
Дочитал исключительно интересный двухтомник Александра Маркова про эволюцию человека. Интересный в том числе тем, что тезисы и аргументация местами вызвали сильное чувство несогласия, а оно всегда плодотворно: будит мысль, заставляет искать опровержения или по-новому глядеть на материал, лежащий в основе своей собственной концепции. В апреле-мае, как будет время после окончания лекционного курса, напишу про все свои возражения, а сейчас про одно, самое очевидное.
Автор доказывает положительную связь между размером мозга и интеллектом, чему посвящена главка «Анатоль Франс ничего не доказывает». Правда, она начинается не с данных, а с утверждений «Положительная корреляция между размером мозга и интеллектом бесспорно существует». Здесь ждёшь какой – то пример, убедительную картинку в доказательство этой бесспорности – скажем, вот вам размер немецких или японских мозгов, вот какая-то мера интеллекта их носителей (скажем, сумма академической успеваемости), вот смотрите сами, корреляция.
Но картинки нет, и возникают сомнения, тем более что широко известны доводы против существования этой «бесспорной корреляции». Это отнюдь не только Анатоль Франс, хотя его и многих других великих людей с малым размером мозга никак нельзя считать статистическими выбросами. И потому, что самый крупный/самый мелкий мозги были извлечены из черепов идиотов, и число великих людей с мозгом выше среднего примерно соответствует числу великих людей с мозгом ниже среднего, а большинство гениев/крупных талантов имеют-таки средний мозг для своей популяции. В.П.Эфроимсон в своей «выборке патобиографий великих людей», использованной им для поиска биолого-генетических детерминант интенсивной умственной деятельности и, соответственно, гениальности, очень хотел показать такую зависимость, но так и не смог; он осторожно пишет о «высоколобости» и «гигантолобости» у значительной части изученных им гениев и талантов, т.е. повышенный размер лобных долей, а не мозга в целом, и то эта корреляция не значима, на уровне тенденции. Собственно, Эфроимсон и показал, что у гениев «не мозг больше, а мотивация выше», что он связывал с разными веществами — стимуляторами умственной деятельности, в т.ч. с повышенной концентрацией мочевой кислоты в крови, благодаря чему они думатель загружают работой везде, всегда и на полную мощь.
То есть, скорее всего размер мозга, как любой другой признак, в популяциях распределён про оптимальной кривой, в том числе потому, что мозг, очень энергоёмкая ткань, иметь крупные мозги энергетически невыгодно, энергозатраты на развитие и функционирование мозга связаны с его размером аллометрически, степенной зависимостью, т.е. с увеличением мозга будут возрастать быстрее, чем «ум», связанный линейно. Т.е. даже если предположить в качестве отправной точки рассуждения, что гипотеза автора верна, рассмотрение платы и выигрыша от увеличения мозга в филогенезе приведёт к выводу, что главный тренд здесь связан не с увеличением массы, а с усложнением архитектуры, позволяющей в том же объёме разместить больше рабочих элементов и создать тренд миниатюризации устройств. Более совершенные компьютеры не больше, а меньше менее совершенных именно в силу совершенствования архитектуры; поэтому в когнитивных науках успешно используется компьютерная метафора (хотя, как у всякой метафоры, её «область значений» ограничена), а в эволюции мозга в ряду гоминид от неандертальцев до сапиенсов начинается небольшой, но значимый тренд уменьшения размеров мозга.
Но это логические возражения против названной корреляции, куда важней прямые аргументы против существования положительной зависимости позитивной корреляции между размером мозга и умственной деятельностью. Ведь что означает скоррелированность признаков Х и У? Чем больше мозг, тем (в среднем) выше интеллект во всём диапазоне размеров мозга, наблюдаемых в выборке, по которой устанавливается корреляция. И наоборот – чем меньше мозг, тем ниже интеллект в той же выборке. Однако хорошо известно, что у грацильных лилипутов (сохранивших пропорциональность сложения) мозг уменьшился радикально, но интеллект полностью сохранен и часто бывает повышенным, со способностью к научной деятельности. И среди обычных людей радикально малый мозг при сохранных умственных способностях бывает не так уж и редко (в соответствии с нормальной кривой, полагаю): помню, на докладе С.Дробышевского в ИОГЕНе про тенденции эволюции мозга в ряду гоминид он рассказывал про вполне обычного в интеллектуальном плане банковского служащего из ФРГ, у которого мозг по объёму был как у шимпанзе.

Другой пример палеонтологический – известные «хоббиты» с о.Флорес имеют мозги, сократившиеся до размера австралопитековых, однако по уровню культуры и степени развития разных зон мозга они соответствуют архантропам, от которых, видимо, и произошли. Т.е. и тут «структура» важнее «кучи», а сложность организации мозга важнее размеров.
Убедительные доводы «против» даёт одомашнивание птиц и млекопитающих. Редукция мозга вообще характерна для всех одомашненных форм за исключением Mus musculus, причём это не результат аллометрии при изменении в весе тела. Однако домашние особи отнюдь не становятся глупее своих диких предков. Напротив, у них развивается «несобственный» и «вынесенный во внешнюю среду, организованную человеком» ум: ориентируясь на созданные нами социальные подсказки – как жесты и интонации человека, так и детали антропогенной среды – они могут решать когнитивные задачи недоступные диким предкам. В том числе по такого рода решениям собаки опережают шимпанзе и т.д.
Так, например, при доместикации морских свинок Cavia aperea f. porcellus по сравнению с дикими предками мозг уменьшился в среднем на 13% за 4500 лет. И в работе Lars Lewejohann et al. (2011), опубликованной во Frontiers in Zoology, исследовали, действительно ли домашние морсвинки пропорционально глупее диких. Оказалось, нет – умнее. Одомашнивание привело к уменьшению агрессивности свинок, возрастанию у них социопозитивного поведения, усилению поведения ухаживания у самцов и сниженной реакции на стресс (всё как обычно при доместикации). Однако до сих пор когнитивных исследований со сравнением диких с домашними было мало: известна лишь одна работа, где в задачах на дискриминацию фигур единственная дикая морская свинка обнаруживала худшие результаты, чем домашние
Домашних и диких морских свинок, самцов и самок, тестировали на решение задач, связанных с пространственной памятью в водном лабиринте Морриса. Во-первых, оценивали скорость плавания, во – вторых, нахождение подводной платформы с запрыгиванием на неё.
Материал и методы. Для опытов брались животных в возрасте от 35 до 45 дней; всего использовано 15 домашних самцов, 13 домашних самок, 13 диких самцов, 13 диких самок. Домашние свинки – продукт скрещивания разных линий, дикие отловлены в Province, Буэнос-Айрес, Аргентина, в 1974 и 1995 гг. Их разведение в неволе не привело к существенным изменениям в поведении и ответе на гормональный стресс. Каждый день животные взвешивались, причём было подтверждено, что они нормально набирали вес несмотря на тестирование.
Дикие морские свинки – обитатели открытых пространств, использующее короткие периоды кормления и зоны, густо покрытые растительностью, пронизанные сложной сетью тропинок. Это предполагает развитую пространственную память.
Морские свинки могли выучить простые лабиринты или научиться различать разные стимулы, но всё же оказались неудобны для этих экспериментов, даже несмотря на то, что являются удобной моделью для болезни Альцгеймера. Но в лабиринте Морриса они учатся хорошо. Тут требуется найти платформу, скрытую под поверхностью воды в круглом бассейне, окружённом различными пространственными ориентирами. Одомашненные морские свинки хорошо плавают и справляются с этим заданием.
Лабиринт представлял собой круглый бассейн с диаметром 160 см, наполненный водой. Для пространственной ориентации внутрь на белые стенки бассейна помещались разные геометрические фигуры из чёрной клейкой плёнки. Целью было найти платформу в 20 см диаметром, сделанную из полупрозрачного акрилового стекла. Платформа была скрыта в 2 см под поверхностью воды в центре одного из квадрантов, в 30 см от стены.
Тестирование начинали в возрасте 35 дней. Во время обучения свинок тестировали 10 раз за пять дней (два теста в день). Каждый тест длился не более 45 с и начинался с того, что свинку помещали в воду с головой, развёрнутой в сторону от того квадранта, где находилась платформа. В это время позиция платформы была постоянной. Если животное находило платформу за 45 с, его оставляли там на 15 с. Если нет – оно помещалось на платформу руками. Если животное оставалось на платформе менее 15 с, его помещали на платформу ещё дважды. В промежутке между двумя испытаниями все жевотные помещались обратно в домик с возможностью обсушиться в течение 5 мин. Через пять дней обучения животные тестировались в течение 60 с в бассейне без платформы. Считалось, что выработаны пространственные ассоциации, если свинка проводила больше времени в «правильном» квадранте. Через пять минут проводился тест, в котором платформа помещалась в противоположный старому квадрант. Если животное плыло со скоростью, меньшей 0,5 км/ч, то оно изымалось из опытов (но был лишь один самец домашней свинки, который так «дрейфовал»).
Изучать морских свинок очень тяжело из-за их флегматичности. В этом плане водный лабиринт их активизирует. Природные местообитания диких свинок – это обычно влажные места, такие как берега озёр и небольших ручьёв.
Результаты. Самцы и самки домашних свинок достоверно превосходили диких по скорости обучения (рис.1). Только для домашних свинок была характерна достоверная пространственная ассоциация с позицией платформы, для диких наблюдались иные стратегии поиска.
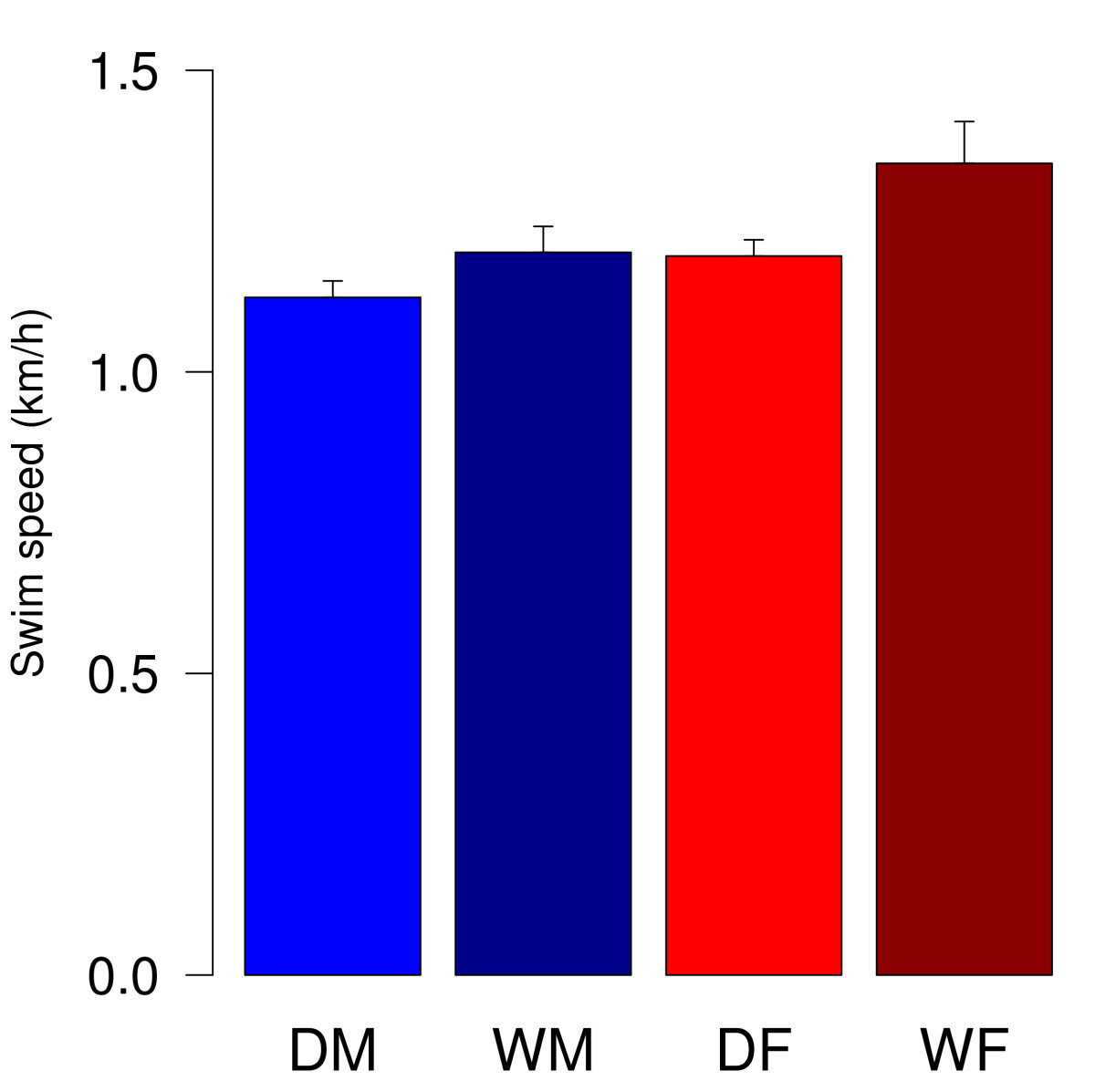
Рис.1. Swim speed in the Morris water maze task. Data represent mean (+SEM) speeds of trial one to ten. DM = male domestic guinea pigs (n = 15), DF = female domestic guinea pigs (n = 13), WM = male wild cavies (n = 13), WF = female wild cavies (n = 13). ANOVA revealed significant effects of sex and domestication (sex: F1, 50 = 6.38, p = 0.015; domestication: F1,50 = 5.21, p = 0.027).
Свинки хорошо плавают (домашние самцы – 1,12 км/ч, домашние самки – 1,19 км/ч, дикие самцы – 1,18 км/ч, дикие самки 1,35 км/ч. И дикие, и домашние свинки находили спрятанную платформу. Во время первой стадии эксперимента характеризовали время нахождения платформы и длину пути, которую свинка проплывала до неё. У домашних свинок эта длина была существенно меньше, чем у диких. Также они несколько быстрее находили платформу (рис.2).
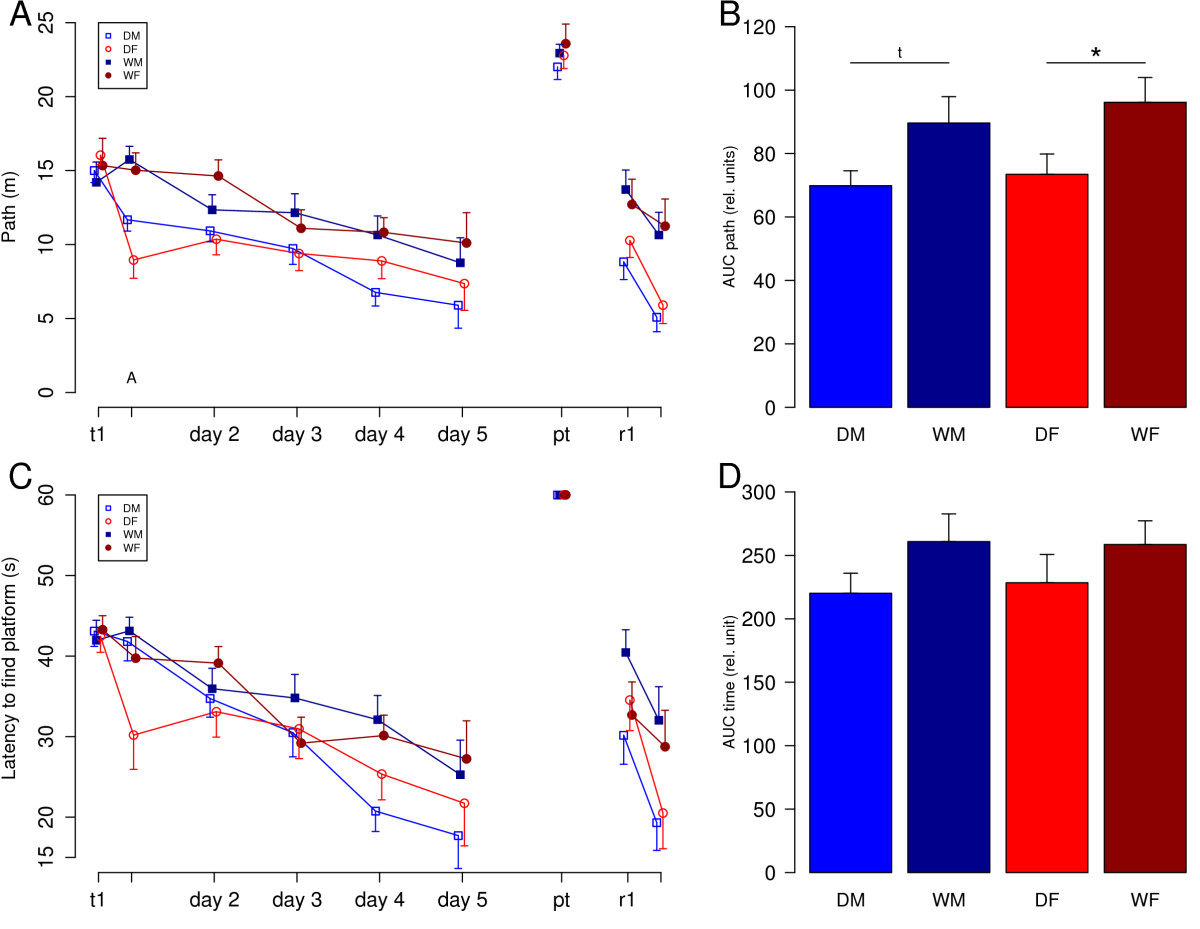
Рис.2. Learning performance in the Morris water maze task. DM = male domestic guinea pigs (n = 15), DF = female domestic guinea pigs (n = 13), WM = male wild cavies (n = 13), WF = female wild cavies (n = 13). A) Learning curve analyzed by path length. The two trials of the first day are depicted separately. Data for day two to day five are combined values of two trials per day. In the probe trial (pt) the platform was removed. Two retention trials (r1 and r2) were conducted on the same day with the platform being moved to the opposite quadrant of the pool. Data represent means and SEMs. B) Comparison of the groups by analysis of the areas under the learning curves (AUC) of the parameter ‘path length’ calculated for the acquisition phase (trial one, day one to trial ten, day 5). Data represent means + SEM. ANOVA revealed a significant effect of domestication (F1, 47 = 9.32, p < 0.01). Statistical symbols of post hoc analysis are depicted in the figure. * = p < 0.05, t = p < 0.1. C) Learning curve analyzed by latency to find the platform. D) Comparison of the groups by analysis of the areas under the learning curves (AUC) of the parameter ‘latency’ calculated for the acquisition phase (trial one, day one to trial ten, day 5). ANOVA revealed effects of domestication as a statistical trend (F1, 47 = 3.02, p = 0.09).
После того, как платформу убирали и свинок тестировали 60с, домашние свинки (как самки, так и самцы) проводили в квадранте, где она раньше была, достоверно больше, чем ¼ (15 мин) времени. Для самцов диких свинок различия были менее значительны, для самок достоверных различий не было.
Когда платформа перемещалась в противоположный квадрант, эффективность обучения оценивалась по изменению времени поиска и длины пути между двумя испытаниями. Статистический анализ выявил достоверное убывание длины пути для всех, кроме диких самок, аналогично выглядела и картина для времени поиска. Для домашних морских свинок и время поиска, и длина пути в последнем тесте были достоверно меньше (рис.3).
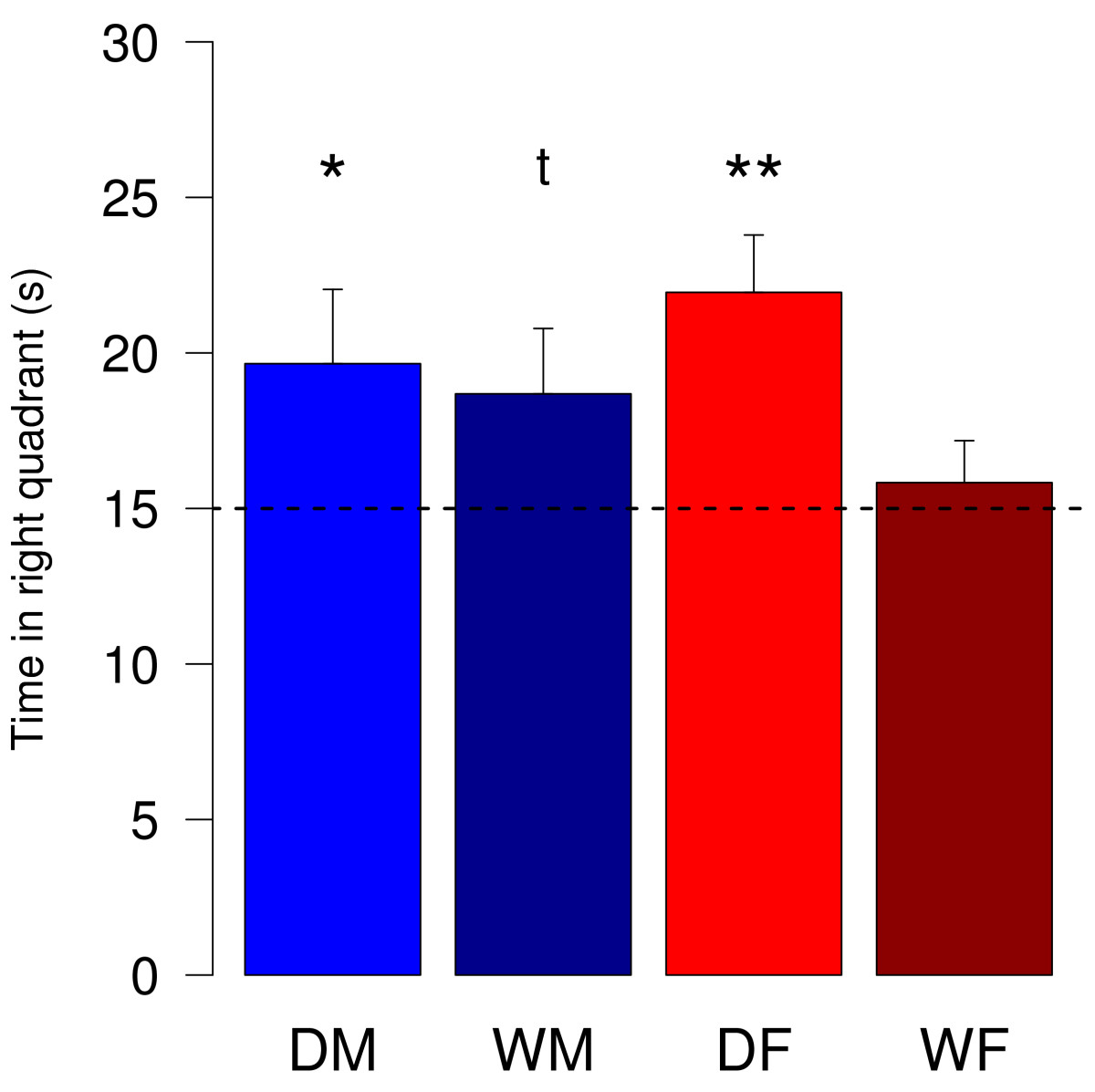
Рис.3. Time spent in the right quadrant of the Morris water maze. In the probe trial the platform was removed and animals explored the pool for 60 s. Data represent mean (+SEM) time the animals were recorded to be in the quadrant of the pool where the platform used to be in previous trials. DM = male domestic guinea pigs (n = 15), DF = female domestic guinea pigs (n = 13), WM = male wild cavies (n = 13), WF = female wild cavies (n = 13). Statistics: one-sample t-test testing the deviation from chance-level (dotted line). ** = p < 0.01, * = p < 0.05, t = p < 0.1.
У диких свинок более сильная реакция на стресс. В другом исследовании та линия, которая сильней реагировала на стресс, чаще показывала поведение «дрейфования». Но в этой работе дикие свинки плыли быстрее, чем домашние. Но со способностями к решению задачи это не коррелировало. Домашние свинки превосходили их в этом плане. Если время решения ими задачи было и ненамного меньше, то путь до платформы они преодолевали гораздо меньший. Тем не менее, характер кривых для диких свинок показывал, что они тоже учатся её решению. Однако данные для случаев, когда платформа была удалена, поддерживают идею, что дикие свинки используют при этом другие навыки (не пространственные). Возможно, что именно высокая скорость, которую развивали дикие свинки, позволяла им использовать какую-то непространственную стратегию. Несмотря на это разница во времени поиска с домашними у них была меньше 10 с, так что их стратегия оказывалась достаточно эффективной. Для изучения собственно пространственной памяти у диких свинок мы намерены применить бОльший бассейн.
Таким образом, одомашнивание может влиять на используемую животными стратегию. Случаи, когда одомашненные животные более успешно выполняли задания, чем дикие, описаны. Так, одомашненные лисы лучше понимали используемые людьми жесты, хотя дикие лучше выполняли задания, не связанные с социальными взаимодействиями. Собаки превосходили волков по способности распознавать коммуникативные сигналы людей. В то же время волки, воспитанные с повседневным общением с людьми, превосходили в этом отношении собак, хотя, возможно, волки были просто более мотивированы к выполнению задания.
Исследовательское поведение у домашних свинок резко сокращено по сравнению с дикими. Поэтому использование сухого лабиринта может приводить к противоположным результатам. В то же время домашние свинки менее нервозны и чувствительны, что приводит к снижению реакции на стресс, а это, в свою очередь, может влиять на обучение и память.
***
Надо сказать, что развитие современных людей из архантропов (также как развитие миролюбивых-эгалитарных карликовых шимпанзе с выдающимися «лингвистическими» способностями из иерархичных-агрессивных обыкновенных шимпанзе) можно описать как доместикацию, которую производили женщины. Соответственно, по мере увеличения социальности людей по сравнению с шимпанзиным «нулём отсчёта» и гиперразвития способности к решению задач путём подражания, доходящей до «чрезмерного подражания» у детей, всё больше возрастала роль «вынесенного ума», опирающегося на социальные взаимодействия и знаки социальной среды, а необходимость в большом мозге уменьшалась – тем более что и усложнение его действовало в ту же сторону.
P.S. Вообще для современных людей «мозг» символизирует «ум» (тогда как в традиционной культуре ум – это чувство), поэтому сравнение размеров мозгов часто несвободно от систематических ошибок, обусловленных доводами вненаучного характера. «Среди интересов Поля Брока (Paul Broca), французского ученого-краниолога XIX века (его имя носит поле Брока, участок лобной доли мозга, отвечающие за речь и часто повреждающийся при инсультах), была привычка измерять мозг, и его всегда возмущал тот факт, что немецкие мозги были граммов на сто тяжелее, чем французские. Поэтому он решил, что при измерении веса мозга во внимание должны приниматься другие факторы, например общий вес тела: это объясняло, к его удовлетворению, больший вес немецких мозгов. Но для своей выдающейся работы о том, что у мужчин мозг больше, чем у женщин, он такой поправки не сделал. Случайно или намеренно, кто знает?» (Бен Голдакр. «Обман в науке», за книжку спасибо homers_wife).
UPD. Как я понимаю, данные, приведённые catta в «Размер и его значение» поддерживают скорей мою т.з., чем автора книги:
«Вернемся к волнующей теме эволоции мозга. Меня спрашивали о двух вещах: о пропорциональности роста разных областей коры и об увеличении размера мозга как такового. Я сейчас напишу про размеры, просто потому что попалась хорошая работа на эту тему: Suzana Herculano-Houzel. Not All Brains Are Made the Same: New Views on Brain Scaling in Evolution// Brain, Behavior and Evolution 201; 78: 22–36
В эволюционном ряду млекопитающих размер мозга постоянно растет. Поэтому вопрос связи функций мозга с размером кажется на первый взгляд вполне тривиальным: чем больше, тем лучше. Согласно традиционной точке зрения, по мере увеличения мозга должно происходить следующее:
— очевидное увеличение количества нейронов
— укрупнение нейронов ( потому что удлинение связи требует увеличения размера клетки для поддержения того же качества передачи сигнала)
— рост числа глиальнйих клеток в пересчете на один нейрон (глиальные клетки занимаются обеспечением нужд нейронов, и они в размере не растут, поэтому на крупный нейрон их нужно больше), то есть, должна падать плотность распределения нейронов в ткани мозга.
Площадь коры должна расти быстрее, чем размер всех остальных структур включая мозжечок, т.к. эволюция мозга мыслится прежде всего как увеличение мощности коры.Автор анализирует все перечисленные тенденции на примере 28 видов млекопитающих из трех разных групп: Насекомоядные, Грызунообразные (грызуны и зайцеобразные), Приматы и родственные приматам Tупайи.
Вопреки существующему мнению об общности закономерностей эволюционного развития мозга оказалось, что мозг увеличивался не одним и тем же способом, плотность нейронов и отношение количества глии к количеству нейронов на ту же массу мозга может сильно различаться, так что того же размера мозг у разных млекопитающих может вмещать различнoe количество нейронов. В дополнение к тому выяснилось, что кора и мозжечок с одинаковой силой концентрируют нейроны по мере роста мозга, то есть эволюционный тренд на самом деле заключается не в росте коры, a в росте коры и мозжечка. Единственный параметр, пропорционально меняющийся с увеличением мозга независимо от группы животных — число глиальных клеток. Тут все просто: чем больше масса мозга, тем больше глиальных клеток.
Вес мозга в целом коррелирует с размером тела. Однако, эта зависимость также не одинакова по группам животных: приматы всегда имеют более крупный мозг, чем грызунообразные и насекомоядные тех же размеров.
Но cамое главное — различия в количестве нейронов. При той же массе мозга приматы имеют большее количество нейронов по сравнению с остальными животными. (Эти различия наблюдаются и между другими группами, например, насекомоядные на ту же массу тела имеют больше нейронов, чем грызунообразные. Ежики умные, в общем.) Кора мозга аналогичных размеров может очень сильно отличаться по плотности нейронов в разных группах, например кора массой 17.7 г у агути содержит около 795 миллионов нейронов, a кора совиной обезьяны массой 15.5 г — 1.5 миллиарда нейронов. Чем больше масса коры, тем сильнее отличие приматов от остальных групп животных. То же самое действительно и для увеличения мозжечка. При этом у всех животных не исключая человека кора обычно содержит около 20% всех нейронов мозга, в то время как мозжечок — около 70%. Bсе остальные мозговые структуры своей плотности не повышают, число нейронов в них растет с размером равномерно для всех животных.
Иными словами, приматaм удалось каким-то образом не только увеличить размер мозга, но и миниатюризировать его структуру. Судя по тому, что у нас размер нейронов в меньшей степени растет с размером мозга, произошел какой-то апгрейд в эффективности передачи сигнала.
А сам по себе размер ничего еще не гарантирует».
Источник wolf_kitses
Рекомендуем прочесть
 Как Европа открыла для себя Московскую Русь, а потом закрыла
Как Европа открыла для себя Московскую Русь, а потом закрыла COVID-19 вскрывает правду
COVID-19 вскрывает правду «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве «Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)
«Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)- Социальная сегрегация в городах
 «Жесты», вόроны и шалашники
«Жесты», вόроны и шалашники- Банковское дело и эволюция целей банковского регулирования
 Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей
