodon optatum). Будучи размером с носорога, этот зверь на самом деле является родственником вомбата и коалы. Дипротодоны были полностью истреблены древними охотниками. Изображение с сайта www.physorg.com
Группа австралийских ученых недавно опубликовала в журнале Science результаты исследования, убедительно показавшего, что в Австралии (по крайней мере, в северо-восточной ее части) именно охота древнего человека, а не изменение климата, стала причиной исчезновения ряда крупных травоядных млекопитающих. Выпадение же из экосистемы этих животных привело к существенному изменению не только растительности данного региона, но и всего ландшафта.
Материалом для анализа послужили две колонки (керна) озерно-болотных отложений, взятых в «кратере Линча» в тропической северо-восточной части Австралии. Сейчас этот кратер никак не выражен — судя по фотографиям, это ровная местность, покрытая травянистой растительностью. Но на самом деле там когда-то существовало окруженное лесом озеро, потом превратившееся в болото. Одна колонка (№1) включала отложения, образовавшиеся примерно за 100 тыс. лет (с 130 до 24 тыс. лет назад). Другая (№2), проанализированная более детально, охватила период в 50 тыс. лет (от 54 до 3 тыс. лет назад). Для каждой колонки получена детальная картина послойного распределения пыльцы разных растений (отдельно оценены покрытосеменные и голосеменные), частиц древесного угля (свидетельства пожаров), а также содержания спор грибов рода Sporormiella. Присутствие последних — очень важный показатель, поскольку прорасти и дать плодовые тела эти споры могут, только если попадут в помёт, оставленный крупными травоядными животными (такие вырастающие на экскрементах грибы называют копрофилами). Соответственно, исчезновение грибов Sporormiella — свидетельство исчезновения крупных травоядных млекопитающих (см. об этом подробнее Вымирание мамонтов и мастодонтов в Северной Америке могло быть причиной, но не следствием изменения растительности, «Элементы», 30.11.2009).

Однако 41 тыс. лет назад условия резко меняются. Споры Sporormiella практически перестают встречаться. По времени это совпадает как раз с исчезновением мегафауны в ископаемых остатках в других местах Австралии. В изученных отложениях резко возрастает количество древесного угля, что свидетельствует о большом количестве пожаров (скорее всего, вызванных уже деятельность человека). Дождевые леса уступают место склерофильной растительности. Исследователи подчеркивают, что отмеченные резкие изменения в характере растительного покрова происходили на фоне в общем не меняющегося климата. Решающим фактором было, по-видимому, истребление крупных травоядных животных, которые выедали большое количество веточного корма, а кроме того разрыхляли почву, способствуя интенсивному круговороту элементов минерального питания. В отсутствие крупных животных на земле скапливалось много горючего растительного материала, который легко возгорался уже не только от редких молний, но и от костров первобытных охотников (не исключено, что они использовали и активные палы растительности).
Все эти процессы довольно быстро охватили Австралию и привели к существенному изменению доминирующих ландшафтов. Именно тогда при возрастании засушливости климата дождевые леса во многих местах сменились очень сухими лесами из деревьев с ярко выраженными склерофильными чертами. Современные эвкалиптовые леса — это, по сути, леса уже такого типа.
Таким образом, истребление первобытными охотниками крупных травоядных животных, по мнению авторов обсуждаемой статьи, явилось основной причиной не только вымирания самих этих животных, но и последовавшей за этим радикальной трансформации доминирующих ландшафтов.
Источники:
1) Susan Rule, Barry W. Brook, Simon G. Haberle, Chris S. M. Turney, A. Peter Kershaw, Christopher N. Johnson. The aftermath of Megafaunal extinction: ecosystem transformation in Pleistocene Australia // Science. 2012. V. 335. P. 1483–1486.
2) Matt McGlone. The hunters did it // Science. 2012. V. 335. P. 1452–1453.
Источник Элементы.ру
Вымирание крупных хищников в Восточной Африке лучше объясняется эволюцией гоминид, чем климатом
Александр Марков

Рис. 1. Некоторые крупные хищники, когда-то обитавшие в Восточной Африке вместе с гоминидами. 1) Саблезубая кошка Dinophelis sp. (тащит убитого австралопитека). В изученном районе в рассматриваемое время встречалось как минимум три вида этого рода. Возраст самых старых находок 4,23–4,07 млн лет, самых молодых 1,0–0,9 млн лет. Рисунок Романа Евсеева с сайта oursociety.ru. 2) Саблезубая кошка Homotherium sp. (от 4,35–4,1 до 1,64–1,39 млн лет). Рисунок © Peter Hutzler с сайта www.newdinosaurs.com. 3) Гигантская гиена Pachycrocuta sp. (от 3,85–3,63 до 3,2–3,0 млн лет). Рисунок © Mauricio Antón с сайта scientificamerican.com. 4) Гигантская выдра Enhydriodon dikikae. В изученном районе водилось 4 вида этого рода, и все были крупные. Возраст самых древних находок гигантских выдр на этой территории — 4,23–4,07, самых молодых — 2–1,88 млн лет
Связь вымирания крупных животных, в том числе хищников, с расселением Homo sapiens хорошо документирована для позднего плейстоцена и голоцена, особенно в Австралии и Америке. Скрупулезный анализ восточноафриканской палеонтологической летописи за последние 4 млн лет показал, что и в более древние эпохи эволюция гоминид, скорее всего, была важной причиной снижения разнообразия крупных хищников. Динамика их вымирания в Восточной Африке, по-видимому, никак не связана с колебаниями климата, однако модель, связывающая темпы вымирания хищников с ростом мозга гоминид, хорошо согласуется с имеющимися данными. Возможно, вначале гоминиды подрывали кормовую базу крупных хищников, воруя у них добычу, а потом — охотясь на крупных травоядных.
Палеонтологи накопили немало данных, указывающих на важную роль поздних представителей рода Homo (в первую очередь, конечно, Homo sapiens, но не только) в вымирании крупных животных в позднечетвертичное время, то есть в последние 130 тысяч лет (см.: Главной причиной позднечетвертичного вымирания все-таки были люди, а не климат, «Элементы», 09.06.2014).
Неоднократно высказывалась гипотеза, что и в более древние времена деятельность гоминид тоже могла способствовать вымиранию крупных животных, в том числе хищников. Конечно, вряд ли австралопитеки, хабилисы или даже эректусы систематически охотились на крупных хищников. Но они могли охотиться на крупных травоядных, подрывая тем самым кормовую базу хищников. Кроме того, еще до изобретения эффективных способов охоты на крупную дичь гоминиды могли заниматься агрессивным клептопаразитизмом, то есть отгонять хищников от их добычи. Некоторые антропологи предполагают, что для этого могли использоваться колючие ветки (и, конечно, были необходимы слаженные коллективные действия бесстрашных двуногих обезьян). Даже если ранние гоминиды не охотились на крупную дичь и не вступали в открытую конфронтацию с хищниками, они могли потихоньку воровать добычу у каких-нибудь зазевавшихся гомотериев — это тоже могло негативно сказаться на долгосрочных перспективах выживания последних.
Понемногу накапливаются эмпирические данные, согласующиеся с гипотезой о влиянии древних гоминид на вымирание крупных хищников (L. Werdelin, M. E. Lewis, 2013. Temporal Change in Functional Richness and Evenness in the Eastern African Plio-Pleistocene Carnivoran Guild). В статье, опубликованной 13 января в журнале Ecology Letters, шведские палеонтологи привели новые аргументы в пользу этой гипотезы.
Авторы проанализировали данные по относительно небольшому, но исключительно хорошо изученному в палеонтологическом отношении району Восточной Африки (рис. 2, h). Этот район, охватывающий часть территорий Эфиопии, Кении и Танзании, на протяжении последних четырех миллионов лет был важным центром эволюции гоминид. Ископаемая летопись гоминид здесь весьма богата и прослеживается от ардипитеков, живших около 4 млн назад, до современного человека включительно.
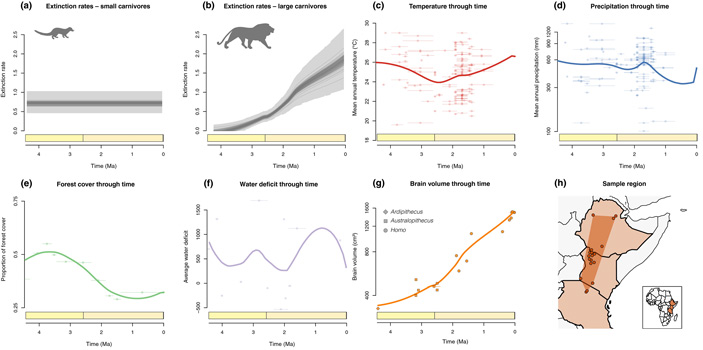
Рис. 2. Вымирание восточноафриканских хищников и его возможные причины. a, b — модели, лучше всего описывающие динамику вымирания мелких (а) и крупных (b) хищников. По горизонтальной оси — время в млн лет назад, по вертикальной — темп вымирания (число вымирающих видов в расчете на эволюционную линию за миллион лет). Для мелких хищников оптимальной оказалась модель с постоянной скоростью вымирания видов, для крупных — модель, предполагающая линейную зависимость скорости вымирания хищников от объема мозга местных гоминид. c, d, e, f — оценки температуры, количества осадков, площади лесов и дефицита влаги в изучаемом районе (дефицит воды оценивался по соотношению изотопов кислорода в зубной эмали травоядных, см.: S. A. Blumenthal et al., 2017. Aridity and hominin environments). g — объем мозга местных гоминид (ромбик — ардипитек, квадратики — австралопитеки, кружочки — Homo). h — карта изученного района, кружочками показаны местонахождения ископаемой фауны (палеоклиматические реконструкции основаны на данных из тех же местонахождений). Рисунок из обсуждаемой статьи в Ecology letters
Местная плиоценовая и плейстоценовая фауна хищных млекопитающих изучена очень подробно, причем один из авторов обсуждаемой статьи, Ларс Верделин (Lars Werdelin), внес важный вклад в ее изучение. Именно он собрал и выверил массив данных по ископаемым восточноафриканским хищникам, на котором основано исследование. То, что все находки проверялись и интерпретировались одним и тем же специалистом — важное достоинство работы. Это позволило минимизировать неопределенность, часто возникающую в подобных исследованиях из-за того, что у разных палеонтологов могут быть разные взгляды на систематику изучаемых групп и таксономическую принадлежность находок.
Массив включает 88 видов хищных млекопитающих, живших на изучаемой территории в течение последних 4 млн лет, и 279 «нахождений» (occurences), то есть комбинаций «место/время/вид животного». Лишь девять из 88 видов до сих пор здесь живут, все остальные вымерли (рис. 1). Среди вымерших крупных хищников — многие виды гиен, гигантские выдры, куницы и циветы, саблезубые кошки и медведи Agriotherium sp. Полный список видов и датировок приведен в дополнительных материалах к обсуждаемой статье.
Все хищники были подразделены на крупных (с массой тела более 21 кг) и мелких. Такое разделение является более или менее общепринятым среди специалистов по экологии хищных зверей, поскольку отражает существенные различия в способах добычи пропитания и спектре доступных жертв. Мелкие хищники, как правило, охотятся на множество видов мелких жертв, затрачивая на каждую охоту не слишком много времени и сил. Крупные хищники полагаются в основном на крупную добычу, поэтому охотятся реже, но с гораздо большими затратами. Эти различия важны в контексте предполагаемого влияния гоминид: например, клептопаразитизм вряд ли способен сильно навредить мелким хищникам, но может представлять серьезную проблему для крупных.
Данные по динамике разнообразия хищников сопоставлялись с опубликованными палеоклиматическими реконструкциями (рис. 2, c–f). Важно, что данные, на которых основаны эти реконструкции, собирались в тех же точках, что и ископаемая фауна.
Палеонтологические данные, особенно по континентальной (а не морской) фауне, всегда страдают неполнотой. К тому же видовая принадлежность находок часто сомнительна, да и датировки не бывают абсолютно точными. Однако для хищников рассматриваемого района и временного интервала все эти погрешности относительно невелики. С оставшейся неопределенностью авторы боролись всеми доступными статистическими способами, включая новые методы и компьютерные программы, основанные на байесовской статистике и разработанные самим авторами (D. Silvestro et al., 2019. Improved estimation of macroevolutionary rates from fossil data using a Bayesian framework).
Реконструированная динамика видового разнообразия крупных хищников в рассматриваемом районе показана на рис. 3.
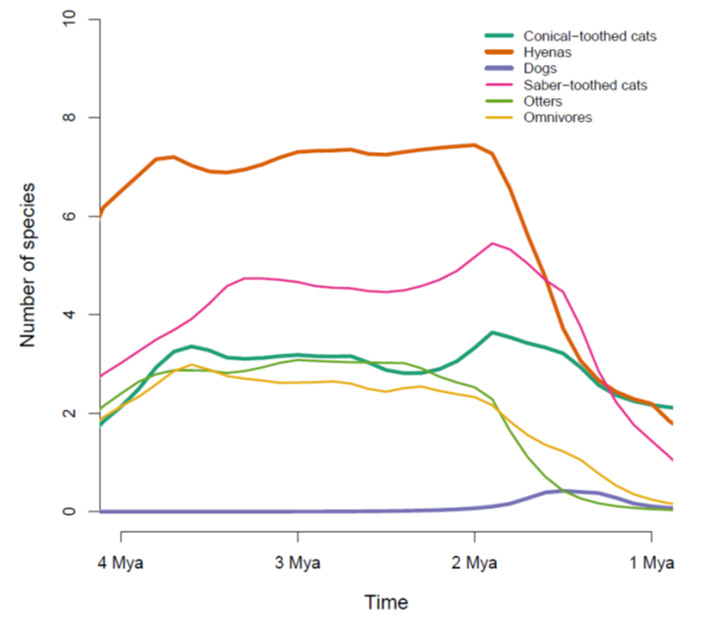
Рис. 3. Реконструкция динамики видового разнообразия крупных хищников в рассматриваемом районе. Показаны данные по шести важнейшим группам: кошачьи с коническими зубами, то есть несаблезубые (Conical-toothed cats), гиены (Hyenas), псовые (Dogs), саблезубые кошки (Saber-toothed cats), гигантские выдры (otters) и «всеядные» (Omnivores). В последнюю группу входят вымерший медведь Agriotherium, гигантская куница и три вида гигантских цивет. По горизонтальной оси — время в млн лет назад, по вертикальной — число видов. То, что это число обычно нецелое, связано с тем, что разнообразие реконструировалось сложными статистическими методами на основе заведомо неполной выборки и неточных датировок. Толстыми линиями показаны группы, существующие по сей день, тонкими — исчезнувшие. Рисунок из дополнительных материалов к обсуждаемой статье в Ecology Letters
В целом данные указывают на высокое разнообразие крупных хищников от 4 до 2 млн лет назад и на быстрый последующий упадок. Примечательно, что упадок происходил почти синхронно во всех группах, за исключением разве что псовых (которые, впрочем, никогда не играли большой роли в местных экосистемах). Это трудно объяснить какими бы то ни было внутренними закономерностями эволюции крупных хищников, не привлекая внешних факторов, таких как климатические изменения или, например, эволюция гоминид.
Изощренный статистический анализ показал то, что в общем-то и так видно на приведенных рисунках: никакой связи между вымиранием крупных хищников и колебаниями климата (температуры, количества осадков) не прослеживается. Из рассмотренных переменных только две могут претендовать на роль факторов, способствовавших упадку крупных хищников: прогрессивная эволюция гоминид, отразившаяся в увеличении объема мозга (рис. 2, g), и сокращение площади лесов (рис. 2, e).
Именно это и показали примененные авторами статистические методы. Из большого числа рассмотренных моделей, связывающих динамику вымирания с теми или иными факторами (рассматривались также модели, предполагающие постоянную скорость вымирания либо одно-два спонтанных изменения этой скорости на протяжении рассмотренного интервала), для крупных хищников самыми правдоподобными оказались модели, предполагающие прямую зависимость скорости вымирания от объема мозга гоминид, либо (с меньшей поддержкой) обратную зависимость от площади лесов.
Для мелких хищников наибольшую поддержку получила модель, предполагающая неизменную во времени скорость вымирания. Авторы не обнаружили никаких указаний на то, что скорость вымирания мелких хищников существенно менялась в течение последних четырех миллионов лет. А ведь если бы дело было в климатических изменениях, то скорости вымирания крупных и мелких животных должны были бы, по идее, меняться согласованно (см.: Позднечетвертичное вымирание крупных млекопитающих не имело аналогов в предшествующие эпохи, «Элементы», 23.04.2018). Но в действительности скорость вымирания мелких хищников не менялась, а крупных — росла.
Таким образом, результаты показывают, что упадок крупных хищников в Восточной Африке мог быть связан либо с эволюцией гоминид, либо с сокращением лесов. Разделить эти две гипотезы на основе данных только по одному району не удается, потому что между ростом мозга гоминид и сокращением лесов в Восточной Африке прослеживается сильная корреляция (оба процесса шли параллельно: мозг рос, а леса сокращались).
Чтобы разрешить эту дилемму, авторы привлекли данные по соотношению крупных и мелких хищников в разных районах современной Африки. Идея была в том, что те факторы, которые вызвали упадок крупных (но не мелких) хищников в Восточной Африке в плейстоцене, наверное, должны и сегодня приводить к пониженной доле крупных видов в общем разнообразии хищных млекопитающих.
Анализ показал, что в наши дни доля крупных хищников в разных районах Африки связана отрицательной зависимостью со степенью антропогенного воздействия и количеством осадков, но практически не зависит от температуры. Влияние площади лесов тоже прослеживается, хотя и слабое. Опираясь на эти современные корреляции между долей крупных хищников и осадками или площадью лесов, авторы попытались «предсказать» динамику доли крупных хищников за последние 4 млн лет (без учета антропогенного фактора). Получилось, что если бы соотношение крупных и мелких хищников «управлялось» осадками и площадью лесов так же, как в наши дни, то оно должно было бы оставаться примерно постоянным в плиоцене и плейстоцене. Это явно противоречит наблюдаемому упадку крупных хищников (на фоне сохранения разнообразия мелких) в последние 2 млн лет. Это можно рассматривать как косвенный аргумент в пользу того, что главной причиной упадка крупных хищников были все-таки люди.
Еще один аргумент удалось получить, сравнив африканские данные с аналогичной (хотя и менее детальной и выверенной) информацией по плиоцену и плейстоцену Северной Америки. Дело в том, что в течение нескольких последних миллионов лет сокращение лесов происходило не только в Африке: это был глобальный процесс, захвативший в том числе и Северную Америку. Однако в Северной Америке вплоть до самого конца плейстоцена не было гоминид.
Палеонтологические данные показывают, что сокращение лесов в Северной Америке не сопровождалось упадком крупных хищников. Даже наоборот: доля крупных хищников в общем разнообразии североамериканских хищных млекопитающих увеличилась в плейстоцене по сравнению с плиоценом (от 34–37% до 42–43%). Это делает еще менее правдоподобной гипотезу о том, что упадок восточноафриканских крупных хищников был вызван сокращением лесов.
Таким образом, полученные результаты свидетельствуют в пользу важной роли древних гоминид в вымирании восточноафриканских крупных хищников. Пока трудно сказать, каким был основной механизм влияния гоминид на хищников (охота, клептопаразитизм или что-то еще), и когда началось это влияние. Авторы попробовали разобраться в этом, сравнивая модели двух типов: первые предполагают, что скорость вымирания крупных хищников всегда была прямо пропорциональна объему мозга гоминид, а вторые — что гоминиды начали влиять на вымирание хищников не сразу, а лишь начиная с какого-то момента (будь то момент изобретения охоты на крупных животных, приручение огня или что-то еще). В итоге модели второго типа проиграли в статистическом состязании моделям первого типа. Таким образом, нет оснований говорить, что некие примитивные варианты поведения гоминид еще не влияли на вымирание хищников, а более продвинутые стали влиять. Похоже, это влияние началось еще в очень давние времена, когда ни о какой охоте на крупную дичь не могло быть и речи.
Может показаться, что авторы делают слишком смелые выводы на основе пусть и тщательно собранного и детально изученного, но всё-таки скудного материала. Напомним, что речь в конечном счете идет всего лишь о 279 палеонтологических находках (привязках того или иного вида хищников к месту и времени). Однако такая ситуация как для палеонтологии, так и для археологии палеолита не только типична, но и совершенно неизбежна: материал всегда катастрофически неполон, датировки приблизительны, видовые определения часто спорны. В данном случае исследователи сделали всё возможное, чтобы извлечь из имеющегося материала максимум полезной информации.
Источник: Søren Faurby, Daniele Silvestro, Lars Werdelin, Alexandre Antonelli. Brain expansion in early hominins predicts carnivore extinctions in East Africa // Ecology Letters. 13 January 2020. DOI: 10.1111/ele.13451
Источник Элементы.ру
Овцебык против северного оленя: и древние охотники
В продолжение темы плейстоценового перепромысла
В последнем ЖОБ’е есть хорошее доказательство сабжа. Дело в том, что в ходе плейстоцен-голоценового вымирания идёт выпадение сильнейших конкурентов, вроде мамонта с окружением, и сохранение слабейших, менее специализированных, вроде сайгака с северным оленем. Понятно, такое требует подозревать роль охоты. Работа И.С.Шереметева, С.Б.Розенфельд, Т.П.Сипко и А.Р.Груздева «Вымирание крупных травоядных млекопитающих: нишевые параметры сосуществующих в изоляции овцебыка Ovibos moschatus и северного оленя Ranfiger tarandus» (№1, т.75, с.62-73) подтверждает эти подозрения на примере северного оленя и овцебыка.
Как отмечают авторы, наиболее серьёзные доводы в пользу антропогенной гипотезы вымирания и против климатической таковы:
1. Рост числа ископаемых находок, расширение возможности датировок, уточнение датировок всё чётче показывают, что вымираниям мегафауны в конкретном регионе предшествовало поведение человека, и именно изобрётшего более убийственные орудия. Для разных регионов, где ситуация была спорной, данные всё чётче склоняются в пользу перепромысла, и нигде – от антропогенной гипотезы к климатической.
2. Сравнительно крупноразмерные виды вымирали быстрее мелкоразмерных, что соответствует стратегии и первобытной, и воременной охоты.
3. Массивность костных отложений, т.н. кухонные отбросы показывают внушительность охотничьих запросов древнего человека и более чем значительный масштаб истребления.
4. В увязке климатических и геологических событий с вымиранием мегафауны заложен порочный круг аргументации. Восстановление климатологами последовательности смен сильно зависит от данных об изменениях растительности и животного мира (на этой стадии антропогенная гипотеза не только не отведена, но в последнее время получает новые подтверждения). А дальше палеонтологии с биогеографами, не задумываясь, используют эти реконструкции для своей аргументации.
5. Изменения климата далеко не всегда совпадают с вымиранием конкретных видов млекопитающих в данном регионе. Поскольку потепления и похолодания происходили периодически, почти всегда было несколько таких же изменений климата и ранее, однако не приводивших к исчезновению данного вида.
6. Смены растительности (в рамках климатических гипотез опосредующих воздействие изменений на мегафауну) не предшествуют, а сопровождают процесс вымирания. Что естественно, учитывая эдификаторную роль мегафауны в тундростепях и других населённых ею травянистых биомах.
Сделать выбор между гипотезами можно, анализируя роль межвидовой конкуренции между более крупными и специализированными видами травоядных млекопитающих (особо склонными к вымиранию в плейстоцене) и менее специализированными, размером поменьше. В биоценотическом плане наиболее важны три аспекта конкуренции: сходство ресурсных спектров (перекрывание ниш), недостаток используемых сообща ресурсов (когда для выживания популяции особенно важны ресурсы, находящиеся в исключительном использовании одного вида и недоступные второму) и собственно результат конкуренции в виде вымирания одних травоядных и выживания других. Недостаток используемых сообща ресурсов равно используется в обоих гипотезах. Согласно климатической, изменения растительности происходили так, что площадь местообитаний сократилась, уменьшив свободное от хищника пространство, согласно антропогенной, распространившийся человек сократил площадь местообитаний, доступных для травоядных.
Тогда ключевым из интересующих нас факторов прошлого остаётся перекрывание ниш, допущение о котором обосновывается высоким разнообразием копытных мамонтовой фауны. Во всяком случае, современные травоядные, ранее входившие в её состав, показывают значительное перекрывание ниш. Само по себе доказательство организующей роли межвидовой конкуренции в сообществах крупных млекопитающих не удаётся использовать за или против одной из гипотез вымирания. Напротив, доказательство обратного прямо доказывают решающую роль первобытного перепромысла, т.е. антропогенного фактора.
Действительно, в современных сообществах травоядных, сильно разреженных человеком, даже в Африке вне нацпарков, последствий межвидовой конкуренции за корм обнаружить не удаётся. Возможно, здесь кроется основная причина нехватки эмпирических доказательств организующей роли соперничества. А вот охраняемые сообщества травоядных там же демонстрируют сверхизобилие сильнейших соперников. Тогда перепромысел травоядных мамонтовой фауны, как всякая катастрофа, не позволяет этим видам (крупным и специализированным) достичь плотности популяций, близкой к пределу ёмкости среды и соперничать за ресурсы. Поэтому всякие факты, демонстрирующие вымирание сильнейшего соперника прежде слабейшего, однозначно подтверждают гипотезу о преимущественно антропогенной причине вымирания, общее представление о котором можно даже сформулировать точнее: не просто сокращение ёмкости среды, а поголовное истребление.
Что и было показано авторами на паре видов мамонтовой фауны, которые сохранились до настоящего времени в условиях, ненамного отличающихся от существовавших в конце плейстоцена – начале голоцена. Это популяции овцебыка Ovibos moschatus и северного оленя Ranfiger tarandus на арктических островах вроде о.Врангеля. Заповедный режим, изоляция, ограниченность территории острова исключают взаимодействие с прочими крупными травоядными, делают конкуренцию этих видов неизбежной и сводят на нет возможные преимущества одного из соперников, не связанные со структурой ниши. Скажем, здесь не переместишься на участки, где соперник отсутствует. В этом плане остров с парой конкурентов, сообща используемыми ресурсами разных видов растений и пространством мало отличается от микрокосма Гаузе.
В плейстоцене ареал северного оленя в Евразии достигал побережья Средиземного и Чёрного морей, а овцебыка – севера Монголии и Казахстана. Дальше произошли 3 события, ставшие ключевыми в решении вопроса о роли соперничества:
1) Оба вида сократили ареал, но в разной степени. Северный олень не так значительно (хоть ушёл из умеренной полосы, но живёт на двух континентах), а вот овцебык очень сильно. Если не считать мест интродукций, он остался только в Гренландии и на севере Канады.
2) Рост численности и расширение ареала овцебыка в конце ХХ века, когда проблемы охраны и интродукций стали усиленно решаться.
3) На арктических островах в условиях охраны тенденции изменения численности этих двух видов полностью противоположны описанному в 1). Численность особей северного оленя уменьшается, овцебыка же возрастает. Что наблюдалось в сходных условиях на 3-х арктических островах:
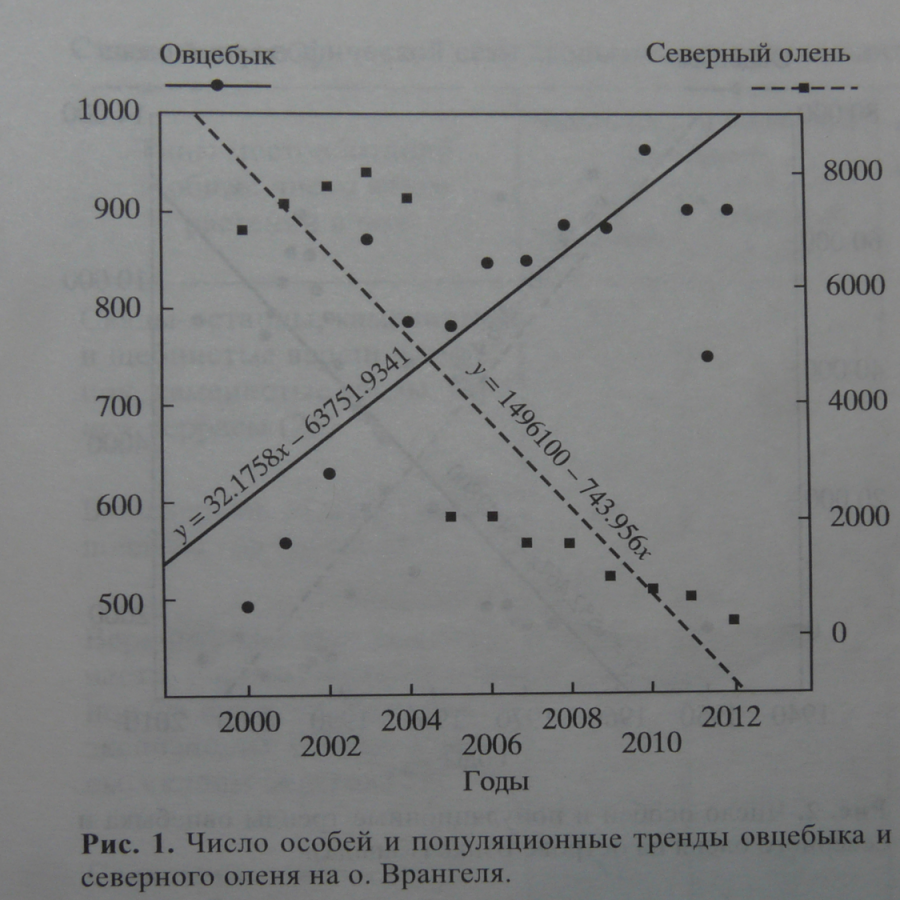
— на о.Врангеля. Помимо того, что здесь нет охоты, крупные хищники, значимые для наших копытных – волки и росомахи – заходили всего несколько раз. Т.е. отсутствие выборочного истребления людьми и крупными хищниками уравнивает условия, в которых находятся оба вида. После образования заповедника в 1975 г. популяция оленей была искусственно сокращена с 6 тыс. до 1.5 тыс.голов, дальше была стабильной, и начала сокращаться после выпусков сразу начавшего размножаться овцебыка в 2003 г.
С 2004 по 2007 г. популяция северных оленей самостоятельно сократилась до 0.45-0.5 тыс., и этот процесс продолжается. Численность же овцебыков подросла с 860 к концу 2003 г. до 960 к 2010 г., после чего рост замедлился. Т.е. самостоятельное снижение популяции северного оленя случилось тогда, когда численность овцебыков росла (рис.1).
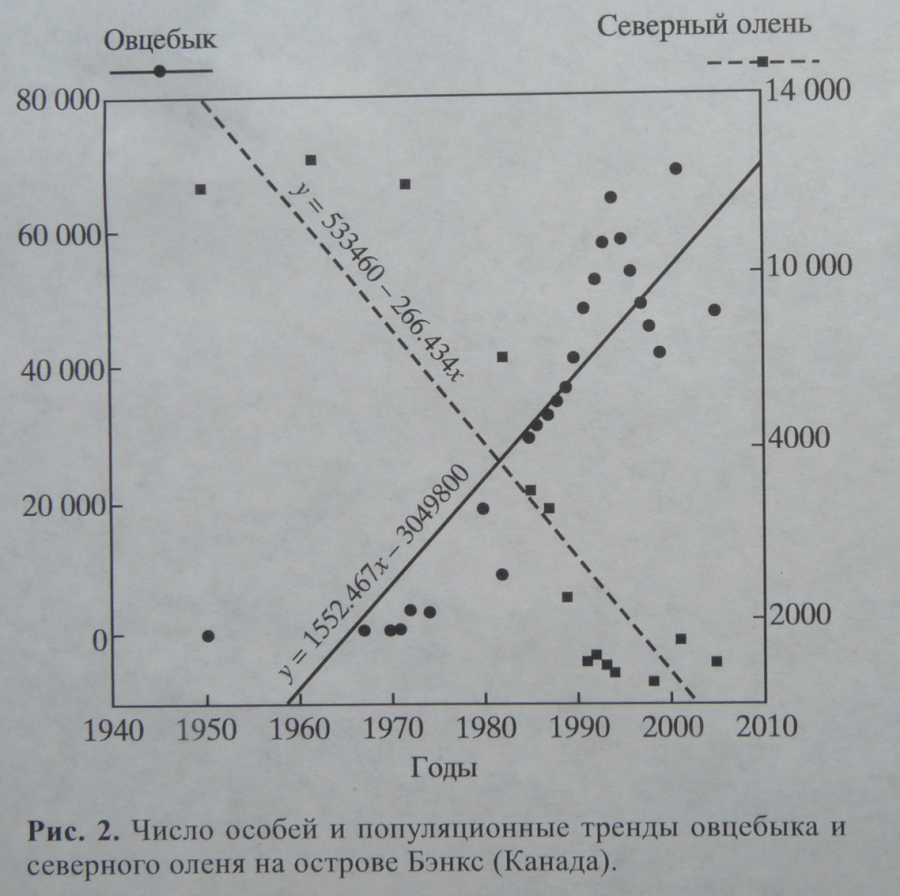
 - на о. Бэнкс в канадском секторе Арктики (730 с.ш., 1210 з.д.) данные виды сосуществуют с плейстоцена. Введение охранных мер вызвало увеличение численности овцебыка, после чего произошла депрессия численности северного оленя (рис.2).
- на о. Бэнкс в канадском секторе Арктики (730 с.ш., 1210 з.д.) данные виды сосуществуют с плейстоцена. Введение охранных мер вызвало увеличение численности овцебыка, после чего произошла депрессия численности северного оленя (рис.2).
 - на о.Нунивак (Аляска, 600 с.ш., 1660 з.д.) северные олени и овцебыки были завезены в 1930-х и 1940-х гг. соответственно. Более поздний рост популяции овцебыка здесь также совпадает с падением численности северного оленя (рис.3).
- на о.Нунивак (Аляска, 600 с.ш., 1660 з.д.) северные олени и овцебыки были завезены в 1930-х и 1940-х гг. соответственно. Более поздний рост популяции овцебыка здесь также совпадает с падением численности северного оленя (рис.3).
 И наоборот, отсутствие охраны (т.е. варварский перепромысел первобытных охотников) вызвало избирательное истребление овцебыка в Евразии и на большей части ареала в Америке – где северный олень в основном сохранился. А вот заповедный режим, т.е. отсутствие избирательного истребления обеспечил рост численности овцебыка с сокращением численности оленя; и мероприятия по реинтродукции овцебыка на территории, заселённые северным оленем, в основном успешны.
И наоборот, отсутствие охраны (т.е. варварский перепромысел первобытных охотников) вызвало избирательное истребление овцебыка в Евразии и на большей части ареала в Америке – где северный олень в основном сохранился. А вот заповедный режим, т.е. отсутствие избирательного истребления обеспечил рост численности овцебыка с сокращением численности оленя; и мероприятия по реинтродукции овцебыка на территории, заселённые северным оленем, в основном успешны.
То есть, можно считать, что овцебык в отсутствие охотников гарантированно выигрывает конкуренцию у оленя, то есть по численности преобладает вид травоядного с более широкой нишей и большей долей кормовых растений, используемых исключительно им. За счёт чего так получается?
Оба вида на о.Врангеля достаточно подвижны, что бы освоить пастбища всей территории острова, хотя олень сильно подвижней, вследствие его стратегии, противоположной овцебычьей (этот вид относительно оседлый, разница в подвижности на порядок – 120 км и 2000 км дальность перемещенй). Если коротко, типы питания травоядных образуют градиент, полюсами которого выступают неселективно стригущий и селективно выщипывающий типы питания (англ. grazing & browsing). Понятно, что разные виды на этом градиенте находятся где-то между полюсами, но ближе к одному из них. Северный олень – это скорее неселективно стригущий, овцебык – селективно выщипывающий, что задаёт различия в их составе диет.
Важный момент – основу питания обоих видов составляют злаки, осоки и ивы. Преимущественное питание ягелем для оленя на о.Врангеля не характерно, что соответствует условиям древних тундростепей, где растительные ассоциации были в основном злаковые, а не мохово-лишайниковые, как в современной тундре. Число используемых сообща видов растений возрастает параллельно росту их общего числа в спектре кормов, т.е. чем больше видов кормовых растений есть в данном местообитании, тем больший % из них оба вида потребляют совместно. Число кормовых растений, используемых исключительно первым и вторым видом, также возрастает с ростом их общего числа в данном местообитании, но у овцебыка много сильнее, чем у оленя (ранговые коэффициенты Спирмена = 0.922 и 0.779). Общее число кормовых растений у овцебыка больше, чем у оленя, в 1.31-2,15 раза в разных местообитаниях. Превосходство в числе кормовых растений, используемых исключительно данным видом, у овцебыка ещё выше – 2.33-8.5 раза и более. У оленя таких видов корма не более 5, в одном из местообитаний их нет вовсе, у овцебыка же – от 7 до 28 (рис.4 и статья данных авторов в «Зоологическом журнале»).
 Полученные данные однозначно характеризуют овцебыка как более мощного конкурента, что связано с большей специализацией кормовой стратегии, и лучшим противостоянием самому трудному периоды – гололёду, когда оба вида прилагают усилия по добыче растений из-под ледяной корки. Мы видим, что в условиях охраны по численности устойчиво доминирует более специализированный (=более мощный) конкурент с более широкой нишей и большей долей эксклюзивно используемых ресурсов. Больше того, наличие многочисленных популяций северного оленя отнюдь не препятствует реинтродукции более мощного конкурента.
Полученные данные однозначно характеризуют овцебыка как более мощного конкурента, что связано с большей специализацией кормовой стратегии, и лучшим противостоянием самому трудному периоды – гололёду, когда оба вида прилагают усилия по добыче растений из-под ледяной корки. Мы видим, что в условиях охраны по численности устойчиво доминирует более специализированный (=более мощный) конкурент с более широкой нишей и большей долей эксклюзивно используемых ресурсов. Больше того, наличие многочисленных популяций северного оленя отнюдь не препятствует реинтродукции более мощного конкурента.
Данные о стравливании стадами северного оленя пастищ на о.Врангеля в период до сокращения численности популяций и проявление результата соперничества в виде смены доминанта говорят о близости обоих популяций к некому пределу насыщения. Теоретически в этих случаях виды могли бы выйти из конкуренции, дифференцировав местообитания (и экониши) так, чтобы в одних из них было сильно больше число видов растений, используемых исключительно овцебыком, а в других, неперекрывающихся – оленем. Но нет, эта часть спектра питания везде больше у овцебыка, так что дифференцировать местообитания и уйти от конкуренции они не могут (если бы жили на материке, то видимо, дифференциации помогла бы разность в подвижности, но с острова куда уйдёшь?).
Если бы на островах было наоборот, то преобладание более слабого (менее специализированного) конкурента объяснимо погодными условиями, т.е. это довод в пользу климатической гипотезы. А так, вымирание более сильного конкурента – овцебыка с сохранением более слабого – северного оленя на большей части ареала – прямое и непосредственной доказательство перепромысла. Наконец, овцебык лучше приспособлен к климату Арктики, чем олень (крупные размеры, лучшая термоизолирующая способность меха, меньшее воздействие гололёда, кормовая стратегия и пр.). Кормовые местообитания овцебыка, имеющиеся на о.Врангеля, до сих пор занимают значительные территории на евразийском материке, а до временной границы плейстоцен-голоцен они были распространены ещё дальше к югу. Т.е. тогда популяции овцебыка были скорей всего жизнеспособней оленьих.
Возражением данному выводу может быть тот факт, что причиной неудач интродукции овцебыков на Шпицбергене, в Исландии и Швеции называли конкуренции с северным оленем. Однако более состоятельной кажется объяснение неудач воспалением лёгких, которым овцебыки легко заражаются во влажном климате в зоне влияния Гольфстрима. Во всех трёх случаях, кроме способствующего климата, есть источник заражения – человеческие поселения.
Иными словами, первобытный человек выступил в роли хищника, выборочно истреблявшего более крупную и исходно более многочисленную в Арктике жертву (одновременно ослабляя выжившую часть популяции, в силу специфики воздействия охотников, противоположной воздействию природных хищников).
——————————-
Внимательные читатели могут спросить – как же так, у птиц-печников межвидовая конкуренция не структурирует отношения (и дивергенцию) близких видов, а вот для этих двух видов – да, и видимо, в сообществах копытных эдификаторов травянистых биомов – тоже да. Есть ли тут некоторое общее правило? Увы, так бывает со всеми обобщениями в биологии, от «законов» до «правил» и «эмпирических зависимостей» (их интересная классификация дана в «Экологии» Розенберга, Мозгового и Гелашвили).
Скажем, в своё время Барт и Белл для разных видов млекопитающих пытались проверить гипотезы Чёрной королевы и конкуренции сибсов, используя оценки общей частоты рекомбинации. По первой, уровень рекомбинации должен быть тем больше, чем больше время смены поколений. Чем больше последнее, тем выше вероятность, что потомки будут жить в иных условиях, чем родители, соответственно, родительские комбинации аллелей не сохраняют адаптивные преимущества = надо рекомбинировать. Если время смены поколений невелико – то наоборот, потомки живут в том же мире, с теми же критериями приспособленности. По второй гипотезе, уровень рекомбинации пропорционален числу потомков. Типа чем больше детёнышей в помёте, тем больше они должны различаться, чтобы не конкурировать друг с другом = надо рекомбинировать.
В качестве показателей уровня рекомбинации использовали превышение числа хиазм над гаплоидным числом хромосом (Burt, Bell, 1987). Оно оказалось высоко – и положительно связано с временем смены поколений, при отсутствии связи с величиной помёта. Т.е. гипотеза Чёрной королевы подтверждается, а конкуренции сибсов – отвергается.
Однако П.М.Бородин с соавторами показывает, что эти оценки частоты рекомбинации неточны. Сейчас разработаны более точные — цитологические методы, основанные на использовании меченых флуорохромами антител к SCP3, белку боковых элементов синаптонемного комплекса и MLH1, эукариотическому гомологу бактериального белка мисматч репарации. Было показано, что MLH1 входит в состав зрелых рекомбинационных узелков. Число и распределение точек локализации MLH1 в пахитенных клетках точно соответствует числу и распределению хиазм в диакинезе мейоза. Установлено, что частота рекомбинации между двумя сцепленными генами, оцененная на основе частоты точек локализации MLH1 между FISH-пробами к этим генам, совпадает с оценкой частоты кроссинговера между ними, полученной в генетическом эксперименте.
И вот с использованием иммунофлуоресцентного метода детекции сайтов рекомбинации авторы получили точные и несмещенные оценки рекомбинационных характеристик геномов на репрезентативных выборках клеток у 21 видов млекопитающих, сравнив их с данными еще по 15 видам. 40% из них составляли полёвки п/сем Arvicolinae, которых анализировали отдельно.
И вот на выборке из 15 видов полёвок не подтвердились ни гипотеза Чёрной королевы, ни конкуренции сибсов – уровень рекомбинации не коррелировал ни с временем смены поколений, ни с числом потомков. Число обменов на клетку практически совпадало с гаплоидным числом хромосом, то есть было минимально необходимым для их расхождения. Поскольку эти виды характеризуются быстрой сменой поколений, отбор был, по-видимому, направлен на консервацию адаптивных аллельных комбинаций и уменьшение комбинативной изменчивости. Это было достигнуто как за счёт увеличения кроссверной интерференции[1], так и за счёт накопления перестроек, уменьшающих число хромосом — Робертсоновских и тандемных слияний.
А вот у всех прочих видов, включённых в метаанализ, гипотеза Чёрной королевы подтверждается (с отвержением конкуренции сибсов, как у Барта и Белла). Время смены поколений сильно и высоко значимо скоррелировано с превышением числа кроссоверов над гаплоидным числом хромосом, при незначимости связи с размером помёта. По-видимому, при увеличении времени смены поколений отбор приводил к повышению уровня рекомбинации. При этом само повышение уровня рекомбинации могло достигаться за счёт снижения интерференции (с.24-25).
Иными словами, одно и то же общее правило – Чёрной королевы – работает для многих видов млекопитающих, и не работает для полёвок. А почему так – определяется «конструкцией» кариотипа тех и других, особенности которой определяют разные способы решения одной и той же эволюционной проблемы. Т.е. «конструкция» эволюционирующих таксонов, от морфологии особей до пространственно-этологической структуры видовых популяций, и присущие ей «конструктивные противоречия» определяют, выполняются ли для них общие правила, или нет, или до какой степени, если да.
Источник wolf_kitses
Рекомендуем прочесть
 Некоторые пути сохранения биосферы при урбанизации
Некоторые пути сохранения биосферы при урбанизации Оптимизация рациона пойдет на пользу здоровью людей и планеты
Оптимизация рациона пойдет на пользу здоровью людей и планеты Стадии деградации дикой природы под антропогенным воздействием
Стадии деградации дикой природы под антропогенным воздействием Исчерпание ресурсов и истощение почв
Исчерпание ресурсов и истощение почв Предисловие книги о глобальном экологическом кризисе
Предисловие книги о глобальном экологическом кризисе История охраны природы, 1945-ныне
История охраны природы, 1945-ныне Из истории охраны и разграбления природных богатств в России
Из истории охраны и разграбления природных богатств в России Рыба наперечёт
Рыба наперечёт
