
Схема рисунка полости рта у птенцов: (а) доминиканской вдовушки (Vidua macroura) и астрильдовых: африканских b – Estrilda astrild и с - Amandara subflava и d – австралийской Stizoptera bichenovii. Источник Hauber, Kilner, 2007
Резюме. Врождённые системы реагирования, реализующие инстинкт, всегда содержат элемент распознавания образов: отсюда потенциальная «сигнальность» инстинктов. В обзоре представлены материалы, обосновывающие данный вывод, и показывающие, что коммуникативное функционирование релизеров двойственно и противоречиво: хороший стимул — плохой сигнал и наоборот. Хотя релизеры «зовут» знаковыми стимулами, прилагательное здесь намного важней существительного. Стимул исходно — заострённая палка, непосредственно (уколом, тычком или толчком) «принуждающая» к определённым ответам, сигнал обеспечивает определённый ответ информацией, считанной с носителя сигнала, т.е. «без принуждения», в условиях свободы выбора. В единицах поведения и единицах коммуникации птиц и других позвоночных — ритуализированных демонстрациях сигнал, информирующий реципиента, оставляющий ему свободу выбора поведения, здесь амальгамирован со стимулом, «принудившим» его же к определённым ответам. Поскольку второе реализуется лишь при условии первого, обратное же неверно, функциональные исследования неизменно показывают, что они антагонистичны, а не синергичны.
***
Так, при ухаживании самка зеленоватого тритона Notophtalmus viredescens демонстрирует брюхо, заполненное яйцами, самец ей — внутренние поверхности задних конечностей с роговыми подушечками, небольшие ямки на скулах и хвост с увеличенной высокой складкой. Реакция самки на его демонстрацию даёт два сценария поведения самца: когда самка остановится рядом, показ быстро закончится и самец перейдёт к следующей фазе ухаживания, когда удаляется, самец следует за ней, стараясь обхватить передними лапками (рис.1), и интенсивно демонстрирует. При низкой реактивности самки продолжительность демонстраций примерно в 25 раз выше: в конце концов, движение самки прочь замедляется, самцу удаётся её нагнать и схватить[1].
Увеличивая интенсивность своей стимуляции, самец «поднимает» реактивность самки до уровня, «переводящего» её на следующий этап ухаживания – сближение с демонстрирующими самцами, чтобы выбрать одного и взаимодействовать дальше именно с ним вплоть до передачи сперматофора (Verrel, 1987). Интенсивность воздействия при таком «обращении внимания» самки на самцов пока что важней точности воспроизведения форм демонстраций. Не случайно она ведёт к характерным «сбоям» — привлечению незрелых самок, самок близкого вида и т.п. «посторонних объектов», негодных для ухаживания.
Когда же под действием соответствующей стимуляции самка уже мотивирована к сближению с самцами и к оценке их демонстраций «с точки зрения» последующей копуляции, точность воспроизведения их специфических форм делается решающей, а интенсивность стимуляции не так важна, см. далее про ухаживание девятииглых колюшек рода Pungitius.

Рисунок 1. Ухаживание у зеленоватых тритонов. Самец обхватывает самку и демонстрирует перед ней (сверху); другие могут присоединиться и также демонстрировать, или попробовать вытеснить данного самца. Фотограф К.Грейсон, Вики.
Так или иначе, релизер действует своей специфической формой: даже когда «принуждающее» воздействие присутствует, оно реализуется лишь при условии, что а) предъявление демонстрации-релизера достаточно стереотипно, т. е. образующие её телодвижения так «чертят» образ, воспринимаемый реципиентом, что он инвариантен к изменениям контекста, б) этот образ устойчиво воспринят реципиентом вопреки разнообразным «помехам», внешним и внутренним.
Так, исследовали индикаторы эмоционального возбуждения в структуре оборонительных криков большой песчанки Rhombomys opimus, и их действие на агрессора. Конфликт провоцировали, ссаживая зверьков на нейтральной территории. После короткого периода борьбы, в разной степени ритуализованной, один (победитель) преследовал второго, показывавшего подчинение и издававшего оборонительные крики. Сокращение дистанции между ними меняло эмоциональное состояние обоих, что сразу отображалось в пропорциональных ему изменениях вокализаций: падала длительность оборонительных звуков, их энергия смещалась в область высоких частот, выросла основная частота и глубина частотной модуляции тональных звуков.
Данные изменения в криках испуганной, обороняющейся песчанки прямо «отталкивали» наступающую, увеличивая дистанцию между ними сразу и резко. Важно, что крики больших песчанок, обладающие более специализированной акустической формой, эффективней ингибировали агрессию по сравнению с менее определёнными криками светлой Gerbillus perpallidus[2] (Володин и др., 1998; Доронина, Володин, 2007).
Третий пример двойственного действия релизеров — как сигналов и специфических стимулов, где второе реализуемо лишь при условии первого, но не наоборот, даёт булькающая вокализация черноголовых гаичек Parus atricapillus (рис.2А). С одной стороны, бульканье — внешнее выражение агрессивности, т. е. способности птицы силой «очистить» кормушку, заставить товарок отступить и ждать своей очереди. С другой, его акустическая форма — предиктор большей или меньшей способности изгонять других особей, в «сквозной» оценке которой «заинтересованы» все совместно кочующие члены зимних стай, а также бродячие гаички (Baker et al., 1991).
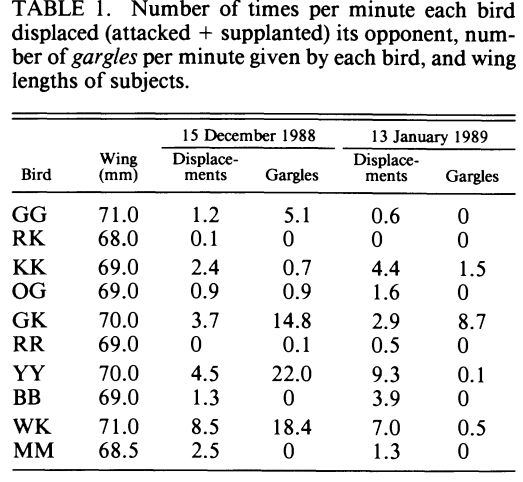
Анализ показывает, что бульканье связано со стремлением птиц доминировать в конкурентной ситуации, скажем, согнать другую или брать корм, не обращая ни на кого внимания. В конфликтах между самцами гаичек всегда побеждает булькающий (рис.2Б). При ссаживании незнакомых птиц на нейтральной территории доминирование устанавливается посредством борьбы. В её начале гаички обмениваются булькающими сигналами, чередуя их с открытой агрессией по отношению к оппоненту (Baker et al., 1991). «Бульканье» здесь — достоверный маркер агрессивности, почему выступает сигналом состояния, предиктором большего или меньшего «объёма агрессии», который животное может «обрушить» для вытеснения или подчинения данной особи.
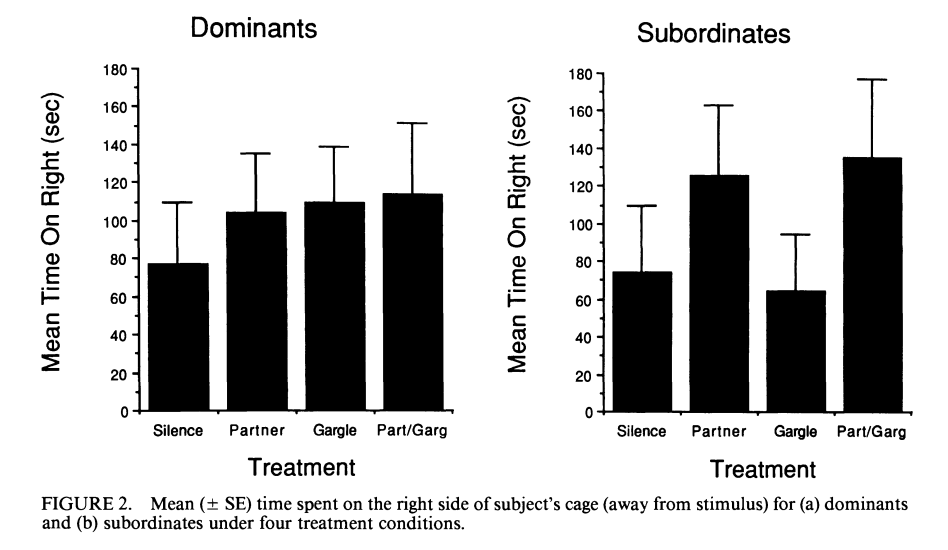
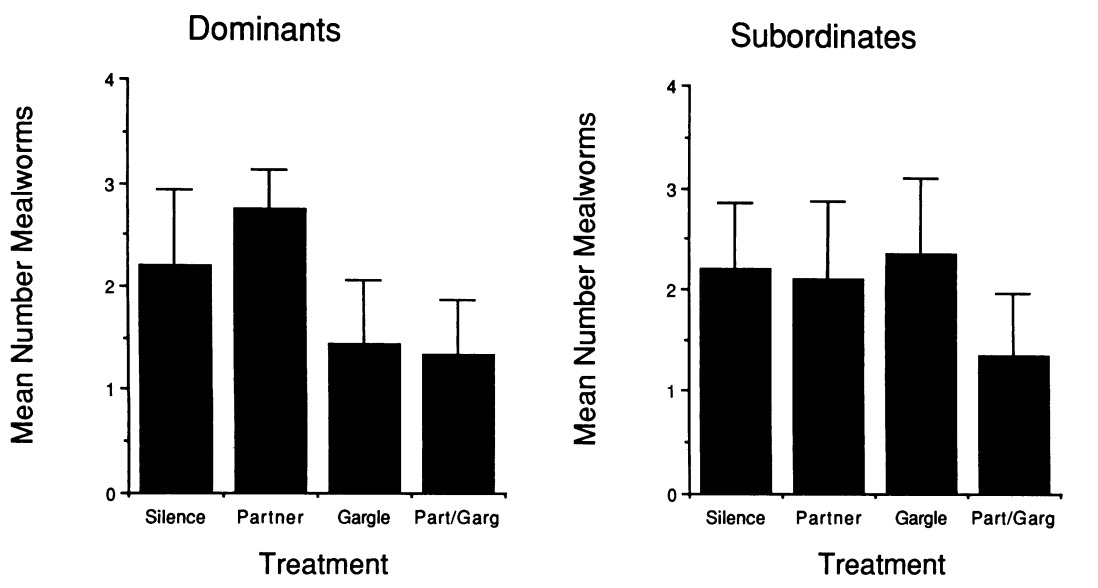
В какой-то момент одна гаичка прекращает сопротивление, а вторая её изгоняет одним бульканьем, без открытой агрессии: здесь появляется доминирование. В устойчивых зимних стаях синиц бульканье — знак присутствия доминанта[3]: «доверие» к такому сигналу сравнимо с реакцией на само появление другой особи и её агрессию, сигнал даже поэффективней (рис.2В). Проигрывание «бульканий» подчинённого препятствовало кормлению доминанта, а его предъявление никак не влияло. Подчинённый, наоборот, не реагировал на «бульканья» доминанта, но освобождал кормушку, увидев его вживую (Baker et al., 1991).
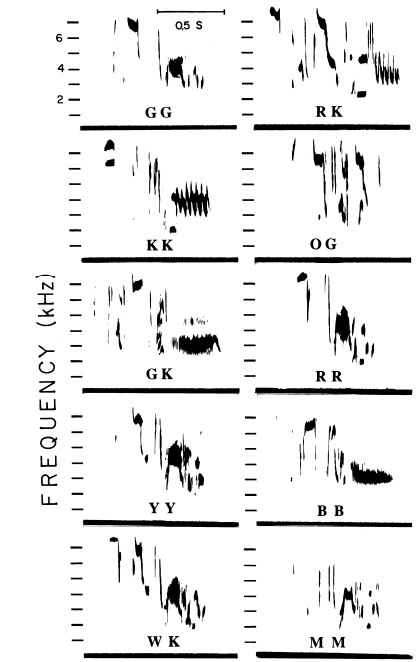
Рисунок 2. Булькающая вокализация как сигнал состояния гаичек. А. «Бульканья» разных птиц, дальше использованные как стимулы в опытах с двумя клетками. Б. Связь агрессии с бульканьем. В таблице — время, в течение которого каждая синица была атакована и вытеснена, за 2 выбранных дня, 15.12.1988 и 13.01.1989. Для каждой особи (строки) показаны длина крыла, число бульканий и вытеснений/мин. В. Данные опытов, показывающих влияние бульканий и/или непосредственного присутствия партнёра на посещение кормушки. Птица находится в одной части клетки, в другой — закрытое блюдечко с мучными червями. Правую сторону или озвучивают бульканьем, или в соседней клетке вживую присутствует синица-противник, или то и другое вместе, контроль – ничего не случается. В определённый момент крышку снимают на 280 с., после чего подсчитывают время, проведённое птицами в той половине, что дальше от корма и проигрываемых звуков (В1, с), а также число съеденных хрущаков (В2). Столбцы В1-В2 слева направо: контроль, партнёр, бульканья, то и другое вместе, слева доминанты, справа подчинённые. Из: Baker et al., 1991.
б.
в1.
в2.
Видно, что «бульканье» имеет самостоятельное значение «прогноза» вероятности эффективного подавления сочленов по группе, отдельное от маркёра агрессивности особи (на основе которого, видимо, развился этот сигнал). Дальше, все индивиды в стае гаичек и «зрители», стремящиеся туда включиться[4], вполне «доверяют» информации идеального характера, передаваемой сигналами. «Доверие» проявляется в том, что обе особи, донор и реципиент, выбирают поведение на следующем «шаге» взаимодействия, исходя именно из неё, часто в противоположность «давлению» собственных агрессивных побуждений и «противодавлению» агрессии оппонента.
На этом примере видим, что действие релизеров двойственно и противоречиво. Сигнал, информирующий реципиента, оставляющий ему свободу выбора поведения, здесь амальгамирован со стимулом, «принудившим» его же к определённым ответам. Поскольку второе реализуется лишь при условии первого, обратное же неверно, функциональные исследования неизменно показывают, что они антагонистичны, а не синергичны, хороший стимул — плохой сигнал, и наоборот.
Так, «зигзагообразный танец» самцов малой южной колюшки Pungitius platygaster[5] вполне эффективен как знаковый стимул. Несмотря на снижение демонстративности и заметности по сравнению с гомологичным сигналом девятииглой колюшки P.pungitius, он хорошо привлекает и самок близкого вида, и не готовых к нересту самок своего. В опытах по скрещиванию самцов P.platygaster с самками P.pungitius 3 из 8 предложенных самок были незрелы (отказывались нереститься с самцами своей формы), но самцам малой южной колюшки удалось не только подвести их в гнездо, но побудить проникнуть в него несколько раз подряд. При нересте P.pungitius подобное редкость, не чаще 1 раза на 50 нерестов (Гомелюк, 1979).

Близкие виды, малая южная и девятииглая колюшки.

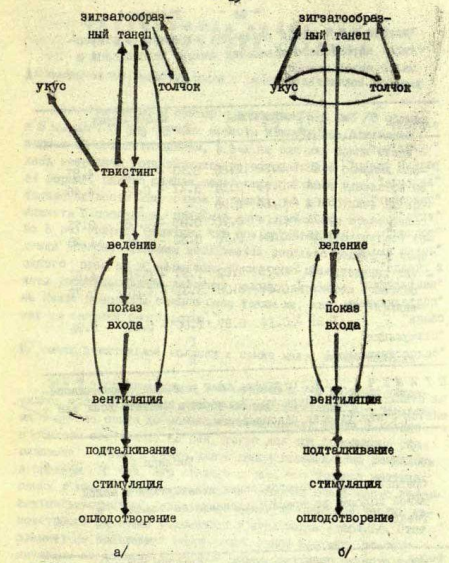
Рисунок 3. Ухаживание девятииглой колюшки. А. Демонстрации партнёров, последовательно реализующие инстинкт: а) зигзагообразный танец; б) твистинг, в) ведение самки, г) показ входа в гнездо, д) вентиляция гнезда, ж) подталкивание входящей самки, з) стимуляция вымета самкой икры, е) оплодотворение. Б. При идентичности демонстраций а)-е) P.pungitius и P.platygaster различаются структурой последовательностей: отказ самки следовать к гнезду побуждал самца её кусать и толкать, затем он повторяет зигзагообразный танец у первого вида и твистинг у второго.
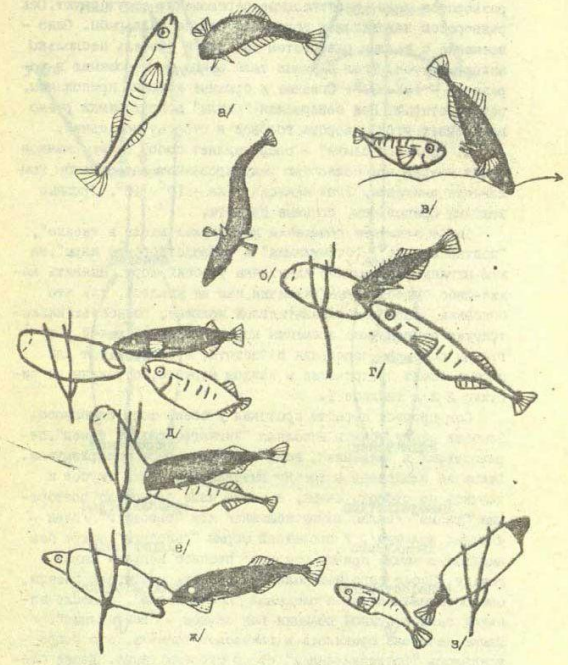
Не менее эффективен «зигзагообразный танец» девятииглой колюшки: резкие скачки угольно-чёрной, с синевато-белыми колючками рыбы привлекают даже не вполне готовых к нересту самок, зрелые плывут с дистанции до 2 м. Однако сильные стимулы — плохие сигналы. В смешанных парах с самками малой южной колюшки возникает «сбой» с нарушением согласованности действий партнёров. «Ведение» конспецифичной самки к гнезду у P.pungitius часто прерывается «твистингом» (рис.3): быстрыми S-образными изгибаниями тела на месте то вправо, то влево, иногда с разворотом вдоль длинной оси тела на угол от 150 до 300. Демонстрация видоспецифична, она одна из самых частых в ухаживании.
«Твистинг» не привлекает самок P.platygaster: при его исполнении они останавливались, едва начав следовать после зигзагообразного танца. Продолжить движение самок побуждает множественное повторение «танца» или «ведения», но самцы P.pungitius склонны повторять «твистинг» снова и снова. Среагируй малые южные колюшки на «твистинг» как на сигнал «сбоя коммуникации», ошибка б исправилась, и взаимодействие продолжилось дальше: но они ждут адекватной стимуляции и не находят её. Возникший «сбой» оценим количественно, потерями времени и энергии из-за возникшей рассогласованности или величиной «упущенной прибыли» (шансом привлечь лучшего самца и отнереститься с ним). Колюшки, особенно малая южная, живут при высокой угрозе нападения хищных рыб и птиц, любое затягивание процесса увеличит риск гибели. А нерест смешанной пары сильно затягивается, от начала опыта до проникновения самки в гнездо в среднем проходит 68 с, в гомогамных спариваниях – 10 с.
При «сбоях» ухаживания в конспецифической паре твистинг повторяется снова и снова, пока самка не приблизится к самцу и не последует за ним или, наоборот, не уплывёт прочь. Его исполнение — фильтр, отделяющий зрелых самок от неготовых к нересту. Подчеркну: те и другие равно чувствительны к стимуляции специфическими демонстрациями самца, но лишь зрелые воспринимают их как сигналы-предикторы программы действий на следующем этапе процесса и поэтому координирующие поведение партнёров. Действительно, сравнение гомологичных демонстраций обоих видов показывает, что у P.platygaster усилено действие их как сигналов, но ослаблено как релизеров, чему способствует больший пресс хищников (Гомелюк, 1979).
Не менее ярко противоречие между сигнализацией и воздействием видно в релизерах кукушат, побуждающих приёмных родителей их кормить, вопреки большим размерам, непохожести на своих птенцов и на их гибель из-за выбрасывания. Правда, выбрасывание птенцов хозяина имеет свою цену: оставшись один, кукушонок стимулирует кормиться явно слабее, чем несколько птенцов в выводке своими яркими зевами[6], приёмные родители носят ему корм хуже, чем полному выводку.
Проблема решается большей стимуляцией, более продолжительными/частыми криками, более ярким зевом у кукушат и пр., либо большим совпадением их сигналов с таковыми птенцов вида-хозяина, но не тем и другим вместе. Так, птенцы обыкновенной кукушки Cuculus canorus выпрашивают при помощи особенно интенсивно издаваемого крика, чем компенсируют дефицит визуальной стимуляции (хотя его зев и ярок, но меньше по площади, чем зевы 4-5 птенцов вида-хозяина).

Рисунок 4. Длиннохвостый коэль (сверху) и бронзовая кукушка (снизу).

Скажем, пищевые сигналы птенцов двух видов кукушек — длиннохвостого коэля Eudynamys taitensis из Новой Зеландии и австиралийско-океанической бронзовой Chrysococcyx lucidus (рис.4) крайне сходны с таковыми птенцов хозяев: почти одинакова основная и максимальная частоты позывок, ритмическая организация лишь чуть-чуть отличается. Птенцы и кукушек, и видов-хозяев равно интенсифицировали вокализацию при прилёте родителей, но сигналы кукушат значимо длиннее, что дополнительно побуждает кормить (Нумеров, 2003).
У ширококрылой кукушки Cuculus fugax отношение веса птенца паразита к суммарному весу птенцов хозяев примерно такое же, что у обыкновенной, т. е. её птенцу тоже нужно усилить выпрашивание, но здесь эта задача решена необычно (Tanaka, Ueda, 2005). На нижней стороне крыльев птенца развиваются ложные зевы — пятна образом окрашенной кожи, демонстрируемые при выпрашивании вместе с реальным зевом для дополнительной стимуляции родителей, благодаря чему те несут больше корма. См. рис.5А: слёток ширококрылой кукушки разевает рот, открывает крылья и трепещет ими, так что «ложные зевы» на них предстают в увеличенными именно в моменты кормления приёмным родителем (синехвосткой Tarsiger cyanurus).

Рисунок 5А. Кукушонок демонстрирует «ложные зевы»
Авторы проверили эту гипотезу экспериментально. Если птенец так стимулирует принос корма, интенсивность демонстрации «ложных зевов» должна зависеть от остроты голода. Действительно, чем голоднее кукушата, тем быстрей учащались демонстрации крыловых пятен, по разным траекториям у разных птиц, но с выходом на одно и то же плато, которое, видимо, обеспечивает должный эффект (мера пищевой мотивации — интервал между приносами корма, рис.5Б).
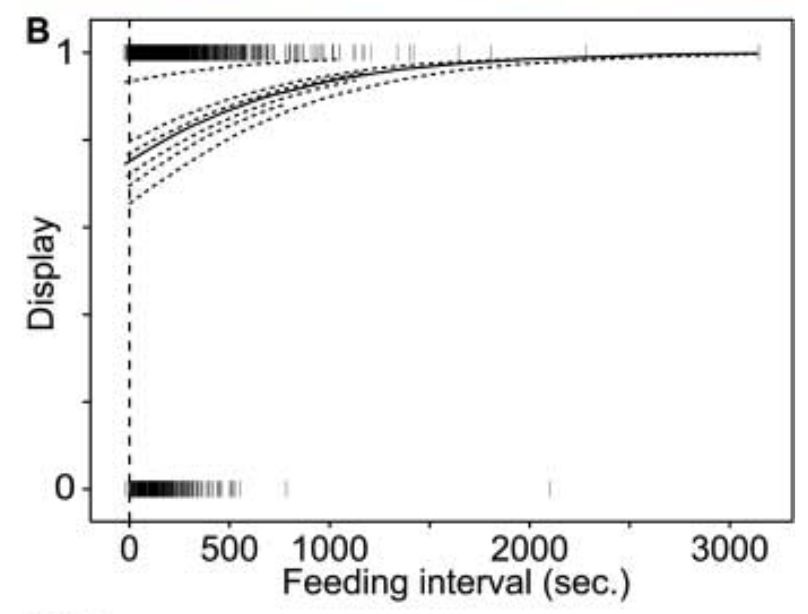
Рисунок 5Б. Зависимость демонстрации «ложных зевов» от интервалов между кормлениями, с.
Авторы предположили, что кукушата демонстрируют пятна не просто так, а чтобы их кормили больше и чаще; для проверки поставили опыт с окрашиванием в 3-х вариантах:
1) контроль, ничего с птенцами не делали,
2) контроль 2: имитировали закрашивание пятен прозрачным раствором,
3) реально закрасили крыловые пятна.
Дальше анализировали различия в интенсивности приносов корма птенцам (раз в ч) во всех 3-х группах (рис.5В): она значимо снижена в 3), но не в 1)-2), хотя там птенцов брали в руки, тревожили и пр.
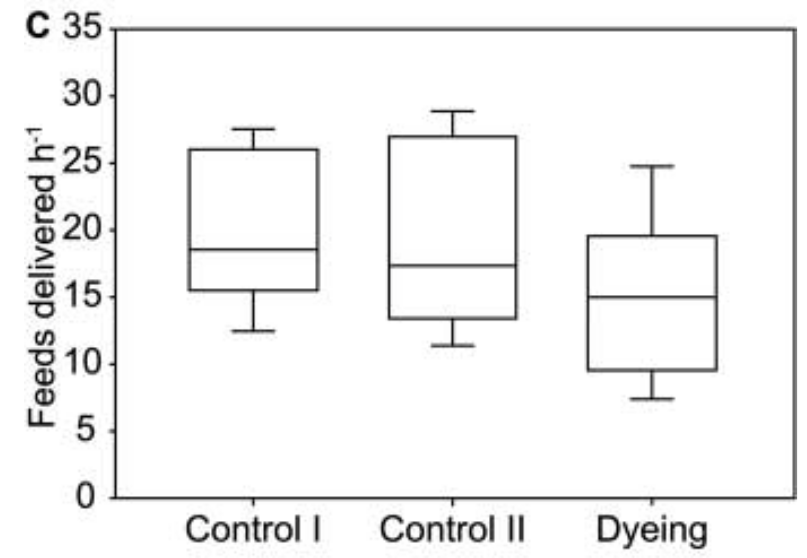
Рисунок 5В. Частота кормлений кукушат с закрашенным «ложным зевом» сравнительно с контрольными группами.
Прямые наблюдения также показывают, что приёмные родители время от времени сбиваются и пытаются сунуть корм в пятна на крыльях (Tanaka et al., 2005, рис.2 и табл.1 работы). Поэтому ложный зев действительно создаёт сверхстимулы, «подстёгивающие» приёмных родителей. Хотя крыловые пятна не точно повторяют рисунок настоящего зева (точнее совсем не повторяют, рис.5А), это неважно: внутренность гнёзд всех трёх видов-хозяев ширококрылой кукушки (синехвостки Tarsiger cyanurus, синей мухоловки Cyanoptila cyanomelana и синего соловья Luscinia cyane) достаточно тёмная. Низкая видимость не позволяет родителям отличить настоящий зев от «ложных зевов» на крыльях и движет почаще кормить единственного птенца, как будто их несколько.
Таким образом, птенцы обыкновенной кукушки эксплуатируют хозяев вокальной «обманкой», ширококрылой же — визуальной, видимо, потому что куда уязвимей для хищников. Все их хозяева гнездятся на земле, и хищные млекопитающие разоряют ~ 50% гнёзд в районе наблюдений. Поэтому их птенцы и птенцы паразита издают пищевые крики намного реже, и кричат тише; ложные зевы вообще позволяют эксплуатировать приёмных родителей молча (Tanaka, Ueda, 2005).
На этом основании авторы предлагают новую классификацию сигналов, используемых птенцами гнездовых паразитов для обмана хозяев. Они предлагают различать сигнальную и сенсорную эксплуатацию. Первая паразитирует на сигналах, реально используемых птенцами вида-хозяина для общения с родителем, они могут быть воспроизведены точно (мимикрия) или усилены, превратившись в сверхстимул. Вторая использует сигнал, несвойственный виду-хозяину, но так и в таком контексте, что тот гарантированно путается.
Данные К.Д.Танака и К.Уэда поддерживают «еретическую» точку зрения на окраску зевов птенцов у других гнездовых паразитов, райских вдовушек cем. Viduidae. Как известно, у каждого вида вдовушек зев птенцов точно соответствует раскраске зева птенцов соответствующего вида хозяев, освоение новых видов-хозяев влечёт за собой акты видообразования, с расщеплением прежних видов. Птенцы, попадающие в чужое гнездо, остаются без корма и гибнут от голода. Традиционно такое сходство интерпретировали как необходимое для распознавания своего вида и несмешивания с чужими, но недавно возникла более убедительная точка зрения, что такое точное совпадение нужно для лучшей стимуляции приёмных родителей, чтоб почаще кормили.
Примечания
[1]Verrell P.A. The complex courtship of the red-spotted newt// Bull. Chicago Herpetol. Soc. 1987. V.22. №6-7. P.119-120
[2]См. Доронина Л.О., Володин И.А. Индикаторы эмоционального возбуждения в структуре оборонительных криков большой песчанки (Rhombomys opimus)// IV Всероссийская конф. по поведению животных. Тезисы докл. М.: Товарищество научных изданий КМК, 2007. C.165-166.
[3]И даже символ, поскольку его означаемое (уровень агрессивности) невыводимо из означающего (акустической формы бульканий, представленной на сонаграммах) для наивного наблюдателя, не включённого в эту систему коммуникации: сперва он должен расшифровать данный сигнал.
[4]скажем, бродячие нерезидентные особи (англ.floaters), присутствующие в популяции гаичек наравне с иерархическими группами, занимающими определённые территории в течение всей осени и зимы. Группы образованы парами, гнездившимися на данном участке, и подчинёнными им молодыми особями, присоединившимися во время позднелетних перемещений, иногда также нерезидентов, могущих присоединиться на протяжении осени и зимы.
[5]Дальнейший рассказ и иллюстрации по: Гомелюк В.Е. Дивергенция полового поведения на примере двух представителей рода Pungitius Coste// Динамика популяций и поведение позвоночных животных Латвийской ССР. Рига: Латвийский Госуниверситет им. П.Стучки, 1979. С.6-25.
[6] Одно из проявлений закона гетерогенной суммации Зайца (нем. Reizsummenregel, англ. law of heterogeneous summation, А.Seitz, 1940, 1941): общий эффект комбинации из качественно разных ключевых стимулов, на которые реагирует один и тот же врожденный разрешающий механизм, является суммой эффектов каждого из этих стимулов, действующих по отдельности. Здесь он выполняется, ибо зевы птенцов «работают» только как стимул, а не сигнал, где закон нарушается: значащие единицы (инвариант формы соответствующих демонстраций) составляются из незначащих элементов (факт 21).
Рекомендуем прочесть
 Как птицы обучаются инстинктивному поведению
Как птицы обучаются инстинктивному поведению Про танец как изоморфизм инстинктивной коммуникации
Про танец как изоморфизм инстинктивной коммуникации Ещё раз об инстинктах
Ещё раз об инстинктах Инстинктивное vs выученное/биологическое vs социальное
Инстинктивное vs выученное/биологическое vs социальное Беседа об инстинктах
Беседа об инстинктах Про «фантомный инстинкт»
Про «фантомный инстинкт» Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных «Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования
«Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования
