
Альфред Зайц (1905-1982).
Резюме. Рассказано о законе гетерогенной суммации, открытом Альфредом Зайцем: он был первым учеником Лоренца, но его жизнь и научные результаты изрядно забылись, поэтому также рассказана биография. Суть закона: общий эффект комбинации из качественно различных ключевых стимулов, на которые реагирует один и тот же врожденный разрешающий механизм = сумме эффектов каждого из релизеров, действующих по отдельности; каждая составляющая сложного стимула, обладающего релизерным эффектом, действует независимо от других; значимый образ партнёра воспринимается не как целое, но сложен суммой отдельных специфических черт, каждая из которых действует как отдельный релизер: все вместе они присутствуют в данном образе, но отнюдь не исчерпывают его. Закон отчасти нарушается, когда речь идёт о демонстрациях — знаковых стимулах, их организованном обмене в процессе коммуникации, а не просто о визуальном образе компаньона. Целостный образ демонстрации как релизер эффективней, чем сумма её компонент, т.е. действие составляющих сложного сигнала не аддитивно, а мультипликативно. Степень нарушения прямо пропорциональна «сигнальности» и «знаковости» демонстраций: когда демонстрации из стимулов превращаются в произвольный знак, закон Зайца не выполняется вовсе, значащие единицы (инвариант формы соответствующих демонстраций) составляются из незначащих элементов, элементарных двигательных актов или других моделей движений. Для знаков – сигнальное воздействие частей равно нулю, они нужны только для образования целого, которое и осуществляет всё воздействие полностью.
Биография
Немецкий этолог и орнитолог Альфред Зайц (Alfred Seitz) родился 21 февраля 1905 года в Гернсхайме, Великое герцогство Гессен; умер 2 августа 1982 года в Нюрнберге. Он вырос в Вене, где отец был директором крупного завода по производству буры. На заводской территории был пруд с небольшими тростниковыми зарослями, где гнездились малые выпи (волчки) Ixobrychus minutus. Как говорил мой учитель Геннадий Николаевич Симкин, учёного делают именно и только открытия, нужно узнать что-то новое про естественную историю птиц, что до этого не знали другие. Первое открытие А.Зайца было про выкармливание птенцов у волчков. Рано заинтересовавшись фотографированием поведения птиц, по окончанию средней школы в 1924 г. он выучился на фотографа, сумел приучить волчков к своему присутствию, благодаря чему наблюдал их размножение с близкого расстояния.
Как и прочие цапли, волчок насиживает кладку с первого яйца: учитывая большое число яиц, самый старший птенец в выводке часто на добрую неделю старше самого младшего. У других цапель — серых Ardea cinerea, средних белых Egretta intermedia или рыжих A.purpurea — много меньшая разница в возрасте между птенцами практически лишает самого младшего шансов на выживание: родители дают корм в первую очередь старшим, активней всего выпрашивающим. Во всяком случае, разница размеров птенцов перед вылетом здесь значительна и хорошо заметна. Однако оперяющиеся птенцы волчков почти одинакового размера, притом что их много в выводке.
Зайц выяснил, в чем загадка. Его прекрасные фотографии показали, что принёсшие корм взрослые выборочно дают его самым маленьким, даже когда те спят и не выпрашивают, и будят их для кормления их будят. Старшим, более крупным и активно выпрашивающим, приходится ждать, пока не покормят малых и слабых, даже если те спят и приходится их будить.
 Первые вылупившиеся птенцы волчка
Первые вылупившиеся птенцы волчка
В 1920е гг. Альфред занимался коммерцией, одновременно участвуя в этологических исследованиях Конрада Лоренца, став его первым учеником (второй — Николаас Тинберген, сооснователь сравнительной этологии). Он был не только сотрудником, но и близким другом Лоренца. В том числе и как фото/кинооператор, снявший первые фильмы по социальному поведению птиц. Один из них 1937 г., про серого гуся, доступен доселе в Немецкой национальной научно-технической библиотеке (DIN) онлайн. Его Зайц снял на старой плёнке Gevärt-Film, подаренной Венским анатомическим институтом, на дряхлую камеру, снимавшую ровно 12 с. с каждого запуска (на холоде несколько дольше, с соответствующим снижением частоты кадров). Однако он до сих пор один из самых востребованных.
Когда Зайц закончил изучение зоологии в Вене, Лоренц предложил ему защищать диссертацию по этологии образования пар у цихлид, ибо в Зоологическом институте в Вене хватало аквариумов (правда поначалу открыт был лишь верхний ряд трехэтажной конструкции аквариума). Он долго сопротивлялся, будучи орнитологом душой и телом, однако всего за несколько недель узнал цихлид настолько же хорошо, как птиц, и понимал их поведение так же тонко. Его две работы по образованию пар у двух видов цихлид вышедшие в Zeitschrift für Tierpsychologie 1940 и 1942 гг. [1 и 2], стали классическими. Один вид тогда звался Astatotilapia strigigena, сейчас систематическое положение этих рыб, увы, непонятно [пишет Лоренц в 1965 г., сейчас их относят к Astatotilapia bloyeti], другой — Hemichromis bimaculatus [хемихромис-красавец. Прим.публ.].

 Astatotilapia strigigena и Hemichromis bimaculatus
Astatotilapia strigigena и Hemichromis bimaculatus
В 1939 г. Зайц защитил диссертацию по цихлидам, в рамках которой ему посчастливилось открыть настоящий закон природы, именно, закон гетерогенной суммации (нём. Reizsummenregel, англ. Law of heterogenic summation): общий эффект комбинации из качественно разных ключевых стимулов (социальных релизеров), на которые реагирует один и тот же врожденный разрешающий механизм (нем. Angeborenes auslösendes Schema, AAS) = суммой эффектов каждого из релизеров, действующих по отдельно. Этот закон очаровал Лоренца, именно в этом время показавшего важность распознавания образов для действенности ключевых раздражителей, их действие как гештальтов (Lorenz, 1939), однако тогда не был опубликован.
В 1949 г. другой ученик Лоренца, Хайнц Прехтль, усложнил представление о внутренней организации врожденного разрешающего механизма. Он обнаружил, что инстинктивное действие, будучи предъявленным несколько раз подряд, перестаёт возникать, однако немедля возобновляется в ответ на другой релизер, относящийся к тому же действию, и перед этим не предъявлявшийся1.
Из этого был сделан вывод, что во врожденном разрешающем механизме может возникать частичная истощаемость, относящаяся к его реакции на один из ключевых стимулов, причём нетождественная истощаемости моторного ответа.

Открытие законов в поведенческих исследованиях бывает нечасто, обычно всё ограничивается описанием наблюдаемого. В том же году д-р Зайц стал куратором государственного музея Бургенланда в г. Айзенштадт в Австрии. В 1940 году его призвали в вермахт; по счастью, служить до 1945 г. ему довелось в Кёнигсберге, и поначалу научным сотрудником у Конрада Лоренца, назначенном в том же году профессором сравнительной психологии на кафедре психологии философского факультета Кёнигсбергского университета (Лоренц — последний профессор кафедры Канта). Правда, в 1942 г. потом Лоренц отбыл в Польшу, где совершал военные преступления, а в 1944 г. на Восточный фронт, но это уже совсем другая история…
В плену Зайц пробыл недолго в Новосибирске; уже в 1946 году во Франкфурт-на-Майне он помогал Бернхарду Гржимеку в реконструкции Франкфуртского зоопарка. Тогда же он опубликовал статью о наземном брачном поведении чибиса Vanellus vanellus [малозаметном, в противоположность воздушным демонстрациям] (ZfT2 1950. Bd.7. S.275–286) и о взаимоотношениях с компаньонами в «мире» [большого] кроншнепа Numenius arquata (ZfT 1950. Bd.7. S.402–417). Будучи ассистентом, он во Франкфуртском зоологическом саду начал свою работу по [сравнительной этологии в связи с филогенетическими связями] псовых (ZfT 1950. Bd.7., S.1–46), успешно продолжив её впоследствии с в Нюрнберге, чему способствовали наблюдения за видами в коллекции зоопарка. В 1950 году он стал директором Нюрнбергского зоопарка: всякий знающий зоосад тогда и сейчас [т.е. в 1965 г., прим.публ.], оценит масштабы усилий и размеры свершëнного.
За время его работы, помимо прочего, были реконструированы обезьянник, вольеры для львов, фламинго и кенгуру, вольер для бегемотов и вольер для тюленей, а также основано Общество друзей Нюрнбергского зоопарка. К 1959 году популяция животных достигла довоенного уровня, а в 1964 году жирафы впервые переехали в недавно построенный жирафятник.
Несмотря на огромную нагрузку, связанную с реконструкцией этого великолепного зоопарка, за 14 лет Зайц внес значительный вклад в науку. В Нюрнберге он завершил начатое во Франкфурт исследование восприятия форм и различения размеров у восточно-прусских овец породы ландрас3 (ZfT 8, 423–441); продолжил детально изучать сравнительную этологию псовых: исследовал енотовидную собаку Nyctereutes, богатую примитивными признаками и близкородственную енотовым Procyonidae4 (ZfT 12, 463–489). Следуя устоявшейся традиции Хайнрота, он начал изучение каждого вида с выращивания одной или нескольких особей в изоляции с самых ранних лет или даже с рождения5. Следует отметить, что он это делал всегда в собственном доме, при преданной поддержке своей жены Марии, не стеснявшейся растить у себя даже белого медведя, от чего сохранились почётные шрамы.
Дальнейшие исследования по сравнительной этологии псовых (ZfT 16, 747–771) пролили новый свет на всё ещё весьма неясные родствнные связи в группе, включая происхождение домашней собаки. Детальное изучение золотистого шакала установило неверность точек зрения Хайнрота и Лоренца, что золотистый шакал Canis aureus главнейший предок домашней собаки. Характерный шакалий вой в той или иной форме реализовался бы у домашних собак, содержи они значимую долю шакальей крови. С другой стороны, плодовитость гибридов от скрещивания разных псовых (того же золотистого шакала с койотом Canis latrans), продемонстрированное Зайцем, подтверждает возможность полифилетического происхождения домашней собаки [сейчас эта точка зрения опровергнута молекулярными данными — только волк, в том числе для басенджи].

Басенджи, африканская нелающая собака
Зайц предположил, что басенджи, африканская нелающая собака, единственная порода с кровью золотистого шакала [сегодня мы знаем, что это не так, однако басенджи, как динго и хаски, были приручены до появления с/х, когда жили только охотники-собиратели]. Как и многие смотрители зоопарков, и как наш учитель [Оскар] Хейнрот, Зайц накопил огромный массив данных наблюдений, дополненных фотодокументами, на публикацию которых нет хватает времени. За годы работы в Нюрнберге Зайц также воспитал отличных учеников, о чём свидетельствуют, например, работы Хаймбургера (ZfT Bd. 16. H.104–113 и Bd. 18. H.265–284).
Все жители Нюрнберга, все друзья Нюрнбергского зоопарка справедливо сочтут большой потерей достижение Зайцем установленного законом пенсионного возраста: ему это слишком рано. Однако его коллеги-этологи утешатся мыслью, что будет время обработать и опубликовать огромный фактический материал, собранный за время работы в зоопарке. Нас беспокоит только то, найдутся ли ответственные органы, которые предоставят этому специалисту, высоко ценимому всеми нами, человеку, материальные возможности и дальше разводить свои объекты, исследовать их и пр.
Источники
Конрад Лоренц: 60 лет Альфреду Зайцу// Zeitschrift für Tierpsychologie. 1965. Bd. 22. H. 5, S. 600–602.
Закон гетерогенной суммации — когда выполняется, когда нет?
Коротко повторю суть закона: общий эффект комбинации из качественно различных ключевых стимулов, на которые реагирует один и тот же врожденный разрешающий механизм = сумме эффектов каждого из релизеров, действующих по отдельности; каждая составляющая сложного стимула, обладающего релизерным эффектом, действует независимо от других; значимый образ партнёра воспринимается не как целое, но сложен суммой отдельных специфических черт, каждая из которых действует как отдельный релизер: все вместе они присутствующих в данном образе, но отнюдь не исчерпывают его.
Лоренц в 1965 г. полагал что
«только недавно oбширные корреляционные исследования Heiligenberg показали, что в отдельных случаях эффекты двух ключевых стимулов не аддитивны, а мультипликативны, во всех остальных оно полностью подтверждается».
Скажем, G.P. Baerends в исследовании, поданном в сборник к 75-летию Отто Кёлера, показал, что закону гетерогенной суммации подчиняются и реакции на выученные раздражители: эффект комплексного воздействия равен сумме эффектов каждой из компонент. Это еще одно проявление известного правила параллельности инстинктивного и выученного поведения, как в отношении формы, так и функции.
Зайц работал с цихлидами, где практически все визуальные демонстрации (ухаживания и угрозы) складываются минимум из трёх компонентов. Первый это собственно демонстрация, то есть ритуализированные телодвижения или их комплексы, внешний образ которых разительно отличается от образа рыбки во время обыденной позы, повседневной активности и т.п. Второй компонент — это рисунок на теле рыбы, разнообразные пятна и полосы: почти все они имеют сигнальное значение, но у разных особей варьирует как их размер, так и (для рыб это главное) угол наклона полос, особенно образующих лицевую «маску» относительно линии движения рыбы и линии демонстрирования при исполнении демонстраций. А третий – это раскраска, как бы «интенсивность заливки» структурных элементов рисунка. Соответственно, действенность сигнала (измеряемая интенсивностью специфической реакции на его предъявления) есть некая функция от выраженности всех трёх элементов, и важно исследовать, может ли недоразвитие одного компонента компенсироваться переразвитием другого?
Закон гетерогенной суммации говорит нам – да, может, но лишь для стимулов и не для знаков (причём для знаковых стимулов – релизеров классических этологов – хуже, чем для «просто» специфических стимулов). Согласно ему, общий эффект комплексного стимула или комплексной демонстрации равен алгебраической сумме эффектов воздействия отдельных компонент – элементарных стимулов. То есть наблюдается аддитивность действия таких релизеров, как форма яйца, внешний облик брачных украшений партнёра, или образ демонстрации, «предъявляющей» украшения – все их можно подразделить на несколько составных частей, потенциально с собственным сигнальным эффектом у каждой.
Так, размер кладки у серебристых чаек Larus argentatus фиксирован – три яйца. Если кладку уменьшить искусственно, чайки затаскивают в гнездо и поддельные яйца (положенные на край гнезда или не слишком далеко от него), когда те в достаточной степени сходны с настоящими. «Степень достаточности» сходства предмета на краю гнезда с собственным яйцом задана определённым сочетанием таких признаков формы, как окраска и размеры яйца, они же составляющие комплексного стимула. Если предмет соответствует сочетанию, он распознаётся как «своё яйцо» и затаскивается в гнездо (рис.1). Воздействие первой и второй компонент аддитивно — некоторая «нетипичность окраски» компенсируется сверх-типичным размером и формой, и наоборот (Baerends, Krujt, 1973).
 А.
А.
Б.
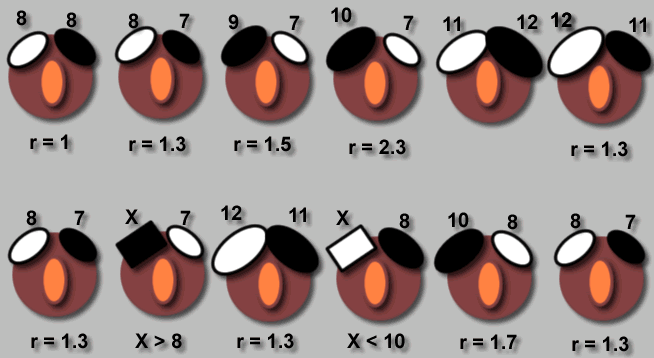
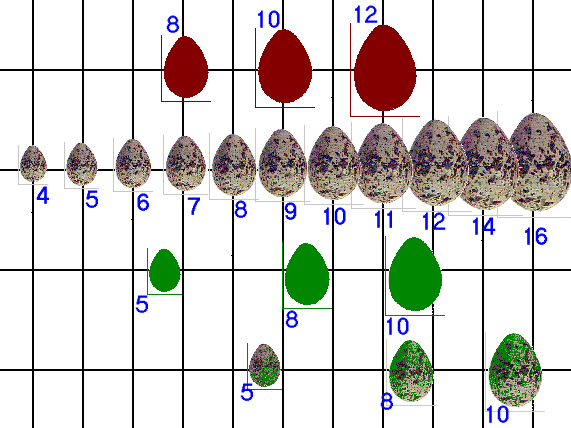
Рисунок 1. Аддитивность воздействия составляющих комплексного стимула, заставляющих серебристых чаек распознавать овальный предмет как «своё яйцо», с последующим закатыванием в гнездо.
А. Количественный метод оценки поддельного яйца. Обозначения. Кружок — гнездо с одним яйцом внутри него и двумя поддельными яйцами на краю, между которыми чайка делает выбор. Цифры 7-12 – число поддельных яиц, r — соотношение между размерами двух поддельных яиц на краю гнезда. X — модель, подлежащая измерению. Выбранное поддельное яйцо показано черным цветом.
Примечание. I. Количественная оценка предпочтений в расположении яйца. Первый рисунок (верхний угол слева) показывает, что предпочтение оказано правому яйцу. Это предпочтение сохраняется, когда с правой стороны положено яйцо меньших размеров (на следующем рисунке в серии). Последовательный ряд проб показывает, что предпочтение положения можно выразить величинами между r = 1,3 и r = 1,5. II. Количественная оценка модели X. Предпочитаемое значение Х лежит между размерами 8 и 9.
Б. – результаты анализа параметров, определяющих предпочтение чаек. Обозначения. Средние значения для разных типов поддельных яиц (А-Г) даны по отношению к ряду стандартных размеров r. А – яйца коричневые с крапинками, в форме кирпичиков; Б — коричневые без крапинок, яйцевидные; В — зеленые без крапинок, яйцевидные; Г — зеленые с крапинками, яйцевидные. 4-16 — величина яйца. Равные расстояния между точками на шкале указывают на равные значения r.
Heiligenberg с сотрудниками (Leong et al., 1969) изучил релизеры территориальной и/или брачной агрессии у цихлид Haplochromis burtoni. Взрослые самцы обитали в аквариумах с молодью: самец может напасть на них, но те легко удирают, конфликтов не происходит. «Базовый» уровень агрессии оценивали по числу атак на молодь за 15 мин до начала тестирования. Дальше через стекло им предъявляли чучело взрослого самца (30 с: самец обычно спокойно смотрит на неё), и в следующие 30 мин регистрировали число атак на молодь (мера подъёма агрессии, вызванной видом потенциального соперника). Для количественной оценки воздействия разных составляющих образа соперника как социального релизера меняли наличие и положение трёх составляющих последнего — полосы через глаз, пятна за глазом и рисунок тела/плавников (серии муляжей I-IV, fig.6, 8, 10, 12 в: Leong et al., 1969).
Мерой реактивности в ответе на релизер был подъём агрессивности в отношении молоди после демонстрации чучела; среднее число атак сперва подскакивает, а затем возвращается к исходному уровню, отражая индивидуальные различия между самцами (рис.2, 3).
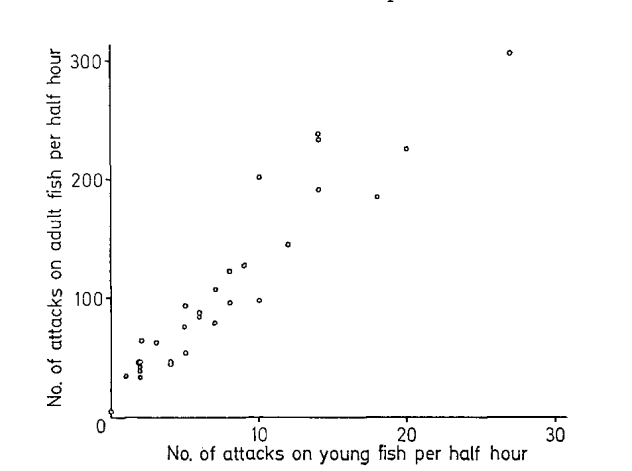 Рисунок 2. Интенсивность атак на взрослых рыб (ордината) и на молодь (ось абсцисс) на протяжении получаса положительно связаны между собой. Оба параметра годятся как мера готовности атаковать.
Рисунок 2. Интенсивность атак на взрослых рыб (ордината) и на молодь (ось абсцисс) на протяжении получаса положительно связаны между собой. Оба параметра годятся как мера готовности атаковать.
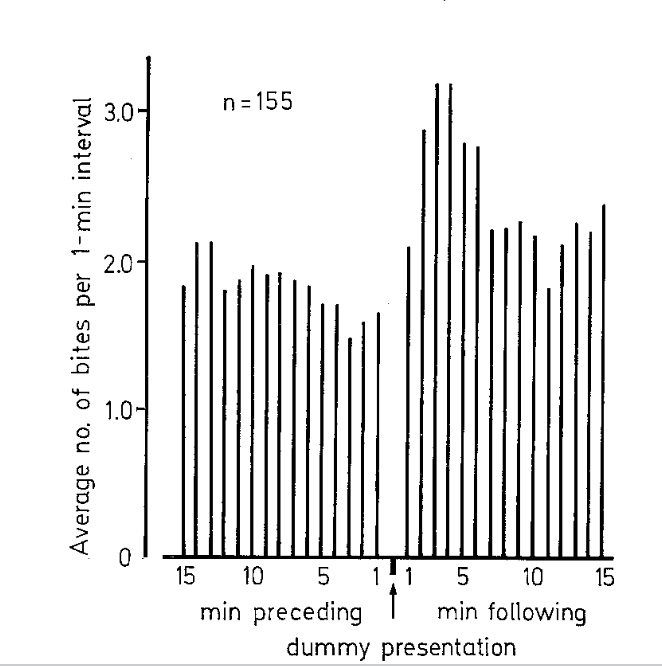 Рисунок 3. Среднее число укусов в мин (абсцисса) подскакивает после предъявления муляжа, затем возвращается к исходному уровню, данные по 400 с интервалами «до» и «после».
Рисунок 3. Среднее число укусов в мин (абсцисса) подскакивает после предъявления муляжа, затем возвращается к исходному уровню, данные по 400 с интервалами «до» и «после».
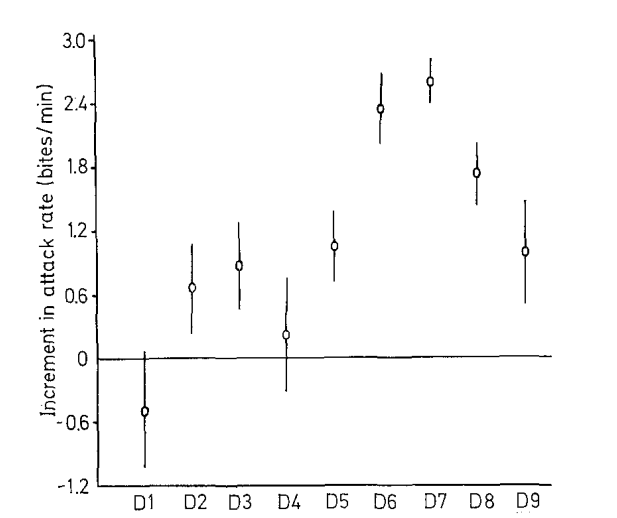
Выяснилось, что для действенности образа достаточно двух компонент: чёрной полосы через глаз и оранжевого пятна за глазом, при несущественности третьего. Полоса, предъявленная без пятна, усиливает агрессию на 2,79 ударов/мин, пятно без полосы снижает на 1,77 ударов/мин. Демонстрация чучела с обоими признаками учащает атаки на 1,08 ударов/мин, т.е. на алгебраическую сумму эффектов воздействия полосы и пятна (2,79 + (–1,77)), они аддитивны в вызывании агрессии (Leong et al., 1969). Влияние цветовых узоров на теле рыбки изучено с предъявлением четырёх рядов моделей: видно, что одни элементы усиливают, другие же ослабляют прирост агрессии на рис.4, их эффект везде аддитивен (рис.5).
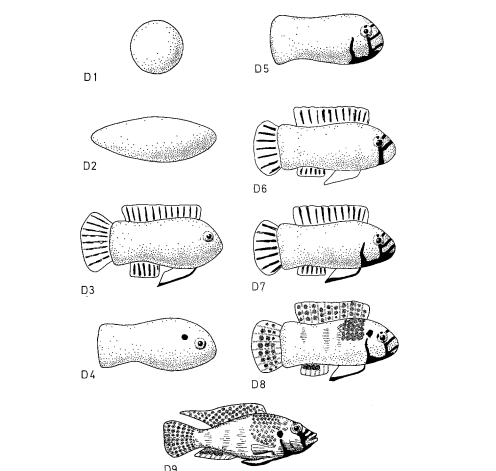
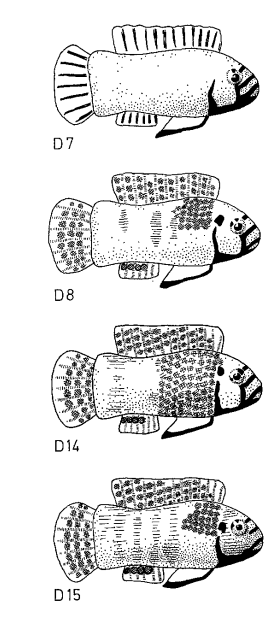
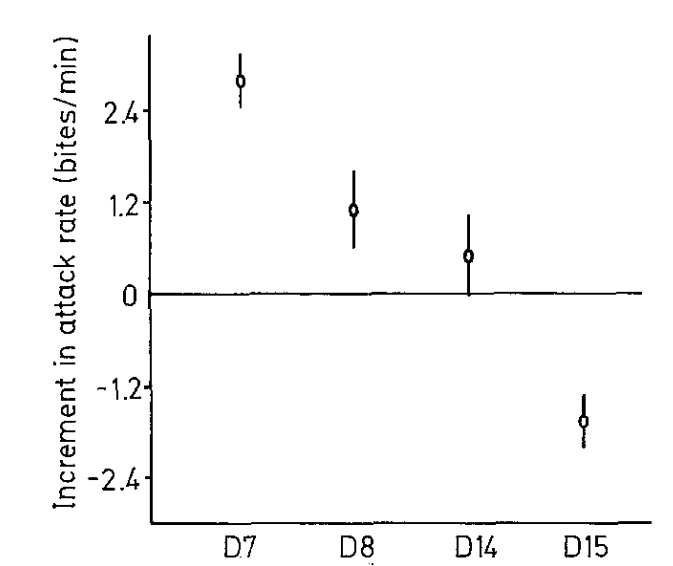
Первый ряд включал 9 моделей, форма и паттерны окраски которых различаются степенью приближения к рыбке в готовности охранять территорию, второй — 4 модели, различающиеся наличием иди отсутствием оранжевого пятна в сочетании с природной окраской, третий — 3, различающиеся «степенью заливки» черных полос на туловище, четвёртый — 4, в ряду которых растёт распространение оранжевого цвета на туловище. Графики для всех 4х вариантов показывают изменения частоты ударов после предъявления соответствующих моделей, средняя ± дисперсия. Видно, что одни составляющие окраски усиливают агрессивный ответ, другие ослабляют, третьи же несущественны (рис.5I-IV). Видимо, эти цветовые узоры поддерживают уровень готовности к атаке среди территориальных самцов в колонии в определённых пределах (Leong et al., 1969).
 I.
I.
 II.
II.
 III.
III.
 IV.
IV.
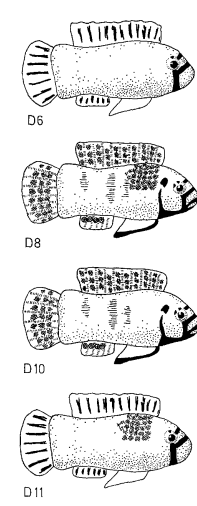
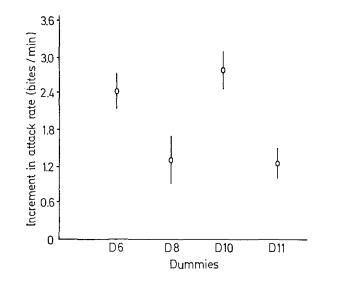
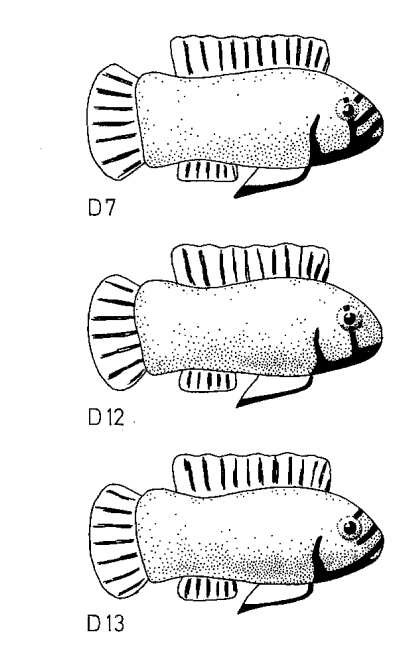
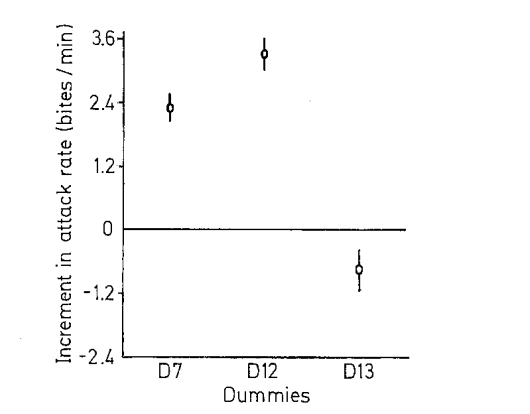
Рисунок 5. Воздействие 4-х серий моделей на уровень агрессивности. I. Внешний вид первой серии моделей (D1-D9, заливка чёрным — черный цвет, клетчатым — синий, горизонтальные полосы — оранжевый) и изменения уровня агрессивности при их предъявлении (абсцисса — тип модели, ордината — прирост агрессивности, ударов/мин). II. То же для второй серии (D6, D8, D10-11, обозначения те же). III. То же для третьей (D7, D12, D13). IV. То же для четвёртой (D7, D8, D14, D15).
Аналогичный эффект оказывает угол ориентации чёрной полосы через глаз: вызываемая агрессия ослабляется пропорционально отклонению от типичного положения, определенную роль играет ориентация всего туловища (рис.6, 7).
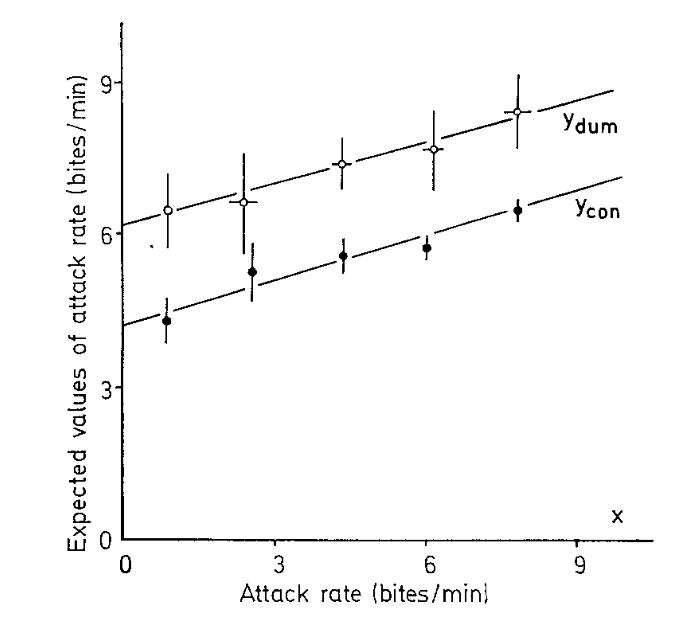 Рисунок 4. Ожидаемая частота нападений самцов H.burtoni (ордината) в зависимости от предшествующей частоты (укусов/мин, абсцисса), данные по 400 с интервалам «до» и «после». Эти данные показывают, что по сравнению с контрольной (Ycon) ситуацией предъявление искусственной рыбы-соперника (Ydum) оказывает аддитивное действие на частоту нападений.
Рисунок 4. Ожидаемая частота нападений самцов H.burtoni (ордината) в зависимости от предшествующей частоты (укусов/мин, абсцисса), данные по 400 с интервалам «до» и «после». Эти данные показывают, что по сравнению с контрольной (Ycon) ситуацией предъявление искусственной рыбы-соперника (Ydum) оказывает аддитивное действие на частоту нападений.
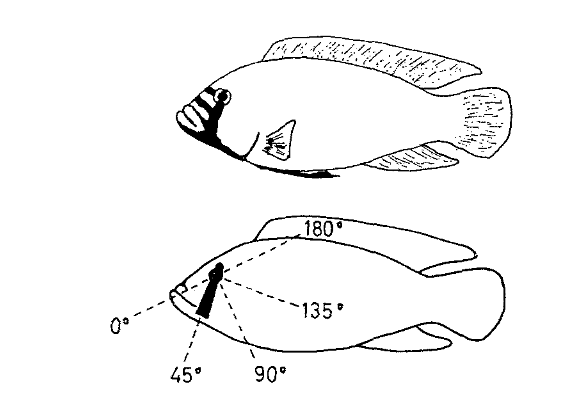
Рисунок 6. Сверху настоящий самец хаплохромиса, снизу муляж с черной «глазной полоской» из фольги, которую можно вращать вокруг центра глаза. Для тестирования использовали 5 положений полоски, различающихся углом между линией полоски и профилем линией лба (или рыла).
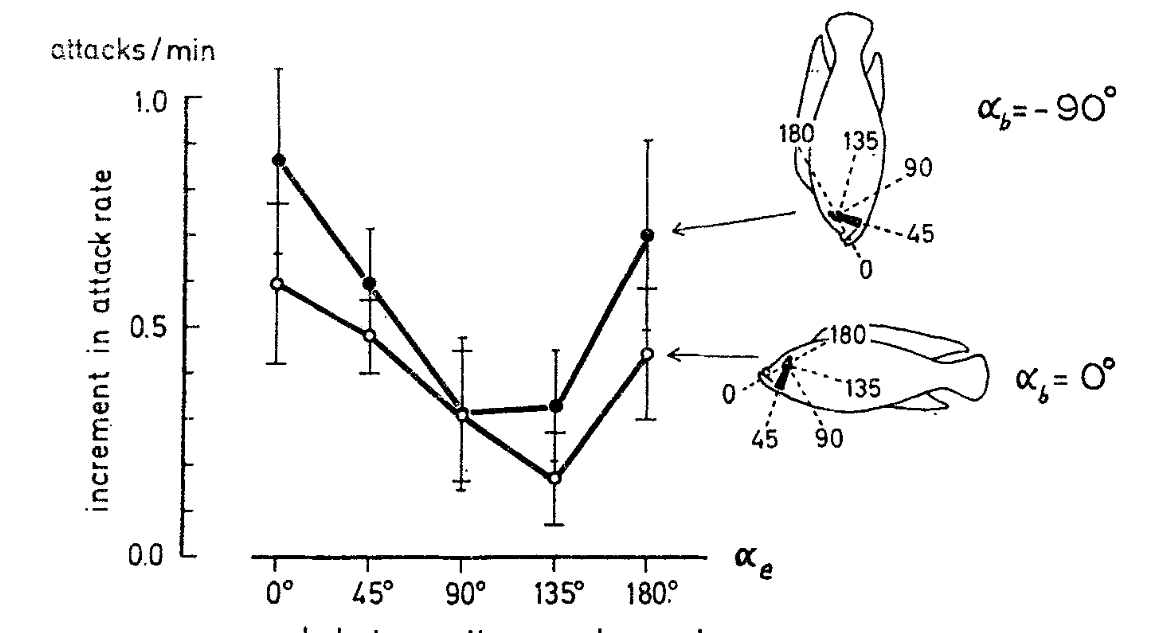 Рисунок 7. Среднее приращение частоты атак (ордината, ударов/мин) как функция угла наклона чёрной полоски относительно профиля лба (или рыла) модели. Белые кружки — модель предъявлена ориентированной горизонтально, чёрные — вертикально головой вниз, «усы» — дисперсия средних. Объединены данные по 6 разным самцам.
Рисунок 7. Среднее приращение частоты атак (ордината, ударов/мин) как функция угла наклона чёрной полоски относительно профиля лба (или рыла) модели. Белые кружки — модель предъявлена ориентированной горизонтально, чёрные — вертикально головой вниз, «усы» — дисперсия средних. Объединены данные по 6 разным самцам.
Используя тот же дизайн опытов, Heiligenberg et al. (1972, 1976) изучили влияние ориентации чёрной полосы через глаз и положения туловища Haplochromis burtoni на реализованную агрессивность рыбки (рис.5, 6). Общий подъем частоты ударов исчезал примерно через 3 мин после предъявления модели; её снижение в первую минуту предъявления вызвано общим снижением активности впервые столкнувшейся с ней рыбки. Первую изменяли, вращая полоску фольги вокруг глаза, второе меняли от обычного до вертикального головой вниз (рис.6). Уровень агрессивности в следующем интервале по 5 мин линейно связана с таковой в предыдущем (эффект саморазогревания). Предъявление модели его дополнительно увеличивает (белые кружки), как бы «переводя» на выше расположенную прямую, параллельную данной (черные, средняя ± дисперсия относительно обоих координат), а иногда ниже. Величина сдвига соответствует эффекту модели.
Использовали 5 разных углов наклона относительно оси тела предъявляемой модели - 00, 450, 900, 1350 и 1800 (n=68, 261, 57, 179 и 75 для горизонтального и n=58, 159, 59, 158 и 59 для вертикального положения модели при предъявлении, рис.7): наибольшее приращение агрессивности происходит при параллельности чёрной полосы профилю лба, минимум ударов достаётся модели при её перпендикулярности. Данные изменения не зависят от положения тела модели, однако расположение «головой вниз» дополнительно увеличивает частоту атак.
Аддитивность эффектов отдельных составляющих комплексного стимула объясняет их взаимозаменяемость в вызывании нужной реакции на комплекс как целое. Основываясь на законе гетерогенной суммации, можно строить изоклины эффективности стимуляции, отражающие разное соотношение вкладов альтернативных компонент в реакцию на выходе, и по их форме судить, насколько закон соблюдён в каждом конкретном случае.
Так, эффективность брачной стимуляции самцом гуппи самки определяется двумя частично взаимозамемяемыми параметрами – развитостью специфического рисунка в задней части тела самца и интенсивностью собственно брачных демонстраций, с гиперболической зависимостью между ними (рис.8).
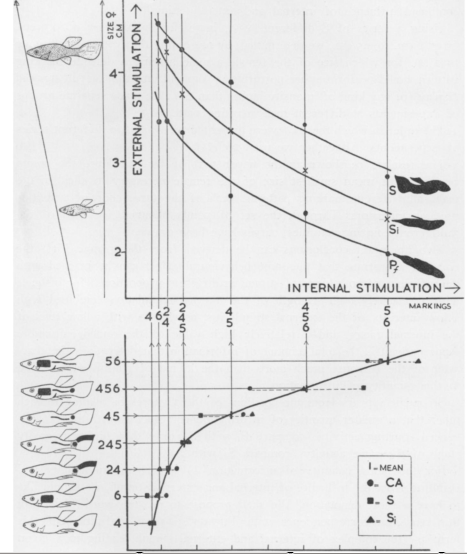
Рисунок 8. Изоклины сигнальной силы в ухаживании самца гуппи Lebistes reticulates (сверху). Снизу – калибровочная кривая для оценки уровня сексуальной мотивации по развитости цветного рисунка на его теле, развитость разных элементов рисунка задаёт «место» на оси абсцисс. По: Baerends et al., 1955, fig.24.
Обозначения. Сверху. Ось У – сила внешней стимуляции (возрастающий снизу вверх размер самки), ось Х – сила внутренней стимуляции самца (уровень сексуальной мотивации), суммирование обоих факторов детерминирует форму выражения брачных демонстраций самца (развитость сигмоидных демонстраций). Линии – изоклины сигнальной силы демонстраций самца (позирование – минимальная, полусигмоидная – средняя, сигмоидная – максимальная). Примечание. Сигмоидная поза (или «степень сигмоидности» положения тела самца) – наиболее ритуализированное (и наиболее интенсивное) исполнение специфической демонстрации хвостом, которой самец останавливает плывущую самку и побуждает её принять ухаживание.
Гиперболические изоклины рис.8 показывают разные уровни интенсивности стимуляции партнёра (и разные уровни эффективности поведения самца) в процессе ухаживания. Вдоль каждой изоклины разное соотношение вкладов составляющих гетерогенного сигнала обеспечивает одну общую интенсивность стимуляции, которая определяется как сигнальная сила, оцениваемая по вероятности и выраженности специфической реакции на неё (McFarland, Houston, 1981). Гиперболы изоклин сигнальной силы вместо прямых показывают частичное нарушение закона гетерогенной суммации в данном конкретном случае.
Вообще, когда речь идёт о демонстрациях — знаковых стимулах, их организованном обмене в процессе коммуникации, а не просто о визуальном образе компаньона, закон гетерогенной суммации отчасти нарушается. Целостный образ демонстрации как релизер эффективней, чем сумма её компонент, т.е. действие составляющих сложного сигнала не аддитивно, а мультипликативно.
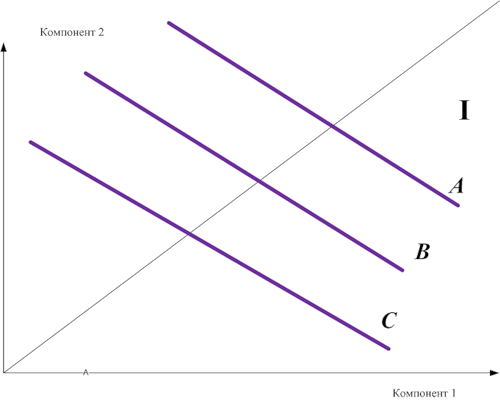
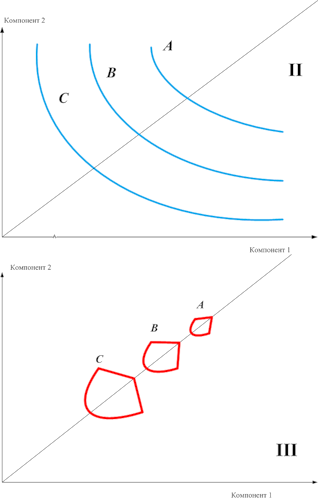
Строго аддитивное воздействие окраски и размеров яйца на запуск поведения закатывания у чаек можно представить общим индексом оценки яйца и изобразить в виде параллельных изоклин. Скажем, так как на схеме рисунка 9.I и рис. 9II-III; везде сигнальная сила стимула А выше, чем стимула В, В больше, чем С, комплексные стимулы А, В и С образованы одними и теми же компонентами 1 и 2 (абсцисса и ордината). Аналогичные изоклины сигнальной силы можно строить для любых комплексных стимулов, а их форма показывает, насколько точно выполняется закон гетерогенной суммации – наиболее точно, если изоклины представляют собой прямые (рис.9.I), менее точно в случае гиперболы (рис.9.II) и совсем не выполняется, если гипербола «редуцируется в точку» (рис.9.III).
Анализ рис.1-7 показывает, что для комплексных стимулов этот закон выполняется в точности, для знаковых стимулов — уже с нарушением: сигнальный эффект целого здесь уже больше суммы сигнального воздействия частей. Если демонстрация функционирует не только как стимул, но и как сигнал: не только подвергает партнёра специфической стимуляции, но и оказывает некий сигнальный эффект через специфический образ демонстрации, репрезентируемый партнёру, то закон гетерогенной суммации нарушается. Изоклины сигнальной силы при использовании знаковых стимулов уже будут гиперболическими, а не линейными (рис.9II).
 A.
A.
Б-В.
Рисунок 9. Зависимость соблюдения закона гетерогенной суммации от категории сигнальных средств, используемых для образования комплексного стимула. А/I. Используются специфические стимулы – закон соблюдается в точности. B/II. Используются знаковые стимулы – закон частично нарушается. C/III. Используются знаки — закон не соблюдается вовсе.
Степень нарушения прямо пропорциональна «сигнальности» и «знаковости» демонстраций. С ростом последней сигнальный эффект специфических образов демонстрации по значимости всё больше и больше превосходит «след» от воздействия демонстраций как стимулов (тем более что последний формируется медленней, чем доходит сигнал). В этом случае закон гетерогенной суммации нарушается больше и больше и, когда демонстрации из стимулов превращаются в произвольный знак, не выполняется вовсе, значащие единицы (инвариант формы соответствующих демонстраций) составляются из незначащих элементов, элементарных двигательных актов или других моделей движений. Для знаков – сигнальное воздействие частей равно нулю, они нужны только для образования целого, которое и осуществляет всё воздействие полностью (рис.3III).
Примечания
1Поэтому большинство коммуникативных сигналов инстинктивной природы мультимодальны – визуальные демонстрации соединяются с акустическими или, скажем, вибрационными, но связка нестрогая, чисто статистическая.
2 Zeitschrift für Tierpsychologie, с 1990х годов это журнал Ethology
3грубошëрстная померанская порода, бывшая и в Восточной Пруссии.
4Сейчас это мнение оставлено, сходство с енотами чисто конвергентное. Больше всего она родственна настоящим лисицам Vulpes spp.
5чтобы определить безусловно врождённые формы поведения, это так называемый каспар-хаузер подход.
Рекомендуем прочесть
 «Жесты», вόроны и шалашники
«Жесты», вόроны и шалашники Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей Вольфганг Шлейдт. Коммуникация животных. Поиски понимания сложности поведения
Вольфганг Шлейдт. Коммуникация животных. Поиски понимания сложности поведения «Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования
«Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования «Работа» релизеров в коммуникации животных: хороший стимул – плохой сигнал, и наоборот
«Работа» релизеров в коммуникации животных: хороший стимул – плохой сигнал, и наоборот- Опасности коммуникации
- Риск демонстрирования как подтверждение «сигнальности» демонстраций
