
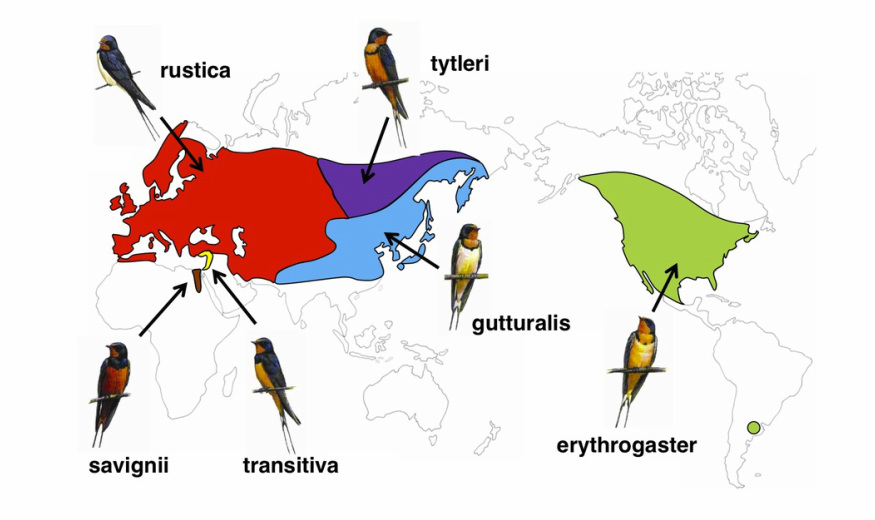
Ареал и подвидовая дифференциация деревенской ласточки. Источник
В продолжение темы, как конкуренция портит науку:
Грантовая система, сплавленная с «библиометрическим джихадом» (оценка продуктивности исследователей по публикациям-импактам-цитированиям), с одной стороны, рождает «плохие практики», вроде описанных в полевой зоологии, с другой — снижает воспроизводимость исследований, с третьей — достаточно часто делает лидерами направления и «образцами исследования», в куновском смысле, скажем так, не лучшие варианты, c четвёртой — способствует развитию имитативной науки. Первое и второе известны и достаточно широко обсуждаются; ниже мы рассказываем про третье на примере такого модного направления, как социобиологические интерпретации полового отбора из книги Е.Н.Панова про сабж. Про четвёртое будет дальше.
«Панов Е.Н.
Идея полового отбора была выдвинута Дарвином в попытке объяснить причины, лежащие в основе резко выраженного полового диморфизма у животных. Он, как правило, состоит в том, что ко всем тем признакам, которые присутствуют у самок, у самцов добавляется еще нечто такое, что, по мнению классика, не является необходимым для выживания особи. Если бы самцу не нужно было бы оставить потомство, он бы вполне мог обойтись без того, что Дарвин называл, в частности, яркой окраской (gaudy coloration) и всевозможными украшениями (various ornaments). Если бы не эти качества, которые самцы используют в конкуренции из-за самок, «они во всех прочих отношениях, если сравнивать их самками, были бы столь же хорошо, как и те, приспособлены к повседневному существованию (for their ordinary habits of life)» (Darwin, 1871: 257).
Если остановиться, до поры до времени, на тех структурах, которые классик считал «украшениями», возникает вопрос, где провести границу между этой категорией и всем тем, что необходимо самцу для собственного успешного существования. Далее в этом разделе я собираюсь показать, что ответ на этот вопрос совсем не так прост, как может показаться на первый взгляд.
В конце 1980-х гг. один из апологетов «теории полового отбора», датчанин Андрес Мёллер, посчитал, что к числу таких «украшений» следует отнести удлиненные перья хвоста у самцов деревенской ласточки Hirundo rustica. В европейских популяциях этого вида длина хвоста самцов составляет в среднем 103.4 мм (разброс значений 89-119 мм), самок 86.9 мм (70-99 мм). Как видно, имеется небольшое перекрывание. Длина крайних рулевых у самцов — около 6 см, у самок — около 4.5 см. Иными словами, крайние рулевые самок составляют в среднем около 75% от их длины у самцов (Cramp, 1988)[1]. Самцы и самки окрашены одинаково и различаются только по этому признаку.
Гипотеза 1: удлиненные перья хвоста у самцов деревенской ласточки как продукт полового отбора
Содержание
- 1 Гипотеза 1: удлиненные перья хвоста у самцов деревенской ласточки как продукт полового отбора
- 2 Гипотеза 2: удлиненные перья хвоста у самцов деревенской ласточки снижают приспособленность их обладателей
- 3 Гипотеза 3: самки ласточек предпочитают самцов с симметрично удлиненными крайними рулевыми
- 4 Оценка результатов Мёллера в 1990-е гг.
- 5 Вопросы, возникшие относительно достоверности выводов Мёллера
- 6 Как вопрос о сущности удлиненных перьев хвоста у деревенской ласточки видится с более широких биологических позиций
- 6.1 Строение хвоста деревенской ласточки способствует улучшению маневренности полета
- 6.2 Удлиненные крайние рулевые не увеличивают стоимости (cost) локомоторной активности.
- 6.3 Длина крайних рулевых у самца деревенской ласточки не может служить для самок показателем «перспективности» самца.
- 6.4 Самка деревенской ласточки выбирает не самца, а гнездовой участок, где есть прошлогодние гнезда.
- 7 P.S. Книга Е.Н.Панова больше о минусах «конкурентной науки», чем о дефектах теории полового отбора
В период с 1988 по 1992 г. Моллер опубликовал 10 статей на тему полового отбора у этих ласточек[2]. В первой из них он без колебаний заявил, что его исследование полностью подтверждает взгляды Дарвина и Фишера. На основе экспериментов по искусственному удлинению хвостовых перьев самцов он пришел к выводу, что особи, подвергшиеся этой операции, быстрее приобретают половых партнеров и достигают большего репродуктивного успеха (Møller, 1988).
Гипотеза 2: удлиненные перья хвоста у самцов деревенской ласточки снижают приспособленность их обладателей
Коль скоро, по Дарвину, половой отбор действует наперекор естественному, те признаки, которые обязаны своим происхождением половому отбору, должны, по его мнению, создавать их носителям дополнительные трудности, в частности потому, что повышают заметности таких особей для хищников (см. 1.1, III). Именно эту идею попытался подтвердить Мёллер в своей следующей статье на интересующую нас тему. Он полагал, что подтвердил ее справедливость экспериментально при помощи все той же методики изменения длины рулевых перьев. Он пишет:
«Я показал, что …самки отдают предпочтение самцам с удлиненными перьями перед теми, у которых они укорочены. Но [первые] …добывают более мелких жертв с пониженной пищевой рентабельностью, [нежели вторые]. Снижение эффективности кормодобывания у самцов с удлиненными перьями хвоста увеличивает дефекты рулевых, вероятно (probably) из-за ухудшения питания во время линьки. У этих самцов при линьке также уменьшается длина хвоста. За счет этого происходят потери их приспособленности: более позднее начало размножения на следующий год и, следовательно, снижение репродуктивного успеха в это время» (Møller. 1989: 132).
При чтении этой статьи напрашивается вопрос, каким должен быть первичный эмпирический материал, чтобы сделать все эти выводы, и можно ли получить репрезентативные выборки (например, по размерам жертв у двух категорий самцов) при использовании применявшихся экспериментальных методов? Как мы сможем убедиться далее, сомнения на этот счет совсем не лишены основания.
В середине 1970-х гг. идея высокой стоимости самцовых «украшений» была переформулирована в так называемый «принцип гандикапа» (см. 1.1, 111). Его автор А. Захави еще раз представил расширенный вариант этих своих построений 18 лет спустя (Zahavi, 1975. 1993)[3]. Так или иначе, они стали весьма популярными в западной литературе к моменту начала активной деятельности Мёллера. Чтобы не отстать от моды, он выпускает в соавторстве с одним из своих единомышленников статью под названием «Дифференциальная стоимость (costs) вторичных половых признаков: экспериментальная проверка принципа гандикапа» (Møller, de Lope, 1994).
Гипотеза 3: самки ласточек предпочитают самцов с симметрично удлиненными крайними рулевыми
Один из постулатов «теории» полового отбора состоит в том, что генетически качественные самцы обладают более крупные и более симметричными сигнальными структурами, а низкокачественные — меньшими по размерам и асимметричными (см. l.l, 111). Попыткам верификации этих взглядов была посвящена очередная статья Мёллера. В ней он пишет:
«Я манипулировал независимо двумя признаками: длиной крайних рулевых и степенью их симметрии у самцов деревенской ласточки, чтобы выяснить, какой из них самки используют при выборе полового партнера. Самцы с удлиненными, симметричными перьями приобретают половых партнеров раньше тех, у которых эти перья укорочены и асимметричны. Таким образом, самки при выборе партнеров расценивают как величину украшения, так и его симметричный характер в качестве надежного показателя качества самца» (Møller. 1992: 238).
Эта заметка, опубликованная на двух страницах журнала Nature, была через два года представлена в расширенном виде в журнале Evolution (Møller, 1994).
Оценка результатов Мёллера в 1990-е гг.
Как полагал Андерссон, имея в виду все три гипотезы, рассмотренные выше, «Половой отбор на орнамент вторичных половых признаков был на примере деревенской ласточки экспериментально продемонстрирован [Мёллером]» (Andersson 1994: 159). Впрочем, этот автор отмечает ряд отличий между результатами Мёллера и теми, что были получены в ходе аналогичного исследования, проведенного в Канаде (Smith, Montgomerie, 1991: Smith et al.. 1991). По их данным, самцы с удлиненными перьями начинают гнездиться раньше, чем те, хвосты которых были укорочены, но это не приводит к повышению репродуктивного успеха первых. Андерссон объясняет эти различия между датской и канадской популяциями различиями в использованных экспериментальных процедурах, а также тем, что во второй из них «половой отбор не столь силен», как в первой. По данным ДНК-дактилоскопии, в гнездах самцов с удлиненными хвостами менее чем две трети птенцов оказались их прямыми потомками, тогда как самцы с укороченными хвостами оказались отцами всех птенцов, находившихся в их гнездах. Эти различия объясняли тем, что «удлинение хвостов, возможно, подавляет (hamper) способность самцов препятствовать их самкам вступать во внебрачные копуляции»[4] (Andersson 1994: 159). Интересно, что у контрольных самцов канадской популяции (не подвергавшихся манипуляциям) обнаружена обратная зависимость между длиной крайних рулевых и пропорцией внебрачных птенцов в гнезде (Smith et al., 1991).
Вопросы, возникшие относительно достоверности выводов Мёллера
В конце 1990-х гг. у двух исследователей из Университета Копенгагена возникли сомнения по поводу утверждения, согласно которому степень асимметрии в проявлении признака может служить показателем приспособленности его носителя. Они обратились к статье Мёллера с соавторами, которая подытоживала данные первого за 1990-1996 гг., где такая взаимосвязь декларировалась наиболее категорично (Møller et al., 1999). Заподозрив неладное при прочтении статьи, Г. Начман и К. Хеллер попросили Моллера прислать им первичные данные. После длительной переписки тот весьма неохотно все же выполнил их просьбу, но в присланном материале все измерения были даны скопом, без разбивки на выборки по годам, как это было сделано в таблице в оригинале статьи.
Оказалось, в частности, что не все величины в присланном материале совпадают с приведенными в этой таблице. Грамотная статистическая обработка первичных данных показала, что выводы Моллера были неадекватными. Результаты проделанной ими работы Начман и Хеллер отразили в статье под названием «Флюктуирующая асимметрия как показатель приспособленности: причинность или статистический артефакт?» (Nachman, Heller, 1999).
Материалы, полученные этими двумя авторами от Мёллера, были дополнительно проанализированы Дж. Раболом, который разбил всех самцов на три группы с разными показателями асимметрии (менее 20, 18 и 12 мм) и сравнил множественные логистические регрессии для них и для всей выборки в целом. Как пишет этот автор «..коэффициенты регрессии оказались отрицательными для всех четырех выборок, тогда как в таблице из статьи Моллера с соавторами они показаны положительными для каждого из семи лет. Возможно ли, — вопрошает автор, — чтобы негативный коэффициент для выборки в целом складывался из позитивных для каждого из семи лет исследований?» (Rabøl, 2007).
Поскольку это был уже второй такой случай в биографии Моллера, осенью 2003 г. он был обвинен датским научным сообществом (в лице Danish Committees on Scientific Dishonesty) в фальсификации фактов[5] и лишен права отлова птиц для научных исследований (там же; Borrell, 2007). Теперь читателю станет понятно, почему я с определенным недоверием отношусь к первым публикациям этого автора, где говорится о распознавания самками самцов, чьи крайние рулевые различаются на доли сантиметра[6]. Как писал Дж. Вилкинсон из Университета Мэриленда по поводу сказанного Мёллером о роли асимметрии этих перьев у ласточек.
«Результаты были слишком поразительными (amazing), чтобы в них охотно поверили. Именно это и заставило нас ближе присмотреться к сказанному в статье» и найти в ней противоречия между текстом и графиками (Borrell, 2007).
Как вопрос о сущности удлиненных перьев хвоста у деревенской ласточки видится с более широких биологических позиций
В одной из недавних статей на эту тему сказано:
«Изучение морфологии и гнездовой биологии ласточки касатки, проводимое Андерсом Мёллером и другими на протяжении последних трех десятилетий, породило несметное количество (incredible amount) заявлений о роли крайних рулевых у особей этого вида в половом отборе. Однако недавние исследования динамики полета ласточек дают основание полагать, что эволюция этих структур была движима скорее естественным, нежели половым отбором» (аноним, http://people.bu.edu/msoren/ReviewPaperExample.pdf).
Далее я рассмотрю те изменения в трактовках интересующего нас явления, которые стали возможными после некоторого ослабления «тисков» парадигмы полового отбора. В истории науки это происходит всегда, когда господствующая доктрина утрачивает новизну и становится общим местом. Ее давление на умы ученых начинает ослабевать (по принципу привыкания к стимулу в этологической концепции поведения), так что появляется возможность подумать об альтернативных объяснениях, зачастую много более реалистичных. В данном случае речь идет о роли крайних рулевых в осуществлении повседневной активности ласточек; о том, являются ли эти структуры показателем качества самца и индикатором его будущего репродуктивного успеха; а также о некоторых более частных моментах, ставящих под сомнение всю концепцию полового отбора.

Строение хвоста деревенской ласточки способствует улучшению маневренности полета
В серии работ британских исследователей (Evans, Thomas, 1997; Evans, 1998; Buchanan, Evans, 2000; и др.) они использовали тот же метод манипуляций с длиной крайних рулевых, которому следовал Мёллер (рис. 2.5). Было показано, что их присутствие улучшает аэродинамические показатели полета ласточек. Как ранее установил Норберг (Norberg, 1994), они обеспечивают подъем на малых скоростях и повышают угол атаки, если рулевые расправлены веером. Анализ видеозаписей полета позволил оценить ряд его характеристик, в частности, средний показатель изменений частоты виражей. Ученым удалось показать, что аэродинамический оптимум достигается при длине крайних рулевых в пределах от интактного их состояния до увеличенного на 2 см. Таким образом, как полагают авторы, если считать, что эти структуры эволюционировали в основном под действием естественного отбора на эффективность полета, то за счет полового отбора они могли, в принципе, удлиниться не более чем на 1 см (Buchanan, Evans, 2000).
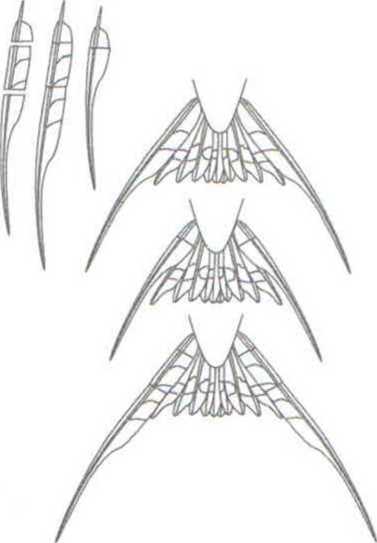
Рис. 2.5. Схема манипуляций с крайними рулевыми хвоста деревенской ласточки (обрезание и приклеивание обрезанных фрагментов). Из: Evans, Thomas, 1997.
Речь идет об эффективности полета при ловле ласточками их жертв. Известно, что в питании деревенской ласточки преобладают насекомые достоверно более крупные, чем у тех видов птиц, кормящихся на лету, которые лишены удлиненных крайних рулевых (ласточки городская Delichon urbica и береговая Riparia riparia[7], а также стриж Apus apus – Архипенко и др., 1968; Bryant, Turner, 1982). А охота на крупных летающих насекомых требует гораздо большей маневренности полета, чем ловля мелких (Cramp, 1988: 267).
В то время как Иванс с коллегами в работах, о которых речь шла выше, допускают, что в развитие удлиненных рулевых перьев у деревенской ласточки наравне с естественным отбором мог участвовать также половой отбор, в последующей статье с участием тех же авторов эта идея не отрицается, а как бы уходит на задний план. Здесь определенно заявлено, что «… крайние рулевые (tail streamers) могли первоначально возникнуть на основе естественного отбора ради увеличения маневренности полета», и что воззрения, согласно которым эти структуры следует рассматривать в качестве гандикапа, неубедительны (Park et al., 2001: 446).
Удлиненные крайние рулевые не увеличивают стоимости (cost) локомоторной активности.
Один из постулатов «теории» полового отбора состоит в том, что структуры, выработанные на его основе, должны обладать высокой стоимостью, создающей дополнительную нагрузку на организм. В отношении деревенской ласточки это попыталась доказать группа исследователей с участием Моллера (Saino et al., 1997). Они измеряли показатель гематокрита[8] у самцов с искусственно измененной длиной крайних рулевых. Идея состояла в том, что у тех из них, у которых эти перья удлинены, этот показатель должен быть выше, чем у самцов с укороченными перьями. Именно такой результат был достигнут в одной из двух исследованных популяций. В резюме к статье сказано: «Сравнительно высокий показатель гематокрита у самцов с экспериментально удлиненными орнаментными перьями может быть ответом на увеличение энергетических требований и повышенного потребления кислорода при полете, обусловленных морфологией их хвоста».
Аналогичное исследование было проведено восемью годами позже в Шотландии (Cuervo, de Ayala, 2005). Исследователи исходили из тех же предположений, что и авторы предыдущей работы, но получили совершенно иные результаты. Они установили, что уровень гематокрита наиболее высок у ласточек по окончании весенних миграций (интенсивная локомоция), но снижается по ходу репродуктивного сезона. Как оказалось, у птиц с интактными длинными хвостами, родительский вклад которых был невелик (невысокая локомоторная активность), показатели гематокрита снижались более значительно, чем у особей с первоначально более короткими крайними рулевыми (они были еще более укорочены на 21 мм), родительский вклад которых был близок к максимуму (высокая локомоторная активность). Как пишут авторы, «Эти результаты не подтверждают гипотезу, согласно которой крайние рулевые у деревенской ласточки удлинялись под действием полового отбора». Они добавляют: «… по крайней мере в изученной популяции», и что именно географическая изменчивость может быть причиной несовпадения полученных данных с результатами более ранних исследований. Однако, возможно и более простое объяснение, которое я приведу в комментарии к следующему пункту 3.
Длина крайних рулевых у самца деревенской ласточки не может служить для самок показателем «перспективности» самца.
К. Нейман с соавторами недавно повторили в штате Нью Йорк (США) исследования европейских орнитологов, работая с птицами местного подвида Hirundo rustica erythrogaster. Они намеревались решить те самые задачи, которые более десятилетия назад были поставлены на повестку дня Мёллером. Результаты этого исследования не подтвердили ни один из тезисов, которые следовали из материалов последнего. Эти авторы пишут, что длина крайних рулевых самца не может служить показателем 1) его успеха у самок, 2) репродуктивного успеха в данном сезоне и 3) присутствия и количества внебрачных птенцов в его гнезде (Neumanctal„ 2007).
Эти авторы объясняют полное несовпадение своих данных с теми, что были получены Мёллером и его коллегами следующим образом:
«Наши результаты подтверждают сложившееся мнение (recent suggestions), согласно которому у данного вида функции половых сигналов варьируют географически».
Однако объяснение может быть намного более простым. В своем обзоре исследований по избирательности самок к облику самцов у деревенских ласточек в Европе они цитируют исключительно работы Мёллера, опубликованные им самим в период между 1988 и 1995 г. (пять статей), либо в соавторстве (Мøller, Tegelstrӧm, 1997; Saino, Primmer, Ellegren, Мøller, 1997). Можно полагать, что подход американских исследователей отличался от того, который, как известно, практиковался Мёллером (неясно, насколько часто) и состоял не в том, чтобы узнать истину, а в желании подтвердить свои предвзятые идеи (см. выше и 10.3).
Самка деревенской ласточки выбирает не самца, а гнездовой участок, где есть прошлогодние гнезда.
Многократно показано, что самки птиц при выборе места гнездования ориентируются скорее на качество такого участка, нежели па конституцию пребывающего здесь самца. Это относится как к видам с территориальной моногамией, так и к тем, что размножаются на токах. Все это можно считать строго установленным фактом (см. материалы Главы 6).
При ближайшем рассмотрением не исключением оказалась и деревенская ласточка. Это убедительнейшем образом показано в превосходной работе Ребекки Сафран (Safran, 2007), одного из соавторов исследования, рассмотренного выше, в пункте 3. Из приведенного там текста следует, что самки американского подвида деревенской ласточки не отдают очевидного предпочтения самцам с наиболее длинными крайними рулевыми. Сафран попыталась выяснить, существует ли предпочтение самок к наиболее ярко окрашенным самцам. Ответ и здесь оказался отрицательным. В задачу автора входило также проверить справедливость утверждения, высказанного Мёллером и Биркхедом (Møller, Birkhead, 1993), согласно которому внебрачные копуляции у птиц более обычны при высокой плотности гнездования. Она пишет:
«На основе наблюдений и с использованием экспериментальных техник я проверила справедливость гипотез, согласно которым выбор места гнездования самок обусловлена (1) их предпочтениям к ярко окрашенным самцам и (2) повышению вероятности участия во внебрачных копуляциях в местах коллективного гнездования, но не нашла подтверждения им».
Все оказалось гораздо проще. Особи разных полов концентрируются в таких местах, где наиболее вероятно найти прошлогодние гнезда. Эти места наиболее предпочтительны для самок, гнездящихся впервые (Safran, 2007: 1360; рис. 2.6).
«Таким образом, – пишет Сафран, – не удалось показать, что окраска самцов как таковая, а также прочие показатели (indicators) их качества, которые я принимала во внимание, оказывают серьезное влияние на выбор самками мест гнездования. Вероятно, самка выбирает самца, который обладает территорией, где располагается старое гнездо и останавливается здесь как можно раньше. Поскольку внебрачные копуляции есть норма (prevalent) у данного вида, можно полагать, что самка сначала выбирает место гнездования, а лишь затем вносит поправки при контактах с [потенциальными] половыми партнерами» (Safran, 2007: 1365, 1366)».
Половой отбор: теория или миф? М.: Т-во научн.изд. КМК, 2015. раздел 2.2. С.59-66.

P.S. Книга Е.Н.Панова больше о минусах «конкурентной науки», чем о дефектах теории полового отбора
Подчеркну: это и другие слабые и/или ложные доказательства действия полового отбора, во множестве собранные Евгением Никролаевичем в книге, показывают отнюдь не отсутствие этого феномена в природе, и не его тождество с естественным отбором,как хочется думать автору.
[я сам могу добавить парочку, на больших фрегатах Fregata minor и королевских пингвинах Aptenodytes patagonicus. Вполне корректно показав преимущества самцов с более развитыми «украшениями» в конкуренции с другими самцами (в первом случае) или их предпочтения самками (во втором) авторы успокаиваются и думают, что дело сделано, и не доводят доказательство действия полового отбора до конца — демонстрации, что самцы с более развитыми «украшениями» а) чаще спариваются, чем с менее развитыми и б) эти спаривания реально увеличивают репродуктивный успех таких особей. Потому что явное преимущество особей, оказывающихся «лучшими» на одном из этапов реализации репродуктивного потенциала, может быть перечёркнуто или обращено в противоположность на следующих, как в случае с «сигналами овуляции» у людей.]
Поэтому, старательно и не без блеска деконструируя додумки, преувеличения и артефакты (увы, слишком часто встречающиеся среди исследований этого рода, причины чего см.ниже), он как-то забывает упомянуть более чем разнообразные исследования, где действенность полового отбора показана строго. См. их для птиц, амфибий, грызунов, насекомых, снова птиц и т.д.
Другое дело, что вопреки социобиологическ
Во-вторых, в общем случае половой отбор симметричен у самцов и самок; те и другие интенсивно конкурируют за особей противоположного пола, создавая сравнимые вариансы по приспособленност
Увы, существенная часть исследований в этой области имеет «больные места», в которые противник теории может радостно ткнуть. Как и «ласточкин хвост», они вполне заслуживают «ювеналова битча», роль которого Е.Н.Панов выполнил на отлично. Почему? Книга Евгения Николаевича показывает — из-за минусов «конкурентной науки»: оценка исследований по «импакта
Что крайне прискорбно потому, что сильно продляет жизнь «некачественных» концептов, для которых уже известны опровергающие их данные, от наиболее общих («отбор родичей», «правило Бейтмана») до вполне частных («обмен еды на секс», «сигналы овуляции»). Или которые подтверждаются лишь в определённых условиях, однако претендуют на универсальность.
Верно и обратное: «ложные практики», созданные господством конкурентной науки, даже в наиболее безобидном варианте («оформлении» интересных и тщательно полученных данных терминологией модных концепций, с подгонкой результатов под их выводы) сильно облегчили задачу Евгению Николаевичу — доказать вопреки очевидности , что «полового отбора не существует». Ему было достаточно перебрать большинство исследований, обнаружить невязки данных и/или методов с концептуальной основой и интерпретацией результатов (не говоря об откровенно некачественных работах, наличие которых «в ряду прочих» дискредитирует данное направление особенно сильно) и сказать «вот тут сомнителен метод, тут натянуты выводы — как можно утверждать существование Х на столь шаткой основе?». Одновременно он опустил те (немногие в силу вышесказанного) работы, где действие полового отбора показано строго — и дело сделано; так что по этой схеме можно «опровергать» очень многое.
Фактически вся длинная книждка Е.Н. — это демонстрация системных изъянов «западной» организации науки больше, чем теории полового отбора. А также критика социобиологическ
Примечания
[1] Или даже 85%, по утверждению Андерссона (Andersson, 1994: 159).
[2] Собственно, говоря, именно многочисленные статьи этого автора в значительной степени стимулировали возрожден интереса к теме полового отбора (см. ниже, 2.4).
[3] О надуманности этих взглядов, ведущих в методологический тупик см. Панов, 2012б, глава 1
4 Произвольность и беспочвенность такого «объяснения» самоочевидна.
[5]Мёллера уличили также в плагиате.
[6]Андерссон. уверенный в реальности полового отбора, изложил их в своей книге как доказанный факт.
[7] Средняя масса жертв составляет 6.01 ± 5.64 мг у деревенской ласточки. 1.88 ± 1.49 у городской и 1.26 ± 1.28 у береговой. В питании последнего вида быстро летающие двукрылые весом от 90 до 120 мг и выше отсутствуют полностью, тогда как у деревенской ласточки они составляют 72.1-83.3% добычи (Bryant, Turner, 1982).
[8] Гематокрит – часть объёма крови, приходящаяся на эритроциты или же на все ее форменные элементы (эритроциты, лейкоциты, тромбоциты). Показатели гематокрита выражаются в процентах к общему объему крови.
Рекомендуем прочесть
 Разбор популярных мифов про охрану природы
Разбор популярных мифов про охрану природы «Кровавые когти и клыки профессора Пинкера». Ч.1.
«Кровавые когти и клыки профессора Пинкера». Ч.1. Бедность, коллективистская культура и успехи в учёбе африканцев
Бедность, коллективистская культура и успехи в учёбе африканцев Экологические аспекты гидростроительства и идеи Виттфогеля
Экологические аспекты гидростроительства и идеи Виттфогеля Науку коррумпируют не только фармкомпании, но капитализм в целом
Науку коррумпируют не только фармкомпании, но капитализм в целом Про «приручение вирусов»
Про «приручение вирусов» «Неудобное наследство…» и социальная ответственность учёных
«Неудобное наследство…» и социальная ответственность учёных Взлёт и падение универсальных «базовых эмоций» Экмана
Взлёт и падение универсальных «базовых эмоций» Экмана
