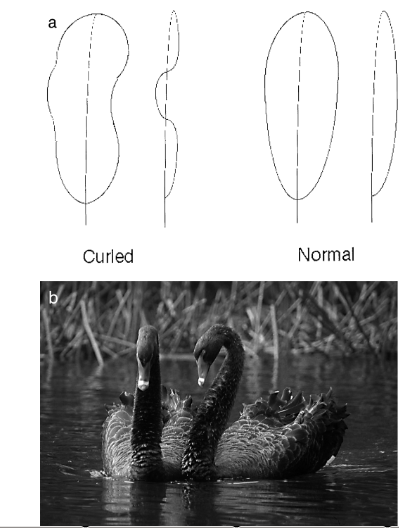
Рисунок 1. Кудрявые перья чёрного лебедя (слева) в сравнении с нормальными, вид сбоку и спереди
Что мне не нравится в социобиологии — она превращает исследования поведения в упражнение по части счастливых догадок. На что б ни упал взор социобиолога — внебрачные копуляции у моногамных видов, систему расселения молодняка, помощничество у птиц и млекопитающих, обмен демонстрациями при общении — везде сразу оказывается, что это чем-то адаптивно и выгодно, да ещё не для особей, худо-бедно способных к саморегуляции и прочим “ответам” на “вызовы” изменений среды, а для «эгоистических генов», которым на окружение наплевать.
А ведь сложную систему нельзя оптимизировать более чем по одному параметру.
Именно его изменения адаптивны, а прочие представляют собой либо подстройки по корреляциям, чтобы организм выжил, несмотря на изменения какого-то одного (ключевого) параметра отбором, или, ещё хуже, просто дестабилизацию и дезорганизацию вследствие слишком быстрого или жёсткого отбора, которой приписывать адаптивность можно лишь через анализ тенденциозно собранных данных.
И какие же шансы, что взгляд исследователя, в начале работы заведомо не понимающего «устройство» социальной жизни данного вида, упадёт именно на её ключевые моменты, дающие адаптивную выгоду? Ничтожно малые, тем более что в социобиологических публикациях отрицательный результат (мол, мы анализировали эту особенность поведения — но нет, не адаптивно, просто последствие чего-то другого или дезорганизация) вовсе отсутствует. Поэтому не удивительно что данные в пользу самых разных социобиологических построений, от родственного отбора до «неверные самки ищут лучшие гены», сплошь и рядом оказываются или некачественными, или обусловленными контекстом, а не самим соответствующим феноменом.
Панадаптационизм (иронично названный в своё время Ричардом Левонтиным “парадигма Панглоссса”) — он до добра не доводит (см.1-2-3), особенно в ситуации, когда генетиками надёжно показано, что отбору подвергаются фенотипы, а не отдельные гены. Знаменитые опыты М.М.Камшилова показывают, что даже когда последнее имеет место быть, результат определяется тем, в составе какой генной сети отбираемый ген функционирует в данных условиях среды, к которым идёт приспособление. Поэтому идея «эгоистичного гена» в ряде случаев (достаточно ограниченных) может быть полезной фикцией, но не онтологией и тем более не универсальным объяснением.
Ещё хуже, что простые догадки у социобиологов обретают “научность”, т.е. воспринимаются как выводы из высокой теории, почему исследователи настаивают на них, даже когда они опровергнуты. И даже в случаях, когда они опровергли их собственноручно, как с возможной значимостью украшающих перьев чёрных лебедей в половом отборе. Kraaijeveld et. (2014) проверили и отвергли эту гипотезу, показав, что число кудрявых перьев поддерживается социальным отбором, нло казуистически продолжали настаивать на половом. Что дальше дезориентирует авторов обзорных статей, если они склонны лишь переписывать основные утверждения из цитируемых работ, не вникая в характер полученных данных (как это сделано, например, в монографиях Е.Н.Панова, в отношении статей, к которым он настроен критически). Увы, то что в них пишется, часто бывает не выводом из данных или их обобщением, но декларации на верность школе или парадигме, вроде социобиологической в статьях, обсуждаемых ниже (асимметричной теории полового отбора, соответствующей наблюдениям, прямо скажем, не очень).
У чёрного лебедя Cygnus atratus по ходу полового созревания (в двухлетнем возрасте) на крыльях вырастают специальные «украшения» — кудрявые перья, которые придают этому виду красиво-романтический облик. Перья есть у самцов и самок, их 7-22 на крыло, их число увеличивается в процессе созревания.
Демонстрация кудрявых перьев происходит во время специфического «вращения», выступающего частью территориальной агрессии пары при охране соответствующего пространства от чужаков (подобное «вращение» есть у родственного лебедя-шипуна Cygnus olor). У обоих видов паре лучше иметь территорию, чем нет: в территориальных парах сравнительно с бродячими значимо больше кудрявых перьев, тогда как отличия по размерам и массе незначимы.
Основное различие бюджетов времени партнёров в периоды до размножения (т.е. ухаживания в образовавшейся паре) и вождения выводка — самцы больше времени проводят в позе угрозы, т.е. демонстрируют эти перья. В период до размножения самцы также участвуют в большем числе конфликтов, хотя общая частота последних не различается между периодами.
Анализ 153 конфликтов показывает, что самцы с большей вероятностью выиграют их, чем самки (χ24 = 55.17, p = .0001), птицы в парах вероятнее, чем непарные (χ24 = 44.74, p = .0001); с большим количеством кудрявых перьев вероятнее, чем с меньшим (χ24 = 15.32, p = .0004; везде рассчитано по данным табл.3). Однако при совместном рассмотрении всех 3 переменных контекста влияние каждой незначимо (везде Р>0.3). Из 40 конфликтах птиц одного пола и брачного статуса 70% выиграли особи с большим количеством кудрявых перьев (рис.5).
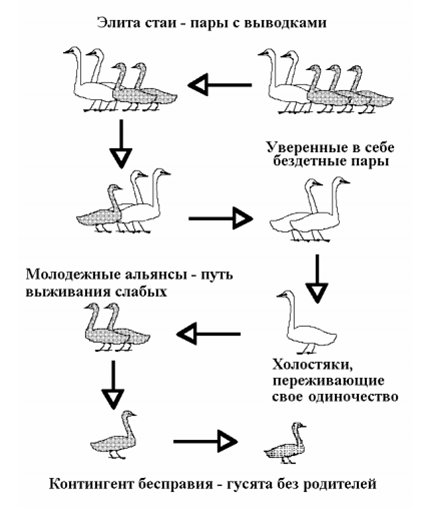
Векторы доминирования в многосемейных стаях гусеобразных. Из: Иваницкий, 2002
Поэтому у самцов эти украшения развиты лучше (самец у лебедей, гусей и полугусей несёт большую нагрузку по охране участка и вождению выводка), более украшенные особи могут отстоять и удерживать большую территорию, то есть обеспечить птенцам лучшие кормовые условия и достичь большей выживаемости птенцов. Территориальные пары в сравнении с бродячими (link).
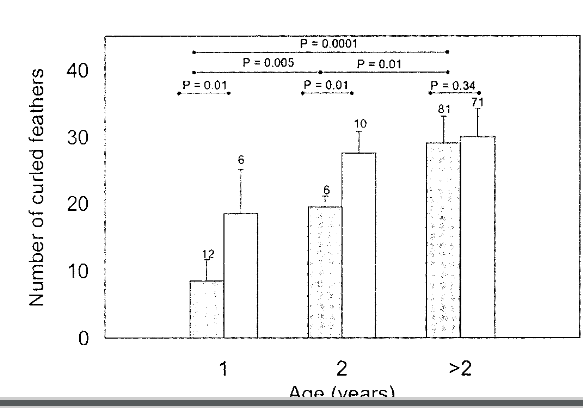 Рисунок 2. Возрастные и межполовые различия в числе кудрявых перьев у чёрного лебедя. Показано их среднее число на крыло (ордината) у птиц известного пола и возраста ± квартильное отклонение. Самки: заштрихованные столбики, самцы: открытые. Каждую особь отнесли лишь к одной категории, чтобы максимально увеличить размер выборки для птиц второго года жизни. Показана значимость различий средних между самцами и самками одного возраста (U-критерий Манна-Уитни). Они также значимы между возрастами у самок, но не самцов. Обратите внимание, что самцы и самки старше двух лет не отличаются по числу кудрявых перьев, в противоположность другим морфологическим признакам (табл.1).
Рисунок 2. Возрастные и межполовые различия в числе кудрявых перьев у чёрного лебедя. Показано их среднее число на крыло (ордината) у птиц известного пола и возраста ± квартильное отклонение. Самки: заштрихованные столбики, самцы: открытые. Каждую особь отнесли лишь к одной категории, чтобы максимально увеличить размер выборки для птиц второго года жизни. Показана значимость различий средних между самцами и самками одного возраста (U-критерий Манна-Уитни). Они также значимы между возрастами у самок, но не самцов. Обратите внимание, что самцы и самки старше двух лет не отличаются по числу кудрявых перьев, в противоположность другим морфологическим признакам (табл.1).
Таблица 1. Половой диморфизм по числу кудрявых перьев и другим морфологическим признакам у взрослых черных лебедей (3 года и старше).
Признаки — строки сверху вниз: число кудрявых перьев, длина клюва, общая длина головы, длина крыла, длина цевки (всё в см), масса тела, кг. Представлены средние ± ошибки средней, коэффициент вариации CV, %. Столбцы: самцы, самки, индекс полового диморфизма SDI, оцененный как разница средних значений признака у самца и самки, отнесенная у самочьей средней. Сравнения числа кудрявых перьев даны по U-критерию Манна-Уитни, другим признакам по t-критерию.
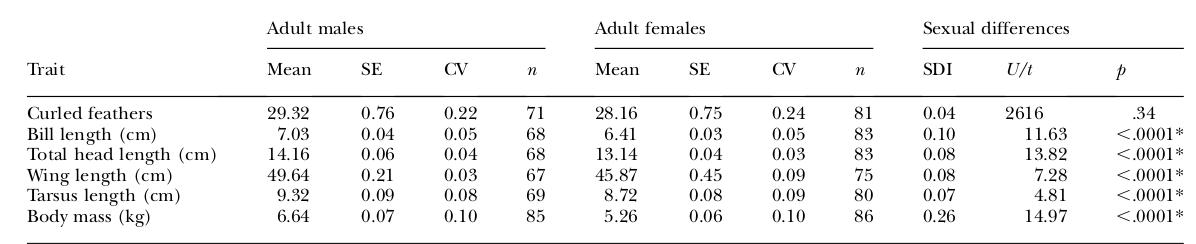 *различия значимы при p <0,05, с использованием поправки Бонферрони (см. Rice, 1989).
*различия значимы при p <0,05, с использованием поправки Бонферрони (см. Rice, 1989).
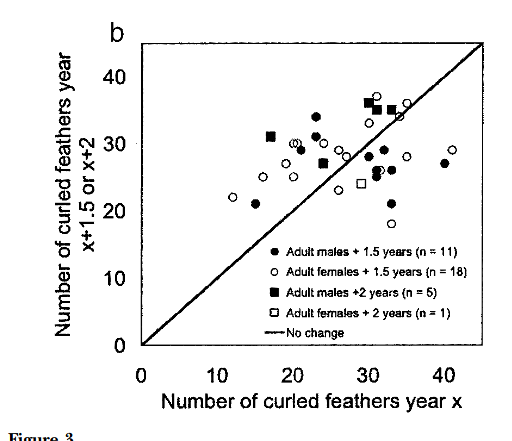
Рисунок 3а. Изменения числа кудрявых перьев у одних и тех же птиц обоего пола в последовательные годы, а. изменения через год: зимой 2001 г. в зависимости от их числа зимой 2000 г. b. изменения через 1,5 или 2 года: зимой 2000 г. или 2002 г. в зависимости от их числа летом 1999 и зимой 2000 г.
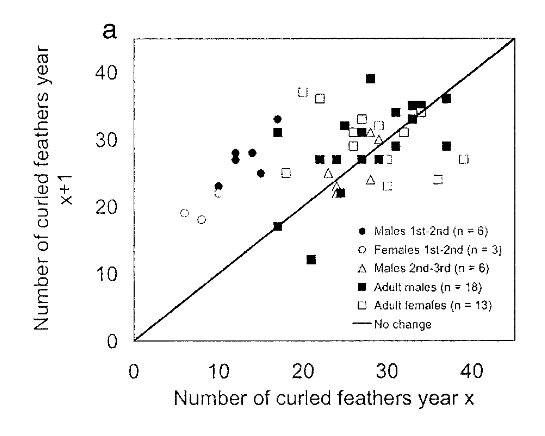
Рисунок 3b
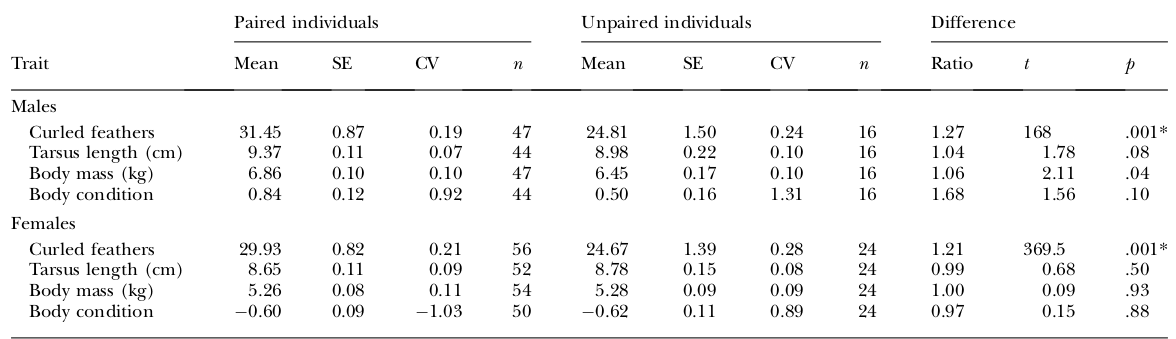
Таблица 2. Сравнение чёрных лебедей, находящихся в парах или нет, самцов и самок, по ряду морфологических признаков (строки сверху вниз — число завитых перьев, длина цевки, масса тела, физические кондиции. Показаны средние ± ошибка, коэффициент вариации, и разница между обоими категориями (еë величина и значимость по t- критерию).
Казалось, всё ясно – «украшения» важный сигнал-мессенджер социального отбора, действующего на соответствующие «ячейки общества» лебедей — пары с выводками (его действие их «выстраивает» в систему агрессивного доминирования), а никак не особи, ищущие партнёра и оцениваемые половым отбором. Но нет, авторы попытались по инерции и/или следуя моде объяснить ситуацию половым отбором, может быть потому, что у птиц, успешно образовавших пару и размножившихся, «украшения» сильно более развиты, чем у птиц без пары. Но поскольку у этого вида эффективное удержание территории – обязательное условие размножения, а не образования пары как такового – пара может занять территорию и потом, возникают сомнения.
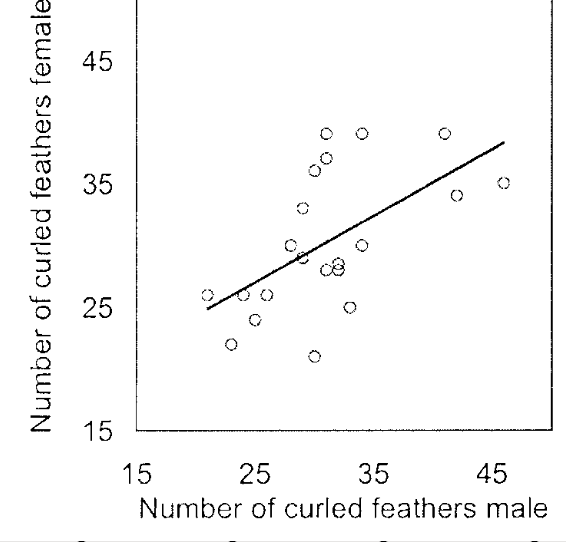 Рисунок 4. Взаимосвязь числа кудрявых перьев у самца и самки в одной паре (ранговая корреляция Спирмена rs =0,61, n = 22, p = 003).
Рисунок 4. Взаимосвязь числа кудрявых перьев у самца и самки в одной паре (ранговая корреляция Спирмена rs =0,61, n = 22, p = 003).
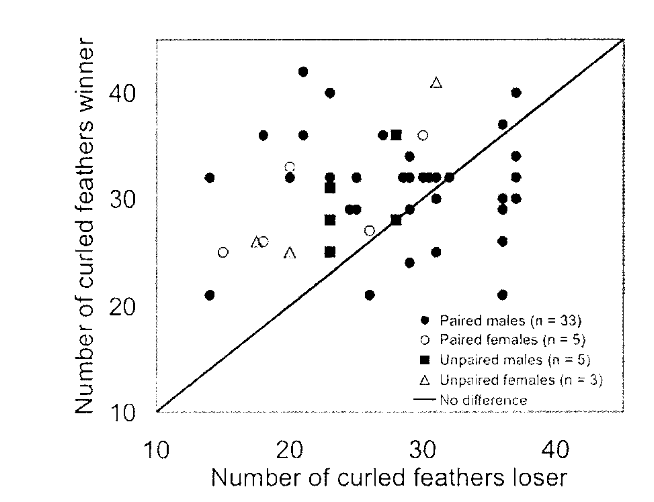
Рисунок 5. Взаимосвязь числа кудрявых перьев выигравших и проигравших в агонистических взаимодействиях лебедей (чёрное самцы, белое самки, кружки — в паре, квадратики и треугольники одиночки).
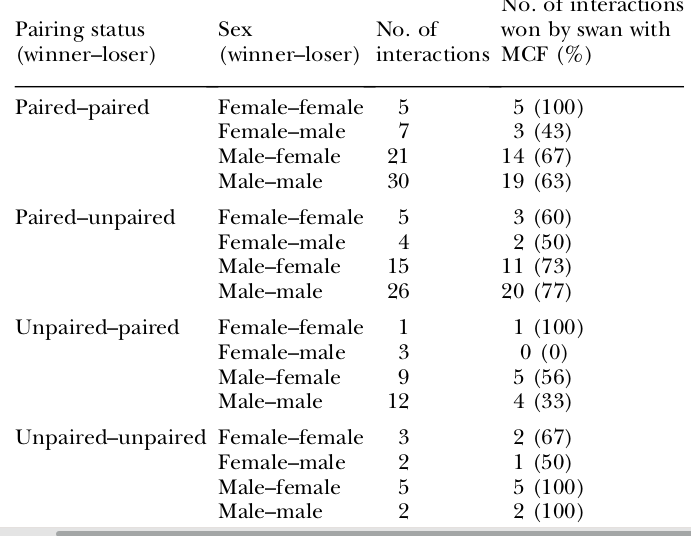
Таблица 3. Исходы агонистических взаимодействий между лебедями с разным числом кудрявых перьев. Строки — парный статус участников: находящиеся в паре против находящихся, то же против одиночек, одиночки против находящихся, одиночки против одиночек. В каждой строке показан половой состав участников: самки против самок, самки против самцов, самцы против самок и самцы против самцов. Следующие 2 столбца: общее число столкновений и % из них, выигранный птицами с большим числом кудрявых перьев.
Чтоб проверить, имеет отношение половой отбор к развитию «украшений» или нет, исследовали характер отцовства на 332 птенцах в 85 выводках лебедей за 3 сезона с использованием 8 полиморфных микросателлитных маркеров. У вида оказались распространены внутривидовой выводковый паразитизм (<5% птенцов в год) и внепарное отцовство. Последнее весьма развито: за 3 года 10-17% (в среднем 15,1%) всех лебедят появились в результате адюльтера. В 27-40% выводков (в среднем 37,6%) был, по крайней мере, один «неродной» птенец, что существенно выше, чем у других видов водоплавающих птиц со сходным жизненным циклом.
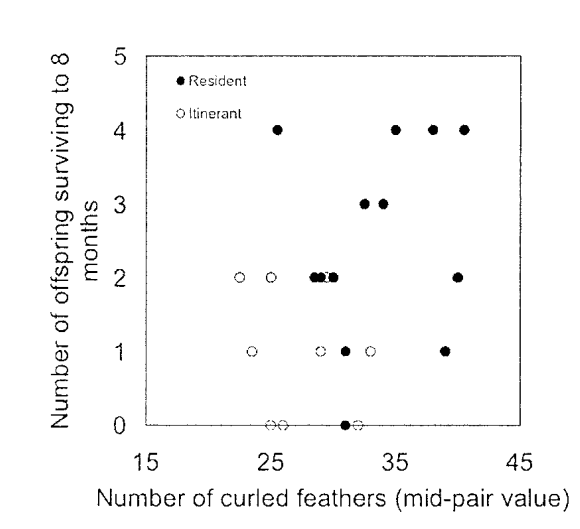
Рисунок 6. Число лебедят, выживших на протяжении минимум 8 мес., в зависимости от среднего числа кудрявых перьев родительской пары. Залитые точки — территориальные пары, полые — странствующие. Зависимость в целом отсутствует, при том что у территориальных пар таких перьев больше.
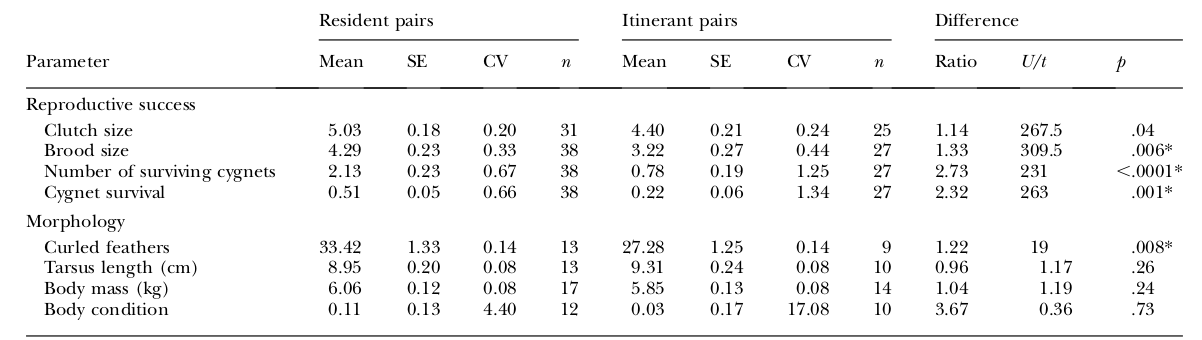
Таблица 5. Различия в репродуктивном успехе и морфологии (средние значения по обоим партнерам) постоянных vs бродячих пар (левый и центральный столбцы, правый — различия между ними по этим параметрам и его значимость, индекс Манна-Уитни U). Строки сверху вниз: параметры репродуктивного успеха — размеры кладки, выводка, число выживших лебедят и их выживаемость; параметры морфологии — число кудрявых перьев, длина цевки, см.; масса тела, кг; физическое состояние, баллы.
Однако разнокачественность особей по развитию кудрявых перьев оказывается незначима что при определении интенсивности их участия во внебрачных копуляциях, что при определении вероятности выкармливания ими чужих птенцов. Притом что разнокачественность особей по развитости кудрявых перьев очень высока, их разнокачественность по репродуктивному успеху существенно ниже, и внебрачное отцовство – в отличие от размера успешно удерживаемой территории – вносит существенно меньший вклад в вариабельность последнего (link). Другой аргумент в пользу социального отбора: выживаемость лебедят у территориальных пар в 2 раза выше, чем у бродячих, в первую очередь потому, что из-за большей успешности в территориальных конфликтах в первом случае птицы могут дольше и эффективней кормиться, как это отмечено и у малого лебедя Cygnus bewicki с белощёкой казаркой (Black, Owen, 1989; Scott, 1980). Если у белощёкой казарки преобладают крупные семьи, и в рамках вышеописанной социальной организации больший выводок позволяет им лучше сохранить территорию, то у чёрного лебедя число завитых перьев значимо лучше предсказывало владение территорией, чем размер выводка.
Увы, даже после соответствующих результатов авторы продолжают считать, что они подтверждают теорию полового отбора, просто он делается «взаимным», и, чтобы «спасти» это мнение от несоответствия данным, объясняют непонятное неизвестным, мол, кудрявые перья значимы в общей с гусями триумфальной церемонии, а через неё — и в образовании пары. А дальше переходят к следующему объяснению, также неподкреплённому данными: мол, самцы с большим количеством кудрявых перьев лучше защищают ресурс, важный для птенцов, самцы этого вида в норме больше насиживают, ergo, самки интенсивно конкурируют между собой за более качественных (=сильнее украшенных) самцов, как это описано у камышницы Gallinula chloropus (Petrie, 1983).
Однако для чёрного лебедя такого рода данные отсутствуют, притом что при образовании пары птицы сближаются с несколькими потенциальными партнёрами, тестируют их «качества» в процессе ухаживания, прежде чем делают окончательный выбор (как и другие виды гусей с постоянными парами, вроде белощёкой казарки Branta bernicla Choudhury and Black, 1993; Van der Jeugd and Blaakmeer, 2001). Накручивая одно непроверенное (и маловероятное) объяснение на другое, авторы дальше предполагают, что такое «тестирование» может быть, продолжается и после образования пары, на стадии её консолидации… — хотя это уже «хайли лайкли«, прямо противоречащее низкой частоте «разводов» в данной популяции чёрного лебедя.
Далее авторы справедливо указывают, что успешность «лучше украшенных» особей в территориальных конфликтах особенно адаптивна в наиболее плотных популяциях, создаёт там давление отбора в сторону увеличения числа кудрявых перьев, и именно чёрные лебеди [и шипуны, у которых такие перья отсутствуют], в отличие от других лебедей, часто гнездятся колониями — однако не замечают, что это довод против эффекта полового отбора и за эффект социального.
Аналогичная ситуация наблюдается, когда «украшение» есть только у одного пола. У пеганок – это красный нарост на клюве самца. Пеганки, гнездящиеся в норах – моногамные полугуси, самец также охраняет и водит выводок, как у лебедей. Самцы с большим наростом более успешны в охране территорий, которые весной имеют кормовое значение. Их партнёрши всяко выигрывают: могут больше кормиться, проявлять меньше бдительности и пр., поскольку к успешным самцам после нескольких поражений противники вторгаются редко. И наоборот, пары, где нарост на клюве самца был небольшой, часто втягивались в изматывающие конфликты с соседями.
Поэтому самки предпочитают самцов с большим наростом, но последние не пользуются этим для максимизации внебрачных спариваний – последние вполне присутствуют, как и у чёрного лебедя, но с развитием «украшений» их вероятность не связана. То есть особи конкурируют за лучшие «позиции» в социальной системе, «конструкция» которой для животных в сообществе, совместно реализующих брачные или иные инстинкты, суть большая реальность, чем непосредственный успех каждой из них в виде спаривания, выгонки противника и пр. Незамечание этого «слона» — главнейший (и неустранимый) дефект социобиологии как исследовательской программы, конкурирующей с таковой сравнительной этологии, он ответственен за куда меньшую состоятельность построений первой в сравнении со второй.
Рекомендуем прочесть
 Два механизма «копирования поведения»
Два механизма «копирования поведения» Ещё раз про «пернатых многожёнцев»
Ещё раз про «пернатых многожёнцев» Внутри- и межвидовое копирование поведения, на примере открывания молочных бутылок синицами
Внутри- и межвидовое копирование поведения, на примере открывания молочных бутылок синицами «Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования
«Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования Устойчивые морфологические типы в эволюции птиц
Устойчивые морфологические типы в эволюции птиц Социальная организация гусеобразных (Аnseriformes): базовые принципы и основные направления эволюции
Социальная организация гусеобразных (Аnseriformes): базовые принципы и основные направления эволюции Против «обмена еды на секс»
Против «обмена еды на секс» Взаимоотношения полов у позвоночных и модели полового отбора
Взаимоотношения полов у позвоночных и модели полового отбора
