

О.В.Смирнова, Н.А.Торопова
I. Необходимость корректировки существующих способов охраны природы и природопользования
1. Последствия традиционного природопользования
Последствия существующих способов традиционного природопользования: потери биоразнообразия, снижение продуктивности и обеднение почв, снижение климатической стабильности подробно описаны в монографии М.В.Бобровского (2010) на основании анализа многочисленных источников.
Появляющиеся данные о высокой продуктивности экосистем на ранних этапах восстановления, противоречат представлениям о деградации биогеоценотического покрова в процессе традиционного природопользования. Причина в том, что после сильного нарушения (например, выжигания леса в цикле подсечно-огневого земледелия) первое поколение растений-эдификаторов реализует весь блок питания, накопленный экосистемой в течение предшествующего развития нескольких поколений в популяциях всех членов биоты, и именно поэтому достигает высокой продуктивности. Исчерпание ресурсов первым поколением приводит к резкому снижению продукционной и остальных функций экосистем, поэтому для достижения экосистемой исходного состояния необходимо спонтанное (не нарушаемое внешними воздействиями) развитие в течение нескольких поколений. Это положение еще не осознано природопользователями, поэтому они не пытаются прогнозировать развитие экосистем значительно дальше «сбора урожая» первого поколения. В то же время, уже накоплен огромный блок данных о потере продуктивности, снижении биологического разнообразия и почвенного богатства после многократных нарушений лесных, луговых и степных экосистем в циклах традиционного природопользования: подсеки, перелога, сбора лесной подстилки, распашки, выпаса и сенокошения. Эти явления постоянно поддерживают экосистемы в раннесукцессионном состоянии, что препятствует им в полной мере реализовать экосистемные функции (Смирнова и др., 1997; Сукцессионные процессы…, 1999; Оценка и сохранение…, 2000; Восточноевропейские…, 2004; Мониторинг…, 2008; Бобровский, 2010).
2. Последствия введения заповедного режима
Наиболее заметное следствие введения заповедного режима на землях, ранее испытывавших мощные антропогенные воздействия (прямые или косвенные) — падение биологического разнообразия после перехода экосистем из раннесукцессионного в позднесукцессионное состояние (Смирнова и др., 1997; Сукцессионные процессы…, 1999; Оценка и сохранение…, 2000; Восточноевропейские…, 2004; Мониторинг…, 2008). Это следствие введения заповедного режима достаточно хорошо документировано в степных заповедниках, экосистемный покров которых был представлен пастбищными экосистемами. Конкретные работы по этой тематике приведены в сводке В.Е.Соколова с соавторами (1997).
Природную регуляцию доисторических степей осуществляли дикие стадные животные-фитофаги, сейчас она не может быть полностью воспроизведена ни выпасом домашних животных, ни сенокошением, ни палами. Однако без этих воздействий на землях степных заповедников катастрофически быстро снижается уровень таксономического разнообразия, зарегистрированный до заповедания. Менее заметен этот процесс в лесных заповедниках, поскольку опыт заповедания значительно короче длительности жизни поколений большинства видов позднесукцессионных деревьев (Сукцессионные процессы…, 1999; Оценка и сохранение…, 2000).
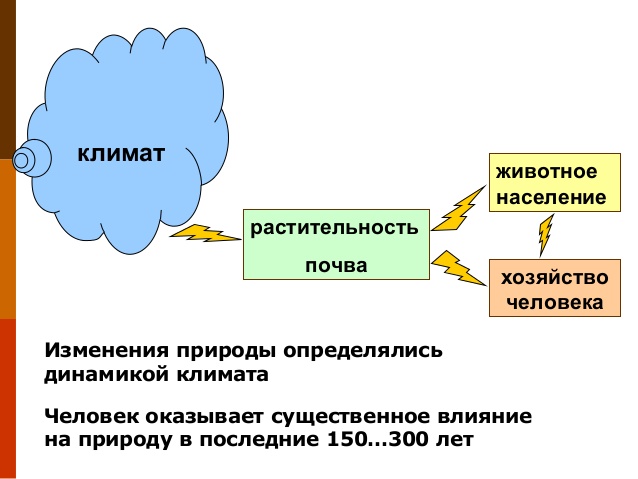
Можно констатировать, что оба способа общения с природой (использование «воспроизводимых» ресурсов и отказ от их использования — заповедание) не позволяют достичь желаемого эффекта, необходимого для устойчивого существования цивилизаций, т.е. возможно полной в современных условиях реализации экологических функций экосистемного покрова. Анализируя современные тенденции деградации биоты биосферы в целом, можно заключить, что для сохранения человечества наиболее значима функция биотической оптимизации климата и гидрологического режима, затем — функции поддержания максимально возможного биологического разнообразия и максимально возможной продуктивности. Постепенно приходит осознание, что сохранение краснокнижных видов или редких экосистем невозможно не только в результате их прямого уничтожения человеком, но и вследствие антропогенно инициированных изменений климатических режимов разного уровня: от локального до глобального.
3. Необходимость разработки принципиально новой, единой стратегии охраны природы и природопользования
В теоретических построениях современных авторов (Горшков и др., 2001; Горшков, Макарьева, 2001, 2006; Мониторинг…, 2008; Методические подходы…, 2010) подчеркнута значимость максимально полной реализации экологических функций биогеоценотического покрова как основного условия его устойчивого существования. При этом наибольшее значение придается функции биотической оптимизации климата как основы поддержания и/или восстановлении максимально возможного биологического разнообразия и максимально возможной продуктивности биогеоценотического покрова и его элементов (экосистем). Однако при решении актуальных задач охраны природы и природопользования степень реализации этой функции, как и всех остальных, не принимают во внимание, что приводит к ошибочным прогнозам направления и темпов изменения биологического разнообразия, как при смене способов природопользования, так и при введении режима охраны.
Основная причина существующего положения – отсутствие четкого осознания различий между потенциями систем любого уровня организации и их реальным состоянием, обусловленным, как правило, многотысячелетней и неоднонаправленной историей использования.
В настоящее время реальное состояние биот экосистем (т.е. совокупности локальных популяций всех видов, обитающих в экосистеме) таково, что они почти полностью лишены возможности преобразовывать абиотические компоненты, т.е. создавать для видов, потенциально способных существовать на данной территории, оптимальный климатический, гидрологический и почвенный режим. В связи с этим, ни сохранение потенциального биологического разнообразия, ни получение максимальной продукции природных экосистем в большинстве регионов при существующих способах общения с природой практически невозможно.
II. Теоретические основы единой стратегии охраны природы и природопользования
Теоретические положения новой стратегии мы формулируем на основе синтеза концепций: оконной мозаики лесных (детритных) экосистем (gap-mosaic concept), мозаично-циклической организации лесных, степных и пустынных (детритных и пастбищных) экосистем (mosaic-cycle concept of ecosystems), а также концепции мощных средопреобразователей экосистем (эдификаторов, ключевых видов — key species, экосистемных инженеров — ecosystem engineers). В качестве примеров приведены некоторые обзорные работы, посвященные этим концепциям (Коротков, 1991; The ecology…, 1985; Oldemann, 1990; Yamamoto, 1992; Kuuluvainen, 1994; Jones et al., 1994; Восточноевропейские леса…, 1994; 2004; McCarthy, 2001; Disturbance dynamics…, 2002; Hastings et al., 2008; Мониторинг.., 2008; Смирнова, Торопова, 2008).
Gap-mosaic» concept возникла при исследовании лесов тропического и умеренного пояса, которые развивались в спонтанном режиме в течение жизни многих поколений деревьев разных видов (Watt, 1925; Коротков, 1991; McCarthy, 2001). В результате непрерывных потоков поколений деревьев в лесных экосистемах постоянно возникают новые местообитания разных размеров, конфигурации и темпов развития, что определяет внутренне обусловленную гетерогенность экосистем и создает условия для совместного обитания экологических различных видов из разных трофических групп (Восточноевропейские…, 1994, 2004; Смирнова, 2004; Смирнова, Торопова, 2008).


Местообитания малых размеров или microsites (площади которых составляют несколько м2) возникают в процессе жизни и смерти особей: это подкроновые пространства деревьев разных видов и разных этапов онтогенеза, межкроновые пространства, элементы ветровально-почвенных комплексов (ВПК): бугры, западины, валежины, формирующиеся в результате падения крупного мертвого дерева, которое сопровождается выворачиванием кома земли. Местообитания более крупных размеров или mezosites (площади которых составляют несколько десятков м2) формируется в процессе зарастания прорывов в пологе леса («окон» или gaps), которые образуются в результате смерти и падения небольшой группы старых деревьев по завершению онтогенеза. В окне формируется новое поколение деревьев одного или нескольких видов, особи которых проходят последовательные этапы развития вплоть до естественной смерти деревьев от старости и болезней. Для описания развития населения окон с момента формирования предложено использовать термин «онтогенетическая парцелла», поскольку различать окна разных этапов развития удобно по скопления особей деревьев одного или нескольких видов близкого онтогенетического состояния. Развитие онтогенетических парцелл исследовано в восточноевропейских неморальных, гемибореальных и бореальных лесах и определены их основные параметры (Восточноевропейские, 1994; 2004).
Mosaic-cycle concept of ecosystems развилась на основе концепции «gap-mosaic», когда представления о биотической гетерогенности среды, создаваемой в процессе потоков поколений деревьев разных видов были использованы при изучении популяционной жизни животных, а затем и представителей иных царств. Эти представления позволили выявить общие принципы организации детритных, и пастбищных экосистем: устойчивое существование большого числа экологически и биологически различных видов в одной и той же экосистеме становится возможным в результате непрерывного преобразования среды обитания («нарушений» — «disturbance» в рассматриваемой концепции) в результате жизни и смерти особей и их групп в популяциях. Смерть и падение старых деревьев; уничтожение подроста, формирование зоогенных полян стадными копытными; строительство хаток, плотин и систем прудов бобрами; колониальные постройки сурков; формирование окон в древостое хвое- и листогрызущими насекомыми и древоразрушающими грибами и пр. приводит к созданию в экосистемах качественно новых местообитаний разного размера: от нано- до макро-местообитаний, которые заселяют экологически различные виды (The mosaic-cycle …, 1991; Восточноевропейские…, 1994; Смирнова, 1998).
Поскольку термин нарушение («disturbance») очень широко используют в современной литературе, необходимо сделать пояснения. В общем смысле нарушение рассматривают как любое событие, которое приводит к разрушению «экосистемы в целом или сообщества, или к изменению популяционной структуры конкретного вида (видов), или изменяет доступность ресурсов и физическую среду» (Disturbance Dynamics, 2002, р. .5). При этом необходимо четко различать эндогенные по отношению к биогеоценозу (экосистеме) и экзогенные нарушения (Смирнова, Торопова, 2008). Эндогенные нарушения – это результат жизни и смерти особей деревьев, крупных стадных копытных, бобров, сурков, термитов и пр. Эти нарушения локальны и формируют мозаики местообитаний относительно небольших размеров (по сравнению с площадью экосистемы в целом), где есть необходимые условия для устойчивого существования всех природных видов и, следовательно, для поддержания высокого уровня биоразнообразия (структурного и таксономического). Именно небольшие размеры элементов мозаики определяет возможность постоянного заноса во вновь возникающие местообитания и приживания зачатков большинства или всех видов-членов сообщества (The ecology …, 1985; Восточноевропейские.., 2004).
Исходя из этимологии слова «нарушение», можно заключить, что его использование при описании естественных проявлений жизни в экосистемах и сообществах некорректно, его целесообразно заменить термином «средопреобразование».

Экзогенные нарушения, т.е. истинные нарушения, – это результат воздействия на экосистемы внешних факторов разной интенсивности и длительности (от кострища в лесу до концентрированных вырубок). Довольно часто в результате экзогенных нарушений природного или антропогенного характера появляются местообитания, размеры которых значительно больше, чем размеры местообитаний, возникающих в процессе средопреобразующей деятельности растений, животных, древоразрушающих грибов и пр. При этом гетерогенность, определяемая эндогенными процессами, нивелируется, и живой (биоценотический) покров становится относительно гомогенным. Именно крупные размеры экзогенно возникших местообитаний — одна из основных причин неполного восстановления биоразнообразия в течение длительного периода после нарушений, поскольку возможности большинства видов расселяться меньше их размеров, иногда на порядки (Удра, 1988, 1990; Методические подходы…, 2010).
Key species concept и ecosystem engineers concept возникли в связи с необходимостью ранжировать виды по степени значимости в процессах структурно-функциональной организации и динамики экосистем. Решая эту задачу, авторы выделили в особые группы виды, средопреобразующие воздействия популяций которых на среду экосистемы приводят к формированию разных местообитаний, необходимых для устойчивого существования популяций большого числа подчиненных видов.
Предложены разные наименования мощных средопреобразователей, наиболее широко распространены два: “keystone species” (ключевые виды) и “ecosystem engineers” (экосистемные инженеры) (Jones et all, 1994, 1997; Gurney, Lawton, 1996; Crain, Bertness, 2006; Gutiérrez, Jones, 2006; Hastings et al., 2008). Исходным можно считать понятие «эдификатор» (aedificator — строитель сообщества, Braun-Blanquet, Pavillard, 1925) широко использованное В.Н.Сукачевым применительно к растениям (1928; 1975). По его представлениям, эдификатор – вид растений, чье присутствие в биогеоценозе заметно меняет экологические режимы: освещенность, влажность, температуру, химический состав почвы, воды и воздуха и др.
В мировой литературе нет единогласия по поводу использования терминов “ecosystem engineers” и “keystone species”, ибо они еще активно разрабатываются. Однако по мере сопоставления характеристик популяций эдификаторов, ключевых видов, экосистемных инженеров, полученных в различных экосистемах для разных видов растений и животных, очевидно, что их целесообразно использовать как синонимы (Смирнова, 2004).
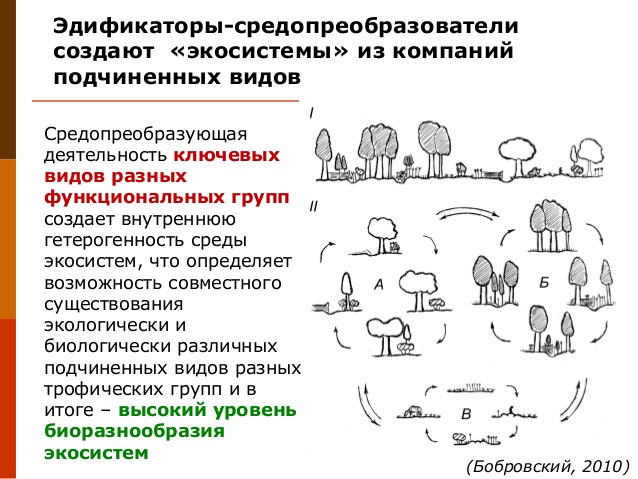
Суть концепций ключевых видов или экосистемных инженеров на современном уровне знаний: каждый вид в экосистеме преобразует среду в процессе формирования потоков поколений в популяции. Однако интенсивность средопреобразующей деятельности различна: наиболее мощные средопреобразователи определяют и постоянно поддерживают организацию экосистемы в целом, в то время менее мощные средопреобразователи определяют и поддерживают отдельные элементы экосистем. В качестве примеров наиболее мощных воздействий обычно приводят строительную деятельность бобров в долинных ландшафтах, формирование мозаики окон возобновления в лесах в результате смерти старых крупных деревьев и пр. Можно заключить, что ключевыми целесообразно считать виды, которые в процессе жизнедеятельности наиболее значительно, по сравнению с остальными видами в экосистеме, преобразуют биотическую и абиотическую среду, что ведет к изменению гидрологического, температурного, светового режимов; микро-, мезорельефа; строения почвенного покрова. В свою очередь, эти преобразования дают возможность жить в одной экосистеме экологически и биологически различным видам и, в итоге, определяют высокий уровень биоразнообразия.
Таким образом, результат популяционной жизни ключевых видов — создание гетерогенной среды экосистем. На локальном уровне это определяет возможность совместного обитания элементарных популяций подчиненных видов, отличающихся по экологическим потребностям. На региональном уровне преобразование среды, производимое наиболее крупными ключевыми видами, по масштабам и характеру изменений сопоставимо с мезоклиматическими изменениями.

По особенностям воздействия на среду ключевые виды могут быть объединены в небольшое число функциональных групп. В настоящее время описаны функциональные группы ключевых видов детритных и пастбищных экосистем водораздельных ландшафтов, а также экосистем пойменных ландшафтов (Смирнова, 1998; 2004; Восточноевропейские…, 2004).
III. Основные положения теории организации экосистемного покрова и составляющих его элементов — экосистем и их комплексов
1. Основу познания структурно-функциональной организации экосистемного покрова составляют представления о потенциях и позициях систем.
Потенции системы – это ее свойства, полностью проявляющиеся при спонтанном (эндогенно обусловленном) развитии в оптимальных условиях. Если такое развитие нарушается внешними (экзогенными по отношению к системе) воздействиями, и/или оно осуществляется в неоптимальных условиях, то система проявляет лишь часть потенциальных свойств, и это характеризует ее позиции в конкретных условиях. Описание развития системы, осуществляемого путем полной реализации потенций в конкретных условиях, представляет собой удобную модель, своего рода «эталонный вариант», необходимый для оценки степени отклонения конкретных систем от эталона и выяснения причин этих отклонений.
2. Экосистемный покров и его элементы достигают полной реализации потенций за счет активной средопреобразующей роли деятельности биоты; при этом максимально полно выполняются основные экологические функции: биотическая оптимизация климата и гидрологический режим, максимально возможное биологическое разнообразие и продуктивность. Активная роль биоты проявляются не только на локальном, но на региональном и глобальном уровнях. В случае если биота Земли полностью реализует потенции, она выступает как основной климаторегулирующий фактор на планете (Горшков, Макарьева, 2001, 2006).
3. Живой покров биосферы и биота каждой экосистемы представляет собой множество взаимодействующих популяций разных видов из разных трофических групп. Основной механизм, определяющий возможность биоты активно преобразовывать среду, — потоки поколений в популяциях ключевых видов. Их средопреобразующая деятельность создает гетерогенность среды экосистем, что определяет возможность совместного существования экологически и биологически различных подчиненных видов разных трофических групп и в итоге – высокий уровень биоразнообразия. Средопреобразующая деятельность ключевых видов увеличивает разнообразие экологических ниш в экосистемах за счет пространственно-временного перераспределения ресурсов. В итоге уменьшается роль конкурентных и возрастает роль комплементарных и мутуалистических отношений в функционировании экосистемного покрова и его элементов (Мониторинг…, 2008; Методические подходы…, 2010).
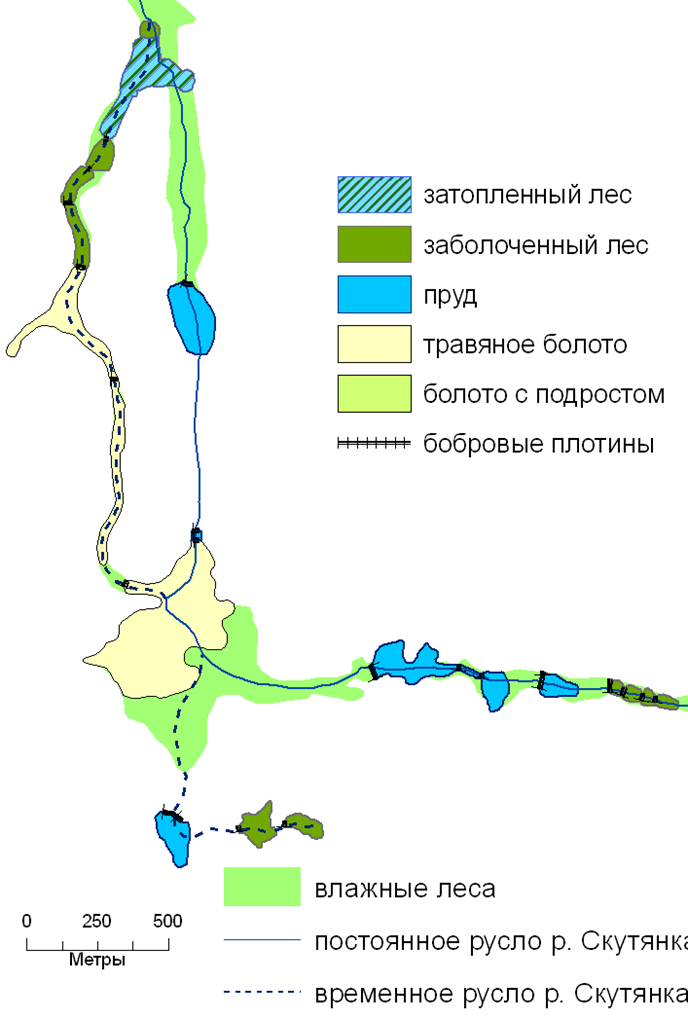
развитие экосистем речных долин после преобразования бобрами. 2004 г.
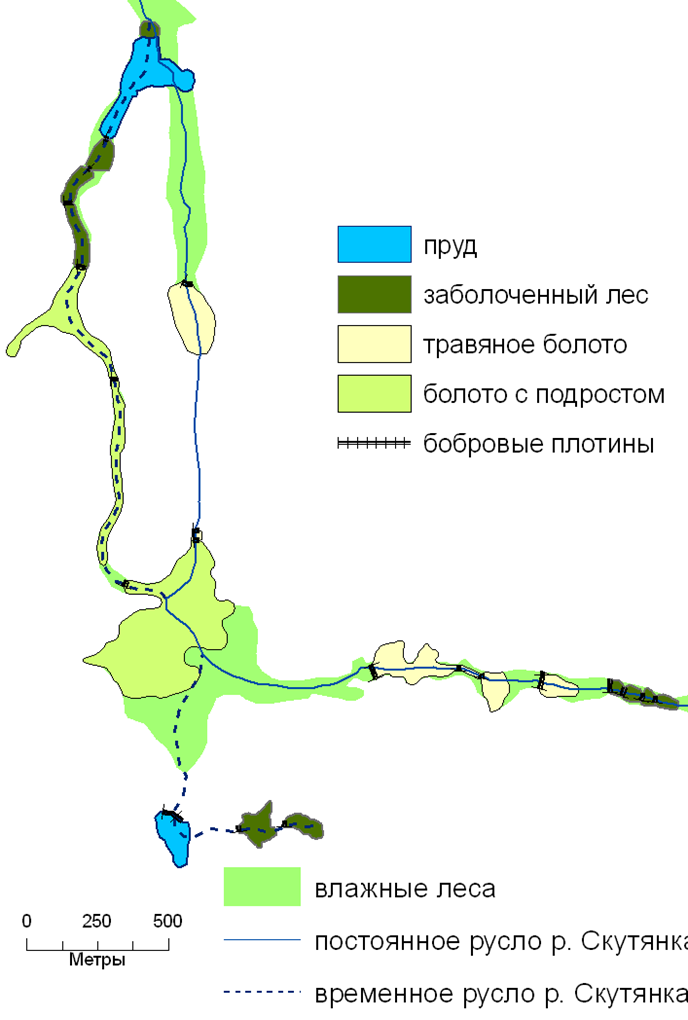
развитие экосистем речных долин после преобразования бобрами. 2008 г.
4. Существуют принципиальные различия между средопреобразующей деятельность природных ключевых видов и воздействиями человека. Способы, интенсивность и размеры воздействий первых сформировались в ходе длительной эволюции, направленной на интенсификацию использования абиотических ресурсов всеми потенциальными обитателями экосистем в целом. Способы, интенсивность и размеры воздействий человека принципиально менялись по мере развития цивилизаций, причем основная тенденция этих изменений – создание преимуществ для избранных видов в ущерб основной части биоты. Такой путь преобразования экосистемного покрова по мере усиления средопреобразующей роли человека, привел к формированию неполноценных экосистем и экосистемного покрова в целом, которые не способны в полной мере реализовать экологические функции (Vera, 2000).
IV. Определение основных понятий синэкологии, необходимых для дальнейшего развития теории организации экосистемного покрова.
1. Экосистема – совокупность взаимодействующих элементарных популяций видов разных трофических групп (биота экосистемы) и активно преобразуемое ими местообитание.
Это определение принципиально отличается по следующим позициям:
1) элемент биоты экосистемы не особь, а популяция, представляющая собой множество особей одного вида; такой подход полностью соответствует системной парадигме и восстанавливает естественный порядок иерархии биосистем;
2) во всех проанализированных нами определениях экосистемы (Смирнова, Торопова, 2008) живое и неживое оценивают как равноправные компоненты, более целесообразно сделать акцент на биоту как на определяющее начало в экосистеме;
3) активное преобразование местообитаний совокупностью популяций всех видов биоты определяет возможность формирования в разных местообитаниях экосистем, биоты которых сходны по составу и структуре.
2. Сукцессия – процесс спонтанного формирования (первичная сукцессия) или восстановления (вторичная сукцессия) потоков поколений в популяциях всех потенциальных членов экосистемы. Можно предположить, что восстановление в экосистеме всех ее потенциальных членов приводит к полной реализации ее потенций.
Значительные расхождения в трактовках понятия «сукцессия» (Миркин и др., 1989; Миркин, Наумова, 1998) связаны с тем, что его используют для объяснения принципиально различных процессов: эндогенного развития, экзогенно-эндогенного развития, экзогенного развития.
Эндогенное развитие начинается после однократного полного или частичного уничтожения предыдущей экосистемы. Оно обусловлено устойчивым потоком поколений в популяциях всех членов биоты после уничтожения предыдущей экосистемы. Этот путь развития назван аутогенная первичная или аутогенная вторичная сукцессия (Одум, 1975).
Экзогенно-эндогенное развитие обусловлено периодическим прерыванием потоков поколений в популяциях (всех или части видов биоты) внешними воздействиями. Если внешние воздействия прекращаются, развитие экосистемы становится эндогенным. Процесс назван аллогенная сукцессия или аллогенное развитие (Миркин и др., 1989; Миркин, Наумова, 1998).
Экзогенное развитие обусловлено постоянным внешним воздействием на потоки поколений всех членов биоты. Оно может быть двух типов. Первый тип – дигрессии – характерен для экосистем, у которых четко различны потенции и позиции; при этом условия среды не препятствуют реализации потенций экосистем после прекращения внешних воздействий. Второй тип (у него еще нет названия) характерен для скоплений особей разных видов, суммативных образований, не обладающих эмерджентными (собственно системными) свойствами, не достигших состояния системы. В фитоценологии они названы экотопически обусловленными группировками видов (Корчагин, 1976). Целесообразно отличать экотопически обусловленные группировки на начальных стадиях первичных сукцессий, которые по мере развития преобразуются в экосистемы, от экотопически обусловленных группировок, которые не могут преобразоваться в экосистемы, ибо их свойства определяют внешние факторы. Приведем несколько примеров. Группировки видов в полярных или жарких пустынях постоянно находятся на пределе возможностей существования, и вследствие этого не в состоянии достичь уровня экосистем. Группировки видов на прирусловых отмелях испытывают периодические воздействия, определяющие их видовой состав и структуру. Разграничение собственно экосистем и экотопически обусловленных группировок необходимо при исследовании процессов развития экосистемного покрова, ибо к группировкам в принципе не приложимы представления о сукцессиях и климаксе.
В современном покрове наибольшие площади занимают экосистемы в состоянии аллогенного развития или дигрессии. Для того, чтобы составить обоснованные прогнозы их развития после прекращения экзогенных воздействий, необходимо сравнивать эти экосистемы с экосистемами, находящимися на разных этапах аутогенных сукцессий и в состоянии квазиклимакса.
3. Основной механизм аутогенной сукцессии – средопреобразующая деятельность ключевых видов. Именно в ходе и в результате ее формируется биотически обусловленная гетерогенная среда экосистемы. Экологическая емкость экосистемы постоянно увеличивается и, как следствие, возрастает экологическое и таксономическое разнообразие. В том случае, если развитие экосистемы осуществляется по типу аутогенной сукцессии, каждый внедряющийся ключевой вид (группа ключевых видов) преобразует внутреннюю среду, увеличивая гетерогенность экосистем, и тем самым обеспечивает сосуществование как предшествующих, так и последующих компаний подчиненных видов. Уничтожение ключевых видов человеком, возрастающее на протяжении всего голоцена, и антропогенно обусловленные разрывы их ареалов приводят к формированию биотически неполноценных экосистем, не способных в полной мере реализовать экологические функции.
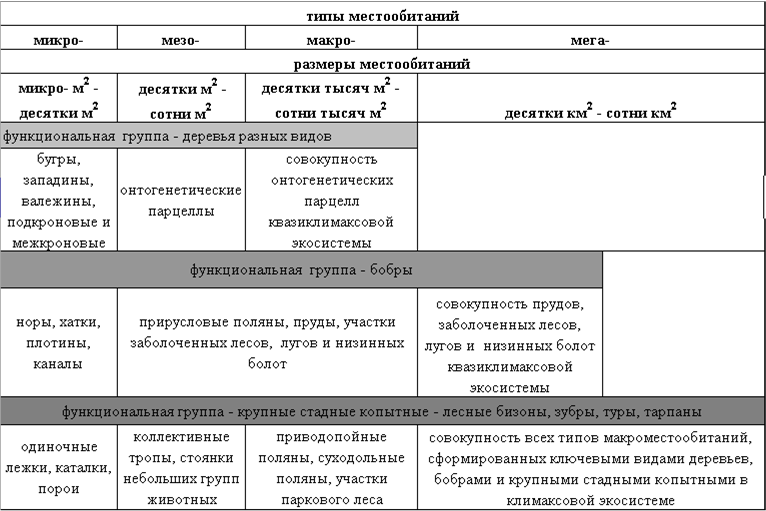
Система местообитаний, создаваемая ключевыми видами разных функциональных групп
4. Собственное (биологическое) время сукцессии – время от начала формирования будущей экосистемы (появление экологически обусловленной группировки) до перехода ее в климаксовое состояние. Его можно измерять календарным временем; а также числом поколений ключевых видов, в результате смены которых экосистема достигает состояния климакса. Одновременное определение обоих показателей позволяет более глубоко понять суть динамических процессов и различать этапы спонтанного развития и экзогенных воздействий. Собственное время сукцессии можно определить лишь при спонтанном развитии экосистемы в ходе аутогенной сукцессии. В случае если аллогенное развитие сменяет аутогенную сукцессию, его длительность можно рассчитать, имея сведения о времени начала антропогенных воздействий на экосистему, о числе антропогенных нарушений и их периодичности.
5. Климакс – такое состояние экосистемы, для которого характерен процесс поддержания устойчивых потоков поколений в популяциях всех потенциальных членов биоты, организуемых средопреобразующей деятельностью ключевых видов. Климаксовыми экосистемами можно считать только такие, в составе которых есть все ключевые виды, потенциально способные в настоящее время обитать на модельной территории. В случае если часть ключевых и связанных с ними подчиненных видов отсутствует, экосистемы правильнее называть квазиклимаксовыми (Смирнова, 2004). В связи с практически полным отсутствием в современном экосистемном покрове истинно климаксовых экосистем необходимо проводить модельные реконструкции их состава, структуры и размеров минимального пространства, необходимого для их устойчивого существования. Нужно также определять состав видов, устойчивые потоки поколений которых ранее поддерживались деятельностью природных ключевых видов, а в настоящее время полностью зависят от человека как наиболее мощного ключевого вида современности (в масштабах от локального до планетарного). В том случае, если исследователи относят экосистемы, постоянно или периодически поддерживаемые экзогенными воздействиями в сукцессивном состоянии, в разряд климаксовых, использование понятия «климакс» становится непродуктивным и принципиально неверным.
6. Собственное пространство климаксовой и квазиклимаксовой экосистем представляет собой совокупность пространств элементарных популяций ключевых и подчиненных видов, при этом часть этих пространств перекрывается. Оно формируется по достижению экосистемой климаксового состояния. Если часть ключевых видов уничтожена, формируется собственное пространство квазиклимаксовой экосистемы.
Представление о собственном пространстве климаксовых и квазиклимаксовых экосистем и о минимальном пространстве, необходимом для выявления их параметров, может быть сформировано на основе определения параметров элементарные популяции ключевых видов. Конкретные расчеты параметров элементарных популяций некоторых ключевых видов деревьев опубликованы (Смирнова и др., 1990; Восточноевропейские…, 1994; Smirnova et al., 2000). Однако для большинства ключевых видов такие расчеты еще надо проводить.
Таким образом, можно заключить, что основные параметры климаксовых и квазиклимаксовых экосистем: размеры (линейные, площадь и объем), состав и структуру можно измерить на основе изучения в природе и/или реконструкции элементарных популяций наиболее мощных ключевых видов и их взаимосвязей с подчиненными видами.
На начальных этапах аутогенных сукцессий границы экосистем можно определить по смене типов предшествующих антропогенных воздействий (следы рубок, пожаров, выпаса скота, мелиорации и пр.); на завершающих этапах – по смене видового состава наиболее мощных ключевых видов. Основные параметры сукцессивных экосистем на каждом этапе развития есть результат взаимодействия экзогенных (в основном — антропогенных) и эндогенных факторов. На начальных этапах развитие экосистем определяют предшествующие нарушения, на завершающих этапах – популяционная жизнь ключевых видов.
V. Основные типы экосистемного покрова
Совершенствование мер охраны природы и природопользования должно быть основано на представлении о глубине деградации экосистемного покрова анализируемой территории. В настоящее время большая часть покрова суши представлена экосистемами в состоянии аллогенного развития или дигрессии, значительно меньшая – экосистемами на ранних стадиях аутогенных сукцессий. Очень небольшие площади занимают экосистемы на поздних стадиях аутогенных сукцессий и в состоянии квазиклимакса. В то же время для оценки возможности выполнения экосистемным покровом экологических функций надо сравнивать конкретные экосистемы и их комплексы с эталонами. Последние в редчайших случаях можно обнаружить в природе или моделировать на основе глубоких знаний биологии и экологии ключевых видов и сопряженных компаний подчиненных видов.

Ельник-зеленомошник, сформировавшийся на заброшенной пашне (И.Шишкин)
Опыт исследования и модельной реконструкции экосистемного покрова лесов Европейской России (Восточноевропейские…, 1994; 2004; Смирнова, 2004; Мониторинг…, 2008; Бобровский, 2010) позволил заключить, что для познания природных закономерностей его организации необходимо различать понятия, обсуждаемые далее.
· Потенциальный покров – это покров, который существовал до начала активных антропогенных преобразований и был сложен климаксовыми экосистемами. Средопреобразующая деятельность локальных популяций полного набора ключевых видов разных трофических групп обеспечивала устойчивое существование экосистемного покрова и полную реализацию его экологических функций. Так, структурно-функциональную организацию потенциального покрова лесного пояса территории Европейской России определяла средопреобразующая деятельность ключевых видов трех функциональных групп. Первая группа – крупных стадные копытные (зубры, туры, тарпаны) — обеспечивала устойчивое существование лесо-луговых и лесо-степных ландшафтов в пределах экосистем, минимальная площадь выявления которых соответствовала и была бы соизмерима площади малого речного бассейна. Вторая группа представлена на территории Европейкой России одним видом – бобром речным, он обеспечивал постоянное поддержание долинных ландшафтов в экосистемах бассейнах малых рек. Третья группа – позднесукцессионных деревьев разных видов (темнохвойных и широколиственных) – обусловливала возможность постоянного поддержания лесных фрагментов водораздельных ландшафтов. Соответственно размеры экосистем потенциального лесного покрова целесообразно определять, исходя из совокупных размеров площадей элементарных популяций наиболее крупных ключевых видов. В лесном поясе Европейской России это были крупные стадные копытные.
Потенциальный покров анализируемой территории был сформирован к началу среднего голоцена и практически полностью уничтожен к середине позднего голоцена (Восточноевропейские…, 2004), однако его модельные реконструкции необходимы для глубокого познания природных механизмов существования экосистемного покрова.
· Восстановленный покров – покров, который может начать формироваться в настоящее время при условии полного прекращения антропогенных воздействий на время, превышающее длительность жизни нескольких поколений наиболее мощных ключевых видов и будет представлен квазиклимаксовыми экосистемами, т.е. экосистемами, в которых сохранилась только часть потенциального набора ключевых видов. В зависимости от того, какие ключевые виды сохранились или какие способны восстановиться на территории, выведенной из хозяйственного использования, облик восстановленного покрова будет различным.
Так, наиболее обычный вариант на территории Европейской России – восстановление собственно лесных экосистем, где среди ключевых есть только позднесукцессионные виды деревьев. В настоящее время, в связи с повсеместно регистрируемым восстановлением природного ареала бобра речного (Гревцев, 2002), возможно восстановление не только собственно лесных экосистем на водоразделах, но и лесо-болотных и прудовых экосистем в долинах малых рек и ручьев (Завьялов и др., 2005; Алейников, 2010).
Прежде, чем делать заключение о наличии восстановленного покрова на конкретной территории, необходимо тщательно исследовать историю ее использования как минимум с начала позднего голоцена, а также реконструировать потенциальные ареалы ключевых видов и оценить возможность их восстановления на анализируемой территории.
При этом следует четко осознавать, что уничтожение большой части ключевых видов привело к гибели большого числа подчиненных видов или к их зависимости от антропогенных преобразований экосистемного покрова, которые в некоторой степени имитировали природную средопреобразующую деятельность уничтоженных ключевых видов. В связи с этим, изъятие того или иного участка экосистемного покрова из существующей системы природопользования может привести к существенному сокращению таксономического и структурного биоразнообразия. При этом реализация иных функций экосистемного покрова, таких как биотическая оптимизация климата и гидрологического режима может становиться более полной, чем в исходном состоянии.
· Современный покров – это покров, который состоит из существующих на текущий момент антропогенно преобразованных экосистем, по преимуществу раннесукцессионных, а также экосистем в состоянии дигрессии. Основной механизм поддержания существующего разнообразия — хозяйственная деятельность человека как главного ключевого вида современности. Для решения существующих проблем охраны природы и природопользования необходима разработка региональных сукцессионных систем на основе единого методологического подхода и согласованной системы методов. Предложен алгоритм исследования экосистемного покрова на основе изложенных ранее представлений (см. раздел VI).
· Оптимальный покров – это покров, который должен и может быть создан, и постоянно поддерживаться человеком в результате единой системы охраны и природопользования, имитирующей воздействия сохранившихся и уничтоженных природных ключевых видов, которые обусловливали возможность полной реализации экологических функций экосистемного покрова. Создание таких систем требует огромных усилий по предварительному изучению современного покрова и огромных вложений капитала и интеллекта для начала реализации принципиально новых подходов общения с природой.
При этом надо сознавать, что существенные практические достижения в реализации новой стратегии могут быть достигнуты только при условии:
· принятия научным сообществом принципиально новой методологии оценки состояния экосистемного покрова и его элементов (экосистем или их комплексов) и интеграции усилий при проведении региональных исследований на основе согласованной системы методов;
· разработки на этой основе региональных моделей единой системы охраны природы и природопользования; их реализация на модельных полигонах, включающих ОППТ и производственные участки экосистемного покрова.
VI. Алгоритм исследования экосистемного покрова лесных территорий
На современном уровне знаний наиболее значимые результаты можно получить при исследовании лесов. Во-первых, это наиболее широко распространенный тип растительности нашей страны, во-вторых, степи и полупустыни как природные образования практически не существуют, и поддержание биоразнообразия целиком зависит от человека, поэтому модельная реконструкции их потенциального покрова значительно сложнее, подобная реконструкция лесного покрова.
Для исследования лесного покрова на основе изложенных теоретических представлений необходимо создать систему длительных стационарных работ, аналогичную организованным В.Н.Сукачевым (1975), координирую задачи, методы и ожидаемые результаты.
Мы предлагаем последовательность действий для оценки состояния экосистемного покрова лесов умеренного пояса:
1. Выбор модельной территории, которая по данным литературы и рекогносцировочным исследованиям наименее преобразована. Такие территории можно обнаружить в заповедниках и иных типах охраняемых территорий (Ярошенко и др., 2001; Aksenov et al., 2002) и анализ ее истории, сопровождающийся картографическим материалом. Глубина ретроспективы как минимум должно быть равна длительности жизни нескольких поколений деревьев позднесукцессионных видов (для лесов умеренного пояса 600-1000 лет). Для этого используют радиоуглеродный анализ углей (многие леса имеют пирогенное прошлое); следы в почве ветровально-почвенных комплексов (ВПК) разных поколений, следы плужной подошвы и корчевания деревьев, определение возраста наиболее старых деревьев, писцовые книги, материалы генерального межевания (Восточноевропейские…, 1994; 2004; Сукцессионные процессы…, 1999; Оценка…, 2000; Мониторинг…, 2008; Бобровский, 2010).
2. Составление списка ключевых видов, ранее обитавших в климаксовых экосистемах модельной территории на основе исторических, археологических и палеонтологических материалов (Восточноевропейские…, 1994, 2004), а также анализ данных региональных археологических отчетов и архивных сведений; расчеты площади выявления климаксовых экосистем модельной территории и определение их положения в структуре ландшафта.
3. Составление списка ключевых видов, ныне обитающих на модельной территории в комплексах сукцессионных систем, сбор недостающих данных об основных параметрах их элементарных популяций и определение минимальной площади выявления квазиклимаксовых экосистем. Составление списка подчиненных видов, обитающих ныне на модельной территории. Разделение их по отношению к свету на две группы: 1) собственно лесные виды, которые существуют в квазиклимаксовых лесах, включая местообитания окон, 2) луговые и степные виды, которые обитают в лесах на ранних этапах сукцессий и на нелесных участках (сенокосные поляны, дороги, залежи). Такие работы проведены на особо охраняемых природных территориях европейской России, но их явно недостаточно для региональных обобщений (Восточноевропейские…, 1994; Сукцессионные процессы…, 1999; Оценка…, 2000). Виды первой группы господствуют на поздних этапах сукцессий и в квазиклимаксе. Среди них надо выделить индикаторные виды, которые наиболее тесно связаны с микроместообитаниями, создаваемыми ключевыми видами. Виды, существующие в широком диапазоне местообитаний, можно считать индифферентными.
4. Членение модельной территории на элементы, размеры которых достаточны для существования в них элементарных популяций ключевых видов доисторических (климаксовых) экосистем. Для лесов умеренного пояса в качестве таковых целесообразно рассматривать бассейны малых рек. Во-первых, здесь есть все разнообразие элементарных ландшафтов: автоморфных, транзитных и аккумулятивных (Глазовская, 2002; Колбовский, 2006), необходимых для реализации жизненных потребностей таких ключевых видов животных, как крупные стадные копытные и бобры; во-вторых, это те элементы суши, где относительно автономно осуществляются вещественно-энергетические процессы (Методические подходы…, 2010).
5. Сбор натурных данных о растительном и почвенном покрове, почвообразующих и подстилающих породах, о рельефе, уровне грунтовых вод малого речного бассейна (бассейнов) модельной территории; сопоставление полученных данных с картографическими материалами. Выделение элементов в каждом из названных слоев, их типизация и сопряженный анализ элементов разных типов в разных слоях; выявление наиболее часто встречающихся сочетаний.
6. Типизация экотопов на основе сопоставления элементов, выделенных в слоях почвообразующих и подстилающих пород, рельефа и уровня грунтовых вод. При этом желательно разработать шкалу балловых оценок глубины преобразования экотопов в результате предшествующей хозяйственной деятельности (мелиорации, смыва грунтов, распашек, пожаров и пр.).
7. Типизация биот экосистем еще не разработана и это одна и сложных задач; на современном уровне возможна типизация растительности, ее можно проводить на основе разных классификаций. В настоящее время в России используют как принятую международным сообществом эколого-флористическую классификация (Миркин, Наумова, 1998; Вебер и др., 2005), так и отечественную доминантную классификацию (Нешатаев, 2001), а также разработанную для европейской России эколого-фитоценотическую классификацию (Мониторинг…, 2008, http://mfd.cepl.rssi.ru/flora). Она позволяет установить соответствие между синтаксонами эколого-флористической и доминантной классификаций, и удобна для оценки сукцессионного статуса экосистем в пределах лесного пояса европейской России (Мониторинг…, 2008).
8. Типизация почв экосистем, отражающих их сукцессионный статус, не разработана; одна из актуальных задач – определение диапазона почвенных разностей в пределах квазиклимаксовой экосистемы, сформированной ключевыми видами только деревьев. При этом наряду со структурными признаками все большее значение приобретает определение уровня питательного режима почв и его дифференциация по типам микроместообитаний (Методические подходы…, 2010).
9. Формирование сукцессионных рядов в пределах экотопов одного типа по степени выраженности системы микро- и мезоместообитаний, развивающихся в процессе жизни и смерти деревьев. Поскольку аутогенные сукцессии длятся в течение жизни нескольких (многих) поколений деревьев, единственная возможность исследовать сукцессионные ряды – рассматривать пространственные ряды как временные. При этом осложнения возникают в связи с невозможностью доказать абсолютную идентичность экотопов экосистем в анализируемом ряду как по их природным особенностям, так и по особенностям предшествующих нарушений. Решение проблемы состоит как во все более детальном проникновении в историю природопользования, так и в увеличении числа исследованных сукцессионных рядов на сходных экотопах для отделения общих закономерностей от частных особенностей. Эту сложную проблему каждый исследователь вынужден решать заново, четко обозначив, до какой степени детальности он считает необходимым оценивать этап развития экосистемы. Для лесных экосистем европейской России на основе исследования растительности и, отчасти, почв разработан комплекс признаков-индикаторов сукцессионного статуса (Мониторинг…, 2008), который можно использовать и для других территорий.
В качестве примера рассмотрим выявленные к настоящему времени признаки-индикаторы сукцессивных и квазиклимаксовых экосистем бореальных лесов Европейской России, которые включают характеристики растительности и почв (Мониторинг…, 2008).
Признаки-индикаторы сукцессионного и квазиклимаксового состояния растительности объединены в две группы: 1 — структурное разнообразие, 2 – таксономическое разнообразие.
Структурное разнообразие синузии деревьев оценивается по набору признаков, характеризующих особенности их популяционной жизни как ключевых видов. Это:
(1) полнота онтогенетических спектров популяций;
(2) присутствие в составе древесного яруса видов разных популяционных стратегий;
(3) полный набор онтогенетических парцелл;
(4) полный набор микросайтов, формирующихся в результате жизни и смерти деревьев.
Структурное разнообразие синузий кустарников, кустарничков и трав оценивается по структуре доминирования, а также по наличию и доминированию видов разных эколого-ценотических групп. Для темнохвойных лесов Европейской России выявлена последовательность смены доминирующих эколого-ценотических групп в ходе сукцессий. Показано, что на завершающих стадиях сукцессий и в квазиклимаксе по числу видов и по покрытию господствуют высокотравные виды из бореальной, неморальной и нитрофильной групп. Структура доминирования меняется от моно- и олигодоминантных синузий на ранней стадии сукцессий к полидоминантным синузиям в квазиклимаксе (Восточноевропейские…, 2004; Смирнова и др., 2006; Мониторинг…, 2008).
Таксономическое разнообразие оценивается
(1)для совокупности всех синузий растений одновременно,
(2) для каждой синузии (деревьев, кустарников, кустарничков и трав среди сосудистых растений, моховидных и лишайников) отдельно. Для этого используют два показателя альфа-разнообразия: видовая насыщенность – число видов на единицу площади и видовое богатство – общее число видов в синузии или в растительном сообществе. От начальной к конечной стадии сукцессии в лесных экосистемах видовая насыщенность и видовое богатство возрастают. На поздних стадиях и в квазиклимаксе (после формирования полноценной парцеллярной структуры и ВПК) значения этих показателей максимальны.
Признаки-индикаторы сукцессионного и квазиклимаксового состояния почв:
1) Структура почвенного профиля: на начальных стадиях сукцессий – формирование протяженного гумусового горизонта как результата жизнедеятельности биоты экосистемы, и наличие контрастных горизонтов; на средних стадиях — формирование мозаичной структуры профиля за счет оборачивания почвы вывалами раннесукцессионных, а затем первым поколением позднесукцессионных видов деревьев; на поздних стадиях и в квазиклимаксе — формирование недифференцированного почвенного профиля, представленного гумусовым горизонтом, в результате многократного оборачивания почвы вывалами позднесукцессионных видов; расположение гумусового горизонта непосредственно на почвообразующей породе. По мере формирования недифференцированного профиля в ходе сукцессий меняются и типы почв.
2) Мощность гумусового горизонта: на начальных стадиях – минимальная, соответствующая глубине проникновения поверхностных всасывающих корней деревьев и трав; на средних стадиях – соответствующая глубине проникновения в почву поверхностных и частично — якорных корней; на поздних стадиях и в квазиклимаксе – максимальная, соответствующая глубине проникновения в почву основной массы якорных корней деревьев.
3) Признаки давности антропогенных воздействий, такие как пожары, распашки, выпас скота в лесу. На начальных стадиях сукцессий после пожаров в почве есть осветленный горизонт со слоями углей; на средних – угли можно найти в отсыпке вывалов раннесукцессионных видов; на поздних и в квазиклимаксе – углей нет или отдельные угли встречаются в нижней части отсыпки вывалов позднесукцессионных видов.
На начальных стадиях сукцессий после распашек есть поверхностный осветленный почвенный горизонт с четкой и ровной границей между пахотным и нижележащим горизонтом; на средних – поверхностный горизонт представлен мозаикой осветленных и гумусированных фрагментов; на поздних – отдельные морфоны гумусового горизонта имеют белесую присыпку – следы преобразованного поверхностного осветленного горизонта (Бобровский, 2004; 2010; Мониторинг…, 2008).
Опыт работы в бореальных и гемибореальных лесах Европейской России показал, что минимальная длительность первичной (пирогенной) аутогенной сукцессии, проводящей к формированию квазиклимакса, измеряется суммарной длительность жизни одного поколения раннесукцессионных и двух-трех поколений позднесукцессионных видов. В астрономическом времени это составляет 600-1000 лет. За это время не только растительность, но и почва могут восстановиться до устойчивого состояния (Методические подходы…, 2010).
Несомненно, углубление представлений о климаксе и сукцессиях как процессах развития экосистем в целом приведет к увеличению числа диагностических признаков, поскольку многие компоненты экосистем не проанализированы в этом отношении. Сложность анализа определяется практически полным отсутствием в лесном поясе Европейской России как квазиклимаксовых экосистем, так и экосистем, претерпевающих автогенное развитие.
Наиболее актуальны сейчас исследования почвенной биоты и почв в популяционных мозаиках древесных видов. Они позволят выявить признаки-индикаторы сукцессионного состояния экосистем в почвенной биоте и углубить представления о сукцессионном статусе почв. Дальнейшие исследования приуроченности подчиненных видов к популяционным мозаикам древесных видов позволят разделить подчиненные виды биоты на группы индикаторных и индифферентных видов, а также описать мозаики (порои кабанов, окна в древостое, создаваемые листо- и хвоегрызущими насекомыми, грибами), создаваемые видами, занимающими промежуточное положение.
10. Размеры постоянных пробных площадей для стационарных исследований сукцессионного состояния экосистем следует определять, учитывая степень их внутриценотической гетерогенности; нецелесообразно закладывать пробные площади одинакового размера для характеристики разных этапов сукцессий. На каждом этапе размер пробной площади определяют методом увеличивающихся площадок (внутри физиономически однотипного контура растительного покрова); но на ранних стадиях аутогенных сукцессий, а также при аллогенном развитии и дигрессиях размер пробной площади увеличивают до тех пор, пока не перестанет увеличиваться число видов высших растений (как наиболее легко учитываемой группы). На средних и поздних стадиях аутогенных сукцессий размер пробной площади увеличивают до тех пор, пока не только видовое, но и структурное разнообразие, представленное ВПК и онтогенетическими парцеллами, не перестанет увеличиваться. На стадии квазиклимакса к уже перечисленным признакам добавляют признак «устойчивое состояние онтогенетических спектров популяций» всех позднесукцессионных видов деревьев (Уранов, Смирнова, 1969; Ценопопуляции…, 1988). На постоянных пробных площадях одновременно надо провести оценку экологических режимов в основных типах ВПК и онтогенетических парцелл; определить их параметры; описать почву и почвенную биоту; наземные растения, лишайники и грибы; определить биомассу видов анализируемых групп и их химический состав.
11. Большая длительность аутогенных сукцессий (по модельным расчетам в бореальных и гемибореальных лесах европейской России – 600-1000 лет) существенно осложняет подбор модельных территорий для их исследования, ибо последние тысячелетия (начиная с бронзового века) – время активных антропогенных преобразований биогеоценотического покрова (Восточноевропейские…, 2004; Мониторинг…, 2008). В связи с этим, необходимо отличать экосистемы, находящиеся на разных этапах аутогенных сукцессий, и экосистемы, претерпевающие аллогенное развитие. Для последних характерно отсутствие согласованных изменений в формировании популяционных мозаик, например, отсутствие элементов ВПК и окон, имматурных и виргинильных онтогенетических парцелл в старовозрастных лесах в связи с выборочными рубками или выпасом скота в лесу; отсутствие подроста позднесукцессионных видов под пологом раннесукцессионных видов в связи с частыми низовыми пожарами и пр. Наиболее значимы для разграничения аутогенных и аллогенных экосистем хорошо документированные сведения об истории природопользования, например, материалы генерального межевания с картами угодий (Оценка…, 2000).
12. Решение задач переориентации лесопользования на более полную реализацию основных экологических функций экосистем и их ландшафтных комплексов может быть получено при математическом моделировании. Среди моделей, основанных на представлениях gap-mosaic concept, в наиболее полной мере представления популяционной биологии учтены в комплексе имитационных моделей FORRUS-S (Чумаченко, 1998; Чумаченко, Смирнова, 2009), который широко используют для решения экологических задач лесопользования (Мониторинг…, 2008). Для его дальнейшего развития необходимо получить более полные натурные данные об изменениях биоты и почв экосистем в ходе аутогенных сукцессий.
Для реализации предложенной последовательности действий надо решить обозначенные далее задачи.
Задачи на уровне экосистем:
1. Сбор натурных данных о составе, структуре, динамике, продуктивности и химическом составе растительности, животного населения, грибов, цианобактерий; о структуре и о физическом и химическом составе почв, почвообразующих и подстилающих пород; об особенностях рельефа и уровне грунтовых вод всех типов экосистем модельной территории.
3. Получение круглогодичной информации об основных экологических параметрах местообитаний, включая параметры микроклимата и гидрологического режима на уровнях как микросайтов и парцелл, так экосистем в целом.
4. Демографический анализ популяций ключевых видов и видов-индикаторов экологических режимов в основных типах лесных экосистем.
5. Составление интегральной базы данных экосистемных параметров.
6. Интегральная оценка сукцессионного статуса лесных экосистем.
7. Разработка моделей динамики лесных экосистем при спонтанном развитии и при разных способах природопользования.
Задачи на уровне комплекса экосистем в пределах модельных речных бассейнов малых рек:
1. Выбор для исследования модельных бассейнов и оценка взаимосвязи растительности, почв и экотопов.
2. Оценка видового разнообразия биот модельных речных бассейнов и сопоставление этих данных с региональным пулом.
3. Определение параметров гидрологического режима (стока и просачивания, уровня снегового покрова) модельных бассейнов разных типов.
4. Определение микроклиматических параметров автономных, транзитных и аккумулятивных ландшафтов в пределах модельных бассейнов разных типов.
5. Количественная оценка круговорота основных элементов минерального питания на территориях модельных бассейнов с эталонным и нарушенным лесным покровом.
6. Создание баз данных биологических и экологических параметров экосистем и их комплексов в пределах модельных бассейнов разных типов
7. Составление современных и прогнозных электронных карт растительности и почв модельных бассейнов разных типов
8. Использование современных карт растительности и почв модельных бассейнов для верификации космоснимков модельных территорий.
9. Модельные расчеты скорости восстановления биоты и почв климаксового типа на территории модельных бассейнов при разной пространственной структуре лесного покрова
Задачи на уровне комплекса малых речных бассейнов
1. Определение размеров, состава и структуры комплекса малых речных бассейнов, способных в полной мере осуществлять основные экологические функции (климаторегулирующую, водоохранную, почвообразующую и функцию сохранения биологического разнообразия) в эталонных природных лесах и в лесах с разными системами лесопользования
2. Разработка моделей динамики лесного покрова комплекса малых речных бассейнов в лесах ООПТ и в промышленных лесах с разными системами природопользования.
3. Подготовка серии эколого-экономических проектов систем природопользования и охраны для лесов модельной территории
VII. Заключение
Пересмотр основных представлений экологии экосистем с позиций популяционной парадигмы позволил
· разработать основные положения теории организации экосистемного покрова и составляющих его элементов: экосистем и их комплексов;
· выделить основные типы экосистемного покрова, сформированного на основе природных закономерностей и разной степени антропогенных преобразований;
· предложить алгоритм исследования экосистемного покрова лесов с помощью стационарных исследований модельных территорий на основе разработанной методологии и согласованной системы методов.
Такой подход, с одной стороны, может стимулировать исследователей к более глубокому проникновению в механизмы структурно-функциональной организации и динамики экосистемного покрова. С другой стороны, он позволяет организовывать экспериментальные работы по реализации полученных представлений в виде региональных моделей единой стратегии охраны природы и природопользования. Необходимо четко осознавать, что реализация предлагаемой стратегии требует огромных усилий и длительного времени и ни в коей мере не противоречит решению конкретных тактических задач по увеличению доли территорий, выведенных из хозяйственного использования.
Литература
1. Алейников А. А. Динамика растительного покрова долин малых рек в результате строительной деятельности бобров / Вестник МГУ Леса – Лесной вестник. 2010. №3. С. 165-168.
2. Бобровский М.В. 2004. Лесные почвы: биотические и антропогенные факторы формирования // О.В. Смирнова (ред.). Восточноевропейские леса. История в голоцене и современность. Кн. 1. М.: Наука. C.381–427.
3. Бобровский М.В. Лесные почвы Европейской России: биотические и антропогенные факторы формирования М. 2010. КМК. 421 с.
4. Вебер Х.Э., Моравец Я., Терийя Ж.-П. Международный кодекс фитосоциологической номенклатуры. 3-е изд. // Растительность России. 2005. № 7. С.3-38.
5. Восточноевропейские леса: история в голоцене и современность / Под ред.О.В. Смирновой. М., 2004. Кн.1. 479 с. Кн.2. 575 с.
6. Восточноевропейские широколиственные леса / Под ред.О.В. Смирновой. М., 1994. 364 с.
7. Глазовская М.А. Геохимические основы типологии и методики исследований природных ландшафтов. Смоленск, 2002. 288 с.
8. Горшков В.Г., Макарьева А.М. К вопросу о возможности физической самоорганизации биологических и экологических систем//Доклады РАН. 2001. №378, С. 570-573.
9. Горшков В.Г., Макарьева А.М., Лосев К.С. Планетарный парниковый эффект и биотическая устойчивость климата Земли //Известия РАЕН. Секция наук о Земле, 2001. №7. C. 62-74.
10. Горшков В. Г., Макарьева А. М. Биотический насос атмосферной влаги, его связь с глобальной атмосферной циркуляцией и значение для круговорота воды на суше. Петербургский институт ядерной физики РАН. Гатчина. 2006. Препринт 2655. 54 с.
11. Гревцев, В. Н. Состояние ресурсов бобра в России, их размещение, использование и воспроизводство // Современные проблемы природопользования, охотоведения и звероводства: Мат. Межд. науч-практ. конф., посвящ. 80-лет. ВНИИОЗ, Киров, 28-31 мая 2001г. — Киров, 2002. — С. 197 — 200.
12. Завьялов, Н. А. Влияние речного бобра на экосистемы малых рек М.: Наука, 2005.186 с.
13. Колбовский Е.Ю. Ландшафтоведение. М., 2006. 408 с.
14. Коротков В.Н. Новая парадигма в лесной экологии // Биологические науки. 1991. №8. С. 7-20.
15. Корчагин А.А. Строение растительных сообществ // Полевая геоботаника. Л.: Наука, 1976. Т. 5. 320 с.
16. Методические подходы к экологической оценке лесного покрова бассейна малой реки / Отв. ред. Л.Б.Заугольнова, Т. Ю. Браславская. М.: 2010. КМК. 482 с.
17. Миркин Б.М., Розенберг Г.С., Наумова Л.Г. Словарь понятий и терминов современной фитоценологии. М.: Наука, 1989. 223 с.
18. Миркин Б.М., Наумова Л.Г. Наука о растительности (история и современное состояние основных концепций). Уфа, 1998. 413 с.
19. Мониторинг биологического разнообразия лесов России /Под ред.А.С.Исаева. М., 2008. 453 с.
20. Нешатаев В.Ю. Проект Всероссийского кодекса фитоценологической номенклатуры // Растительность России. 2001. №1. С. 60-72.
21. Одум Ю. Экология. М.: Мир, 1975. 740 с.
22. Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России (под ред. Л.Б.Заугольновой). М.: Научный мир. 2000. 185 с.
23. Смирнова О.В. Популяционная организация биоценотического покрова лесных ландшафтов // Успехи совр. биол. 1998. № 2. С. 25-39.
24. Смирнова О.В. Методологические подходы и методы оценки климаксового и сукцессионного состояния лесных экосистем (на примере восточноевропейских лесов) // Лесоведение. 2004. №3. С. 15-26.
25. Смирнова О.В., Чистякова А.А., Попадюк Р.В. и др. Популяционная организация растительного покрова лесных территорий (на примере лесов европейской части СССР). Пущино: ОНТИ НЦБИ АНСССР. 1990. 92 с.
26. Смирнова О.В., Заугольнова Л.Б., Попадюк Р.В. Популяционная концепция в биоценологии // Журн. общ. биол. 1993. Т 54, №4. С.438-448.
27. Смирнова О.В., Попадюк Р.В., Заугольнова Л.Б., Ханина Л.Г. Оценка потерь флористического разнообразия в лесной растительности (на примере заповедника «Калужские засеки») // Лесоведение. 1997. № 2. С.27-42.
28. Смирнова О.В., Турубанова С.А., Бобровский М.В., Коротков В.Н., Ханина Л.Г. Реконструкция истории лесного пояса Восточной Европы и проблема поддержания биологического разнообразия // Успехи современной биологии. 2001. Т.121. № 2. с. 144-159
29. Смирнова О.В., Заугольнова Л.Б., Ханина Л.Г., Бобровский М.В., Торопова Н.А. Популяционные и фитоценотические методы анализа биоразнообразия растительного покрова // Сохранение и восстановление биоразнообразия. Учебно-методическое издание. М.: Изд-во НУМЦ, 2002. С. 145 — 194.
30. Смирнова О.В., Торопова Н.А. 2008. Сукцессия и климакс как экосистемный процесс // Успехи соврем. биол. Т.128. №2. С. 129-144.
31. Соколов В.Е., Филонов К.П., Нухимовская Ю.Д., Шадрина Г.Д. Экология заповедных территорий России. М.: Янус-К. 1997. 576 с.
32. Сукачев В.Н. Растительные сообщества: введение в фитосоциологию. М.-Л.: Книга, 1928. 232 с.
33. Сукачев В.Н. Избранные труды. Проблемы фитоценологии. Л.: Наука. 1975. Т.3. 543 с
34. Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия / Под ред. О.В.Смирновой, Е.С.Шапошникова. СПб.: РБО, 1999. 549 с.
35. Удра И.Ф. Расселение растений и вопросы палео- и биогеографии. Киев: Наукова думка, 1988. 200 с.
36. Удра И.Ф. Расселение и миграция древесных растений в умеренном поясе Евразии. Науч. докл. на соис. уч. ст. д.б.н. М., 1990. 39 с.
37. Уранов А.А., Смирнова О.В. Классификация и основные черты развития популяций многолетних растений //Бюл. МОИП. Отд. биол. 1969. Т. 79. Вып. 1. С. 23-30.
38. Ценопопуляции растений (основные понятия и структура). М.: Наука, 1976. 216 с.
39. Чумаченко С.И. Моделирование динамики разновозрастных многовидовых лесных ценозов // Журн. общ. биол., 1998. Т. 59, № 4. С. 363-376.
40. Чумаченко С.И., Смирнова О.В. Моделирование сукцессионной динамики насаждений //Лесоведение 2009.№6, С. 3-17.
41. Ярошенко А.Ю., Потапов П.В., Турубанова С.А. Малонарушенные лесные территории европейского севера России, М., 2001. 75 с.
42. Aksenov D., Dobrynin D., Dubinin M. Atlas of Russia’s Intact Forest Landscapes. Moscow, 2002. 182 p.
43. Braun-Blauquet J, Pavillard J. Vocabulaire de sociologie vegetale. 2 ed. Montpellier, 1925, 22 p.
44. Crain C.M. & Bertness M.D. Ecosystem engineering across environmental gradients: implications for conservation and management // Bioscience. 2006. Vol. 56, P. 211–218.
45. Disturbance Dynamics in Boreal Forest: Defining the Ecological Basis of Restoration and Management of Biodiversity. 2002. Vol. 36, №1, 447 p.
46. Gutiérrez J.L., Jones C.G. Physical ecosystem engineers as agents of biogeochemical heterogeneity // BioScience. 2006. Vol. 56, P. 227–236.
47. Gurney, W.S.C. & Lawton, J.H. The population dynamics of ecosystem engineers. // Oikos, 1996, V.76, P. 273–283
48. Hastings A., Byers J.E., Crooks J.A., Cuddington K., Jones C.G., Lambrinos J.G., Talley T.S., Wilson W.G. Ecosystem engineering in space and time. Review and synthesis // Ecology letters. 2008 Vol. 10, P. 153-164.
49. Jones C.G, Lawton JH and Shachak M. Organisms as ecosystem engineers // Oikos. 1994. Vol. 69, Р. 373-386
50. Jones C.G, Lawton J.H. and Shachak M. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 1997. 78: 1946-1957
51. Kuuluvainen T. Gap disturbance, ground microtopography, and the regeneration dynamics of boreal coniferous forests in Finland: a review // Ann. Zool. Fennici. 1994. Vol. 31. P. 35–51.
52. McCarthy J. Gap dynamics of forest trees: A review with particular attention to boreal forest // Environ. Rev., 2001, Vol. 9, P 1-59.
53. Smirnova, O.V., Zaugolnova, L.B., Istomina, I.I., Khanina, L.G. Population mosaic cycles in forest ecosystems. In: Proceedings IAVS Symposium. IAVS, Opulus Press, Uppsala. 2000. P. 108-112.
54. The ecology of natural disturbance and patch dynamics. Orlando, 1985. 472 p.
55. The mosaic-cycle concept of ecosystem. /Ed. Remmert H. 1991. Berlin, Heidelberg, N-Y. 168 p.
56. Tropical trees as living systems. Cambridge etc: Univ. press,1978. XXVIII, 675 p.
57. Vera F.W.M. Grazing ecology and forest history. Oxon-New York: CABI Publ., 2000. 506 p.
58. т Watt A.S. 1925. On the ecology of British beech woods, with special reference to their regeneration. II. The development and structure of beech communities on the Sussex Downs // J. Ecol. Vol.13. P.27–73.
59. Yamamoto S. 1992. The gap theory in forest dynamics // J. Plant Res. Vol.105. P.375–383.
Hасширенный текст доклада на семинаре «Актуальные задачи Биологии охраны природы на Северо-Западе Российской Федерации», г. Вологда, 2 декабря 2010 г.
Источник naturschutz
Рекомендуем прочесть
 Причина гибели сайгаков в Казахстане
Причина гибели сайгаков в Казахстане Закон о «запрете» заемного труда дискриминирует одиноких и многодетных родителей
Закон о «запрете» заемного труда дискриминирует одиноких и многодетных родителей Экосистемные последствия лесоэксплуатации: Восточная Финляндия vs Карелия
Экосистемные последствия лесоэксплуатации: Восточная Финляндия vs Карелия Приступ франкофобии
Приступ франкофобии Влияние «Великой Рецессии» на неравенство материального положения в обществе с учетом возрастной и расовой принадлежности
Влияние «Великой Рецессии» на неравенство материального положения в обществе с учетом возрастной и расовой принадлежности Горим…
Горим… «На женском дне»: старая статья на фоне советского акмэ
«На женском дне»: старая статья на фоне советского акмэ Социально-когнитивные способности антропоидов: эволюционные предпосылки нравственных основ сотрудничества у людей
Социально-когнитивные способности антропоидов: эволюционные предпосылки нравственных основ сотрудничества у людей
