
Прекрасные расписные малюры (Malurus cyaneus, близкие родственники блестящих расписных малюров, которым также свойственны внебрачные копуляции), у гнезда, из которого выглядывает птенец паразитирующей на малюрах бронзовой кукушки (Chalcites sp.). Рисунок известного художника-анималиста и орнитолога Джона Гульда из статьи K. L. Buchanan, A. Cockburn, 2013. Fairy-wrens and their relatives (Maluridae) as model organisms in evolutionary ecology: the scientific legacy of Ian Rowley and Eleanor Russel
Австралийские птицы — синие расписные крапивники или малюры Malurus cyaneus могут гнездиться просто территориальными парами или коммунально, с помощниками — неразмножающимися особями, выкармливающими потомство размножающейся пары. Строят крупные шарообразные гнёзда, тёплые и тёмные внутри, что важно для нашего изложения.
Обычно хозяева гнездовых паразитов пытаются помешать откладке яиц кукушкой или воловьей птицей — атакуют, окрикивают, и гонят, всяко препятствуя проникновению к гнезду.

Кукушки, паразитирующие на прекрасном расписном малюре: рыжехвостая…

…и блестящая
Малюры же не решаются — может быть потому, что паразитирующая на них местная бронзовая кукушка Chrysococcyx basalis, напоминает ястребов, охотящихся на крапивников, ошибка может стоить жизни. Поэтому при откладке яиц кукушки легко обманывают крапивников, те практически не пытаются им помешать. В месте работы Langmore et al., 2003 (парк Campbell, Канберра, Австралия) 19-37% гнезд малюров содержат яйцо кукушки (данные 1999-2002 гг., N=349 гнёзд).
Малюры распознают и отвергают птенцов, а не яйца кукушек
А дальше реакция малюров зависит от стадии их репродукции. 13 из 15 кукушачьих яиц, отложенных до яйцекладки малюров, втаптывались хозяевами в подстилку гнезда, гарантируя невылупление паразита, но 52 из 53, появившихся в дни яйцекладки, были приняты (G-критерий знаков, G2 =47,5, P < 0.0001). А дальше самое интересное: если кукушка откладывала яйцо после начала инкубации, родители оставляли кладку (7 из 8 случаев), но когда кукушечьи яйца подкладывали сами исследователи, малюры приняли 26 из 27 яиц (G2 =23,1, P < 0.0001). Следовательно, ключевое различие — образ самой кукушки, увидев которую у гнезда (что более вероятно во время насиживания, чем до), родители оставляют кладку, обрекая паразита на смерть. Такая судьба постигает до 40% гнёзд с птенцами Ch.basalis, специализирующейся на данном виде, и 100% — с птенцами бронзовой кукушки Chrysococcyx lucidus, использующей его нерегулярно (Langmore et al., 2003). С учётом обоих защитных мер малюров, эффективный уровень паразитизма составляет 16-32%.
Дальше подробно исследовали, распознают ли малюры чужие яйца и кукушонка в гнезде, и как защищаются в этих случаях. Подкладывание чужих яиц, сходных или резко отличных по окраске от их собственных, показало что нет, не распознают, тем более что их яйца и паразита весьма похожи.
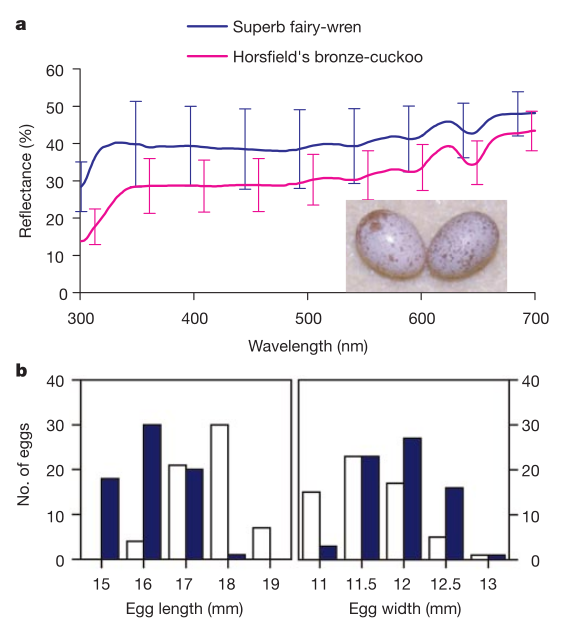 Рисунок 1. Яйца прекрасного расписного малюра и рыжехвостой кукушки почти идентичны. а. Различия в окраске, показано среднее (± стандартное отклонение, SD) спектральное отражение яиц первого (синяя линия, N = 26 из 16 кладок) и второго (розовая, N = 9) видов, длина волны в нм (абсцисса). Ордината — отражательная способность, %, SD рассчитана по вариациям внутри и между яйцами. Яйца кукушек и малюров весьма сходны, первые отражают несколько хуже (анализ главных компонентов, 9 яиц на вид, PC1: F1,16 = 7,80, P = 0,013), видимо, из-за большего числа темных пятен на остром конце яйца. Врезка — яйца малюра слева) и кукушки (справа). б. Распределение яиц бронзовых кукушек Хорсфилда (белые столбцы) и малюров (синие) по длине (слева, мм) и ширине (справа, мм).
Рисунок 1. Яйца прекрасного расписного малюра и рыжехвостой кукушки почти идентичны. а. Различия в окраске, показано среднее (± стандартное отклонение, SD) спектральное отражение яиц первого (синяя линия, N = 26 из 16 кладок) и второго (розовая, N = 9) видов, длина волны в нм (абсцисса). Ордината — отражательная способность, %, SD рассчитана по вариациям внутри и между яйцами. Яйца кукушек и малюров весьма сходны, первые отражают несколько хуже (анализ главных компонентов, 9 яиц на вид, PC1: F1,16 = 7,80, P = 0,013), видимо, из-за большего числа темных пятен на остром конце яйца. Врезка — яйца малюра слева) и кукушки (справа). б. Распределение яиц бронзовых кукушек Хорсфилда (белые столбцы) и малюров (синие) по длине (слева, мм) и ширине (справа, мм).
Дальше исследовали, отличают малюры когда-нибудь чужие яйца от своих. При яйцекладке в их гнезда добавляли раскрашенное яйцо, отличающееся от их собственного цветом, рисунком или размером. Малюры не отвергали яйца необычного цвета или рисунка (выбросив 3 из 16 яиц, крашеных в голубое, 2 из 11 в пятнистое, 4 из 18 яиц исходного цвета малюра, рис.1), 0 из 7 крашенных в коричневое были оставлены или выброшены; χ² = 1,8, df=3, P = 0,62; см. также ссылку 8), но значимо чаще бросали кладки с яйцами крупнее их собственных. Так, они отвергли 7 из 15 более крупных яиц, но лишь 1 из 11 более мелких (те и другие цвета пергамента; G2 = 4,2, P = 0,04).
Тусклый свет в шарообразном гнезде малюра (средняя освещённость ± SE = 976 + 105 лк, N = 107 гнезд) мог способствовать тактильному, а не визуальному распознаванию чужих яиц, стратегии, наблюдаемой у других видов с шарообразными гнёздами5,9. Правда, это плохая защита от рыжехвостой кукушки, несущей яйца близких размеров к яйцам малюров (рис.1). Следовательно, когда кукушки откладывают яйца одновременно с малюрами, те ничего не могут им противопоставить.
А вот появившихся кукушат малюры бросают (11 из 29 или 11 из 42, если с разоренными гнездами), однако собственных птенцов никогда (N = 95 гнезд, 18 из них под ежедневным наблюдением). Вероятность бросания кукушонка не зависит ни от того, вылупились до этого птенцы хозяина или нет (N = 37, G2 = 0,83, df=1, P = 0,36), ни от месяца вылупления, т. е. времени года (с сентября по январь, G2 = 1,9, df=4, P = 0,7).

Во всех 11 кукушонка бросали в возрасте 3–6 дней (N = 7). Самка прекращала его кормить, строила новое гнездо и спаривалась. Хотя самец (или самцы в коммунальной группе) иногда кормил кукушонка до трех дней (5 случаев), в конце концов он умирал от холода или голода. Труп обычно съедали муравьи-мясоеды Iridomyrmex purpureus. Следовательно, малюры отвергают чужих птенцов, те не гибнут как бы сами собой, из-за менее эффективного выпрашивания по сравнению с птенцами хозяина, как это показано для ряда видов гнездовых паразитов, не выбрасывающих птенцов хозяев15–17.
Авторы дальше исследовали способы распознавания малюрами кукушат. Они манипулировали кладками (см. Методы) так, чтобы после вылупления в гнездах находился 1 птенец рыжехвостой (N = 20) или птенец блестящей кукушки (N = 8), или один птенец малюра (N = 14). Птенцов могли бросить во всех трёх случаях, и можно подумать, что самки распознают кукушат просто потому, что те остаются одни в гнезде. Однако явно важные и другие признаки, ибо птенцов блестящей кукушки бросали всегда, тогда как малюра и рыжехвостой кукушки — с существенно меньшей вероятностью, причём одинаковой для птенцов хозяина и специализирующегося на нём паразита (рис.2).
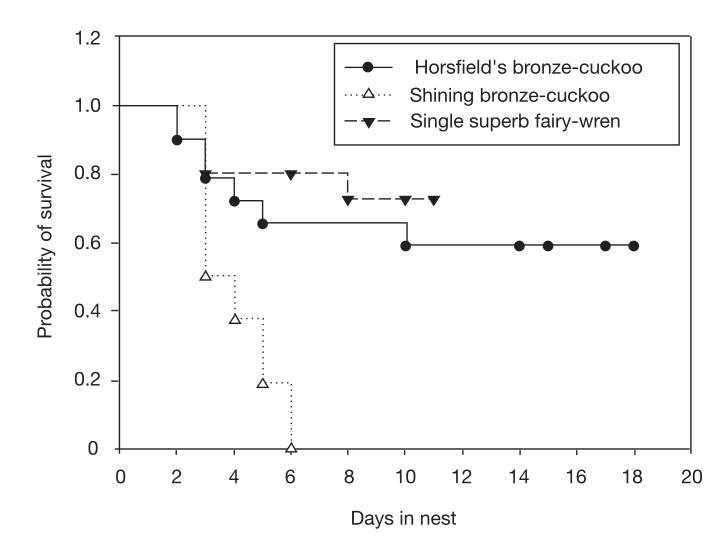 Рисунок 2. Кривые выживаемости птенцов кукушек (рыжехвостой, чёрные точки, и бронзовой, белые треугольнички) и одиночных птенцов малюров (чёрные треугольнички) в своём гнезде без кукушек (кривые Каплана–Мейера, ось Х дни, У — шанс дожить до данного дня=1-шанс, что родители оставят гнездо). Различия между видами значимы (χ² = 9,01, df=2, P = 0,01). Риски быть брошенным одиночных птенцов малюров значимо не отличались от таковых бронзовых кукушек, специализирующихся на этом виде (коэффициент риска = 0,82, 95% доверительные интервалы 0,4, 1,59), при высоко значимых отличиях у блестящих кукушек (коэффициент риска = 2,44, 95% доверительные интервалы 1,21, 4,85).
Рисунок 2. Кривые выживаемости птенцов кукушек (рыжехвостой, чёрные точки, и бронзовой, белые треугольнички) и одиночных птенцов малюров (чёрные треугольнички) в своём гнезде без кукушек (кривые Каплана–Мейера, ось Х дни, У — шанс дожить до данного дня=1-шанс, что родители оставят гнездо). Различия между видами значимы (χ² = 9,01, df=2, P = 0,01). Риски быть брошенным одиночных птенцов малюров значимо не отличались от таковых бронзовых кукушек, специализирующихся на этом виде (коэффициент риска = 0,82, 95% доверительные интервалы 0,4, 1,59), при высоко значимых отличиях у блестящих кукушек (коэффициент риска = 2,44, 95% доверительные интервалы 1,21, 4,85).
Следовательно, оставление кукушат — приспособительный ответ на гнездовой паразитизм. Опыты рис.2 показали, что оставление кукушат — не просто побочное следствие невыгодности заботы (особенно группой1) об одиночном птенце, иначе малюры одинаково часто бросали бы птенцов всех трёх видов. Если выводки с одним птенцом были невыгодны, то одинаково часто оставляли бы одиноких птенцов всех трех видов, к тому же подобное оставление практикуется всеми самками, а не частью популяции. 10 из них были заражены кукушатами обоих видов, 2 из 5, вырастивших одинокого птенца малюра, впоследствии бросили птенца рыжехвостой кукушки; 5 из 5 вырастивщих его, в дальнейшем бросили птенца блестящей кукушки.
Бросание кукушат — не стратегия против неродственных особей в гнезде: самки бросают их даже при связанности родством со всем своим выводком в незараженных гнездах (N = 40 выводков18), самцы же обычно не родственны птенцам (самки малюров летают спариваться на соседние территории), но старательно кормят их и кукушонка18. Редко паразитирующие блестящие кукушки обычно распознавались малюрами, тогда как специализированный паразит, кукушка рыжехвостая, существенно реже, т. е. защитные механизмы против кукушат, вероятно, вызвали их контрадаптации.
Видимо, визуальные признаки для различения кукушат несущественны. Птенцы блестящей кукушки принадлежат к одной из двух цветовых морф: розовато-желтой, похожей на птенцов малюра (рис.3b), или черной (рис. 3c), заметно отличающейся. Птенцы рыжехвостой кукушки промежуточны по окраске (рис. 3a). Внешне похожих птенцов светлой морфы, однако, малюры постоянно бросают (N = 4), тогда как весьма непохожих птенцов рыжехвостой, наоборот, принимают (14 из 20 случаев; точный тест Фишера, P = 0,02).
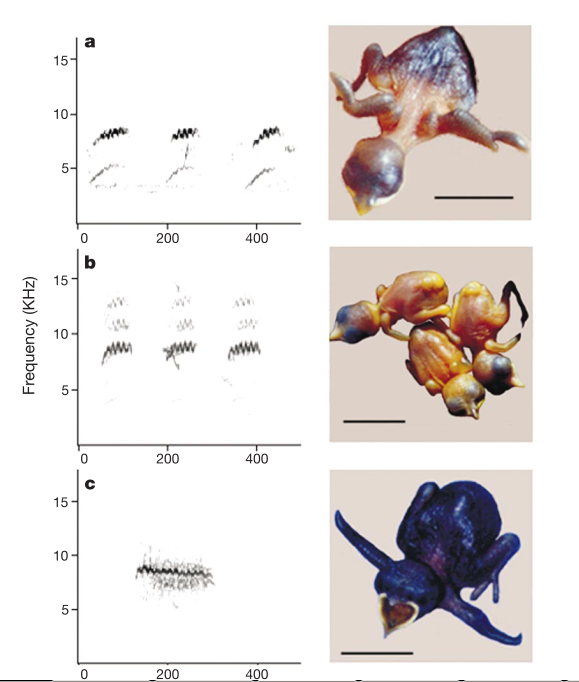
Рисунок 3. Сонограммы криков выпрашивания, 6 день после вылупления: кукушонок рыжехвостой (а, N = 7 записей от 4 птенцов) и бронзовой (с, N = 5 записей от 2 птенцов) кукушек, b. — выводка из 3х птенцов малюров (N = 14 записей от 8 выводков). Врезки а-с — фото птенцов данных видов через 1–2 дня после вылупления. Фото b слева и справа — 2 птенца малюра, в центре птенец светлой морфы бронзовой кукушки. Масштаб 10 мм.
Тогда изучили вокальное распознавание, предположив, что различение криков своих и чужих птенцов малюров способствовало отбору миметических криков выпрашивания у рыжехвостой кукушки, но не блестящей. Анализ сонаграмм криков выпрашивания птенцов всех трёх видов (там измеряли частоту самой громкой части крика, его длину и частотный диапазон) показал куда меньшее сходство с малюрами у птенцов блестящих кукушек светлой морфы, чем у птенцов рыжехвостой (рис.3; см. также ссылку 19).
По мере взросления птенцов всех трёх видов у них значимо не отличались ни пиковая частота (дисперсионный анализ (ANOVA), взаимодействие вида и возраста: F2,21 = 0,40, P = 0,66), ни частотный диапазон вокализаций (ANOVA, взаимодействие вида и возраста: F2,21 = 0,40, P = 0,66). Однако крики кукушат блестящей кукушки значимо длиннее, чем у малюров, и это различие всë явственнее по мере взросления птенцов (ANOVA, взаимодействие вида и возраста: F2,21 = 8,20, P = 0,002; разность групповых средних блестящей кукушки vs малюра значимы, наименьшая значимая разность групповых средних Фишера, PLSD, P = 0,03). А вот птенцы рыжехвостой кукушки значимо не отличаются от малюров по этому признаку (PLSD, P = 0,40).
Поэтому авторы предположили, что различение вокальных сигналов хозяевами могло способствовать отбору миметических призывных криков у бронзовых кукушек Хорсфилда. Еще предстоит определить, врожденна способность хозяина различать кукушат или достигнута обучением. Так, дроздовидные камышовки Acrocephalus arundinaceus запоминают первую кладку и потом отвергают все яйца, на неё непохожие4. Однако это немыслимо у малюров, где распознаются птенцы, а не яйца: слишком уж велика цена их неправильного импринтинга на одинокого птенца кукушки или отказа от собственного птенца, что перевешивает выгоду от избегания эксплуатации6. Данные авторов опровергают гипотезу Лотема6, что принявшие кукушат самки их импринтировали и дальше отвергнут всех прочих птенцов. Самки, принявшие птенца рыжехвостой кукушки, не бросили одинокого птенца малюра в следующем размножении (0 случаев оставления гнезда в шести случаях, биномиальный тест, P = 0,03).
Возможно, среди хозяев кукушек, чьи птенцы губят птенцов хозяина, малюры уникальны по способности бросать кукушат. Стоимость принятия кукушонка ~ вероятности обнаружить его в гнезде, зависящей от интенсивности паразитизма и силы защитных механизмов хозяина на стадии яйца, а поскольку у малюров эти механизмы действуют на следующей стадии, цена гарантированно выше. Скажем, британские тростниковые камышовки Acrocephalus scirpaceus — хозяева европейской кукушки Cuculus canorus — отвергают примерно 20% отложенных им яиц кукушки, т. е. в конечном счёте выкармливает кукушат 5–13% их пар³. Однако в 3х разных популяциях малюров, бывших под наблюдением в Новом Южном Уэльсе, в 13–37% гнёзд был птенец рыжехвостой кукушки (данное исследование; см. также ссылки 20, 21). Даже с учетом неверного времени откладки яиц самками рыжехвостой кукушки, как следствие, оставления гнезд хозяевами, малюры значимо чаще сталкиваются кукушонком, чем тростниковые камышевки.
Ещё по ряду причин малюрам выгодней распознать и отвергнуть птенцов, а не яйца паразита. Их Сезон размножения крайне растянут (три-шесть месяцев22), что значимо больше, чем у типичного хозяина обыкновенной кукушки в Европе (2,5 месяца у тростниковой камышовки, 2,5 месяца23). К тому же малюры легче переносят издержки, созданные ошибочным оставлением одиночного своего птенца: выводков с одним птенцом у них 3,08% (N=1587; А. Кокберн, личное сообщение), т.е. сравнимо с камышевками (3,8% из 2094 выводков3,24), оба вида в среднем выводят 1-2 птенца/гнездо22,23. Однако малюры имеют до трех выводков в год (34,4% самок успешно делают два выводка25), т.е. в среднем 2,6–3,9 птенцов/гнездо14,20,21,25., тогда как второй выводок были лишь у 8–32% камышовок23. Поэтому гибель одного птенца куда меньше скажется на репродуктивном успехе малюров, нежели камышовок.
Резюмируя: авторы предполагают, что распознавание кукушонка возникает только в ответ на преодоление паразитом защитных механизмов, связанных с распознаванием яиц, и лишь при достаточно высоком уровне паразитизма, когда гибель распознанных кукушат перевесит издержки ошибок распознавания, т. е. сезон размножения достаточно длинен для массовых повторных кладок, а плодовитость хозяев достаточно высока , чтобы выдержать «цену» ошибочного оставления своих птенцов.
Птенцы малюров внутри яйца выучивают «талон на обед», а кукушата не могут
Расписные крапивники нашли другое решение — использовать для борьбы с кукушками эволюционное достижение певчих птиц, которого нет у неворобьиных, именно способность к вокальному научению, причём не только в песне, но и в позывах (см.обзор И.Р.Бёме, стр.31-42). Самка крапивник озвучивает развивающиеся эмбрионы специальной вокализацией (в статье написано «поёт, но это, конечно, позывы), которую они выучивают. Она становится «талоном на обед» — птенцы встраивают выученную мелодию в крики выпрашивания, их начинают кормить лишь после его предъявления.
Также некоторые геригоны (Gerygone spp., сем. Шипоклювковых Acanthizidae) выбрасывают визуально имитирующих птенцов Chalcites spp. из гнезда, продолжая заботиться об оставшемся выводке [8, 9]. Прекрасным расписным малюрам недостаточно визуального распознания: хотя кукушата убивают их птенцов, оставшийся птенец рыжехвостой (но не блестящей) кукушки имитирует крики выпрашивания хозяев.
Diane Colombelli-Négrel et al. (2012) исследовали, как малюры распознают кукушонка, несмотря на сходство его криков выпрашивания с их собственными. Они предположили, что малюры распознают кукушонка по акустическому шаблону, причём с гибким и зависящим от опыта порогом отторжения11,17,18, ибо малюры чаще бросают одиночных птенцов именно в годы встречаемости рыжехвостых кукушек в месте их гнездования11. Кукушки же избирательно имитируют крики выпрашивания птенцов хозяина14.
Выяснилось, что прекрасные расписные малюры в борьбе с кукушками используют эволюционное достижение певчих птиц, которого нет у неворобьиных, именно способность к вокальному научению, причём не только в песне, но и в позывах (см.обзор И.Р.Бёме, стр.31-42). Запись 15 инкубируемых кладок в гнёздах по 24 ч/сут показало наличие ранее неописанной вокализации (крик насиживания, incubation call), адресуемой самками развивающимся эмбрионам (рис.4, аудио S1-S3 разных лет и мест).
Самки издавали его в среднем 16 ± 2 раз/ч в конце инкубационного периода, за 4–5 дней до вылупления (что соответствовало 9–10 дню инкубации), прекращая кричать перед вылуплением или сразу после него. 4 из 15 самок издавали инкубационные крики утром в день вылупления яиц, но в последующие дни никогда. Этот трель с 19,1 ± 2,6 отдельными элементами, длительностью 1,8 ± 0,1 с, минимальная частота её составляющих 5782,8 ± 149,3 Гц, максимальная 11011,2 ± 140,5 Гц. Примерно раз в 4 минуты самка адресует её развивающимся эмбрионам, начиная примерно с 9 дня инкубации и до вылупления
Этой озвучки достаточно, чтобы птенцы малюра выучили самочью мелодию и встроили в собственные крики выпрашивания (что выяснилось визуальным анализом их спектрограмм в 12 гнёздах в сравнении с инкубационными криками самок из тех же гнёзд, а подтверждено их кросскорреляционным анализом вкупе с анализом главных компонент, рис.5). Эта мелодия делается их «талоном на обед» — их кормят лишь при наличии данных звуков, кукушата на сие неспособны.
В разных гнёздах птенцы в свои крики выпрашивания включают разные ноты с маминого крика. Получается маркёр принадлежности к одному семейству, дальше используемый в социальной жизни этих птиц. Самки также обучают этому крику своих самцов, включая соответствующую трель в крики, которыми насиживающая самка выпрашивает корм у вернувшегося с ним партнёра, данные по 8 гнёздам 2011 г. (вообще самки активно используют крики выпрашивания с данными «подписями» в социальных взаимодействиях вне гнезда). Благодаря эти вокализациям взрослые различно воспринимают своих и чужих птенцов, кормя первых значительно интенсивней.
При этом у самок с большей высокой частотой инкубационных криков отмечено большее сходство криков с криком выпрашивания птенцов в координатах 2 из 5 главных компонент, использованных в анализе данных вокализаций (линейная регрессия CV1: r = 0,30, t48 = 2,15, p = 0,04; CV2: r = 0,26, t48 = 21,89, p = 0,06; CV3: r = 0,48, t48 = 3,81, p < 0,0001; CV4: r = 0,15, t48 = 1,06, p = 0,29; CV5: r = 0,10, t48 = 0,70, p = 0,49). Усреднение показателей по всем 5 в одну меру показало, что эта последняя — значимый значимый предиктор общей взаимосвязи между структурами тех и других криков — инкубационных матери и выпрашивающих птенцов (r = 0,35, t 48 = 2,57, p = 0,01; см. Таблицу S1, доступную онлайн).
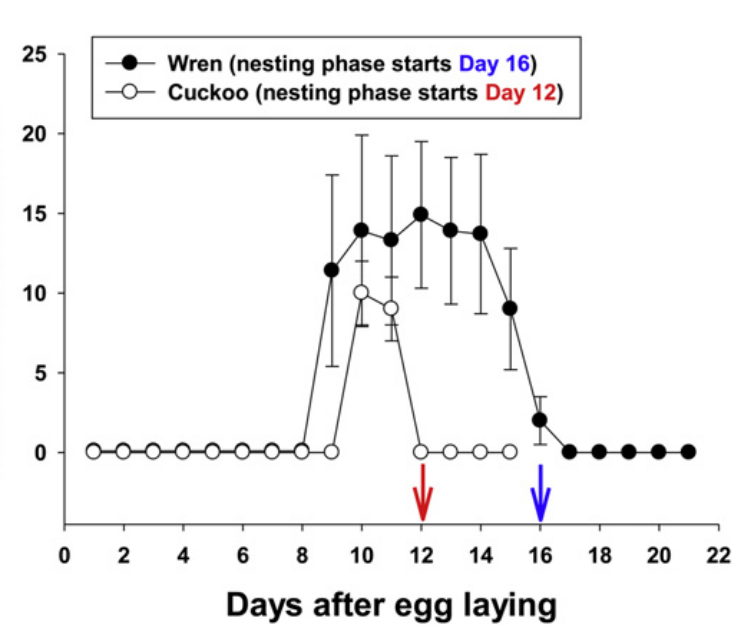
Рисунок 4. Изменения интенсивности инкубационных криков самок малюров в период насиживания (раз/ч, средняя ± SE), дни от начала кладки (различно в заражённых гнездах и нет, N=6 и 2 тех и других, по которым есть ежедневные записи с начала насиживания до начала криков выпрашивающих птенцов, обоих кукушат бросили на 3й день).
Обозначения. Черные точки — птенцы малюров, белые — кукушек Chalcites sp.
Примечания. Малюры насиживают свои яйца ~ 15 дней, кукушечьи ~ 11; начинают насиживать на 10-й день после яйцекладки и прекращают в день вылупления, для гнёзд с кукушонком 12-й (красная стрелка), для незаражённых 16-й (синяя).
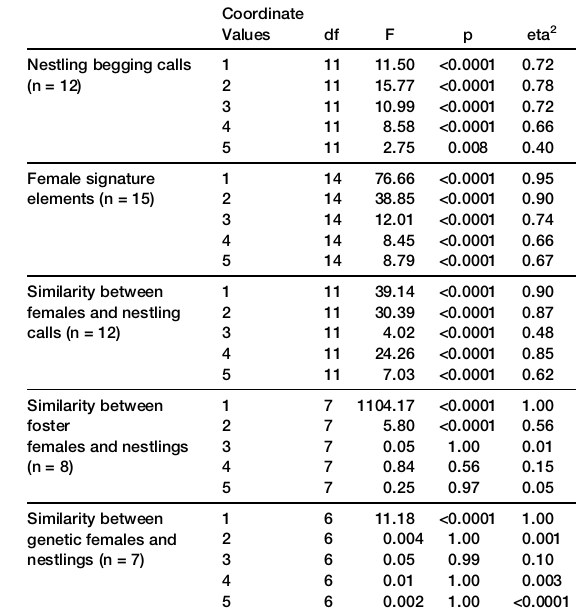
Рисунок 5. Сходство криков выпрашивания птенцов малюров с индивидуальной «подписью» в инкубационных криках выращивающих их самок.
Обозначения. Строки сверху вниз: крики выпрашивания птенцов; инкубационные крики самок; сходство птенцовых криков с самочьими; сходство птенцовых криков с «подписью» их приёмных родителей; то же для сходства с генетическими родителями. Столбцы — результаты многомерного дисперсионного анализа (ANOVA) по первым пяти значениям координат, полученным с помощью анализа главных компонент, df, значимость действия фактора (№ компоненты) на отклик (положение звука в редуцированном пространстве), eta2 — сила влияния фактора на отклик.
Примечания. Авторы выполнили спектрографическую кросс-корреляцию и анализ главных компонент, сравнив этими методами крики выпрашивания птенцов на 3–4 день жизни с «подписью» в инкубационных криках самок на 10–12 день насиживания. Также приведены данные о криках самок и птенцов в опытах с перекладыванием их между гнёздами. Представлены данные по 15 гнездам малюров без кукушат, наблюдавшимся в 2007-2011гг. (три верхние строки табл.) и 8 гнездам, использованным в опытах по перекрестному выкармливанию (две нижние строки).
Гипотезу о заимствовании птенцами малюров индивидуальных «подписей» самок (делающихся «талоном на обед») из их инкубационных криков окончательно подтвердили данные о переносе яиц между гнёздами в инкубации (не позднее 4 дня по окончанию кладки), когда самки еще не издавают криков во время насиживания. В 8 гнёздах малюров авторы перемещали полные кладки — 3 яйца. Действительно, сравнение криков выпрашивания птенцов в этих гнёздах с «подписями» биологических и приёмных матерей (рис.5) показало сугубое сходство первых значений координат с первыми, а не со вторыми, для вторых же значений картина обратная, причём здесь сходство тем больше, чем чаще приёмные матери озвучивали инкубационным криком развивающиеся яйца (рис.6).
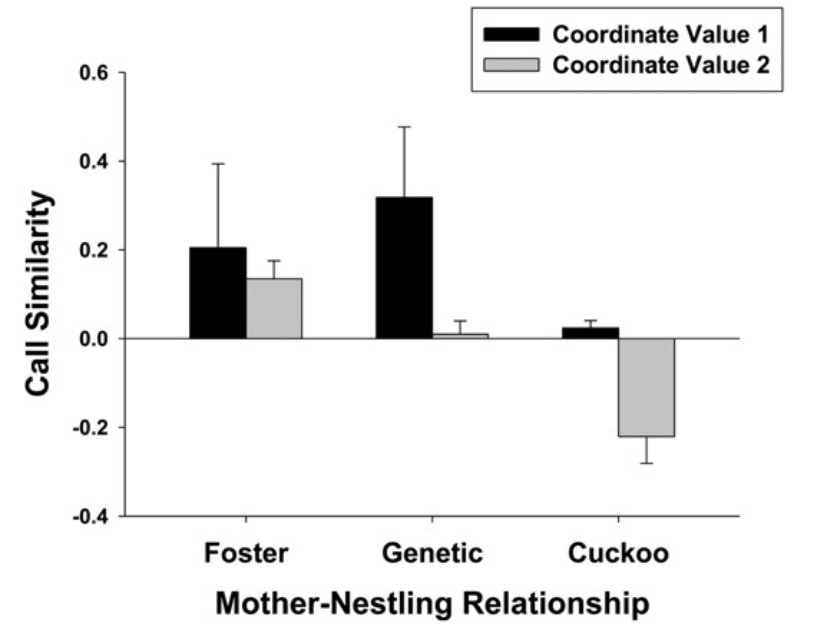
Рисунок 6. Сходство криков 8 птенцов малюра, сменивших гнездо, с криком выпрашивания их приёмных (слева, n=7) и родных (центр, n = 2) родителей, а также рыжехвостых кукушек (n=2).
Обозначения. Представлены средние ± SE совпадения координат сравниваемых криков по первым двум осям главных компонент (CV1 — черные столбцы, CV2 — серые), по ним были максимальны различия вокализаций между родными и приёмными гнёздами.
Сходство с криком выпрашивания рыжехвостой кукушки самое низкое, значимо ниже, чем с биологическими родители у птенцов, сменивших гнездо (рис.6). Перекладкой яиц между гнёздами было показано, что птенцы, выучившие крик приёмных родителей, получают от них корм лучше и больше, чем от биологических. Услышав «неправильные» крики выпрашивания кукушонка, расписные крапивники его почти не кормят, часто тревожатся, а то могут и вовсе разрушить гнездо и начать новую кладку («умри ты, душа моя, с филистимлянами»). См. Colombelli-Négrel et al. (2012).
Помимо того, что кукушонок вряд ли выучит нужные звуки, против него работает разница в фенологии (рис.7). Кукушки подкладывают яйца в гнёзда крапивников достаточно поздно, так что в любом случае не успели бы выучить (примерно на 12й день инкубации у малюров). Cамки, острей чувствующие угрозу гнездового паразитизма (больше видящие и слышащие кукушек, или чаще сталкивавшиеся с ними до этого) интенсивнее «учат» птенцов, а те лучше воспроизводят «пропуск». Поскольку у эмбрионов обучение идёт не очень, самка должна кричать у гнезда много и долго. Соответственно, это рискованно (в первую очередь для гнезда, в меньшей степени для неё). Поэтому «тратятся» на «антикукушечью защиту» не все самки, но лишь непосредственно столкнувшиеся с кукушкой, видящие и слышащие её.
На присутствие взрослых кукушек вблизи гнезда малюры реагируют увеличением шансов бросания одного птенца11,17. Возможно и наоборот: «чужой» крик выпрашивания служит сигналом поиска кукушек в районе гнезда, повышает бдительность малюров и снижает скорость кормления.
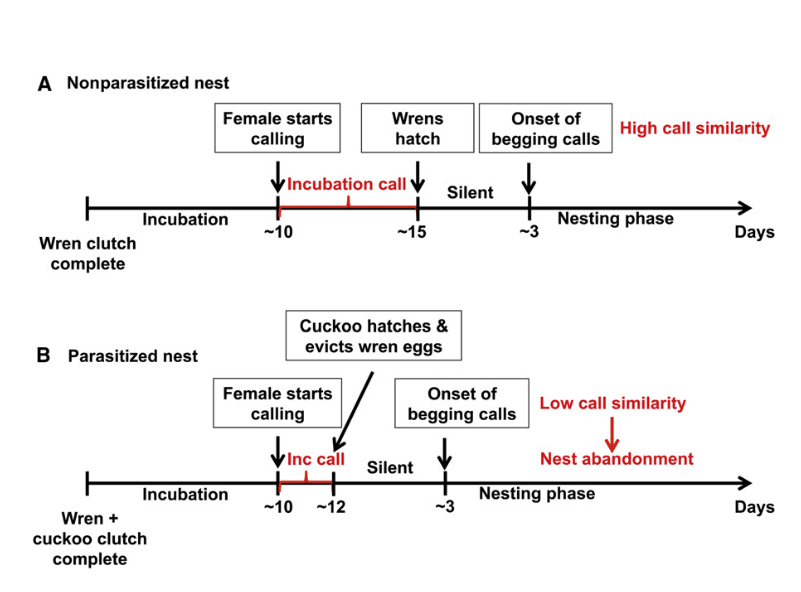 Рисунок 7. Гипотетическая хронология насиживания со «вписыванием» индивидуальных «подписей» из инкубационных криков самок малюра в крики выпрашивания будущих птенцов
Рисунок 7. Гипотетическая хронология насиживания со «вписыванием» индивидуальных «подписей» из инкубационных криков самок малюра в крики выпрашивания будущих птенцов
Обозначения. Цифры на абсциссе — дни, сперва от начала насиживания, после 2й стрелки — от вылупления. А. Кладка без кукушонка, В — с ним. Стрелки слева направо в А — начало инкубационных криков, (красным) длящихся до вылупления (вторая), третья — начало криков выпрашивания птенцов, в В — начало инкубационных криков самки, вылупление кукушонка, выбрасывающего яйца малюра, начало криков выпрашивания кукушонка.
Примечание. Обратите внимание, что эмбрионы малюров слушают инкубационые крики самок ~ 5 дней, эмбрион кукушонка ~ 2 дня, почему в прротивоположность птенцам хозяев гарантированно не выучит индивидуальную «подпись» из инкубационного крика самки и не «встроит» её в собственный крик выпрашивания для использования «талоном на обед».
Далее выяснилшось, что чем чаще самки сталкивались с кукушками, тем важней их птенцам выученный «талон на обед» для получения пищи. Сперва авторы воспроизвели около разных гнёзд крики выпрашивания своих vs чужих птенцов взрослым малюрам (самкам, самцам и помощникам, если коммунальная группа, n=13), затем крики выпрашивания чужих птенцов vs кукушат (n = 18). В обоих случаях для парных сравнений реагирования взрослых птиц подбирали кричащих птенцов одного возраста. Действительно, в ответ на выпрашивание чужих птенцов все взрослые малюры значимо дольше обследовали окрестности гнезда (df = 12; t = 3,00; p = 0,01), реже посещали гнездо (df = 12; t = 22,10; p = 0,05), реже кормили своих (df = 12; t = 3,42; p = 0,005) и больше издавали тревожных криков (df = 12; t = 24,52; p = 0,001).
При этом не выявлено различий в реакции взрослых малюров на выпрашивание чужого птенца или кукушонка (df = 17; все p > 0,06), т. е. «талон на обед» здесь не видовой, а сугубо индивидуальный, его отсутствие всегда воспринимается как наличие паразита. В 8 гнездах, где поменяли потомство, приёмные матери восприняли крик выпрашивания биологически своих птенцов как чужих, не узнали его и реагировали соответственно (все p > 0,09; данные не показаны).
Самый важный момент — сходство акустических форм у птенцовых «талонов» с индивидуальными «подписями» самок, служащее сигналом для разделения «своих» и «чужих», прямо пропорционально частоте самочьих криков; усиливается с возрастом птенцов; управляется риском паразитизма, позитивно влияющим на интенсивность вокализации самок. Также было показано, что птенцы, лучше выучившие «подпись», чьи крики выпрашивания более сходны с самочьими вокализациями, получают от родителей больше корма. Иными словами, порог различения в этой системе не постоянный, но зависимый от контекста; возможно, следующий шаг «гонки вооружений» между малюром и рыжехвостой кукушкой будет тонкая подстройка сенсорных и когнитивных стадий развития кукушонка, чтобы суметь этот порог обойти.

Полосатая шипоклювка
Далее Sonia Kleindorfer et al. (2014) исследовали, могут ли насиживающие самки малюров повысить порог различения в ответ на увеличение риска паразитизма, т.е. увидев или услышав в ближайших окрестностях рыжехвостых кукушек. Для этого гнезда озвучили кукушачьими криками (рис.8а), контроль – крики живущих поблизости полосатых шипоклювок Acanthiza lineata (рис.8b), сравнив интенсивность озвучки инкубационными криками (раз/ч, ордината рис.8) после и до опытов, выполненных в 2х повторностях, в 2012 и 2013 гг.. Оказывается да, могут: в оба сезона размножения самки усиливали подачу инкубационного крика после озвучки криком кукушки, но не шипоклювки (рис.8). Дисперсионный анализ не выявил значимые различия между гнёздами в частоте инкубационных криков до опыта (ни год, ни место наблюдений не влияли); после озвучиваний криком кукушки, но не шипоклювки, их частота значимо возрастает в сравнении с частотой до (парный t-тест, t = 22.936; d.f. = 15; p = 0.010 vs t = 0.788; d.f. = 12; p = 0.446). И чем интенсивней была стимуляция, тем интенсивней и дольше самки малюров озвучивали будущий выводок инкубационным криком. Прямые наблюдения за ними выяснили, что самки, чаще сталкивающиеся с кукушками, издают эти вокализации интенсивней и ближе к яйцам, чем не сталкивающиеся, так что у их птенцов «талон на обед» более действенный.
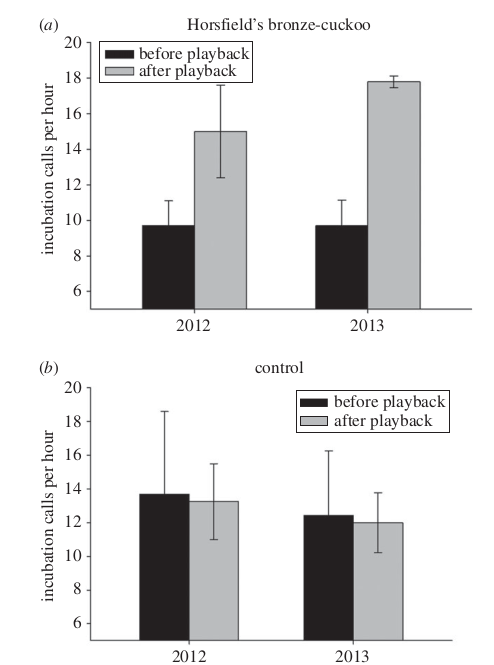
Рисунок 8. Изменения частоты инкубационных криков насиживающих малюров до после озвучивания криком рыжехвостой кукушки (а, n=16) или шипоклювки (b, n=13) в 2 последовательных сезона размножения.
У попугаев, к слову, подобный «талон на обед» развился в персональные идентификаторы птенцов, фактически имена. Они выучиваются от выкармливающих их родителей; точнее, их вокализации выступают матрицей, которую птенец дальше меняет импровизируя, почему его крики оказываются индивидуальной (сходным образом молодые самцы певчих птиц поступают с песнями, выучиваемыми от старых самцов). Дальше они используются для социального общения родителей и потомства в стае, а не только чтобы начать выкармливать тех кого надо, как у малюров.
Примечание
1Как и другие виды малюров, прекрасный расписной практикует коммунальное гнездование: кроме размножающейся пары, на её территории живут несколько более молодых субординантов (обычно самцов), которые сами не размножаются, но участвуют в выкармливании потомства, окрикивании хищника, или охране территории. Самки малюров активно посещают соседние территории, привлекаясь пением владельцев, и спариваются с ними: частота внебрачных копуляций и доля происходящих от них птенцов здесь сравнима с «законными» копуляциями в паре. Самцы, соответственно, стараются спариться с максимальным числом чужих самок.
Рекомендуем прочесть
 Освещение города и гибель птиц
Освещение города и гибель птиц Кто далеко летает, живёт рискованней
Кто далеко летает, живёт рискованней Знание, которое спасает жизнь
Знание, которое спасает жизнь «Работа» релизеров в коммуникации животных: хороший стимул – плохой сигнал, и наоборот
«Работа» релизеров в коммуникации животных: хороший стимул – плохой сигнал, и наоборот Устойчивые морфологические типы в эволюции птиц
Устойчивые морфологические типы в эволюции птиц От «энергетического поворота» к угрозе биоразнообразию — о положении с охраной птиц в Германии
От «энергетического поворота» к угрозе биоразнообразию — о положении с охраной птиц в Германии О повторяющейся эволюции
О повторяющейся эволюции- Как общества служат зеркалами друг другу: схизмогенез у индейских народов США
