
Peter K. McGregor, ведущий исследователь «подглядывания» и коммуникативных сетей у позвоночных.
Резюме. Сравнительная этология основывается на двух главных концептах — реальность существования и существенность форм единиц поведения — демонстраций. Первое означает, что они дискретизированы в потоке поведения, обладают типологической определённостью формы, т.е. она специфична у каждой демонстрации, что делает её узнаваемой для самих особей и для внешнего наблюдателя в независимых актах повторного предъявления: она не условность, рождённая наблюдателем для удобства классификации, и не кажимость, созданная его категоризирующим вниманием. Поэтому формы единиц поведения могут быть выделены и описаны объективно, на чисто структурной основе, без неявного привнесения функциональных моментов в процедуру описания. Второе означает, что специфический результат коммуникации (образование пары, сближение в ней партнёров и спаривание, изгнание захватчика или, наоборот, захват периферии его территории после конфликта) достигается специфическими средствами, каждое из которых вполне определённым образом детерминирует последующие выборы поведения у другого участника (-ов) взаимодействия, как и у самого демонстратора, т.е. оно специализировано в определённом влиянии на него (-них). Эта специализация суть коммуникативная функция единиц поведения: каждое из подобных влияний специфично и статистически устойчиво, почему может быть установлено (а функция охарактеризована) на чисто функциональной основе без неявного привнесения структурных моментов (есть соответствующее влияние, резко меняющее поведение второй особи, или нет, безотносительно конкретных границ демонстраций во времени).
Критики сравнительной этологии, социобиологи на Западе и последователи Е.Н.Панова в нашей стране активно атакуют оба тезиса; по счастью, это везде попытки с негодными средствами. Реальность существования специфических единиц поведения, демонстраций, показана строго ещё в 1980е и многажды подтверждена после; их специализация как знаков или знаковых стимулов, участвующих в коммуникации и этим существенных для особей не меньше, а больше, чем для наблюдателя-этолога, поставленная под сомнение в годы концептуального кризиса сравнительной этологии (1970-80е гг.), постепенно подтверждалась всё больше в последующем, о чём я писал, и не раз (в том числе подтвердилась в анализе риска коммуникации — безусловном достижении критиков сравнительной этологии). Здесь рассказывается об исследования «подглядывания» и коммуникативных сетей в сообществах позвоночных, окончательно подтверждающих достижение специфического результата коммуникации специфическими средствами, опрокидывающем все построения критиков.
***
Независимым (и окончательным) подтверждением сделанного ранее вывода, что видоспецифические демонстрации — не просто релизеры, но образуют «язык» по Ф. де Соссюру1 выступает «подглядывание» (англ. eavesdropping behaviour, Johnstone, 2001). Оказывается, демонстрации и исходы взаимодействий, наступившие вследствие организованных обменов демонстрациями и действиями, т.е. коммуникации, исключительно привлекательны для «третьих особей», «зрителей» взаимодействия. Напрямую не вовлеченные в эти обмены, «зрители» в противоположность участникам отнюдь не подвержены «давлениям» мотивации изнутри и стимуляции извне. Однако они постоянно интересуются поведением и взаимодействиями соседей, концентрируясь на тех же моментах, что «содержательны» для этих последних — какие демонстрации предъявляются, как интенсивно, каковы исходы взаимодействий, наступающих вследствие обмена демонстрациями.
Это изучено нами у расселяющихся сеголеток больших пёстрых дятлов в августе-сентябре при закреплении в в складывающихся внегнездовых группировках2. Несведущие в его территориальной структуре, после прибытия (обычно под вечер) они не перемещаются как хотят, но «выявляют» эту последнюю «подглядыванием». Первые 2-3 ч, иногда сутки, они размещаются в пограничных зонах охраняемых территорий и крайне незаметно: держатся в глубине крон, почти не передвигаются, но интенсивно осматриваются, отслеживая маркировку пространства криком «кик» (тем более что она достигает одного из двух суточных пиков за 1,5-2 ч до захода в ночёвочные дупла), патрулирование границ, случающиеся конфликты и прочие формы территориальной активности владельцев3 . На это уходит 83.5% времени.
На «проходящих» особей вселенцы почти не обращают внимания: контакты с ними неизменно мирные, в августе-сентябре 2006 г. из 84 подобных случаев с 19 птицами отмечено только 2 стычки, в неожиданных встречах «нос к носу». Все зафиксированные (и редкие) передвижения «наблюдателя» происходили в места наиболее интенсивной маркировки или конфликтов, для лучшего их отслеживания. Ночуют новоприбывшие в слабее всего охраняемых частях поселения (из 23 прослеженных ночёвок 13 — на ветвях деревьев, 6 — начали выдалбливать ночёвочное дупло (на это уходит 2-4 дня), 4 — ночевали в дуплянке).
На следующий день сразу, без «проб и ошибок», вселенцы вторгаются именно к тем резидентам, кто «вообще» охраняет пространство менее успешно, чем прочие или чаще всего проигрывал конфликты вчера4. Место вторжения (на периферии территории резидента, избранного для этих претензий) дятлы маркируют «киканьем» 4-5 ч: 54,3% уходит на крики, 20,0% — на осматривания и перелёты, на кормление только 11,9%. Перелетая в пределах площади 40-50 м2, птица раскрывает полностью хвост и предъявляет 6ю пару рулевых новым соседям5. Это сигнал угрозы, точнее, владения данным пространством и максимальной готовности его защитить в территориальном конфликте6. Молодые всё это проделывают как можно ближе к центрам социальной активности оседлых членов поселения, найденным при «подглядывании»: гА = 0.94, Р < 0.01 (Фридман, 1996).
В разреженных поселениях (<16-20 особей/км2) владельцы не обращают внимания на эти претензии: сеголетка атакуют лишь при маркировке непосредственно рядом с их «кузницами» и ночёвочными дуплами. В плотных же поселениях (>20 и особенно >30 особей/км2) соседи вторгаются, инициируя конфликты: чем плотней, тем быстрей их реакция, а конфликты ожесточённей (обычно после раунда маркировки, но может её и прервать).
Птицы молча перелетают по веткам, демонстрируя друг другу крайние рулевые и вытягивая шею под углом 45°: «киканья» сеголетка — лишь если вторжение прервёт маркировку. Длительность подобных контактов 25-30 мин. После этого в 20% случаев оседлый дятел атакует вселенца, что в 80% случаев приводит к изгнанию последнего (п = 50). Чем меньше времени пробыл мигрант в данной точке, тем меньше вероятность её удержать (г = 0.828, Р < 0.05). Если прежние резиденты заметят сеголетка до начала этого раунда «маркировки с конфликтами», он изгоняется почти гарантированно, если он выстоит от их натиска, то включается в число резидентов (Фридман, 1996).
Самое важное, что «подглядывание» за территориальной активностью соседей, результаты которого структурируют одноимённую активность у «наблюдателя» у больших пёстрых дятлов равно присуще всем резидентам и активным вселенцам, но никогда не встречается у «проходящих» особей: те вполне безучастны и к рекламированию, и к территориальным конфликтам других дятлов. Напротив, первые две категории птиц, когда сами не участвуют во взаимодействиях, то активно отслеживают ход чужих социальных контактов, их исходы и/или использованные демонстрации. По результатам слежения они корректируют поведение в будущих взаимодействиях в сторону большей успешности демонстрирования (что зависит от последовательности выбора демонстраций, моментов их предъявления и смены — см. далее), выбирают «наилучшее время и место для них», и «наиболее удобного» противника. Поэтому чаще «подглядывающие» особи выигрывают территориальные конфликты и дистантные угрозы тем чаще, чем больше «подглядывают», а интенсивность последнего пропорциональна уровню территориальной активности данной особи, прежде всего маркировок и патрулирований границ, и частоте конфликтов её соседей7.
Границы территорий8 краснокрылого трупиала Agelaius phoeniceus меняются достаточно редко, длинные периоды стабильности чередуются с каскадами быстрых перераспределений пространства, охраняемого каждой особью в группировке, и конфигурации границ. Их «передел» как цепная реакция охватывает всё поселение именно потому, что территориальные самцы постоянно отслеживают перемещения и взаимодействия соседей, особенно ход и результат территориальных конфликтов, и вторгаются преимущественно к соседям, только что проигравшим конфликт, одновременно усиленно обороняя собственные границы от птиц, только что победивших противника9.
Опыты с подстановкой чучел самцов на участки краснокрылых трупиалов показали, что соседи много чаще (и успешней) вторгаются на участки особей, проявивших недостаточную агрессивность при атаке чучела или вовсе не реагирующих на данный сигнал. Если сосед-наблюдатель лишь недавно занял участок рядом с особью, подвергнутой эксперименту, то вторжения к ней следовали сразу же после тестирования, если «наблюдатель» занял соседнюю территорию уже давно, изменение границ происходило только на следующий день. Контроль — подстановка чучел восточного тиранна Tyrannus tyrannus, также в демонстративной позе с расправленными крыльями: красные “эполеты” на чучеле трупиала были расправлены и хорошо заметны (Freeman, 1987).

А.

Б.
Рисунок 1. А. Самцы краснокрылого трупиала угрожают друг другу в позе «клюв вверх». Б. Слабое выражение той же позы (готовность защищать это пространство от попыток захвата) у птицы, следящей с наблюдательного пункта (англ. perch site).
Перед экспериментами автор картировал территории по местам демонстрации «клюв вверх» (англ. bill-up или bill-tilting display10, рис.1): границы точно проводятся при одновременном ее предъявлении соседями друг другу (посредине между ними) и примерно в остальных случаях, посредине между ближайшими наблюдательными пунктами, если у каждой птицы их отметили ≥30. Чучела трупиала или тиранна подставляли через несколько недель активной защиты пространства, тестируя как постоянных владельцев данной территории, так и «заместителей», вытеснивших прежних владельцев: опыты прекратили с началом копуляций. При их предъявлении фиксировали наблюдательные пункты соседей или фокальной особи, оказывающиеся за уже установленными границами (т. е. последние ими уже нарушены ради лучшего наблюдения за происходящим). После тестирований картировали все наблюдательные пункты и места демонстраций «клюв вверх» в течение 30 дней, то и другое >0,5 м вглубь чужой территории считали вторжением, если то и другое не встречает сопротивления, границу считали переместившейся (Freeman, 1987).
У самцов-заместителей охраняемое пространство меняется легче: включённость в него конкретных точек фиксировали по двум подряд регистрациям «клюва вверх» или наблюдательным пунктам владельца, утром и вечером одного дня. Тот же критерий (две демонстрации подряд) применяли для картирования вторжений и передвижки границ в связи с тестированием.
В периоды стабильности частота побед в серии конфликтов статистически устойчива у всех резидентов и чётко коррелирует с другими показателями ранга, например, со степенью развития красных эполет самца или средним числом самок в гареме на его территории (Eckert, Weatherhead, 1987a-c). Поэтому победы одних и поражения других птиц значимы как «вторичные сигналы» для «зрителей» на соседних участках. Опытное и контрольное чучела ставили на 20 мин в центр территории, и наблюдали с дистанции в 40-50 м; фиксировали число ударов, нанесённых владельцем (максимум 15), и время, проведённое им на чучеле (максимум 19,25 мин). Обе характеристики территориальной агрессии связаны отрицательно (rs ~ -0,25): их суммированием получили индекс «энергичности охраны пространства» (табл.1), позволяющий сравнить 39 протестированных резидентов по способности монополизировать ресурсы в этом пространстве (англ. Resourse-holding potential).
Таблица 1. Краснокрылые трупиалы оценивают способность друг друга охранять территории, отслеживая энергичность атаки на чучело, и эти оценки влияют на их территориальное поведение в будущем.
|
Интенсивность атак на чучело |
I |
II |
||
|
N |
Y |
N |
Y |
|
|
Отсутствуют, контроль |
12 |
0 |
12 |
0 |
|
Ниже медианы, а |
9 |
3 |
6 |
6 |
|
Выше медианы, а |
13 |
2 |
13 |
2 |
|
Ниже медианы, б |
6 |
6 |
8 |
4 |
|
Выше медианы, б |
10 |
0 |
10 |
0 |
Обозначения: а — постоянные владельцы, б — «заместители», I — Были ли вторжения соседей при тестировании, Y да, N нет, II — были ли сдвиги границ территорий соседей «за счёт» пространства фокальной особи, Y да, N нет.
Чучело тиранна вообще не вызывало атак; после тестирования не было ни вторжений соседей к соответствующим самцам, ни перемещений границ с ними (табл.1). Энергичность атак на чучело трупиала варьировала от 0 (не обращали внимания) до 1,21: резиденты значимо не отличались от «заместителей» ни по средней энергии атак, ни по доле птиц с ненулевым индексом. Высокая энергичность атак на чучело трупиала страхует от вторжений соседей во время опыта и от передвижения границ (не в пользу владельца) после него, низкая — наоборот, провоцирует эти риски для постоянных владельцев и «заместителей» (табл.1). Следовательно, все соседи отслеживают конфликты друг друга (Freeman,1987), фиксируясь на позе «клюв вверх» и других значащих демонстрациях.
Наблюдения за территориальным и песенным поведением меченых радиопередатчиками самцов южного соловья Luscinia megarhynchos показали, что они отслеживают территориальные конфликты соседей, обращая особое внимание на песенные блоки, маркирующие у них ситуации отпора захватчикам11. В опытах с проигрыванием записей песен установили использование владельцами территорий блоков песен соседей наравне с собственными для отпора захватчикам. В опытах их привлекали записью «песенной дуэли» соседа с третьей особью, те вторгались туда, позволяя фиксировать ответы захватчиков на разные единицы владельца. Самый интенсивный ответ был на блоки, уже встречавшиеся в ситуации прошлых отпоров захватчикам, но лишь при его успешности, когда захватчик смолкал, убирался и пел уже на своей территории. Самцы активно вторгались на чужие территории при исполнении там «победоносных» блоков владельца, но никогда — блоков из песни «поверженного» захватчика (Naguib et al., 2004).
Далее «наблюдатели» копируют «победоносные» блоки соседей: они звучат в их собственной песне уже при охране своей территории от подобных захватов. Копирования не происходит, если резидент, за которым «подглядывают» наблюдатели, проигрывает (рис.2). Следовательно, территориальные самцы южного соловья используют структуру пения соседей как «систему раннего предупреждения» о возможных нарушениях сложившейся территориальной структуры группировки захватчиками, а отдельные блоки, связанные с высокой вероятностью победы, — для увеличения собственной способности противостояния этим рискам (Naguib et al., 2004).
А.
Б.
В.
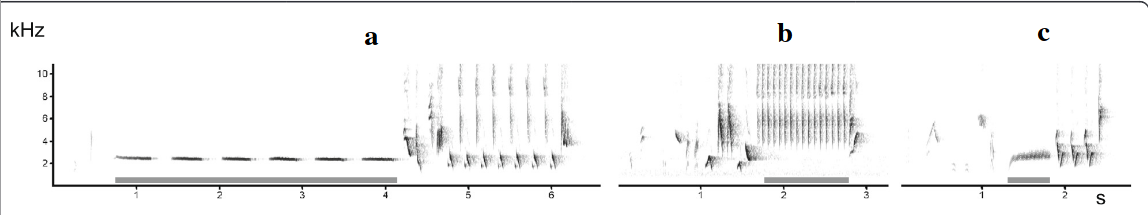
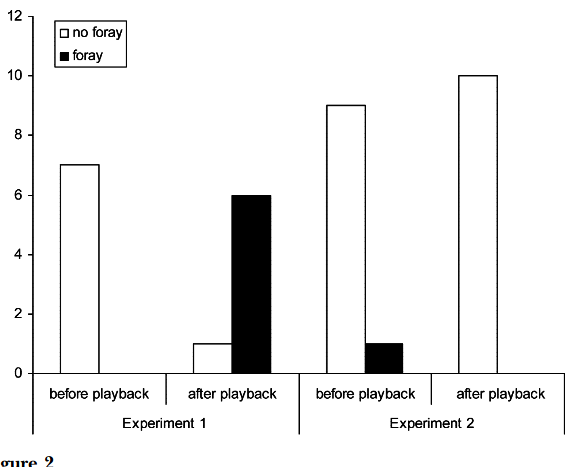
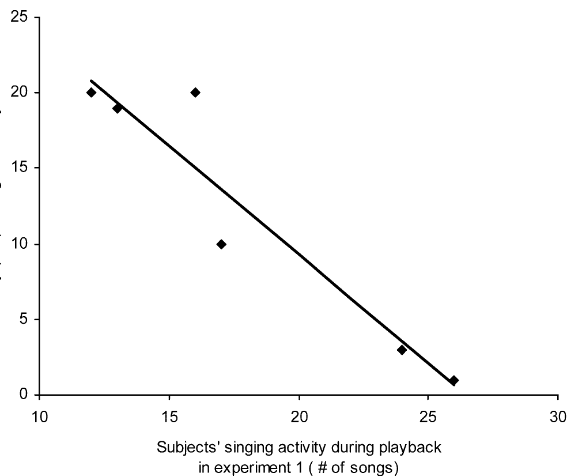
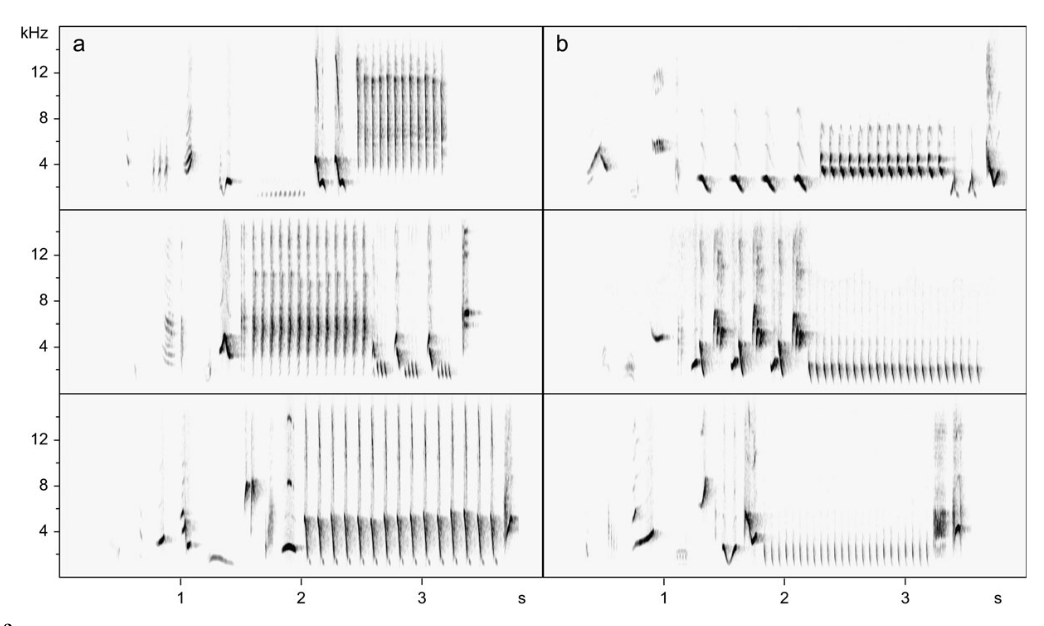
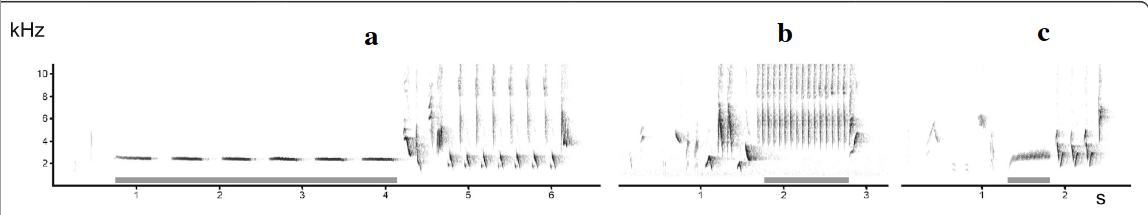
Рисунок 2. «Подглядывание» во взаимодействиях самцов южного соловья. А. Примеры песенных блоков: даны их спектрограммы, серое — образующие их акустические формы. Обозначения. а: свистовая песня — гомотипическая серия вокализаций в узком диапазоне частот почти без частотной модуляции; б: трелевая песня: быстрое повторение вокализаций в широком диапазоне частот при незначительной частотной модуляции; c. жужжащая песня — образована длинным, неповторяющимся жужжанием, создаваемым очень быстрым повторением вокализаций в узком частотном диапазоне на достаточно низких частотах. Б. Число радиопрослеживаемых соседей, вторгнувшихся или нет на фокальную территорию (n=19), белые столбики — число не вторгшихся, чёрные — число вторгшихся. Эксперимент 1 — c 26.04 по 9.05.2002 г., эксперимент 2 — с 26 по 29.04, самцы занимают территории уже несколько дней, на большинстве из них уже поселились самки. Эксперимент 2 имел 2 части: сперва, как в эксперименте 1, песню проигрывали на чужой территории, имитируя вторжение соседей, вторая же часть имитировала отступление захватчика с пением на своей территории: через 2 мин спустя проигрывания на фокальной территории данную песню повторяли как за её пределами, так и за пределами песен соседей, чьё вторжение отслеживалось. Обратите внимание, в этом случае вторжений соседей не происходит, в отличие от предыдущих. В. Чем выше интенсивность пения в проигрываемых записей (абсцисса, № песен в эксперименте 1), тем позже вторгаются соседи на эту территорию (ордината, мин). Источники: А. — Bartsch et al., 2015, Б. — В. — Naguib et al., 2004.
Последующие работы начали выявление сигналов, на основе которых корректируют поведение «подглядывающие» самцы (Schmidt et al., 2004). Различия в конкурентоспособности маркируются большим или меньшим присутствием в песне быстрых широкополосных трелей (рис.3). Их значимо больше в песнях, направленно адресованных оппонентам в ближнем взаимодействии самцов (например, при вторжении на чужие территории), чем в случае пения днём или ночью «в пространство».
Самцам, далее различавшимся по успеху размножения, проигрывали записи песен из двух динамиков, находящихся в 32 м друг от друга, они как бы слышали песенную дуэль двух самцов. «Пение» одного содержало 10 песен с присутствием трелей, чередующихся с 10 песнями без трелей в случайном порядке, второго — те же 20 песен вовсе без трелей. Дальше, используя радиопрослеживание, фиксировали, какой из динамиков вызовет больше вторжений соседей (за 30 мин после проигрывания в сравнении с 30 мин до), как скоро они подойдут поближе, как интенсивно будут петь и пр.
А.
Б.
В.
|
Переменные |
I |
II |
|
|
2) |
115±28 c |
272±45 c |
-0,43 |
|
3) |
273±44 c |
618±55 c |
-0,9 |
|
1) |
4±1 м |
12±2 м |
-0,85 |
|
5) |
64±12 |
35±10 |
0,85 |
|
4) |
30±10 |
11±5 |
0,82 |
Г.
Д.
Е.
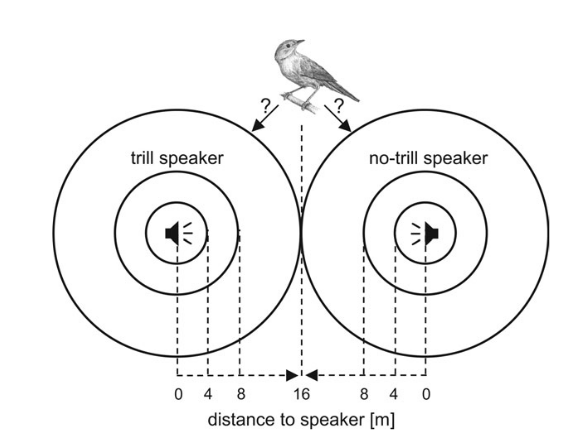
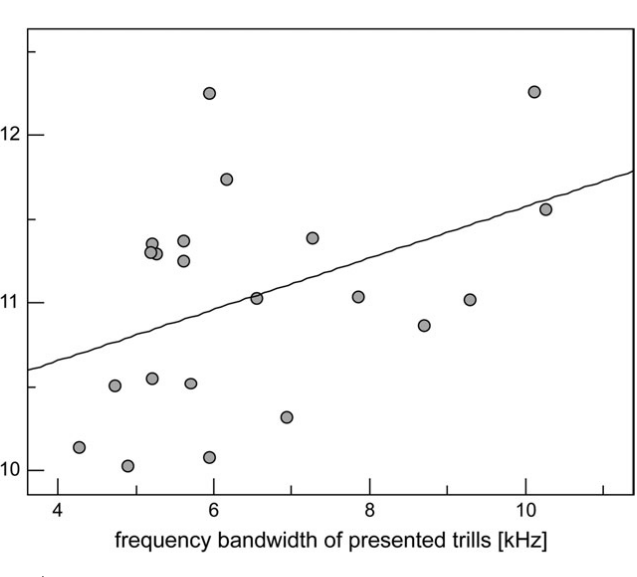
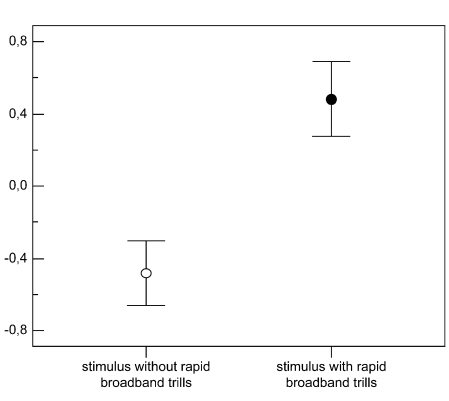
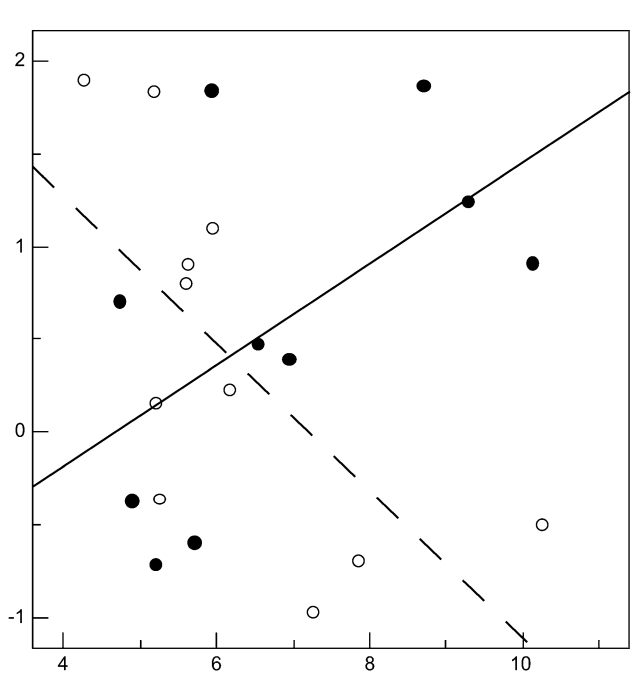
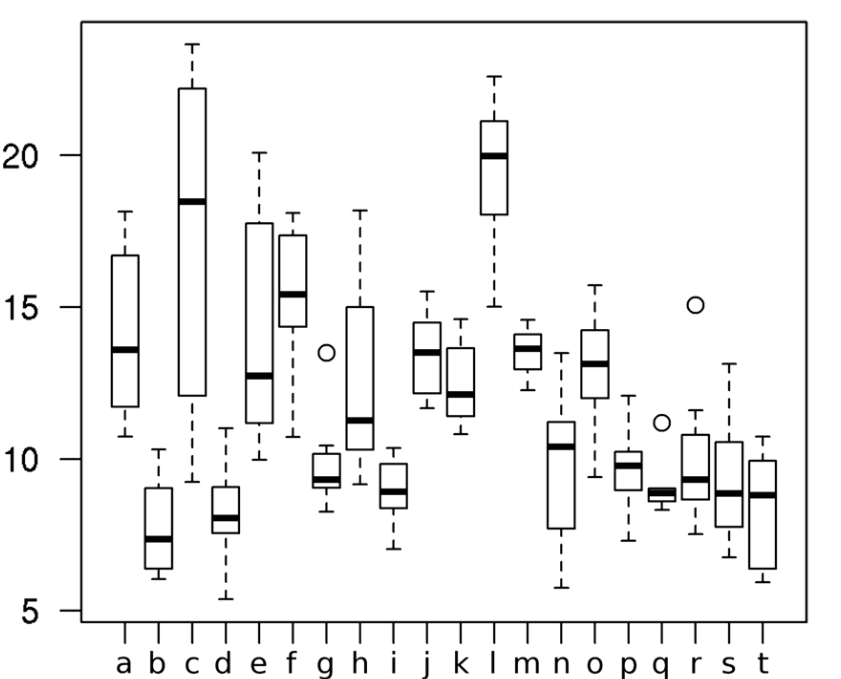
Рисунок 3. Обилие быстрых широкополосных трелей в песнях маркирует уровень территориальной агрессии самцов южного соловья. Обозначения. А. Радиопрослеживаемые самцы выбирают между двумя динамиками с разным составом песен. Пунктир — расстояния до соответствующего динамика. Б. Разные типы песен южного соловья, различающиеся частотой повторения быстрых широкополосных трелей (а), и шириной диапазона частот, в котором звучат последние. Их легко отличить от других типов трелей (b): а звучат намного быстрей (>7 элементов/с) и в большем диапазоне частот (>3 кГц). В. Ширина частотного диапазоне трелей (абсцисса, кГц) позитивно связана со скоростью исполнения трелей в песнии конкретных самцов (ордината, элементов/с), правда, только на уровне тенденции, r = 0,42, n = 21, P = 0,057. Г. Различия в интенсивности ответа самцов на пение с трелями (I) и без них (II), 1)-5) см. текст. Д. Ответ на пение с трелями (слева) значимо интенсивней (ось У, координаты по оси главных компонент). Меньшие координаты показывают скорейшее приближение и более тесное сближение с динамиком. Е. Абсцисса — ширина частотного диапазона тестовых песен, кГц, ордината — интенсивность ответа радиопрослеживаемых самцов: полые кружки и пунктир — неудачников, не сумевших спариться, залитые кружки и линия — успешно приобретших самку. Источник: Schmidt et al., 2008.
Всего тестировали 21 самца. Интенсивность ответа оценивали 5 характеристиками: 1) наибольшее приближение песенных постов вторгнувшихся соседей к каждому динамику, м; 2) время задержки до наибольшего приближения, с; 3) время задержки до приближения с пением в радиусе ≤4 м вокруг каждого из динамиков; 4) число песен, исполненных в радиусе ≤4 м; 5) число песен, исполненных с каждой стороны, т. е. в радиусе ≤16 м. Анализ главных компонент показал объяснимость 62% дисперсии ответа самцов координатами по первой оси (РС1): у параметров скорости ответа на проигрывание и тесноты сближения соседей с динамиком они отрицательные, у числа спетых песен положительные (Schmidt et al., 2008).
Общая интенсивность ответа резидентов значимо больше на пение с трелями, чем без них (рис.2Д), здесь нет различий между соловьями, дальше успешно нашедшими самку, и «неудачниками», так и не образовавшими пару (дисперсионный анализ, F1,18 = 0,016, P = 0,90). Не различается и реакция этих двух групп самцов на частоту повторения трелей в тестовых песнях (F2,15 = 0.7, P = 0.5), при чётких различиях в интенсивности реакции на проигрывание песен с трелями, зависевшей от тонкой структуры последних (F1,17 = 9.1, P = 0.008). Значимая характеристика здесь — ширина частотного диапазона трелей: при его расширении интенсивность ответа неудачников значимо уменьшается, но растёт у самцов, успешно приобретших самку (r = 20.67, n = 11, P = 0.024 vs r = 0.56, n = 10, P = 0.094).
Мы видим, что тонкая структура песен кодирует информацию о «качестве» самца в контексте охраны территории, её направленно воспринимают владельцы соседних участков через «подглядывание» (вместе с «подслушиванием») и на её основании корректируют собственную территориальную активность (Schmidt et al., 2008). В точности то же самое осуществляют самки, оценивающие самцов как потенциальных партнёров во время ночного пения. Анализ поведения 10 радиопрослеживаемых самок L.megarhynchos, отловленных за 70 км и выпущенных в новой местности в начале образования пар, показывает интенсивные ночные «экскурсии» на участки разных самцов12, контрастирующие с дневным пребыванием на одном месте. Посещая участки разных самцов (до 6), они выбирают потенциального партнёра на основе «подслушивания» песен: самца останавливается на одном из участков и дальше сближается с владельцем, образуя пару (Roth et al., 200913).
А.
Б.
В.
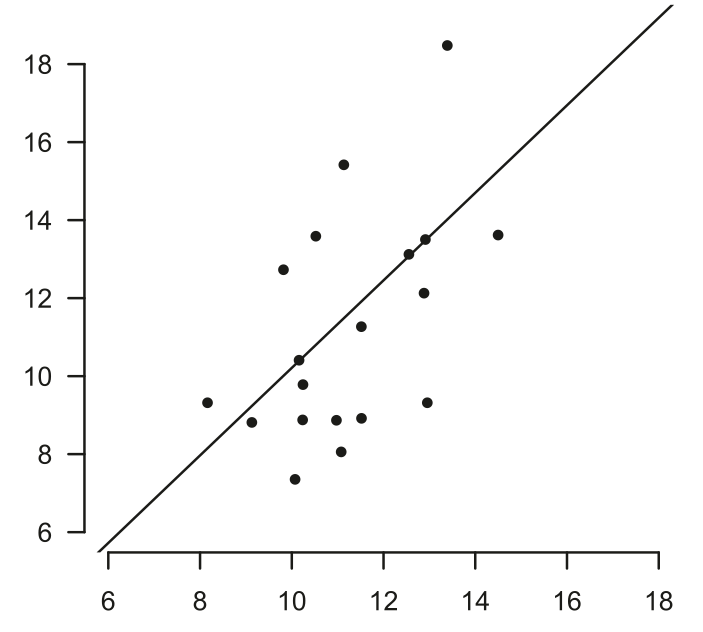
Рисунок. 3. Песенная структура как предиктор отцовских качеств самцов южного соловья. А. Различные типы песен, составляющие самцовый репертуар: серым подчёркнуты характерные для них акустические паттерны. Обозначения. (a): свистовая песня: серия повторяющихся элементов, узкополосная, с малой частотной модуляцией или совсем без неё. (b): трель: быстрая, широкополосная трель из повторяющихся элементов с незначительной частотной модуляцией (c): шумная песня из длинного, неповторяющегося жужжания, созданного очень быстрым повторением вокализаций с низкой частотой и в узком диапазоне частот. Б. Разнокачественность самцов (n=20, каждый столбец — отдельная особь) по интенсивности кормления птенцов (ордината, посещений гнезда/ч). Межсамцовая вариабельность параметра больше, чем изменчивость интенсивности у каждого самца (диаграммы с усами — показаны медианы, 25% и 75% квартили, выбросы). В. Позитивная связь частоты кормления птенцов, предсказанной по песенным характеристикам (абсцисса), с реальной частотой (ордината), для 20 изученных соловьёв. rs = 0,54, P = 0,014). Источник: Bartsch et al., 2015.
Вклад самцов L.megarhynchos в выкармливание примерно равен вкладу самки, интенсивность кормления — их персональная характеристика, не менявшаяся за время наблюдений (ANOVA, n = 20, F = 12,46, P < 0,001). Песенные характеристики — маркёр этой разнокачественности, в изученной популяции весьма значительной (рис.3Б), так что наблюдаемая у них интенсивность кормления птенцов весьма точно соответствует предсказанной (рис.3В). Анализ показывает передачу этого рода информации 4 аспектами сложности песни: средняя длина самого короткого колена (число вокализаций до переключения на другие колена), а также величина репертуара шумных песен, свистовых песен и трелей. Иными словами, лучшие отцы — соловьи, многократно повторяющие один и тот же тип песни, и с более обширным репертуаром каждого такого типа. Их связь с интенсивностью кормления птенцов описывается регрессией с коэффициентами (в порядке снижения) 0,75, 0,46, 0,14 и 0,005 (ранговая корреляция Спирмена rs>0; P<0,05 для n=20) (Bartsch et al., 2015).
Непрерывное слежение компетентных участников коммуникации за всеми процессами и результатами общения соседей, точность «прогнозирования» их территориальных реакций и корректировки собственных демонстраций по поведению для лучшего противостояния «риску вторжения» со стороны победителей14 лучше отражает специфическую территориальную мотивацию животного, чем его собственное участие в конфликтах и экспрессия исполнения им демонстраций. Резко повышенное внимание к «чужим» взаимодействиям, его строгая направленность на сигнальные элементы в потоках соседской активности15 заставляет считать это внимание коммуникативным, а поведение, реализуемое после «подглядываний» — следствием специфического эффекта сигналов, корректирующего прежнюю модель поведения «наблюдателя».
«Подглядывание» и коммуникативная сеть в полной мере присутствуют во взаимодействиях с компаньонами других видов, например, при спасении жизни от хищников. У краснокрылых трупиалов (Agelaius phoeniceus) самцы-владельцы территорий весь сезон размножения издают сигналы предупреждения об опасности с частотой >4-5/мин, в пределе до 40/мин. Обычно они в каждый данный момент издают один и тот же сигнал; если в ответ на опасность какой-то самец резко сменит его, соседи сразу же переходят на новый вариант, чем ретранслируют сигнал по району перемещения возможной опасности. Вывод, что крики тревоги сигнализируют самкам на гнёздах именно о невидимой для них опасности, не об уровне возбуждения или беспокойства самого самца, поддерживается тремя независимыми рядами фактов (Beletsky, 1989):
1) Лишь владельцы участков слушают сигналы друг друга и немедля копируют смену типа вокализации. Нетерриториальные самцы, не включённые в коммуникативную сеть группировки, издают ровно те же сигналы, но только столкнувшись с опасностью напрямую;
2) Резко различны реакции самок на проигрывание повторяющихся и сменяющихся сигналов самца: лишь во втором случае они беспокоились и оставались у гнёзд.
3) Бдительность самок растёт при отлучках самцов с территории (отслеживаемых по отсутствию повторяющихся сигналов поблизости), и снижается в их присутствии (Beletsky, 1989).
Полевые опыты показали кодирование дистанций до хищника или его опасности темпом издавания криков. Для трупиалов, гнездящихся в густых зарослях, то и другое во многом взаимозаменяемо: разная степень опасности от потенциальных хищников требует разных дистанций начала тревоги, но не «запуска» разных реакций беспокойства, отведения или бегства. Птицам предъявляли чучела ондатры, ястреба, виргинского филина и сороки на разном расстоянии от гнездовой территории, а также фиксировали темп вокализации в ответ на приближение наблюдателя. В обоих случаях темп тревожной вокализации обратно пропорционален дистанции до возможной угрозы, падение интенсивности подачи крика с ростом дистанции зависело от степени опасности (Beletsky, 1991).
Здесь повторяющийся сигнал тревоги информирует партнёра о наличии самца и о среднем уровне его бдительности, позволяя самке выбрать оптимальную частоту собственных осматриваний. А вот смена его типа сигнала — знак потенциальной опасности; волны его ретрансляции по поселению показывают направление и расстояние до неё (Beletsky, 1991).
В отличие от социального общения, в контексте защиты от хищников животные часто отслеживают крики тревоги других видов и иную активность, специфически маркирующую опасности, и ориентируются по ним не хуже, чем по видовым сигналам.
Затраты времени на «подглядывание» довольно значительны: оно интенсифицируется при тревоги и стрессе — следствиях потери контроля над ситуацией. Всякое неожиданное приближение компаньона (хищника, угрожающего безопасности, захватчика, претендующего на часть территории, срыв сближения потенциальных партнёров из-за неожиданной агрессии самки и пр.) — признак критической нехватки информации, необходимой для устойчивой жизни и размножения на данном участке, симптом неадекватности поведенческой модели, реализуемой здесь и сейчас.
Благодаря непрерывной системе «подглядываний» все особи в группировке16 связаны в сквозную «коммуникативную сеть» (англ. communication network, McGregor, Peak, 2000; McGregor et al., 2000). Она информирует всех «включенных» в нее о существенных событиях, случившихся в общем пространстве группировки: типах опасных объектов, замеченных одними особями, классах территориальных конфликтов, разрешенных другими etc. (McGregor, Peak, 2000; McGregor et al., 2000). Коммуникативные сети существуют в форме сигналов, их «материальных носителей» — демонстраций, и правил информационного обмена «всех со всеми», опосредованного обменом демонстраций во взаимодействиях (McGregor, 2005; Sih et al., 2009). Они позволяют всем особям, «подключенным» к «сети», эффективно корректировать поведение по событиям, в которые те непосредственно не вовлечены, а часто даже сами не наблюдали, и которые не могли напрямую влиять на их мотивационное состояние.
Вовлекаясь в социальное общение, индивиды «автоматически» функционируют как «узлы» (приемники и передатчики сигнала) в системе обмена сигналами и/или стимулами с положительной обратной связью, из которой рождается коммуникативная сеть. Совокупности определенного типа «узлов», т.е. особей, связанных однородным процессом коммуникации в системное единство (ток, колонию, выводок, группу территорий, охраняемых гнездовыми парами или одиночными особями и пр., в зависимости от форм социальности данного вида в данных обстоятельствах) точно соответствует смыслу лоренцевского понятия der Kumpan.
По данным отслеживаний будущая активность птиц-членов-«сети» (с кем из соседей инициировать взаимодействие, какими демонстрациями его «открыть» и т.п.) корректируется так, чтобы максимизировать вероятность выигрыша при следующих территориальных конфликтах — как в попытках территориальных захватов, так и в обороне от них (Beletski, Oriance, 1987).
Резюмируя: коммуникативные сети, поддерживаемые «подглядыванием», фиксируются везде, где выигрыш взаимодействия зависит от выбора поведения в проблемной ситуации, независимо от его масштабов (от отдельных актов до стратегии поведения в целом). Они неизменно присутствуют в социальном общении позвоночных от костных рыб до птиц и млекопитающих (Dablesteen, 2005; Peak, 2005; Sih et al., 200917): «зрители» по данным «подглядывания» корректируют модель поведения, чем увеличивают его успешность в будущих социальных контактах с обоими участниками отслеженного взаимодействия или (несколько менее) с прочими особями в сообществе18. См. исследования охраны территорий у чернополосой цихлазомы Cichlasoma nigrofasciatum (Keeley, Grant, 1993), бойцовой рыбки Betta splendens (Doutrelant, McGregor, 2000), зелёного меченосца Xiphophorus helleri (Earley, Dugatkin, 2002); ящериц Lacerta saxicola (Целлариус, Целлариус, 2006)); или системы агрессивного доминирования сосновых соек Gymnorhinus cyanocephalus (Paz-y-Mino et al., 2004); у хаплохромисов Бартона Astatotilapia burtoni (Grosenik et al., 2007), во время ухаживания бурых воловьих птиц Molothrus ater (Yokel, Rothstein, 1991; West, King, 1988, 1990).
Везде относительная способность побеждать в стычках (или выигрывать взаимодействия других типов) фиксируется в различии демонстраций победившей и побеждённой особи19; по мере накопления «индивидуальной истории» побед и поражений, всё более различающихся у разных особей в сообществе: по этой причине один и тот же уровень территориальной агрессии разные резиденты «выразят» разными демонстрациями и различно используют их в будущих взаимодействиях с соседями. Разнокачественность индивидов по способности выигрывать взаимодействия (брачные ли, территориальные и т. д.) фактически визуализируется в демонстрациях, делаясь явной для наблюдателя. Он же автоматически (инстинктивно) реагирует на демонстрации, выбирая из особей сообщества наиболее подходящих партнёров для [будущих] стычек, ухаживаний etc., из видового репертуара — те демонстрации, что воздействуют на них эффективней всего.
Так, анализ роли «подглядывания» в агрессии у цихлид Cichlasoma nigrofasciatum20 показывает, что разные телодвижения участников конфликта передают разные «сорта» информации о возможностях действия по отношении к данному оппоненту (какие демонстрации использовать и в какой последовательности для максимизации вероятности победы). Одни демонстрации (lateral display) показывают относительный размер рыбки и, соответственно, развиваемую ею «мощность поведения», другие (noncontact display) — относительную способность бороться (Keely, Grant, 1993). Обе характеристики связаны с эффективностью участия в обмене демонстрациями, точностью и адекватностью ответа на демонстрации партнёра, позволяющие обогнать его в том «повышении планки» ритуализированной угрозы, что обеспечивает победу в конфликте.

Пара чернополосых цихлазом самец справа
Дальше на основании сигнальной информации «зрители» адекватно модифицируют собственное поведение (именно и только угрожающие демонстрации), увеличивая эффективность участия в агонистических взаимодействиях с теми особями, которых они наблюдали и со «средним» членом сообщества в целом. Иными словами, животное не само, собственным мозгом с анализаторами, воспринимает ситуацию решает проблемную задачу и делает вывод. Хотя для стороннего наблюдателя всё выглядит именно так, на деле животные действуют автоматически, как модем, считывающий информацию, и в соответствии с ней меняющий конфигурацию сигнала в обслуживаемой им компьютерной сети.
Здесь более чем уместна и весьма эвристична метафора машины Тьюринга: абстрактной вычислительной машины, созданной, чтобы показать возможность разложения любой интеллектуальной или когнитивной операции на ряд простейших и чисто механических действий. Верно и обратное: осуществимы устройства, демонстрирующие понимание сказанного или выдачу адекватных ответов так точно, что в некотором испытании («тест Тьюринга») будут неотличимы от «настоящего» разума, хотя действуют «чисто механически», без всякого понимания [и/или прочувствования, понимание бывает и эмоциональное]21. Как машина Тьюринга симулирует элементарные операции логического вывода, хорошо описано в книге С.Пинкера «Язык как инстинкт» (1994, с.62-63).
Так порождается парадокс машины Тьюринга: зафиксировав эффективную коммуникацию = адекватное реагирование агента Х на сигнал + эффективный выбор действий на основании сигнальной информации на всём протяжении процесса взаимодействия, невозможно определить, сознательно действует Х или нет. Даже у людей, не говоря про животных, вывод о сознательности участников коммуникации обречён навсегда остаться гипотезой, не станет «теорией сознания» твоих компаньонов, также как выводы об их коммуникативной компетентности, о «сигнальности» и «знаковости» демонстраций и пр. Если эти гипотезы имеют тенденцию подтверждаться в последующих актах взаимодействия, и тем самым увеличивают готовность восприятия демонстраций как знаков – предикторов последующих событий, взаимодействие развивается по определённой траектории, если нет – останавливается и обрывается.
Проблема Тьюринга ясно показывает, что даже у людей, следующих презумпции намеренности и осмысленности всяческих действий, тем более человеческих, непосредственное общение душ невозможно: анализируя поведение партнёров по коммуникации, мы всегда оцениваем не сознательность действий (о ней лишь догадываемся, как и о планах, намерениях, представлениях говорящего), а компетентность владения специфическим языком (в смысле де Соссюра), поддерживающем данный поток общения.
Поэтому как видовые инстинкты всегда социальны и реализуются в коммуникации между индивидами22, «подглядывание», поддерживающее коммуникативную сеть — интегральная часть инстинктивного поведения позвоночных. И наоборот, представления о реализации инстинкта в организованном обмене демонстрациями — социальной коммуникации — стоит основывать не на индивидах, их восприятии ситуации и отреагировании стимулов от других индивидов, но на связывающих их системах (социальные связи, коммуникативная сеть, etc.), и информации, передаваемой сигалами в этих последних. Участники коммуникации, демонстрирующие друг перед другом, и этим ретранслирующие сигналы системы, вкупе с «подглядывающими» за ними «зрителями» коммуникации, в известном смысле представляют собой живые машины Тьюринга (Фридман, 2013).
Видимо, это всеобщее явление, моделируемое в работах Hsu et al., 2006; Sih et al., 2009; неслучайно структуры переднего и среднего мозга, поддерживающие «сеть», консервативны в ряду позвоночных, как и функционирующие там медиаторы (Goodson, 2005). Больше того, степень (и успешность) коррекции собственного поведения по предъявляемым формам демонстраций у «зрителей» взаимодействий сравнима с таковой их непосредственных участников (Фридман, 2017аб).
В норме животное не ждёт появления перечисленных проблем, но активно и регулярно обследует пространство активности, отслеживая поведение компаньонов, концентрируясь не столько на них самих, сколько на вторичных сигналах: социальных взаимодействиях, их демонстрациях и исходах, криках, пении, других следах активности (особенно применительно к другим видам, хищникам или конкурентам). Животные явно исследуют ситуацию и через «подглядывание» извлекают информацию, нужную для реструктурирования собственного поведения, решения вставшей перед ними проблемы. При этом они держатся максимально незаметно, их собственные демонстрации при максимуме концентрации внимания на компаньонах, особенно их демонстрациях, перемещения обеспечивают непрерывность слежения и точность фиксации происходящего.

Короткохвост (короткохвостый сцинк)
У резидентных особей короткохвоста Tiliqua rugosa неожиданное беспокойство от наблюдателя (взятие в руки, обработка в руках, сам факт наблюдения за ящерицей, пугающейся сотрясением почвы и тенью) резко увеличивает частоту рекогносцировочных перемещений по участку в следующие полчаса-час (рис.4). Эффект наиболее выражен у самых активных и разогретых самцов, с максимальной температурой тела (Kerr et al., 2004).
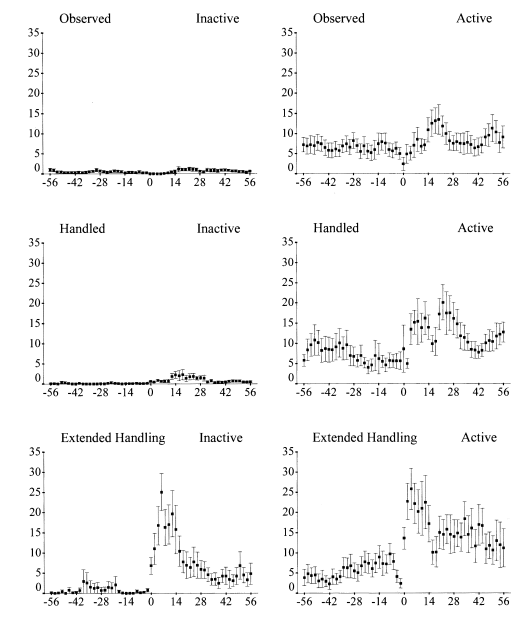
Рисунок 4. Всплески исследовательского поведения активных (справа) и неактивных (слева) короткохвостов после разных способов беспокойства (сверху вниз — наблюдение за сцинком, взятие в руки, обработка в руках).
Обозначения. Ордината — интенсивность обследований, число шагов в двухминутных интервалах за 60 мин до и 60 после беспокойства (точка 0). Показаны средняя SE, n=10.
Источник: Kerr et al., 2004.
В чём эволюционный смысл затрат на «подглядывание», а иногда даже риска при «подключении» к коммуникативной сети, и особенно при ретрансляции сигналов? Единственно возможное объяснение — для получения информации, переданной специфическими формами демонстраций, с последующим использованием её для корректировки моделей собственного поведения в будущих взаимодействиях, особенно в будущих противостояниях отслеживаемым индивидам. Первое столь же (или более) адекватно, что у самих участников взаимодействия, второе сравнимо по эффективности с поведением особи, выигравшей взаимодействие.
Реализация инстинкта — «автоматический» выбор действий на основании сигнальной информации разрешает возникшую проблемную ситуацию лучше (во всех смыслах — точность выбора поведения, действенность выбранного, затраты и безопасность, важные для животных много более, чем для нас), чем собственное слежение особей за развитием той же ситуации и собственный выбор действий вне инстинктивных форм поведения данного вида23. В силу этой «автоматичности» и, безусловно, «сигнальности» или «знаковости» демонстраций, «подглядываемых» соседями, сеть обеспечивает дальнодействие сигналов и событий в системе-социум, выигрышное для всех «включившихся» в коммуникативную сеть, поддерживающих её собственной активностью, скорректированной по данным «подглядываний», тратящих силы и время на ретрансляцию вторичных сигналов о ходе и результате прошедших взаимодействий (а иногда и рискующих в этой связи).
Восприняв сигналы, циркулирующие по «сети», животные, вовлеченные в проблемную ситуацию того типа, о котором сообщает сигнал (контакт с хищником, обнаружение корма, конфликт с потенциальным захватчиком, привлечение потенциального партнера и пр.), могут действовать специфическим образом сразу, «запустив» инстинктивное реагирование на сигнал определенным действием в сочетании с ретрансляцией сигнала дальше. Они «не отвлекаются» на собственное наблюдение за ситуацией и собственные прогнозы будущего развития событий, не загружают этим собственные анализаторы и интеллект, а используют «автоматику» инстинктивного реагирования; однако благодаря сигнальной информации, «автоматичные» действия оказываются точными, адекватными и опережающими.
С точки зрения социобиологии всё это — безусловный альтруизм24, однако реализованный вопреки социобиологическим теориям (где «сети» невозможны или редки из-за «обмана», буде не скреплены отбором родичей или дороговизной коммуникации) и выступающий их мощным фальсификатором. Это противоречие незамечаемо больше десятилетия, отражая парадигмальность науки «вообще» и необходимость смены парадигмы в нашей конкретной дисциплине, именно восстановления сравнительной этологии «на повышенном основании».
Коммуникативные сети в сообществе животных вполне изоморфны рынкам с их специфическими сигналами (цены, курсы акций, решения участников, специально доводимые «до общего сведения»), в которые включены и в которых работают продающие и покупающие. Как и в сообществах позвоночных, они поддерживают рынок своей активностью даже если на данном временном интервале это в убыток; практически повсеместно они одушевляют и персонализируют рынок (границы которого неясны каждому из включённых участников, иногда и исследовтелям), воспринимая и обсуждая его как существо с целеполаганием, намерениями и т. д., которые хочется выяснить25.
Исследование настороженности пушистопёрых дятлов (факт 21) показывает, что включённость в коммуникативную сеть, внутри- или межвидовую, даёт крупную «экономию» на реакциях оборонительного характера (осматривание, окрикивание потенциальной опасности)+выигрыш от большей успешности кормодобывания. «Экономия» достигается за счет того, что у поведение самообеспечения таких особей (не только кормление, но и отдых и чистка) прерывается приемом-передачей сигналов и/или сериями осматриваний только там и тогда, где и когда это необходимо, как в цитированных работах по настороженности D.pubescens.
Это верно для всех контекстов общения; ухаживания и угрозы не меньше, чем предупреждения об опасности. Скажем, «подслушивание» птенцами друг друга и родителей у сипух минимизирует конфликты. Совятам сперва проигрывали обмен криками между птенцами, постарше или помладше, один инициировал, второй немедленно отвечал, показывая больший уровень голода. Прослушивая диалоги, совята определяли возрастную иерархию и, когда во второй раз им проигрывали крики каждой «модели» отдельно, отвечали, бросая вызов, во-первых, птенцу, бывшему инициатором (именно он «угроза»), во-вторых, птенцу помоложе (легче победить) (Dreiss et al., 2013).
Или «подслушивание» позывок «вэд» самцов серой славки Sylvia communis, следующих непосредственно за образованием пар в контексте близкого взаимодействия партнеров, побуждает других самцов удаляться от уже «занятой» самки; напротив, ухаживательные песни самца их привлекают. Издавание позывок «вэд» также препятствует вторжениям других самцов, могущих стать «помехой» ухаживанию в паре, или, как минимум, тормозит их, см. результат опытов по проигрыванию обоих сигналов (Balsby, Dablesteen, 2005).
И эти (немалые!) выгоды доступны всем «подключившимся» к коммуникативной сети и «оплачивающим» членство в ней стереотипным воспроизведением форм видовых демонстраций в собственных телодвижениях, «должных» элементов репертуара в «должные» моменты взаимодействий с сочленами и активными вселенцами (McGregor, 2005). А это не только рискованно, но и затратно, часто идет на пределе мышечных сил, энергетических и метаболических возможностей организма, см. данные по манакинам Manacus vitellinus (Fusani et al., 2007; Barskie et al., 2011), райским птицам Lophorina surerba (Frith, Frith, 1988) и пр.); затраты и риск, как мы знаем, линейно связаны с эффективностью демонстраций.
Из всего этого «честность» сигналов с устойчивостью циркуляции информации по коммуникативной сети следует автоматически. Вполне предсказуемо оказывается, что в сообществе животные «доверяют» тому и другому не меньше, чем каждому отдельному сигналу. Они необратимо «подключаются» к коммуникативной сети и заранее «инвестируют» в ее поддержание, т.е. ориентированы на выигрыш долговременный и условный. Но социобиологи прогнозируют противоположное! Обратимое и гибкое членство, «оплата» лишь после «выигрыша» от участия в сети, а не постоянная, оппортунистическое чередование «кооперации» и «обмана» в демонстративной активности и участие/неучастие во взаимодействиях, максимизирующее кратковременный выигрыш и до.
В реальности все выходит иначе. Естественный отбор в популяции «эгоистических индивидов» работает за «честную коммуникацию», а не против нее. Он последовательно выбраковывает не только «обманщиков», но и «глупцов», не умеющих с нужной скоростью, точностью и эффективностью распознать и отреагировать соответствующий общий сигнал. В первую очередь это особи, склонные к ошибкам в распознавании, с низкой реактивностью или те, кто после сигнала тратят какое-то время на исследование происходящего вместо «запуска» инстинктивного ответа.
Наконец, заблаговременное инвестирование в поддержание общей «сети», способное дать больший выигрыш оппоненту или «третьим» особям, чем «инвестору», — типичная групповая адаптация. С социобиологической точки зрения они, если и появляются, не инвариантны, а сильно зависимы от состава сообщества, стратегий индивидов, ибо неустойчивы к «обману», а «обманщики» будут всегда (см. пример в поведении певчего воробья Melospiza melodia; демонстрация этого — одна из заслуг социобиологии). Преобладающим вариантом и/или нормой они могут стать лишь в особых случаях, когда кооперация исходно «эгоистических» индивидов поддерживается отбором родичей или иным специальным механизмом.
На деле мы видим противоположное. Это возникает всегда, специальных механизмов не требуется, достаточно «просто» использования видовых сигналов — специфических средств коммуникации, обеспечивающих ей специфический результат в полном согласии со сравнительно-этологическими концептами. По этой причине кооперация особей в составе «сети» выступает как норма и, напротив, «обман» с «неучастием» редок и быстро «вычищается» механизмами социального контроля, сильно варьирующими от вида к виду.
Примечания
1Де Соссюр (2004) определяет язык как систему дифференцированных знаков, соответствующих дифференцированным «понятиям» внешнего мира (де Соссюр, 2004) = типам конфликтов интересов, «сталкивающих» особей данного вида между собой и разрешаемых через определённым образом организованный обмен демонстрациями — коммуникацию. Это определение касается только функционирования семиотической системы и ничего не говорит о вовлечённости в него (или нет) представлений, мышления, понимания, и пр. Организованный обмен знаками языка, передающий информацию между участниками коммуникативной сети, благодаря чему разрешаются конфликты интересов в системе, может реализовываться помимо психики участников, лишь поддерживающих ретрансляцию сигналов в сети собственной специфической энергией действия, необходимой в реализации инстинктивного акта, т. е. они превращаются в своего рода модемы в компьютерных сетях. Действительно, что восприятие форм демонстраций как сигналов и отреагирование воспринятого (выбор поведения на оснтове сигнальной информации) происходят «автоматически», то и другое представляет собой последовательные этапы реализации инстинкта; индивиды-участники коммуникации могут быть уподоблены живым машинам Тьюринга: (факт 21 и ниже).
2См. Фридман В.С. Поведение больших пёстрых дятлов при расселении, https://cyberleninka.ru/article/n/povedenie-molodyh-bolshih-pyostryh-dyatlov-dendrocopos-major-pri-rasselenii?ysclid=mj5merrx539789832.
3Их перечень с описанием функционирования при охране внегнездовых территорий D.major см. «Зачем большим пёстрым дятлам всю зиму охранять территории», https://www.socialcompas.com/2020/04/26/zachem-bolshim-pyostrym-dyatlam-vsyu-zimu-ohranyat-territorii-2/
4«Зачем большим пёстрым дятлам…», op.cit.
5Напомню, вне демонстрирования в агонистическом контексте они не видны, даже когда хвост максимально раскрыт из-за высокого возбужлдения или физических усилий. Здесь же их белый цвет, тёмный рисунок явно демонстрируется другим дятлам, для чего сеголеток даже поворачивает: так ведут себя не только они, но и все D.major при закреплении.
6 Подробное описание коммуникации в этих конфликтах см. «Зачем большим пёстрым…», op.cit.
7См. подробней «Зачем большим пёстрым дятлам…», op.cit. Понятно, что это свидетельствует о сигнальной роли во взаимодействиях отдельных единиц-демонстраций или, мягче, включающего их поведения и/или подтверждает возможность использования сигнальной информации (в чём бы она не «содержалась, в первых или вторых) для «опережающего отражения» происходящего наблюдателем, непосредственно в нём не участвующим.
8Самцы занимают их на заросших рогозом (Typha spp.) болотах, жёстко охраняют границы и рекламируют, стараясь привлечь в охраняемое пространство максимум самок (территориальная полигиния).
9Это же видим в плотных поселениях больших пёстрых дятлов в осенне-зимний период, «Зачем большим пёстрым…», op.cit.
10Главная территориальная демонстрация у этого вида трупиалов и близких: голова поднята, клюв устремлён вверх, тело напряжено, красные «эполеты» полностью расправлены, почему максимально заметны, контурное оперение прижато.
11песни южного соловья образуются комбинаторикой крупных единиц — блоков, каждый из которых — последовательность более элементарных вокализаций, фигур и фраз, следующих друг за другом в довольно жёстком порядке. Каждый самец комбинирует блоки по-своему, поэтому индивидуален не только состав песни, но и употребление разных блоков в разной ситуации, подобно позывам у разных пар воронов (Engguist-Düblin, Pfister, 2002). Если алгоритм комбинаторики блоков, образующий песню, характеризует индивида, то разнообразие блоков – локальную популяцию: самцы «черпают из общего резервуара» (Hultsch, Todt, 1989).
12Птицы покрыли в среднем 1,12 км в первую, 1,49 км во вторую ночь, максимально до 4,5 и даже 6 км
13Русский пересказ см. Алексей Гиляров. Соловьихи ищут женихов во время ночных прогулок, https://elementy.ru/novosti_nauki/431022/Solovikhi_ishchut_zhenikhov_vo_vremya_nochnykh_progulok
14У больших пёстрые дятлов данное поведение зависит от прошлой успешности охраны пространства и, в свою очередь, увеличивает ригидность сложившейся стратегии поведения, затрудняет изменения данной успешности, см. «Зачем большим пёстрым дятлам всю зиму…», op.cit.
15Поскольку именно их специфическое действие (в первую очередь как социальных релизеров, но также и через выбор поведения каждым из участников по ним как сигналам) детерминирует интересующие «подглядывателей» исходы взаимодействий; в каждый данным момент для «подглядывателей» и непосредственных участников взаимодействия, фиксирующих смену форм поведения друг у друга, они выступают предикторами будущего течения процесса, в том числе вероятности наступления разного рода исходов (см.«20 фактов об инстинктах», факты 6-7).
16Как резиденты, так и активные вселенцы, пытающиеся здесь занять территории, приобрести партнёра или иным способом «встроиться» в её социальные связи — но не «проходящие» или нетерриториальные особи.
17Все источники здесь по Фридман, 2013, 2017аб.
18Как именно это происходит, см. «Коммуникация животных и машина Тьюринга»
19Подробное описание этой дифференциации репертуаров у больших пёстрых дятлов, охраняющих осенне-зимние территории, и её канализирующего влияния на модели поведения индивидов, всё больше «расходящиеся» между собой, сообразно дифференциации социальных статусов, см. https://www.socialcompas.com/2020/04/26/zachem-bolshim-pyostrym-dyatlam-vsyu-zimu-ohranyat-territorii-2/
20Сегодня её одни авторы относят к роду Anatitlania, другие — к роду Hypsophrys.
21См. Алан Тьюринг. Вычислительные машины и разум// Mind. 1950.
22См. «20 фактов об инстинктах», факт
23Скажем, на основе обучения, рассудочной деятельности, копирования поведения других особей и пр. О существующих здесь возможностях см. Харитонов С.П. «Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования, https://www.socialcompas.com/2020/03/28/ratsionalnoe-i-irratsionalnoe-v-povedenii-ptits-sochetanie-ratsionalno-veroyatnostnoj-i-relizernoj-modelej-povedeniya-pri-prinyatii-resheniya-o-vybore-mesta-gnezdovaniya/
24Мы назовём мягче, групповой адаптацией: выгодная при реализации всеми членами социума и/или популяции, она рискованна и/или затратна для особей, включившихся в эту реализацию, причём «плата» немедленна, а «выигрыш» долговременный. Таковы же реализация инстинкта в целом, включая организованный обмен демонстрациями во взаимодействиях, т. е. коммуникацию. «Альтруизм» лучше оставить за действиями в пользу другого без очевидного выигрыша здесь и сейчас (обычных, например у приматов), здесь выигрыш очевиден и легко измерим. См. Фридман В.С. Групповые адаптации не нуждаются в групповом отборе, https://cyberleninka.ru/article/n/gruppovye-adaptatsii-ne-nuzhdayutsya-v-gruppovom-otbore?ysclid=mkpdegyvzl545939359; «Что не так с социобиологией?».
25Кнорр-Цетина Карин, Брюггер Урс. Рынок как объект привязанности: исследование постсоцальных отношений на финансовых рынках, https://cyberleninka.ru/article/n/rynok-kak-obekt-privyazannosti-issledovanie-postsotsialnyh-otnosheniy-na-finansovyh-rynkah?ysclid=mkmkjl2s3269327164
Рекомендуем прочесть
 «Жесты», вόроны и шалашники
«Жесты», вόроны и шалашники Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных «Работа» релизеров в коммуникации животных: хороший стимул – плохой сигнал, и наоборот
«Работа» релизеров в коммуникации животных: хороший стимул – плохой сигнал, и наоборот Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей Про танец как изоморфизм инстинктивной коммуникации
Про танец как изоморфизм инстинктивной коммуникации- Про смену парадигм в нейрофизиологии
- Закон гетерогенной суммации Альфреда Зайца: когда выполняется, когда нет?
- Риск демонстрирования как подтверждение «сигнальности» демонстраций
