

Красный и жёлтый самец мексиканского чечевичника Carpodacus mexicanus на кормушке в г.Феникс
Аннотация. Урбанизация, распространяющаяся вдоль «городского градиента», «с точки зрения» реагирующих видовых популяций, представляет собой некий вызов, ответ на который подчиняется своего рода правилу Ле Шателье. Вид или приспосабливается (немедленно либо с запозданием в случае возвратной урбанизации), меняя «конструкцию» популяционной системы в сторону большей «открытости» и «проточности» группировок, большей лабильности структуры системы, с соответствующей лабилизацией жизненной стратегии. урбанизация вызывается в каждом регионе в отдельности, и не столько развитостью «центрального ядра», сколько плавностью переходов от него к низовой урбанизации (через города 2-го, 3-го и т.д. уровня в соответствующей агломерационной системе, не случайно так напоминающей планетную).
Чем более консервативной была жизненная стратегия вида первоначально, тем большая плавность переходов потребуется для вызывания урбанизации. В этом случае городской градиент должен быть менее крутым, но более длинным. Если он будет круче – вид начнёт отступать на периферию региона, придерживаясь в процессе отступления исходных биотопов до тех пор, пока городской градиент не придёт в соответствие с параметрами, оптимальными для урбанизации именно данного вида. Тогда запускается возвратная урбанизация (после перестройки популяционной структуры).
Коллега ishs год назад спрашивал меня, “что мешает вяхирю из Хамины жить в Выборге, ведь там же совсем рядом”? Попробую поподробнее рассказать, «какие такие барьеры» мешают птицам урбанизироваться, когда они падают и почему.
Важно подчеркнуть, что подобные «камни преткновения» есть у городских популяций самых разных видов птиц. Их быстрое распространение как бы упирается в невидимую границу и десятилетиями топчется там, не умея освоить, казалось бы, «точно такие же» города по ту сторону. Затем вдруг, в самый неожиданный момент, она преодолевается разом, и урбанизация вида охватывает новые территории.
Скажем, урбанизированные вяхири несколько десятков лет «не могли преодолеть[1]» всего лишь 50 км от польского г.Ольштына, где урбанизация случилась в середине ХХ века, до Калининграда, где она началась лишь в 1990-е[2]. То же относится к вяхирям городов юго-восточной Польши, «не осилившим» небольшую дистанцию до г.Львова, где урбанизация началась только в 90-е и была полностью задокументирована. Сейчас вид быстро урбанизируется по всей правобережной Украине, причём освоение урболандшафта обнаруживается одновременно и в разных местах.

Вяхирь в городе
Почему так, что определяет быструю урбанизацию вида в одном регионе, и медленность (или невозможность) данного процесса – в другом, где города на первый взгляд «точно такие же»?
Тогда я дал наиболее общий ответ, сейчас умею его конкретизировать до возможности предсказания шансов (или, напротив, препятствий) на начало урбанизации данного вида птиц в данном староосвоенном регионе[3]. И, в случае если урбанизационный процесс «запустится» — до прогноза средней скорости и степени полноты урбанизации данного вида.
Если вспомнить классификацию причин Аристотеля, все примерно согласны относительно формальных причин урбанизации «диких» видов птиц. Это попытка расширить спектр местообитаний, освоив новый ландшафт, с его уникальными плюсами, отсутствующими, между прочим, не только в природных ландшафтах, но и в менее динамичных антропогенных.
Другое дело, поскольку этот процесс суть элементарное эволюционное явление, продолжаются споры о его механизме. То ли это селективный процесс, с распространением полезных адаптаций отбором? То ли преадаптация некоторых видов и/или категорий особей к урбосреде (включая прямое приспособление за счёт поведенческой и прочей пластичности)? [Наиболее интересный вариант селекционистской ТЗ даёт парфорсная модель эволюции В.В.Суслова. В.К.].
Или это номогенез – направленное преобразование пространственно-этологической структуры всей региональной популяции вида в ответ на «давление» растущего «городского ядра» на местообитания вида в регионе, и вызванное этим увеличение средового стресса? Или, что то же самое, направленная перестройка «конструкции» системы в сторону большей устойчивости в сильно более нестабильной урбосреде с «островными» и очень динамичными местообитаниями. По ходу эволюции урболандшафта они «делятся», «движутся», исчезают в одних местах и появляются совсем в новых. См.пп.7-8 тут.
Соответственно, население городских популяций всех видов птиц (независимо от исходной биологии особей и «конструкции» популяционных систем в их естественных биотопах) вынуждено постоянно «перебрасываться» из одних поселений в других (без задержки, а лучше всего – с упреждением). Чтобы сполна использовать плюсы урболандшафта, не попадая при этом под удар рисков, не оказываясь в «экологических ловушках» и пр. Соответственно, пространственно-этологическая структура городских популяций самых разных видов птиц преобразуется в одну и ту же сторону, так что «конструктивно» оказывается полной противоположностью структуре популяций негородских (тех же видов): и не только в природных, но и в с/хландшафтах[4].
См.таблицу 1. (полностью сравнение пространственно этологической структуры урбанизированных популяций с внегородскими тут)
| Урбанизированные популяции | «Внегородские» популяции |
| «Проточность» группировок (открытые системы) | Консерватизм группировок (закрытые системы) |
| «Дальний порядок» обмена особями в популяции | «Ближний порядок» обмен особями в популяции |
| Направленная сортировка особей в популяции вдоль «городского градиента» | Только ближние и местные переселения, независимые от стратегии |
| Опережающее отражение сигналов среды, следование за средой без смены стратегии | Простое реагирование на сигналы среды, оппортунистические изменения стратегии ради сохранения членства в группировке |
| Экспоненциальный рост даже в ограниченном пространстве биотопов (плотность населения не зависит от размера «острова»), пределы роста снимаются через дальнейшую урбанизацию | Пределы роста существенны и не преодолеваются популяцией, плотность населения пропорциональная размеру «острова» |
| Наиболее устойчивый параметр популяционной структуры — соотношение приверженцев альтернативных стратегий, связанный с состоянием урбосреды | Наиболее устойчивый параметр популяционной структуры – резидентское «ядро» группировок и % особей «ядра» и «периферии» |
| «Островной синдром» в поведении развит, исключительно высокая толерантность к побеждённым и подчинённым особям, часто на фоне круглогодичной территориальности | «Островной синдром» в поведении не развит, полная интолерантность к побеждённым и подчинённым особям, они удаляются с территории победителя и часто вытесняются из группировки |
| Высокая «открытость» поведения особей к инновациям, постоянная готовности к использованию «нетипичных» местообитаний, корма, мест гнездования в уже существующем индивидуальном пространстве, или к переселению на другие «острова» в связи с динамикой урбосреды. Нечувствительность к «островному» и «барьерному» эффекту и в связи с этим устойчивость к городской среде | Открытость поведения к инновациям низкая, почти нулевая. Особи стараются занять такую территорию, так взаимодействовать с соседями и партнёром, чтобы необходимость «инноваций» была минимальной. «Островной» и «барьерный» эффект эффективно блокируют возможность перемещения раз выбранной территории и/или поиска партнёра за её пределами. Высокая уязвимость при поселении в городе |
| Активная урбанизация вида, формирование городских популяций, демографически обособленных от материнской | В случае поселения в городе – пассивные урбанисты, быстро вытесняющиеся |
Но споры об эволюционных механизмах явления будут вестись ещё долго. Куда интересней установить материальную и действующую причину процесса урбанизации «диких» видов птиц, его modus operandi. Какие (по происхождению) особи принимают участие в нём как? Тут сталкиваются два представления.
1. Урбанизация вида идёт преимущественно путём расселения «городских рас», возникших в каком-то одном месте внутри ареала и затем лишь распространяющихся по соседним городам независимо от жизни «лесных» особей того же вида в природных местообитаниях вокруг городов («модель расселения городских рас»). Скажем, урбанизация чёрного дрозда или вяхиря в Европе[5] описывается так. Городские популяции того и другого вида сперва складываются в 1820-х гг. в Рейнланде и в 1840-х гг. в Париже (почему именно там и тогда – не обсуждается), а затем расселяются по городам, сходным с ближайшим центром урбанизации.
Соответственно, местные «лесные» птицы того же вида во вновь заселяемых регионах в урбанизации участия не принимают. Разве что в самый последний момент, когда местность изменится так, что биотопов «лесной» популяции (в смысле крупных, нефрагментированных массивов природных ландшафтов) останется недостаточно для поддержания сколько-нибудь жизнеспособных популяций. Тогда вид будет представлен преимущественно «городскими» особями, «лесные» же воленс-ноленс примкнут к ним[6]. При таком механизме «волна урбанизации» не может проникнуть в регионы, уж слишком культурно различные – скажем, волны урбанизации, начавшиеся в Западной Европе, останавливаются на границе Восточной.
Почему-то оно очень популярно в Европе, несмотря на довольно слабое соответствие фактам в подавляющем большинстве случаев. Даже урбанизацию чёрного дрозда пытаются объяснить таким образом, хотя для большинства регионов урбанизация местных «лесных» птиц была прямо показана.
2. Модель «урбанизации местных «лесных» особей». «Городское ядро» региона заселяется из лесов, лугов и т.д. природных ландшафтов ближних пригородов, после того, как те окажутся под «давлением» пяти главных видов биотопических изменений, запускаемых в регионе развитием урболадшафта «ядер» агломераций[7].
Это фрагментация массивов природных ландшафтов извне с инсуляризацией изнутри, изоляция с «дроблением» возникающих «островов» и стохастизация их размещения по территории, см. городской градиент рисунка 1 (2-4: инсуляризация и преобразование с краёв идут одновременно с фрагментацией, см.тут). Иными словами, «лоскутное одеяло» природных ландшафтов чем ближе к «ядру», тем сильней фрагментировано извне (дорогами и застройкой), инсуляризируется изнутри (дорожно-тропиночной сетью, просеками, сенокосами и пр.), а полученные «острова» трансформируются с краёв.
Кроме того, по мере «дробления» взаимное расположение новообразованных «островов» природных ландшафтов всё больше стохастизируется: первоначально они располагались группами, 2-3 мелких «острова» вблизи наиболее крупного, то с уменьшением размеров и увеличением дистанции «островов» друг от друга их размещение всё больше оказывается случайным. См.подробнее тут.
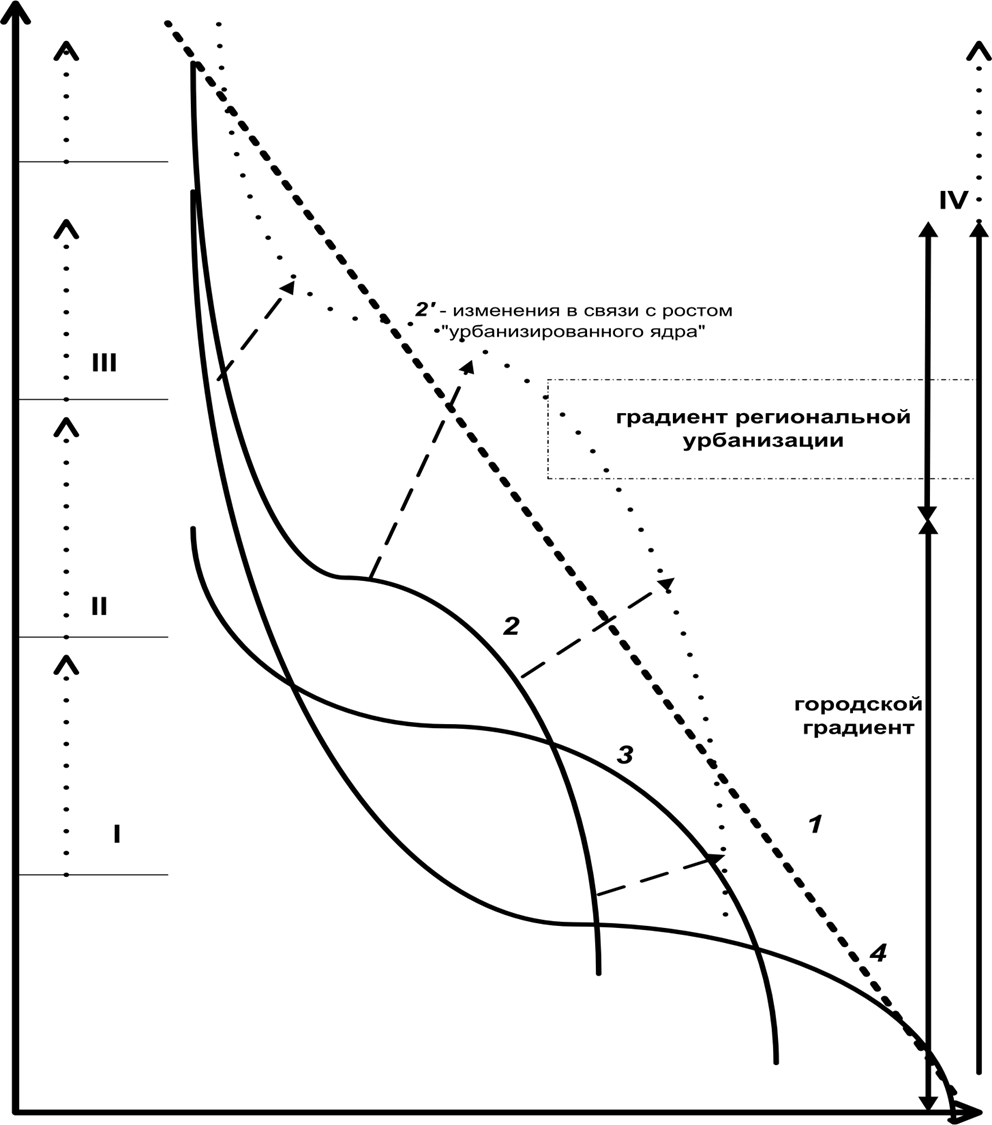
Рисунок 1. Городской градиент: падение степени урбанизации территории (1) и степени изменённости городом природных ландшафтов, представляющих собой естественные местообитания вида (2-4) от центра города (I) к его границе (II) и далее к периферии зоны влияния города, III. IV – радиус центральной агломерации и потенции его изменения в процессе роста урбанизированного «ядра» (штрих-пунктирные стрелки). Пунктирная стрелка – вектор изменений урбосреды в процессе городского развития.
Трансформация видовых местообитаний после попадания в зону воздействия города и включения соответствующих биотопов отдельными участками в структуру урболандшафта (на следующем этапе расширения города). 2 — фрагментация непрерывных местообитаний вида в «зоне влияния» города, образование «островного ландшафта». 3 — изоляция «островов» улицами, коммуникациями, городскими кварталами, усиление «краевого эффекта», сокращение площади отдельных «островов» и увеличение их изолированности. 4 — дробление и разъединение «островов», переход от закономерного расположения «островов» относительно друг друга к целиком стохастическому.
Этими пятью процессами урбанистической трансформации ландшафта исходный «материк» естественных местообитаний превращается в «архипелаг». «Острова» последнего имеют динамику, сугубо неблагоприятную для большинства видов: они сокращаются в площади, всё более изолируются друг от друга. По мере «измельчения осколков» фрагментации они переходят от группового размещения в пространстве[8] ко всё более случайному и независимому друг от друга. Это верно для всех типов фрагментов, возникающих при дроблении «материка»: лесных, луговых, и водно-болотных.
Все пять процессов подобны борзым, «бегущим перед» урбанизационным процессом и «дробящие» исходный ландшафт на фрагменты, позднее включающиеся в структуру собственно города (по мере территориального роста последнего). Частные градиенты, выделенные Б.Клауснитцером, представляют собой как бы проекции общего тренда преобразования местообитаний вдоль «нашего» городского градиента на динамику отдельных типов ландшафтов[9].
Интенсивность всех пяти падает вдоль «городского градиента», протягивающегося по радиальному срезу урбанизированного региона, от «ядер агломерации» к периферии. Малонарушенная периферия начинается за границей зоны влияния города, устанавливающейся средней дальностью выездов горожан на уик – энд. Или по аэрофотоснимкам – линией, за которой чёткие многоугольники лесных, луговых и т.д. природных участков, фрагментированных полями, дорогами и застройкой, сменятся природными границами между ними, которые плавны, извилисты, предполагают экотонные опушечные местообитания и пр.
Город «распространяется в регион» посредством автомобильных дорог и других линейных инфраструктур, регрессивное развитие сети которых стимулирует появление новых «пятен» урбанизации разного масштаба, от микроурботерриторий до участков уже городской застройки, закладки новых микрорайонов или городов-спутников и пр.
Сказанное в равной мере касается «ядер» и зависимых городов агломераций, просто размеры «своих» регионов и пространственные масштабы урбанизации у них разные. Интенсивность антропогенных преобразований ландшафта рис.1 падает центробежно вдоль городского градиента, причём собственно фрагментация с сопутствующими процессами (рис.1, 2) опережает растущее разъединение «островов» неприродным урбанистическим матриксом[10], существенно менее проницаемым для птиц, чем даже с/х ландшафт (рис.1, 3). Последнее, в свою очередь, опережает в пространстве и во времени стохастизацию размещения «островов» (рис.1, 4).
Однако по мере прогресса урбанизации региона появляется и растёт «архипелаг» техногенных аналогов сокращающихся природных местообитаний: пустырей вместо лугов, старых парков и лесопарков вместо лесов, рыбразводных прудов, отстойников и т.д. техногенных водоёмов вместо сокращающихся водно-болотных местообитаний (на них, также как на поймы, приходится максимум рекреационной нагрузки). То есть динамика этого «архипелага» прямо противоположна таковой исходных местообитаний вида – здесь «острова» растут в площади, появляются в новых и неожиданных местах, сливаются между собой и пр. В пределе они формируют сеть новых рукотворных ландшафтов (с вкраплениями природных участков, которые удалось спасти от поглощения городом, вовремя придав им статус ООПТ, культурной достопримечательности и пр.).
Отсюда понятно, что единственное спасение региональных популяций «диких» видов состоит в том, чтобы вовремя «пересесть», полностью или частично, с «островов» естественных местообитаний, сокращающихся, отступающих к периферии региона, на «архипелаг» их антропогенных производных. Или техногенных аналогов. Как ни странно, особенно это верно (и наиболее насущно) для самых уязвимых видов птиц по классификации P.Angelstam & G.Mikusinski – территориально и биотопически консервативных, экологически специализированных, жёстко привязанных к крупным массивам малонарушенных природных ландшафтов (старым лесам, верховым болотам, степным участкам и пр.). Хорошо видно, что при существующих трендах урбанизации староосвоенных регионов Европы, северной Америки, Европейской части СССР и пр. сохранение таких видов в их исходных местообитаниях совершено невероятно. Действительно крупных массивов, могущих поддержать жизнеспособные популяции таких видов – раз, два и обчёлся (один-два на несколько областей).
Так, анализ метапопуляционных моделей показывает, что большинство болот Мещеры недостаточно велики или недостаточно связаны с другими болотами для поддержания жизнеспособных популяций их специфических видов птиц и (в меньшей степени) бабочек[11]. Надо или восстанавливать исходную целостность ландшафта, утраченную в ходе добычи торфа и осушительных мероприятий (скажем, затапливая бывшие с/х земли), или же соответствующие виды «для самосохранения» вынуждены будут осваивать нарушенные болота, приболотные местообитания, заболоченные участки меньших размеров – а там и в сильно изменённый человеком ландшафт выйдут. Как это происходит в ряде регионов с лесным жаворонком (юлой), серым сорокопутом и дербником (в других местах эти виды жёстко придерживаются коренных биотопов – крупных болотных комплексов и сосновых боров). Последний вид, в лесной зоне придерживающийся крупных болотных массивов, в 2005 г. загнездился в г.Иваново, причём сразу 4-мя парами[12].

Дербник, самец
Дальше, оставшиеся крупные массивы слишком изолированы друг от друга. Если популяции соответствующих видов жёстко сохранят строгую привязанность к таким местообитаниям и исходно-консервативную жизненную стратегию (исключающую выход за их пределы), то будут всё более уязвимы к воздействию островного эффекта, так что их вымирание оказывается вопросом времени. Это т.н. отложенное вымирание видов, чьи популяции вдруг оказываются на «архипелаге» вместо материка. Ход событий будет неблагоприятен, даже если все соответствующие крупные массивы будут вовремя выявлены и взяты под охрану, что само по себе очень непросто – природопользователи сопротивляются, и практически всегда существующие ООПТ это то, что удалось у них «выбить», а не то, что оптимально для сохранения целевых видов и сообществ.
В силу названного сопротивления, разного в разных странах[13], существующие ООПТ возникают позже времени, оптимального для начала охраны целевых видов и сообществ, и почти всегда меньше по площади, чем необходимо. Отсюда «спасение утопающих – дело рук самих утопающих»: единственный шанс на невымирание видов из категории угрожаемых, исчезающих, краснокнижных в староосвоенных регионах Европы/Америки/Японии будет смена жизненной стратегии (переход от консерватизма к лабильности, в ходе которого популяционная структура вида преобразуется от «негородской» «конструкции» к городской, см.табл.1).
На счастье, очень многие угрожаемые/краснокнижные виды птиц (про другие группы не знаю, но мыслится, что и там так, с поправкой на разницу в расселительных возможностях и в экологии), способны на такой «поворот». Я готов даже обосновать, что способны все, причём можно прогнозировать «время задержки» между началом сокращения популяции уязвимого вида, вызванного разрушением местообитаний растущими «ядрами» агломераций, до начала возвратной урбанизации (рис.2).
В первый период вид ещё следует исходной консервативной стратегии, во втором же – скачкообразно отказывается от неё, начинает осваивать мелкие «острова» местообитаний, возникшие вследствие фрагментации, а не отступать с них и пр. Время ожидания между первым и вторым, по всей видимости, тем больше, чем консервативней исходная жизненная стратегия вида по сравнению с близкими видами того же рода или той же экологической гильдии, насколько он «выделяется консерватизмом» среди близких родственников. Иными словами, наиболее уязвимые виды староосвоенных регионов[14] по своей биологии суть противоположность «видам-выскочкам» с наиболее лабильной стратегией, первыми урбанизирующихся, быстрей всех расселяющихся по «рукотворным ландшафтам» вроде лесополос в чуждые им природные зоны и пр.
Примеры этого многочисленны. Вот средний дятел; белоспинный дятел; чёрный аист; ряд видов хищных птиц, включая орлов; черноголовый поползень (п.8); и пр. и пр.

«Краснокнижный» средний пёстрый дятел с 1980-х гг. всё чаще встречается в старых парках, сперва на кормушках зимой, а потом и гнездится

После резкого сокращения численности в 1950-1970-х гг. «поворот» к освоению лесных микрофрагментов, лесополос и других «архипелагов» местообитаний антропогенной природы происходит у таких считавшихся урбофобными видов, как орёл-могильник, змееяд, дербник и ряд других.
Так, по Закону об охране природы ГДР 1954 года их даже было запрещено фотографировать без специального разрешения: «пусть лучше будет белое пятно в науке, чем в природе[15]». Запрещали фотографировать беркута, орлана-белохвоста, чёрного аиста, филина, дрофу и даже вóрона. Из-за высокой пугливости этих видов даже столь незначительного воздействия было достаточно, чтобы пара бросала гнездо. Сейчас эти виды, кроме первого, в Европе в большей или меньшей степени восстановили численность после возвратной урбанизации, в ходе которой существенно снизили все составляющие уязвимости для человека, в том числе и пугливость.

Тетерева на кладбище старых самолётов во Внуково

Тетёрка из этой стаи на фоне спирали Бруно
Скажем, сейчас змееяд гнездится и в зелёной зоне г.Киева. Из 5 пар змееядов Киевской области, исследованных К.А.Письменным (2008), 2 гнездились в лесах, интенсивно посещаемых людьми (всего популяция вида в области составляет около 50 пар). Это припойменные леса по правому берегу р.Ирпень, в т.ч. одна — в зелёной зоне г.Киева. Одна из этих 2-х пар в 2005 г. загнездилась в 30 м. от лесовозной дороги, по которой возили лес, вырубавшийся в 200 м от гнезда, и шло движение техники. Успешно выведя птенцов, в 2006 г. они загнездились там же и тоже успешно, несмотря на эксплуатацию дороги, и оставили территорию лишь в 2007 г., когда само гнездо было срублено, но переместились недалеко.
Самец и самка этой пары во время охоты не беспокоились при виде двигающихся и работающих людей на дистанции >40-50 м., в т.ч. потому что в районе охоты заболоченные участки чередуются с частными земельными наделами, где всегда есть работники. Охотничья территория пары включала военный полигон и новую промышленную зону близ с.Новые Петровцы, интенсивно используемые в период 2000-2008 гг. Раз самка добыла ужа на территории завода строительных смесей в рабочий день, в 30 м от административного здания.
Пара, гнездящаяся в пределах города Киева, ежегодно меняла гнездо, видимо из-за исключительного наплыва отдыхающих во время майских праздников, но держалась данной территории. В выходной день 19.08.2007 она успешно кормила птенца, несмотря на исключительный шум, созданный тренировками мотоциклистов в 600 м. на недостроенной дороге Романовка-Киев.
И даже пары, гнездившиеся в малопосещаемых местах, показывают повышенную толерантность к человеку по сравнению с ситуацией 1960-1980-х гг. Пара, гнездившаяся в 2006-2008 гг. у с.Феневичи, 2 года подряд использовала одно и то же гнездо, несмотря на визит человека с целью кольцевания. То есть присутствовавшая ранее урбофобия уменьшилась у всей популяции, а не только у особей, оказавшихся в тесном контакте с хозяйственной деятельностью, людьми и техникой[16].
Орёл-могильник на Юге России, на Украине с 1980-90-х гг. активно осваивает новые, нетипичные местообитаний. Орлы начинают гнездиться в саженых сосняках, достигающих спелого возраста; гнезда страиваются на верхушках старых сосен. Так, обследование р.Северский Донец в пределах Харьковской обл. показал рост численности именно этой, «динамичной»[17] популяции. Там же, на боровой террасе, были найдены новые гнездовые пары в Донецкой, Луганской и Днепропетровской областях.
В 2000-х гг. рост численности «динамичных» популяций вида вызвал освоение новых местообитаний (также антропогенных и «островных») за пределами боровой террасы. Птицы стали гнездиться в лесополосах, посаженных преимущественно в 1950-60-х гг., и достигших спелого возраста. Данная инновация зафиксирована одновременно на двух изолированных друг от друга территориях – в междуречье Северского Донца и в Степном Крыму.
Одновременно вид освоил иные типы лесных микрофрагментов. Так, одна пара заняла группу одиночных тополей среди лугов в низовьях р.Берека Изюмского района Харьковской обл.; две другие облюбовали небольшие байрачные леса в степных балках Барвинковского района (Ветров, Милобог, 2008).
При этом «консервативные» популяции орла-могильника (связанные с исходными видовыми местообитаниями), издавна обитающие в старых дубравах лесостепной зоны центральной части Украины, все эти годы находятся в глубокой депрессии. Настолько глубокой, что даже не наблюдается расселения молодых по степным ландшафтам окрестных областей (Одесской, Николаевской и Кировоградской), вполне подходящим для заселения.
Соответственно, тенденция к восстановлению сплошного ареала вида, чётко обозначившаяся в 1990-2000-е годы, обеспечивается именно и только экспансией «динамичных» популяций. Они обретают неуязвимость к «островному эффекту» и другим следствиям преобразования ландшафтов в ходе региональной урбанизации, которые сокращали численность вида в 1950-1970-х гг., «консервативные» же остаются уязвимыми. Понятно, поддержка каких из них стратегически выгодней для природоохранников[18].

То же самое видим у колпицы, которую долго описывали как осторожную птицу глухих дебрей в плавнях. Вид загнездился на рыбразводных прудах Тернопольской области (причём в 2008 г. одновременно 2 группы орнитологов нашли её на 2-х разных прудах[19]). Что переносит на нашу территорию процесс, уже идущий в Европе. В то же время популяция вида в исходном биотопе, в плавнях, пребывает в глубокой депрессии.
Как писал орнитолог Виталий Грищенко из Каневского заповедника, «это вообще генеральная линия» эволюции для видов, камнем преткновения для которых были взаимоотношения с человеком.
Белые цапли. После прекращения тотального истребления они через несколько десятилетий очухались и начали быстро расселяться, при этом успешно осваивая измененные местообитания. Большая белая цапля сейчас у нас обычный вид на заросших прудах и в верховьях водохранилищ. Небольшие колонии можно найти на многих сельских прудах. Малая белая цапля стала гнездиться на юге в лесополосах.
Примерно то же самое с лебедями. Сейчас на многих сельских прудах можно увидеть как минимум парочку шипунов. А с севера еще движется и кликун. На отдельных прудах, там где самые мощные заросли, умудряются гнездиться и журавли…
Что-то подобное, возможно, намечается с малым бакланом. В последние годы стали отмечаться далекие залеты. Его нашли на зимовке под Черкассами, были залеты аж в Белоруссию. Расселение, насколько я знаю, пока еще не идет, но обычно такое «бродяжничество» ему предшествует. Так что можно ожидать.
О приспособлении хищников мы с Гаврилюком писали еще в материалах 1 конференции молодых орнитологов. Появились гнезда орлана и малого подорлика на сравнительно небольших и легкодоступных деревьях. Гнезда орланов мы находили на средневозрастных соснах с кучей веток от земли не где-нибудь в плавнях, куда никто не полезет, а на склонах оврагов среди полей. В последние годы начали проводить учеты зимующих птиц на крупных рыборазводных прудах. Оказалось, что до замерзания (т.е. до середины декабря — начала января) там держатся десятки орланов. В общей сложности учитывали до 120-150 особей! Орлан после продолжительного спада численности в середине прошлого века сейчас потихоньку ее восстанавливает».
Важно подчеркнуть, что с началом возвратной урбанизации возникает мозаицизм популяции уязвимого вида. «Поворот» к освоению антропогенных фрагментов их исходных ландшафтов в зоне влияния города совершают отнюдь не все популяции. Сперва это делают лишь некоторые («динамичные»): очень быстро показывают рост численности, расширение ареала и лавинообразный рост использования нетипичных биотопов, мест гнездования, способов кормления и пр. Другая часть популяций, часто прямо в соседней местности, остаётся консервативной: по-прежнему держатся за крупные малонарушенные массивы исходных ландшафтов, с избеганием их более фрагментированых и/или изменённых человеком вариантов с жёстким сохранением традиционных для вида биотопов, мест гнездования, способов кормления и поведенческих стереотипов.
В противоположность растущим популяциям первой группы эти остаются в депрессии: не повышают численность даже при эффективной охране, не колонизируют подходящие местообитания, при умеренном нарушении их биотопов уходят из массива без попыток сохранить гнездование в данном районе за счёт менее типичных гнездовых микростаций, способов кормления и пр. Напротив, «динамичные» популяции первой группы в ответ на нарушение и частичное уничтожение местообитаний человеческой деятельностью радикально меняют способы гнездования, кормовые методы, места кормления и т.д. и продолжают гнездиться в изменённых ландшафтах. Легко видеть, что это различие между «динамическими» и «консервативными» популяциями редких видов птиц на дальней периферии в зоне влияния города в точности повторяет различие между более и менее урбанизированными популяциями обычных видов птиц в самом городе (табл.2).
Традиционная стратегия охраны редких видов направлена исключительно на сохранение «консервативных» популяций вида (в крупных массивах по периферии региона). Появление же «динамичных» популяций вида, делающихся всё менее уязвимыми к преобразованию местообитаний вдоль горизонтального градиента, здесь просто не предполагается (до сих пор).
Однако сейчас оказывается, что к возвратной урбанизации после n лет отступления к периферии региона вдоль горизонтального градиента способны все редкие виды птиц, так же, как и обычные – только через разное время ожидания, и если не вымрут. Соответственно, появление «динамичных» популяций, активно осваивающих все более и более фрагментированные местообитания «вверх» по городскому градиенту рис.1, т.е. в благоприятной перспективе делающихся независимыми от нашей охраны, и утрачивающих уязвимость к урбанистической фрагментации ландшафта и сопряжённому с ней спектру антропогенных воздействий, оказывается регулярным событием.
С учётом того, что найти и вовремя заповедать массивы исходных ландшафтов, достаточные для поддержания жизнеспособности «консервативных» популяций этих же видов достаточно трудно (а то и вполне безуспешно из-за эффективного сопротивления природопользователей, см.пример Финляндии), поддержка урбанизации «динамичных» популяций столь же (а то и более) важна, чем сохранение «консервативных». Последнее наиболее значимо в периоды оттеснения вида с ростом урбанизированного «ядра» региона, но с началом возвратной урбанизации появившиеся «динамичные» популяции более перспективны поскольку растут, осваивают антропогенные аналоги природных местообитаний и могут перестать нуждаться в нашей охране.
Ещё пример – во второй половине ХХ века резко снизилась численность лесного голубя-клинтуха, жёстко связанного с широколиственными лесами, не показывавшего признаков урбанизации (в том числе потому, что гнездился лишь в дуплах старых деревьев). В последние годы, одновременно и в разных местах юга России и Украины вид «выходит» в безлесный ландшафт, где гнездится в опорах ЛЭП, бетонных столбах и других аналогичных конструкциях. Уже появились признаки роста численности: надо думать, доселе исчезающий вид теперь восстановит её, причём именно за счёт освоение антропогенно-изменённых территорий[20].


Колпицы на западе Украины загнездились на рыборазводных прудах, хотя сколько писали о них как о скрытных птицах диких дебрей. На сельских прудах Украины большие белые цапли и лебеди-шипуны стали вполне обычными птицами зарослей. В некоторых местах в таких зарослях на прудах умудряются гнездиться даже журавли.
Иными словами, к урбанизации способны все местные виды птиц, но с разной скоростью. Одни немедленно (прямая урбанизация, виды-выскочки). Другие с задержкой, и после долгого периода сокращения популяций под «давлением» урбанизированного «ядра», распространяющегося вдоль «городского градиента», после отступления на периферию региона и пр (возвратная урбанизация, бывшие урбофобы).
Различается и степень урбанизации, достигнутая конкретными видами в конкретном регионе – одни, раньше связанные лишь с крупными массивами старовозрастных дубрав, мелколиственных или таёжных лесов, начинают заселять лесные микрофрагменты, вторичные леса, перелески и старые парки. Другие доходят до рекреационных лесов/лугов/пойм ближних пригородов, осваивают природные территории города, а то и городскую застройку, и могут не только восстановить численность «до вытеснения», но и превысить её.
Иными словами, возвратная урбанизация постепенно захватывает те виды птиц, которые ещё 30-40 лет назад считались перманентным урбофобами. Так или иначе, сейчас окончательно ясно, что редкие и уязвимые виды птиц способны к возвратной урбанизации не меньше обычных и малочисленных. Естественно, это укрепляет модель «урбанизации местных особей» и входит в противоречие с «расселением городских рас».
——————————————————-
Далее, городские градиенты удивительно стабильны. Развитие большинства (если не всех) агломераций представляет собой ортогенез, почему легко предсказуемо на основе, скажем, известных правил «ранг-размер» Ф.Ауэрбаха, фигур размещения центральных мест Вальтера Кристаллера и легко моделируется с использованием системной динамики Джея Форрестера[21].
Раз установившись в начале урбанизации, скорость распространения этого процесса по территории региона постоянна в последующие 120-150 лет; все попытки людей ускорить или замедлить процесс сколько-нибудь значимы лишь короткое время, но несущественны на длинной дистанции, где преобладает автоматизм (также как с демографическим переходом в населении тех же территорий).
Таким образом, для сугубого большинства агломераций крутизна городского градиента и длина соответствующей «наклонной плоскости» (до пересечения с осью абсцисс) — величины постоянные или (что хуже для урбанизации «диких» видов) меняющиеся с постоянной скоростью (рис.1, 2 → 2’). Соответственно, обе эти характеристики урбанизации территории непосредственно связаны с урбанизацией «диких» видов птиц и (по нашей гипотезе) выступают одним из факторов, определяющих его возможность и сроки начала (относительно «точки 0» — начала региональной урбанизации, роста «ядер» агломераций и соответствующего преобразования местообитаний вида в регионе). Именно, они влияют на то, как именно среагирует популяция конкретного вида птиц на прогрессирующее развитие «урбанизированного ядра» региона с индуцируемым им развитием городов спутников или центров 2-го, 3-го и т.д. порядка на периферии агломерации.
Для вида всё вышеназванное «отображается» в преобразовании «материка» исходных местообитаний в «архипелаг». «Острова» последнего чем дальше, тем больше подвержены фрагментации извне, инсуляризции изнутри, преобразованию с краёв, уменьшению и «дроблению» со стохастизацией размещения по территории (рис.1). Сопоставляя их с жизненной стратегией местной популяции до начала урбанизации и вообще хоть какого-то реагирования на данный процесс, можно предсказать характер последней. Жизненная стратегия вида оценивается по степени консерватизма/лабильности популяционной структуры – альтернативных способов организации популяционных систем у птиц и/или их способов реагирования на «вызовы», связанные с естественной (обычно цикличной) или антропогенной (направленной) трансформацией местообитаний.
Оппозиции консерватизм-лабильность видов (или жизненных стратегий особей) определяли по В.М. Галушину и А.В. Кузнецову (1991, Кузнецов, 1993[22]). Это противоположные типы приспособительных реакций популяционной системы (реализованной в определённом ареале) на разнообразные “возмущения”, будь то трансформация местообитаний, обеднение ресурсной базы или периодическое падение доступности ресурсов.
Консервативные виды отличаются стабильной численностью низким потенциалом размножения, способностью существовать на минимуме ресурсов, высокими требованиями к устойчивости ресурсной базы, минимальным динамизмом населения при максимальном разнообразии биотопов (и ресурсов), эксплуатируемых на занятой территории. Лабильные — представляют собой их противоположность. У них высок потенциал размножения, обычны резкие колебания численности, территориальные и биотопические связи весьма изменчивы: население вида может быстро (без запоздания) концентрироваться в местах появления массового корма, будь то размножение грызунов, вспышки численности насекомых фито- или ксилофагов. Лабильные виды предпочитают менее структурированные и более нарушенные местообитания, с флюктуирующими ресурсами, с парцеллярной структурой растительности, сильно варьирующей во времени и в пространстве (via)
——————————————
Как можно понять, будет ли вид в конкретном регионе урбанизироваться сразу, как только его местообитания начнут «захватываться» и «расчленяться» в связи с ростом центрального ядра? Или начнёт отступать к периферии, «уходить» с мелких «островов» на наиболее крупные, а когда, в свою очередь, урбанизационный процесс начнет дробить/уменьшать их – полностью исчезать из зоны влияния города на «материк» местообитаний по периферии региона? Если второе – когда это отступление прекратится, через сколько лет ожидания начнётся возвратная урбанизация, когда вид «переключится» на освоение «островов» антропогенно трансформированных вариантов исходных местообитаний или их техногенных вместо прежней привязанности к «островам» последних с отступлением вместе с ними? Наконец, есть ли виды, настолько урбофобные и/или настолько консервативные территориально/биотопически, что напрочь неспособны к возвратной урбанизации, даже на самом дальнем пределе распространения трансформирующего влияния города (у границ его зоны влияния в регионе), которые лишь «отступают» по городскому градиенту, но не «взбираются вверх»?
Иными словами, возможна ли урбанизация данного вида в данном регионе; если возможна, она будет прямой или возвратной, если второе – то каков период задержки? Происходящее означает начало «движения» местной популяции по городскому градиенту «вверх» вместо прежнего «скатывания вниз», т.е. смену знака процесса популяционной динамики, почему и говорят о переключении (рис.2).
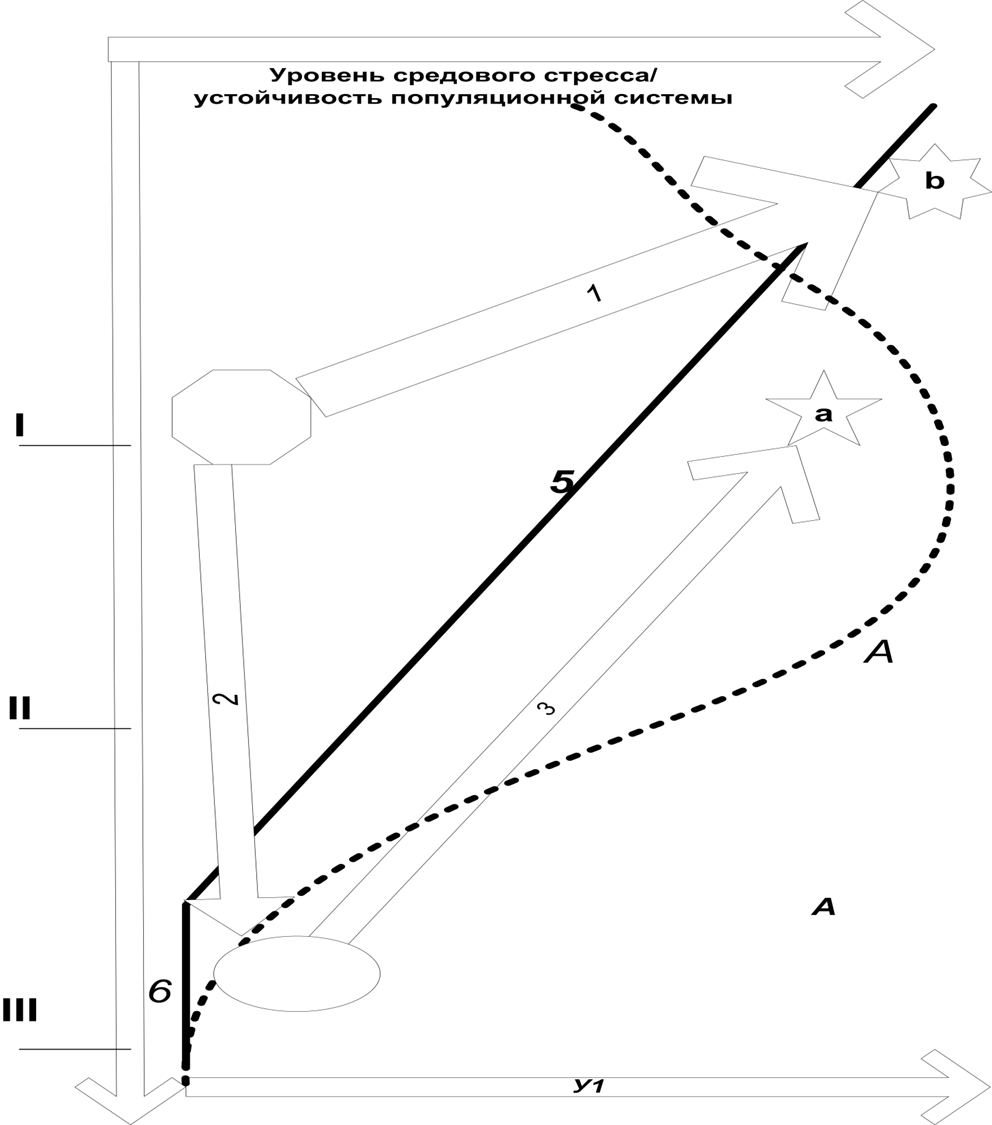
Рисунок 2. Динамика региональных популяций видов в ходе прямой и возвратной урбанизации. Обозначения. Ось Х – городской градиент, вдоль которого а) распространяются преобразования исходных местообитаний «урбанизированным ядром», б) понижается уровень средового стресса для популяции соответствующих видов, созданного процессами 2-4 рис.1, в) «движется» ареал вида в регионе по ходу «оттеснения» на периферию (2) и последующего «возвращения» во фрагментированные и преобразованные местообитания (3, возвратная урбанизация). Оси У-У1 – уровень средового стресса на соответствующем участке «городского градиента» и устойчивость пространственно-этологической структуры популяции, существующей на данном участке градиента, чем выше – тем устойчивей. Стрелки – направления популяционной динамики, заданные изменением устойчивости воспроизводства её пространственно-экологической структуры в локальностях, соответствующих разным участкам градиента. 1 – прямая урбанизация лабильных видов, с быстрым заселением антропогенного «архипелага» местообитаний (звезда b – завершение этого процесса, урбанизированные популяции), 2 — отступление в область минимума средового стресса, на периферию региона, 3 – направленное заселение областей со всё большим и большим уровнем средового стресса по ходу возвратной урбанизации (звезда а – результат этого процесса, восстановленная популяция ранее уязвимого вида, иногда и урбанизированная), овал – более или менее длительное пребывание в оттеснённом состоянии на периферии региона.
Иными словами, при прямой урбанизации средовой стресс некоторый «пусковой» уровень средового стресса (с какими параметрами развития агломерации он может быть связан, см.ниже) немедленно вызывает процесс 2 (рис.2). При возвратной – этот же уровень средового стресса вызывает сперва отступление по городскому градиенту «вниз», пребывание в оттесненном состоянии (1, 3, рис.2), и лишь потом начинается урбанизация. Если, конечно, вид в данном регионе не исчезнет вовсе за время пребывания со сниженной численностью/сокращённым ареалом, распавшимся на отдельные местонахождения. Что хорошо иллюстрируется сравнением близких видов птиц, пытающихся сохраниться на территории города после уничтожения их местообитаний при постоянно идущем преобразовании урболандшафта за счёт гнездования в несвойственных биотопах, нетипичными способами и пр., и немедленно оставляющих данную территорию. См.таблицу 2 отсюда.
Таблица 2. Противоположное реагирование близких видов на локальное уничтожение гнездовых местообитаний (на примере авифауны г.Москвы)
| Вид, остающийся на гнездовании в городе, находящий замену утраченным местообитаниям | Вид, прекращающий гнездование в городе даже при наличии замены утраченным местообитаниям |
| Aythia fuligula | Aythia ferina |
| Podiceps cristatus | P.nigricollis |
| Sterna hirundo, Larus canus | L.ridibundus |
| Gallinula chlorophus | Fulica atra |
| Turdus pilaris | T.iliacus |
| T.merula | T.philomelos |
| Anas platyrhynchos | A.querquedula, A.clypeata |
| Charadrius dubius | Vanellus vanellus |
| Cyanosylvia svecica | Acanthis cannabina |
| Ficedula hypoleuca | F.parva |
| Motacilla citreola | M.flava |
| Dendrocopos minor | D.leucotos |
| Oenanthe oenanthe | Saxicola rubetra |
| Locustella fluviatilis | Locustella naevia |
| Erithacus rubecula | Troglodytes troglodytes |
| Sylvia atricapilla | Sylvia borin |
Примечание. Таблица построена по данным табл.2 книги. Её перечень, безусловно, далек от завершения, а отдельные примеры требуют уточнения. Но факт существования в городской авифауне «пар» экологически близких видов, противоположно реагирующих на антропогенную динамику местообитаний, не вызывает сомнений.
Важно подчеркнуть, что в городе и ближних пригородах виды левого и правого столбцов табл.2 экологически идентичны, поскольку антропогенное нарушение местообитаний, в том числе их урбанистическая трансформация увеличивает перекрывание экологических ниш близких видов, а не уменьшает его как можно ожидать из-за прогнозируемого обострения конкуренции. Впрочем, в силу известной парадоксальности урбоэкосистем (которая создаёт «плюсы» урбанизации для «диких» видов) последнего тоже не получается. Именно в силу различия жизненной стратегии (все «уходящие» виды правого столбца табл.2 консервативней «остающихся» в левом) в момент увеличения средового стресса в данной локальности они реагируют прямо противоположным образом, соответствуя процессам 2 и 1 рис.2
Как произвести тот прогноз, о котором шла речь выше? В его основе – простая идея, что главный минус урбанизации региона для «диких» видов птиц связан отнюдь не с уничтожением местообитаний. Вместо уничтоженных создаются новые, и виды легко могли бы в них «пересесть». Пытаются же они все в периоды роста численности заселять города? а тут зачастую речь идёт об освоении всего лишь «обломков» прежних биотопов, возникающих в процессе антропогенной фрагментации, и дело происходит в «привычном для вида» внегородском ландшафте или рядом с ним. Настоящей проблемой, ведущей к невозможности сохранения видовых популяций в прежних местах обитания, после того, как до них «доберутся» (по мере урбанизации) процессы 2-4 рис.1, оказывается дестабилизация популяционных систем «островным эффектом» и его производными. И чем консервативней жизненная стратегия вида, тем проблема острее, и выше риск вымирания «под прессом» растущего «урбанизированного ядра»
Чем выше уровень «городского градиента», тем больше естественные популяции вида, со свободно реализующейся пространственно-этологической структурой, преобразованы в метапопуляции. Там эта реализация несвободна, а подток особей «подвижного резерва», пополняющих локальные группировки (и влияющий на их долговременное сохранение сильно больше местного размножения и локальных адаптаций), из гарантированного события делается случайным. Соответственно, во всех местных поселениях вида на «архипелаге» вероятность своевременного «заполнения вакансий» владельцев территорий или брачных партнёров чем дальше, тем больше падает, в пределе стремясь к 0; данный процесс идёт тем быстрей, чем сильней «островной эффект», что для данного случая значит — чем выше уровень средового стресса, распространяющегося вдоль городского градиента рис.1-2.
Поэтому без смены жизненной стратегии метапопуляции «консервативных» видов во фрагментированных местообитаниях менее устойчивы и жизнеспособны, чем естественные популяции того же вида в непрерывных, и тем менее, чем сильней фрагментация, островной эффект с краевым. А вот после смены жизненной стратегии, по ходу возвратной урбанизации они перестают быть метапопуляциями и, хотя вновь заселяют «архипелаг» фрагментированных местообитаний в пригородах (а то и в самом городе), утрачивают уязвимость к «островному эффекту». Также, как и (в существенно большей степени) его утратили урбанизированные популяции всех освоивших город видов – чёрного дрозда, сойки, сороки, рябинника и пр.; там и тогда, где это произошло.
Соответственно, урбанизация территории с её характерными фигурами пространственной экспансии урболандшафта в регионе представляет собой существенный стресс для популяционной жизни практически всех видов. Если они сохраняют верность прежним местообитаниям, прежним партнёрам и территориальным связям, то повышенная нестабильность и направленная эволюция городского ландшафта гарантированно «загонит» их гнездовые поселения в «экологическую ловушку», и тем раньше, чем выше консерватизм и уровень городского градиента.
Во внегнездовой период те же факторы препятствуют сохранению связности между местными группировками, нарушая устойчивость потоков переселенцев и нерезидентов (которые управляются и регулируются как направляющими линиями экологического характера, так и сигнальным полем существующих поселений, а то и другое взаимосвязано ослабляется при фрагментации). Из-за этого утрачивается способность к саморегуляции внутри системы, к адекватной и динамичной «переброске» части населения из деградирующих местообитаний во вновь образующиеся перспективные. В урбанизированных же регионах вид выживает в первую очередь за счёт этого динамизма, умения «следовать за средой» при постоянных изменениях последней. Система «рассыпается» на изолированные поселения, «запертые» на изолированных «островах» местообитаний, и выживание каждого из них зависит уже от сугубой случайности, причём вероятность невымирания всё больше и больше снижается от островного эффекта.
Высокий «динамизм» позволяет популяции быстро изменять вид зависимости плотности населения от площади местообитаний с положительной на отрицательную и обратно. При высоком «динамизме» эти изменения связи вида с собственным местообитанием (или, что то же самое, изменения экологической ёмкости местообитания «с точки зрения вида») происходят в ответ на фрагментацию видовых биотопов или нестабильность условий среды. Локальные изменения инициируют быстрые и резкие перераспределения особей по ареалу на значительных территориях внутри ареала, так что «кружево ареала» значительно изменяется (Connor et al., 2000).
«[С.С.Москвитин (2001) показал], что успех внедрения новых видов в район г.Томска (19 видов за ХХ в., в том числе закрепление в южнотаёжной зоне «южных» и «лесостепных» видов) вызван разными причинами или их сочетаниями, но необходимым условием всегда является высокий динамизм внедренцев. Его же демонстрируют виды-потенциальные урбанисты, когда легко и успешно осваивают городские экологические ниши в разных городах внутри видового ареала. Следовательно, «динамизм» популяционной системы вида причинно связан с присутствием того же типа организации популяционной системы, который задаёт устойчивость городских популяций («городской тип», табл.1). Успешная урбанизация «диких» видов есть крайнее (наиболее специализированное) проявление этого «динамизма».
Анализ расселения видов дендрофильной авифауны к югу по пойменным лесам таких рек, как Урал, Дон, Северской Донец, по лесополосам, посаженным в рамках Сталинского плана преобразования природы, вполне подтверждает эту гипотезу. Виды, наиболее склонные к урбанизации, показывают и наибольший динамизм в освоении новосозданных «островных ландшафтов», в расселении по ним к югу, в нелесные пространства степи и даже полупустыни. Это большая синица, зяблик (Fringilla coelebs), лазоревка (Parus caeruleus), мухоловка-пеструшка, чёрный дрозд, зарянка, большой и малый пёстрые дятлы, ушастая сова (Asio otus), вяхирь и пр. – всё потенциальные урбанисты (Белик, 1997).
Их скорость продвижения к югу по лесным «островам» и «полосам» в пойме р.Урал достигает 2-6 км/год км в год. Минимальная скорость расселения зафиксирована у самых крупных из названных видов, вяхиря и большого пёстрого дятла (2-4 км/год). Во второй половине ХХ в. резко возросла скорость расселения всех видов, лидирующих по темпам урбанизации: у большой синицы с 2 до 14 км/год, у лазоревки и зяблика с 2-3 до 5-6 км/год. С той же скоростью неморальные и бореальные виды птиц расселялись в ХХ веке в низовьях Северского Донца (Белик, 1997). Большинство городских популяций этих видов в урбанизированных регионах Европейской части СССР возникали в годы «повышенного динамизма»=наибольшей скорости расселения дендрофильных видов в нелесные зоны.
Показательно, что скорость распространения урбанизационных процессов у таких видов, как чёрный дрозд, вяхирь, рябинник или тетеревятник на 1-2 порядка превышает самые высокие темпы расселения, зафиксированные при расширении ареала вида в природных ландшафтах (то же верно для скоростей расселения по лесополосам и лесным микрофрагментам в нелесном ландшафте). Например, скорости, зарегистрированные при расселении овсянки-дубровника (Emberiza aureola), чечевицы (Carpodacus erythrinus), зелёной пеночки (Phylloscopus trochyloides), синехвостки (Tarsiger cyanurus) в ХХ веке. Аналогичные расширения ареалов здесь требуют десятилетий (Зимин, 2001).
В целом, виды-потенциальные урбанисты за годы по антропогенно-фрагментированным местообитаниям расселяются на расстояние, которое другие виды в непрерывных местообитаниях «проходят» за десятилетия. Поэтому они были названы В.В.Леоновичем (1991) «виды-выскочки». Это значит, что потенциальным урбанистам исходно присущ иной тип организации популяционной системы, наиболее приближающийся к «городским» вариантам организации. Благодаря ему именно в «островном ландшафте», созданном и поддерживаемом человеческой деятельностью (неважно, в городе или за его пределами), все эти виды обретают высокую устойчивость популяций и особый динамизм. Им исходно присущ «городской» тип популяционной организации или близкий к нему (табл.1), тогда как прочие виды региональной авифауны лабилизируют популяционную структуру – и жизненные стратегии индивидов – лишь попав «под давление» урболандшафта». Via
Следовательно, урбанизация, распространяющаяся вдоль «городского градиента», «с точки зрения» реагирующих видовых популяций, представляет собой некий вызов, ответ на который подчиняется своего рода правилу Ле Шателье. Вид или приспосабливается (немедленно либо с запозданием в случае возвратной урбанизации), меняя «конструкцию» популяционной системы в сторону большей «открытости» и «проточности» группировок, большей лабильности структуры системы, с соответствующей лабилизацией жизненной стратегии.
Важно подчеркнуть – здесь отчётливым образом видно, что популяционная система вида реагирует как единое целое[23]. Приспособительное изменение здесь происходит одномоментно и представляет собой направленную перестройку пространственно-этологической структуры популяций всего региона (или как минимум рассоложенных в «точке старта» урбанизации, возвратной или прямой). Больше того, этот процесс достаточно независим от наличия/отсутствия у конкретных особей специальных полезных приспособлений к конкретным факторам урбосреды, к тому городу, к воздействию которого вид приспосабливается. Когда сдвиг в «конструкции» популяционной системы от «внегородского типа» к «городскому» (табл.1) произошёл, популяция может наполняться особями совсем из других регионов, отнюдь не испытывавших воздействия конкретного города, и тем не менее успешно осваивать созданный им «островной» (а потом и урбо-) ландшафт.
Особенно это верно в случае возвратной урбанизации, когда оттеснённый на периферию вид вдруг быстро восстанавливает численность, осваивает фрагментированные местообитания и пр. Поскольку оставшиеся периферийные популяции были невелики (иногда и вполне ничтожны – несколько пар), быстрый рост численности, расширение ареала обеспечиваются включением особей «подвижного резерва» с других территорий. Причём тех, на которых ещё возвратная урбанизация не произошла, вид придерживается традиционных местообитаний, обычного образа жизни, иначе бы эти особи поселялись бы в новых, изменённых местообитаниях, а не включались в периферийные популяции. Однако эти особи, включившись в уже изменённую пространственно-этологическую структуру видовой популяции в данном регионе, активно осваивают фрагментированные местообитания, «движутся вверх» по градиенту рис.2 вплоть до заселения собственно городской территории, как ворон, тетеревятник и те виды, что обсуждаются выше.
Иными словами, при возвратной урбанизации направленная перестройка популяционной системы под «стрессирующим воздействием» урболандшафта здесь существеннее наличия/отсутствия специфических адаптаций особей (тем более что скорость процесса такова, что последние заведомо не успеют распространиться отбором). Во всех случаях урбанизации, как немедленной, так и возвратной, соответственные изменения на уровне особей регистрируются (а тем более увеличиваются в числе либо степени проявления) лишь после перестройки структуры системы и начала «движения» вверх по городскому градиенту у «отступившей» популяции.
Что очевидным образом противоречит традиционной (восходящей к «школьному» варианту СТЭ) модели появления урбанизированных популяций птиц. Там приспособления возникают на уровне особей и распространяются отбором, оба процесса идут тем быстрее и тем более выражены, чем сильней «давит» урбанизация территории на население вида именно в данной локальности. При всей идеологической «правильности» и привычности такой картины, для неё, по-видимому, вовсе нет места – пространственный и временной паттерны урбанизации, что немедленной, что возвратной, не только не согласуются с ней, но прямо противоположны.
Сперва под давлением процесса урбанизации территории, «сила» которого задана «местом» на городском градиенте рис.1-2, меняются характерные скрепы, сцепляющие «эгоистических индивидов» в системное единство – популяцию. и не как-нибудь, а видоспецифическим образом, с характерной структурой отношений социальных, пространственных (к территории) и биотопических. Данное изменение может произойти сразу (прямая урбанизация) или через годы и годы пребывания популяции «под прессом» урбанизации региона (возвратная), за что вид расплачивается долговременным сокращением численности и ареала, но рано или поздно возвратная урбанизация идёт у всех видов.
Повторюсь: даже тех, которых считали исключительно уязвимыми и угрожаемыми в староосвоенных регионах Европы-Америки-Японии их территориального и биотопического консерватизма, «привязывающего» популяцию к крупным массивам старовозрастных лесов, верховых болот и иных исходных ландшафтов. Последние же практически обречены: их можно сохранить как ландшафтный или ботанический объект, но площадь останется недостаточной для поддержания связанных с ними – и поэтому угрожаемых – видов птиц.
Лишь потом изменившиеся отношения внутри популяции меняют поведение и т.д. биологические характеристики индивидов, но не ранее. Популяция приспосабливается к изменениям местообитаний в связи с урбанизацией и осваивает «островной ландшафт» (включая «городское ядро» в его центре) как единое целое, не отдельными особями, которые бы ещё до начала «давления» были бы «наиболее городскими» по своему поведению, экологическим предпочтениям и т.д. деталям биологии.
Соответственно, преадаптированность к урбанизации или, наоборот, урбофобия, проявляются на уровне межвидовых, а не межиндивидуальных различий, причём именно в «конструкции» популяционных систем (большая-меньшая лабильность/консерватизм организации, больший-меньший динамизм в динамике численности), а не в natural history объединяемых ими индивидов.
Легко видеть, что одни и те же виды в разных регионах очень по-разному реагируют на преобразования «их» местообитаний вдоль городского градиента. В одном случае это стимулирует урбанизацию вида (немедленную или возвратную, в зависимости от «крутизны» градиента, см.ниже), в другом – его «отступление» на периферию с неизменной привязанностью к прежним местообитаниям (архипелаг которых последовательно подрывается урбанизацией, почему и приходится «отступать»).
В самом деле, у самых разных видов в настоящий момент происходит заселение разных городских центров, разные популяции находятся на разных стадиях процесса и разделены неурбанизированными популяциями. Например, крупная, с мощным клювом большеклювая ворона полностью урбанизировалась на Хоккайдо, освоив Саппоро и др. крупные города. В Приморье же её урбанизация лишь начинается в крупнейшем городском центре – Владивостоке (сравнимым с Саппоро). В других частях ареала (Забайкалье, север Приморья и пр.) она обитает только в природных местообитаниях, в отличие от мелкой и слабоклювой чёрной вороны[24]. Или же вспомним лёгкую урбанизацию всех видов из сноски 4 в Европе, и медленную, а то и отсутствующую урбанизацию их в Московской области и других староосвоенных регионах Европейской части СССР, с не менее развитыми городскими центрами.

Большеклювая ворона, урбанизировавшаяся на Хоккайдо
Собственно, исходный вопрос ishs был именно об этом. От чего это зависит? Почему по мере развития урбанизации региона разные виды, сперва демонстрирующие вторую реакцию, таки переходят к урбанизации (соответственно, возвратной) если только соответствующая региональная популяция не вымрет «во время отступления»?
Очевидно, чтобы стимулировать контринтуитивный ответ популяционной системы — движение «вверх» по городскому градиенту рис.1-2 вместо более естественного и ожидаемого «цепляния» за видоспецифичные биотопы, сам градиент должен быть некой оптимальной крутизны и длины.
Если меньше – не будет необходимости приспособления, темпы преобразования традиционных биотопов будут недостаточными, чтобы вызвать средовой стресс и вызвать перестройку популяционной структуры, после которой урбанизация возможна. Если больше – средовой стресс будет слишком силён, а его усиление на радиальном срезе от урбанизированного ядра к периферии начнёт возрастать слишком быстро, чтобы искомая перестройка популяционной системы вида стала возможной.
Иными словами, уровень средового стресса (наглядно видимый в динамических характеристиках преобразования «материка» региональных местообитаний вида в «архипелаг» или изменения уже существующих «островов»), чтобы вызвать приспособительные изменения популяционной системы с последующей возвратной урбанизацией, должен распространяться во времени и в пространстве с некой оптимальной скоростью, разной для разных видов. Если она слишком велика, то вызовет отступление на сохранившийся «материк» вместо повышения устойчивости популяционной системы (процесс 2 рис.2), если мала – вид «не заметит» воздействия.
Иными словами, урбанизация вызывается в каждом регионе в отдельности, и не столько развитостью «центрального ядра», сколько плавностью переходов от него к низовой урбанизации (через города 2-го, 3-го ит.д. уровня в соответствующей агломерационной системе, не случайно так напоминающей планетную).
Чем более консервативной была жизненная стратегия вида первоначально, тем большая плавность переходов потребуется для вызывания урбанизации. В этом случае городской градиент должен быть менее крутым, но более длинным. Если он будет круче – вид начнёт отступать на периферию региона, придерживаясь в процессе отступления исходных биотопов до тех пор, пока городской градиент не придёт в соответствие с параметрами, оптимальными для урбанизации именно данного вида. Тогда запускается возвратная урбанизация (после перестройки популяционной структуры).
Поэтому первые урбанизированные популяции чёрного дрозда, вяхиря и др. видов появились именно в тех районах Западной Европы, где были развиты не только (и не столько) крупные центры, сколько низовая урбанизация, где уже к началу ХХ века, а то и раньше реализовались кристаллеровские фигуры. Сказанное особенно верно для тех видов птиц, которые у нас «больше лесные», а в Европе «больше кустарниковые», вроде лесной завирушки с зарянкой.
Следовательно, гипотеза расселения городских рас кажется крайне сомнительной. «За» неё лишь высокие темпы роста численности городских популяций в период образования, при заселении территории данного конкретного города (практически по экспоненте, с последующим выходом на плато и/или депрессией). Однако уже расселение между городами отнюдь не демонстрирует такой же высокой скорости (какой можно было бы ожидать на основании данных о заселении города в процессе урбанизации, но и по хорошо согласующимся с ними данных о расселении «видов-выскочек» по рукотворным ландшафтам вроде лесополос, превышающим максимальные темпы расселения видов в природных ландшафтах). Нет, здесь процесс, как уже говорилось, как бы натыкается на «невидимые преграды», и не может преодолеть их десятилетиями – а затем вдруг скачком захватывает значительную территорию рядом. Иного объяснения, как урбанизации местных особей, происходящей независимо в каждой из агломераций, и управляемых её собственной динамикой развития (рис.1-2), я тут не вижу.
Наконец, окончательный аргумент в пользу модели «урбанизации местных особей» и против «расселения городских рас». В подавляющем большинстве случаев, когда урбанизационный процесс могли проследить с самого начала, от поселения первых пар, он начинался с крупных лесов на окраине города. Затем вид заселял лесопарки внутри города, «двигаясь» от крупных «островов» по всё более мелким и центрально расположенным. Затем, когда (и если) урбанизация вида дошла до конца, он переходил к гнездованию в городской застройке, элементах озеленения вокруг домов и пр.
Так, именно в этом направлении происходит урбанизация пустельги и перепелятника в г.Львове. Оба вида отсутствовали на гнездовании в 1960-е гг., освоение урбосреды начали с крупных «островов» городских лесов и открытых ландшафтов. С 1990-х гг. численность обоих видов растёт, птицы освоили меньшие по площади и более изолированные «острова» соответствующих ландшафтов, но не застройку. Пустельги до сих пор не охотятся среди застройки, хотя это вполне возможно[25]. Другой пример – быстрое расселение канареечного вьюрка по северо-западу СССР в 1960-70-е годы.
Характерно, что, скажем, вяхирь таким путём сперва заселил «урбанизированное ядро» региона, г.Калининград, а затем уже, из этого города, стал осваивать средние и малые города региона[26]. Тот же путь освоения агломерации – из природных ландшафтов в «ядро» вверх по городскому градиенту, а потом уже в центры 2-го, 3-го, 4-го и т.д. уровней – показывают и прочие виды птиц, подтверждая модели рис.2.
В случае расселения городских рас, напротив, наблюдали бы появление первых поселений вида немедленно в центре города, наиболее изменённых местообитаниях. Такое отмечено лишь при расселении уже сформированных городских популяций видов (скажем, горихвостки-чернушки или кольчатой горлицы) в новые области, где вид ранее не обитал никогда (тем более в природных местообитаниях). В случае расселения городских рас скорость занятия ими новых урболандшафтов определяется только уровнем численности и расселительными возможностями первой возникшей городской популяции.
Поскольку при их образовании численность соответствующих видов растёт экспоненциально, способность городских особей к переселению между группировками повышена (как и для нерезидентных птиц из «резерва» возникшей городской расы), то они давно должны бы занять все подходящие города. Примерно как это сделала кольчатая горлица или сирийский дятел в Центральной и Восточной Европе, раз проникнув в соответствующий ландшафт. Обычно такого не наблюдается; урбанизация подавляющего большинства видов, причём именно что хороших урбанистов, как бы спотыкается о невидимые границы, которые не может преодолеть десятилетиями.
Это склоняет нас к тому, что большинство городских популяций большинства видов появляется по модели местной урбанизации наличие, отсутствие, темпы и сроки которой всецело определяются, с одной стороны, жизненной стратегией вида, оцениваемой по степени консерватизма-лабильности. С другой – она представляет собой «ответ» на «вызов», следующий из определённой скорости преобразования «материка» местообитаний вида в «архипелаг» растущим урбанизированным «ядром» региона, его последовательного «дробления», изоляции «островов» и пр. То есть на определённый уровень средового стресса, созданный «давлением» развивающегося урболандшафта на местообитания вида не только внутри, но и вне города.
Поэтому, как таковая, местная урбанизация диких видов птиц не может не зависеть от местных же особенностей урбанизационного процесса, соотношения темпов роста «ядра» агломерации, подчинённых ей городов и низовой урбанизации. Скорее всего, местные особенности урбанизации определяют возможность процесса, а жизненная стратегия вида в сочетании с первыми – сроки начала процесса (относительно «точки 0» — начала урбанизации региона) и его скорость.
Ergo: сумма всех относящихся к делу фактов поддерживает модель местной урбанизации, но не «расселения городских рас», хотя в отдельных случаях последнее тоже бывает. Поэтому даже удивительна высокая популярность первой, но не второй гипотезы на Западе. В частности, так пытаются объяснить урбанизацию чёрного дрозда, хотя исключительная оседлость городских птиц при быстром движении границы урбанизации с запада на восток никак не вяжется с данным объяснением. Хорошо документированных случаев «расселения городских рас» всего три, два из них связаны с завозом человеком (черных дроздов из Познани в Киев, домовых воробьёв в г.Елизово на Камчатке с материка, где ранее обитал только полевой воробей).
Примечания
[1] Кавычки связаны с тем, что в подавляющем большинстве случаев появление новых урбанизированных популяций вызвано урбанизацией местных особей из естественных биотопов, а не заселением новых городов уже сформировавшимися «городскими» птицами. См. обсуждение ниже.
[2] Лыков Е.Л., 2009. Фауна, население и экология гнездящихся птиц городов Центральной Европы : на примере Калининграда : диссертация … кандидата биологических наук. 286 c.
[3] К ним относятся регионы, центры которых оказываются «ядрами» агломераций, а территория более-менее полностью «захватывается» процессом агломерирования за время городского и индустриального развития последних 100 лет. Соответственно, малонарушенные природные территории в таких регионах быстро оттесняются к периферии и сохраняются там лишь отдельными фрагментами в «зонах экономического вакуума» на стыках разных регионов. Однако по мере развития центрального «ядра» его зона влияния «накрывает» всю область, и данные территории подвергаются всё большему риску фрагментации & трансформации местообитаний. Наравне с прочими, уже изменёнными ландшафтами.
[4] Если отношения в популяционной системе вида не изменены в направлении табл.1, вид всё равно формирует городские поселения, особенно в периоды роста численности, и обычно успешно размножается там. Однако большая часть приплод из успешных городских поселений утекает на послегнездовые миграции за город, а это у птиц время запечатления будущих гнездовых биотопов. Соответственно, урбанизация таких видов тормозится даже при успешном размножении тех единичных поселений, что образуются в городе. Последние существуют много лет (и даже десятков лет): однако вопреки эффективному размножению остаются ограничены численно, почти не предпринимают попыток колонизации ближайших подходящих местообитаний и пр.
[5] И других европейских видов, показывающих сильный западно-восточный градиент интенсивности урбанизационных процессов: вяхиря, крапивника, лесной завирушки, черного дрозда, певчего дрозда, дрозда-дерябы, черноголовой славки, пеночки-теньковки, пеночки-веснички, желтоголового королька, красноголового королька, сойки, хохлатой синицы, короткопалой пищухи, зяблика, снегиря. Выявлен в исследовании К. Витт с соавторами (Witt et al., 2005) сравнивших орнитофауну трех больших городов — Гамбурга, Берлина и Варшавы.
[6] Видимо, именно так исчезли природные популяции у домового воробья Passer domesticus и домовой вороны Corvus splendens – единственных видов птиц, у которых вся масса особей представлена городской популяцией, а негородские отсутствуют.
[7] В процессе урбанизации-индустриализации последних 100 лет оно растёт и распространяется сообразно эволюционным моделям урбанизации.
[8] Тогда, когда они ещё были крупными, что увеличивает риск выпадения видов, исходно присутствовавших на «материке», под воздействием «островного эффекта»
[9] «Экология городской фауны». М.: Мир, 1990, стр.151-154. 1-2-3-4.
[10] Понятие матрикса и его проницаемости для особей в метапопуляционной экологии см.тут
[11] Экологические сети и сохранение биоразнообразия Центральной России: исследование на примере торфяных болот Петушинского района / Очагов Д.М., Райнен Р., Бутовский Р.О., Алещенко Г.М., Ерёмкин Г.С., Есенова И.М. – М: ВНИИприроды, 2000. – 80 с.; Бутовский Р.О., Райнен Р., Очагов Д.М., Алещенко Г.М., Мелик-Багдасаров Е.М., Ерёмкин Г.С., Есенова И.М. Сохранение природы торфяных болот центральной и северной Мещеры. – М.: ВНИИЦлесресурс, 2001. – 120 с.
[12] В.Н.Мельников, 2008. Сокола (Falco) в Восточном Верхневолжье//Новiтнi дослiдження соколоподiбних та сов. Матерiали III Мiжнародної наукової конференцiї «Хижi птахи України». Кривий Рiг, 2008. С.249-254.
[13] например, исключительно сильного в Финляндии (1-2), послабее в ФРГ (благодаря мощному и антикапиталистически настроенному «зелёному» движению), сильного в СССР 1955-65-х гг. (хрущёвская «химизация», «борьба с вредными хищниками») и слабого в период после 1967-1972 гг.
[14] бывшие урбофобы по классификации К.Н.Благосклонова (Гнездование и привлечение птиц в сады и парки. М.: изд-во МГУ, 1991). Он делил виды региональной орнитофауны урбофобов, урбофилов и синантропов и полагал виды первой категории неспособными даже к возвратной урбанизации, до конца «цепляющимися» за последние «острова» своих исходных местообитаний, и исчезающими, когда те сократятся настолько, что не смогут поддерживать жизнеспособную популяцию вида. «Бывшие» же они потому, что сейчас всё более проясняется способность их популяций через L лет пребывания «под давлением» урбанизированного «ядра» региона вдруг переключаться на использование фрагментированных местообитаний, которые прежде избегались, заселять сперва местообитания ближних пригородов, а потом и урбанизированные территории, откуда они были вытеснены ранее.
[15] А.С.Мальчевский. Птицы перед микрофоном и фотоаппаратом. Изд.2е. Л.: изд-во ЛГУ, 1976. С.74.
[16] К.А.Письменный, 2008. Гнездование змееяда (Circaetos gallicus (Gm.)) в Киевской области и проблемы его адаптации к хозяйственной деятельности человека // Новiтнi дослiдження соколоподiбних та сов. Матерiали III Мiжнародної наукової конференцiї «Хижi птахи України». Кривий Рiг, 2008. С.307-311.
[17] Динамизм (популяции) — термин С.С.Москвитина (2001). Под ним понимается способность птиц (большая у одних и меньшая у других видов) «дистанционно оценивать ландшафтную картину больших пространств, в совершенстве ориентироваться, своевременно и направленно реагировать на конкретную климатическую, экологическую и антропогенную ситуацию и высокоэффективно пользоваться ими для себя» (Москвитин, 2001). Также см.ниже.
[18] Ветров В.В., Милобог Ю.В., 2008. Распространение могильника (Aquila heliaca Sav.) в степной зоне Украины // Новiтнi дослiдження соколоподiбних та сов. Матерiали III Мiжнародної наукової конференцiї «Хижi птахи України». Кривий Рiг, 2008. С.51-54.
[19] Грищенко В. Н., Дзюбенко Н. В., Бокотей А. А., Яблоновская-Грищенко Е. Д., 2010. Гнездование колпицы на западе Украины // Орнитология в Северной Евразии : материалы 13 Междунар. орнитол. конф. Северной Евразии (Оренбург, 30 апр. — 6 мая 2010 г.): тез. докл. Оренбург. С. 109.
[20] Белик В.П., Ветров В.В., Милобог Ю.В. Ренессанс клинтуха в Восточной Европе: демографический потенциал новой адаптации // Стрепет. 2010. Т. 8. Вып. 1. С. 70-74.; Гаврилюк М.Н. (2009): Гнездование клинтуха в бетонных столбах в Черкасской области. — Беркут. 18 (1-2): 209-210.; Бобенко О.А., Ильюх М.П., Плеснявых А.С., Друп А.И., Друп В.Д., Хохлов А.Н. (2007): Клинтух – новый гнездящийся вид Ставропольского края. — Птицы Кавказа: изучение, охрана и рац. использование. Мат-лы научно-практич. конф., посвящ. 25-летней деятельности северокавказской орнитол. группы. Ставрополь: СГУ. 6-16.
[21] См. его книгу «Динамика развития города» (М.: Прогресс, 1974); к слову, он был руководителем в MIT аспирантов Денниса и Донеллы Медоуз, когда они создавали первые модели пределов роста.
[22] Галушин В.М., Кузнецов А.В., 1991. Концепции жизненных стратегий птиц// Мат-лы Х Всес. орнитол. Конф. Ч.1. Минск, изд-во “Навука i тэхнiка”. С.12-17.; Кузнецов А.В., 1993. Сравнительная характеристика населения хищных птиц в ряду антропогенно — трансформированных территорий Верхневолжья// Автореферат дисc. … канд. биол. наук. М.: МГПИ им. В.И.Ленина. 23 с.
[23] как она представлена в данном регионе – сеть более или менее устойчивых поселений в разных «пятнах» местообитаний, каждый сезон размножения «наполняемых» особями, и не обязательно местными.
[24] Тарасов А.А., 1994. Сравнительная экология массовых видов врановых птиц в антропогенных ландшафтах Приморского края. Дисс.канд.биол.наук. М. 230 с.
[25] Бокотей А.А., Дзюбенко Н.В., 2008. Поширення та динамiка чисельностi денних хижих птихiв у м.Львовi// Новiтнi дослiдження соколоподiбних та сов. Матерiали III Мiжнародної наукової конференцiї «Хижi птахи України». Кривий Рiг, 2008. C.33-38. см.рис.2 в этой работе.
[26] Характерно, что единственный хорошо документированный случай расселения городских рас (вόроны в Прибалтике) одновременно стимулировал местную урбанизацию. «Урбанизация воронов в Прибалтике резко ускорилась в 1940-1960-е гг. после инвазии с запада «городской расы», освоившей пригороды ряда крупных городов. Результатом стало самостоятельное освоение местными птицами мелких и средних городов вместо прежнего избегания антропогенного ландшафта, и общий рост численности популяции (Вилкс, 1971, цит. по Константинов и др., 1985)».
Рекомендуем прочесть
 Как Европа открыла для себя Московскую Русь, а потом закрыла
Как Европа открыла для себя Московскую Русь, а потом закрыла COVID-19 вскрывает правду
COVID-19 вскрывает правду «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве «Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)
«Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)- Социальная сегрегация в городах
 «Жесты», вόроны и шалашники
«Жесты», вόроны и шалашники- Банковское дело и эволюция целей банковского регулирования
 Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей

{kind=link}