

Я много писал, что формы демонстраций ухаживания, угрозы, умиротворения и пр. у животных столь же произвольны, как произволен знак: см. раздел про это в «От стимула к символу». Для более примитивных сигналов-стимулов («указывающих» на мотивацию отправителя) это верно не менее, чем для эволюционно продвинутых сигналов-символов, «указывающих» на ситуацию во внешнем мире животного (одну из категорий проблем, в которую особь «втянута» данным взаимодействием и по ходу последнего вынуждена разрешать). В общем случае внешний вид демонстрации не «показывает» ни её «назначения», ни мотивации отправителя, вопреки антропоморфной интуиции что, скажем, агрессия маркируется попытками «стать больше», «развернуть корпус», «поднять хохол», вообще «стать больше», чтобы «устрашить оппонента». См. хороший пример — исследованная Paweł Ręk & Tomasz S. Osiejuk (2010) сигнализация об агрессии у коростелей Crex crex:
«Видовой крик коростеля «крекс-крекс» состоит из двух слогов, которые самец воспроизводит с разной частотой снова и снова. Первый слегка короче, чем второй. Ключевой параметр ритма креканий, по гипотезе авторов кодирующий уровень агрессивности самца – это интервал между последовательными криками Int2 (a не слогами – Int1). Интересно, что внешний слог в структуре крика представляет собой маркёр индивидуальности птицы и стабилен на протяжении нескольких лет, это как бы её «личная подпись» (Peake et al. 1998; Peake and McGregor 1999). См.рисунок 1 статьи (а – монотонный ритм подачи креканий, b – прерывистый, S1, S2 – продолжительность слогов, см.ниже)».
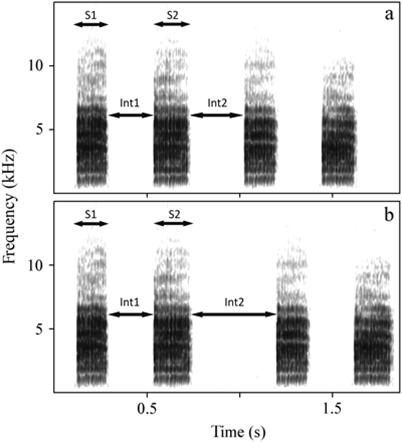
Соответственно, ритм подачи креканий определяется соотношением двух временных интервалом – между криками (Int 2) и между слогами внутри крика (Int 1). Если пауза между криками близка по длине к паузе между слогами, коростель выдаёт монотонный ритм. Когда интервал между криками значимо больше интервала между слогами, выдаётся прерывистый ритм. То есть всё разнообразие переходов от строго монотонного ритма к максимально прерывистому можно охарактеризовать численно (эту переменную называли «ритм», она варьирует от 0,4 до 1,3).
Ритм подачи крика может меняться в течение ночи, в течение сезона и в целом становится более прерывистым за время охраны территории. В совокупности с сезонным эффектом более прерывистый ритм позитивно связан с массой тела самца. Однако, в каждый отдельный момент времени особь свободно меняет ритм подачи крика, в том числе в ответ на крик других коростелей или на проигрывание их криков через магнитофон.
Авторы предположили, что ритм подачи крика – конвенциональный сигнал, указывающий на уровень агрессивности самца, то есть на вероятность удержать территорию в окружении конкретных соседних самцов с их ритмами. Причём изменение ритма в сторону большей прерывистости отражает увеличение агрессивности, в сторону монотонности – уменьшение. Важно подчеркнуть: во втором случае птица тратит на звукопроизведение больше энергии, чем в первом, то есть более эффективный сигнал обходится дешевле. В таком случае во время взаимодействия коростели предпочтительно атакуют соперников, сигнализирующих о сходной агрессивности (кричащих в близком ритме), но летят прочь от противников, сигнализирующих о более высокой агрессивности (кричащих в более прерывистом ритме) и дают возможность бежать противникам, сигнализирующим о низкой агрессивности (кричащих в более монотонном ритме)».
Источник wolf_kitses
Хотя у млекопитающих (и как минимум у некоторых видов птиц) общие с нами индикаторы эмоционального состояния, их выявление — плод долгого изучения, о результатах которого надо знать, «автоматически« наблюдателем это не ловится.
Однако как кажется, здесь есть что возразить. Существует вполне обоснованная теория, почему телодвижения демонстраций, используемых в коммуникации позвоночных, выглядят так, как они выглядят. Это своего рода теория деятельности, только применительно к инстинктивному поведению животных. Она принадлежит К.Н.Благосклонову[1]: согласно ей, в движениях демонстраций «проверяется» умение особей «должным образом» сопрягать своё поведение с таковым социальных компаньонов в ключевых для выживания взаимодействиях[2].
Надо сказать, такая проверка очень важна, ибо особи вне социальных связей — это «эгоистичные индивиды», поддержание их — вещь рискованная и затратная, поэтому связи «натягиваются» (и «обслуживаются» сигналами, особенно самым продвинутым видом их — сигналами-символами) только там, где без этого никуда. Тем не менее «эгоизм» прорывается наружу, создавая разного рода «сбои» в развёртывании взаимодействия, неточности обмена демонстрациями, цена которых — дискоординация при завершающем акте, вроде копуляции (за что платят не-оплодотворением) или агрессии (за что платят ранением).
Поэтому жизненно важно максимально сопрячь действия самца и самки в ухаживании, победителя и побеждённого в агрессии, чтобы получить нужный результат за приемлемое время течения процесса — но сделать это очень и очень непросто. У животных, с их «слепыми, но мудрыми» инстинктами сие получается сильно хуже, чем координация движений танцующих. Это изобилие «ошибок» и «сбоев», к слову, показывает, насколько сигнальные системы животных, кажущиеся изощрёнными, неэффективней и и примитивней нашей.
Другая причина «срыва» взаимодействий — каждый их тип (территориальная агрессия, поиск и «тестирование» партнёров для образования пары, ухаживание в образовавшейся пары и пр.) связан со своим собственным конфликтом мотиваций, преодолеваемым как раз в обменах демонстрациями. См. табл. 10 в «От стимула к символу». Скажем, при ухаживании больших пёстрых дятлов сближение партнёров может быть сорвано выплеском агрессии одного (чаще со стороны самки) или страхом и бегством другого партнёра.
Поэтому важный неспецифический эффект демонстраций, которыми обмениваются компраньоны, состоит в нарастающем «связывании» обоих через стимуляцию подобного подобным, чтобы по ходу процесса 1) их действия синхронизировались друг относительно друга, 2) смены демонстраций у обоих участников всё больше координировались во времени; 3) само же взаимодействие продолжалось далее, с последовательным снижением вероятности остановки или «сбоя» процесса, когда социально связанные и социально зависимые компаньоны под действием вышеописанных причин рискуют, «расцепившись», вновь стать «эгоистическими индивидами». См. «Ещё про «расшифровку «языка животных», ч.1»
А специфический эффект демонстраций состоит в рефлексивном управлении процессом сближения, через передачу информации и «уколы» воздействиями, позволяющие обоим заранее скорректировать будущее поведение сообразно наиболее вероятному будущему выбору поведения у партнёра и тем дифференцировать социальные роли по ходу и после взаимодействия с наибольшей выгодой для обоих (с минимумом временных затрат и риска «сбоев» процесса.
Согласно модели К.Н. Благосклонова отбор тестирует совместное действие, венчающее сближение партнёров, когда последнее преодолело все эти «сбои» с «помехами» и пришло к специфическому результату. Демонстрации представляют эти телодвижения в ритуализованной (вычурной или гротескной) форме, чтобы подчеркнуть «умения», критически важные для «соединения» данных компаньонов в определённую «клеточку» социальной организации: брачную пару, «лестницу иерархии» в группе, основанную на агрессивном доминировании, группировку территориальных резидентов, связанных общей коммуникативной сетью и пр. (типы компаньонов, как писал ещё Лоренц, соответствуют контекстам общения, и формируют каждый свою «клеточку» социальности животных, см. табл.10).
Эти движения кажутся нам самыми «неестественными», ибо обслуживают не витальные потребности особей, что мы видим постоянно и часто, а «сцепление» их друг с другом в процессах коммуникации, воспроизводящих популяционную структуру. Это и бывает нечасто, и требует обойти исходный «эгоизм« индивидов (включая запрет на прямой телесный контакт, поддержание индивидуальной дистанции), связав их видоспецифическими связями. Отсюда и напряжение, и «неестественность» этих действий. Скажем, у поганок ухаживание всегда включает скоординированные нырки за гнездовым материалом и его передачу партнёру (или завершается им). Ритуализируется не ныряние, важное для обычного выживания, а именно передача растительной ветоши другому и скоординированность этого процесса. См. рассказ самого Константина Николаевича:
«Среди разнообразных форм поведения в период размножения можно выделить некоторые жизненно важные, т. е. такие преимущественно врожденные формы поведения, при отсутствии которых у родителей потомство не может появиться или выжить до времени самостоятельного существования. Жизненно важны обычно все черты поведения, связанные с заботой о потомстве и присущие лишь одному из родителей без возможной их взаимозамены. Очевидно, все эти формы поведения совершенствуются под влиянием естественного отбора. Каким же образом осуществляется отбор этих форм поведения? В ряде случаев он происходит не только в момент проявления жизненно важного поведения, но по крайней мере у некоторых птиц и значительно раньше, еще во время брачных игр и ритуалов.
Самец кормит самку
Содержание
Среди разного рода брачных ритуальных демонстраций птиц широко распространено кормление самки самцом (рис. 1).Н.Тинберген (1970) видит в подобном кормлении выражение подчиненности самки самцу. Р. Хайнд (1975) полагает, что
«…оно, вероятно, связано с каким-то весьма специфичным внутренним состоянием, которое к тому же отличается от состояния выпрашивающего корм птенца».
Эти рассуждения весьма неопределенны, не объясняют явления с эволюционной точки зрения.

Рис. 1. Самец лесной завирушки кормит самку. Фото Ф. Штейнбаха
В природе даже такой жизненно необходимый рефлекс, как кормление птенцов, может быть выражен у отдельных особей птиц в резко различной степени. Это можно видеть, сравнивая, например, число прилетов с кормом к птенцам самцов и самок мухоловок-пеструшек в разных гнездах. Картина получается очень пестрой. Доля участия самцов в кормлении птенцов старшего возраста в разных гнездах может изменяться от 30 до 70% (у этих птиц, как и у многих других, доля участия самцов в кормлении птенцов увеличивается с их возрастом). Еще резче разница у пеночек-теньковок. В большинстве пар теньковок самец не кормит птенцов вообще, но некоторые самцы изредка все же приносят им корм. Иногда, однако, попадаются самцы, которые кормят птенцов наравне с самкой.
У подавляющего большинства птиц, как и в двух последних примерах, один партнер, как правило, компенсирует недостаток в поведении другого. Даже в случае гибели одного из партнеров второй способен самостоятельно выкормить птенцов. Но у многих видов функции родителей строго разграничены, и тогда только норма жизненно важных элементов поведения каждого из них обеспечивает сохранение потомства.
Большие синицы, как правило, делают два выводка за лето по 8—12 (и до 18) птенцов в каждом. Кладка, насиживание, выкармливание птенцов в полтора раза длительнее, чем у некоторых других птиц того же размера (Благосклонов, 1969в). Чтобы успеть выкормить второй выводок, самки начинают кладки во время кормления птенцов первого выводка (в сутки откладывается одно яйцо), и ко времени их вылета из гнезда синица уже крепко сидит на второй кладке. Забота о слетках полностью ложится на самца, и, если у него будет кормовая активность ниже нормы, выводок погибнет.
У больших синиц обычно сначала образуется пара, затем самец начинает поиск места для гнездования, самочка лишь следует за ним. Ее роль иная: время от времени она принимает позу просящего корм птенца и подражает его голосу. Самец немедленно реагирует на это поиском насекомых и кормит самку, она замолкает, через некоторое время все повторяется снова. Замечательно, что далеко не всегда самец кормит самку, иногда он, не найдя ничего съедобного, подлетает к ней и «делает вид», что кормит ее, имитируя движения кормления. Очевидно, что в данной игре важен не корм для самки, а реакция самца на крик птенцов — это должно быть кормление. Именно эта важнейшая для сохранения потомства черта поведения самца как бы проверяется при составлении пары, и, если у самца не будет адекватной реакции на крик слетка, пара, очевидно, не составляется. Может возникнуть вопрос, почему цикл иногда не полон. В ритуале может отсутствовать корм. Нам представляется это понятным. Ведь проверяется не способность птицы находить корм, она очевидна, если самец выжил в течение по крайней мере года, а именно способность по сигналу выпрашивания корма отдавать его.
Для большой синицы в норме характерно кормление самцом насиживающей самки. Это считалось элементом стереотипа видового поведения. Однако в гнезде большой синицы с ненормально малой кладкой в 5 яиц самец обычно уже не кормит в период насиживания самку — у нее остается достаточно времени для самостоятельной кормежки (Звенигородская биологическая станция Московского университета, 1974). Следовательно, число яиц в кладке определяет поведение самки, а она стимулирует или не стимулирует самца к кормлению. Поскольку у синиц, особенно у большой, в норме много яиц в кладке, в норме же и кормление самцом, т. е. должна быть развита и способность самца к кормлению. Тем более рационально включить в брачные ритуалы демонстрацию самцом способности кормить не только птенцов, но и самку. Выпрашивание пищи у самца самкой в брачный период имеет место и у других синиц, но только у большой оно выражено очень резко.
В начале брачного периода самец серебристой чайки (Larus argentatus) часто кормит самку. Это действие стимулируется тем, что самка выпрашивает у самца корм совершенно таким же криком, каким требуют корм у родителей подрастающие птенцы, с той только разницей, что у взрослой птицы он получается звонким, а не резким и писклявым, как у малышей (Тинберген, 1975). Далее о роли самца и самки в прокормлении птенцов Н. Тинберген пишет, что самка большую часть времени находится у гнезда, а добычу носит преимущественно самец.

Рис. 2. Серебристые чайки. Фото Ю. Артюхина
Ритуал ухаживания самца за самкой у серебристых чаек сложен (рис. 2). В него включены ритуальный сбор материала для гнезда и демонстрация агрессивности по отношению к самцам, находящимся поблизости. Не есть ли и это демонстрация самцом умений, нужных в семейной жизни: строить гнездо, защищать гнездовую территорию? Стимулирующее воздействие подобных демонстраций становится понятным. Самец как бы заставляет самку воспринимать зрительно элементы будущей семейной жизни, и если он входит в роль заботливого отца семейства, то и самка активно вовлекается в свою роль матери у гнезда: просит корм у самца для еще не существующих птенцов. Ритуальное кормление свойственно всем видам чаек, Н. Тинберген (1970) подробно описывает его у моевки (рис. 3) и озерной чайки. Это естественно, так как, по-видимому, у всех чаек самец — основной кормилец семьи.
У полярных крачек (Sterna paradisea), по данным Н. В. Ан- зигитовой (устн. сообщ.), наблюдается тот же ритуал выпрашивания корма самкой у самца. И здесь самка движениями и голосом подражает птенцу. Можно предполагать особую роль самцов этого вида в выкармливании потомства, что подтверждается прямыми наблюдениями. Самец приносит корм птенцам в 5—10 раз чаще, чем самка, которая часами не отлучается от гнезда. Самки иногда применяют этот прием выпрашивания корма уже во время выкармливания птенцов, когда самец приносит пищу к гнезду. Реакции самца могут быть различны. Он передает рыбку самке, но может отдавать ее и птенцу. Если рыбку получает самка, то опять-таки либо она отдает корм птенцу, либо съедает его сама. Вероятнее всего, что самка таким выпрашиванием корма как бы стимулирует деятельность самца по ловле рыбы, если он выполняет эту свою обязанность недостаточно активно. Ведь так поступают только некоторые самки.

Рис. 3. Обыкновенные моевки на гнездах. Фото С. Миронова
У грачей (Corvus frugilegus) также существует брачный ритуал подражания кормлению птенца самцом (Blume, 1973). М. А. Воинственский (устн. сообщ.) отмечает, что самки грачей постоянно находятся у гнезда, корм птенцам доставляет самец.
У перечисленных видов птиц инициатива ритуального кормления исходит обычно от самки. Она просит корм, и самец должен реагировать на это кормление. Однако бывает и иначе. Свен Нильсетер (1974), шведский фотограф и путешественник, описывает «сватовство» у дымчатых (глупых) крачек на острове Буш-Ки в цепи Флорида-Кис:
«Инициатива к более близкому знакомству исходила от самца, который опускался рядом с понравившейся ему самкой, поворачивал к ней голову, широко раскрывал клюв, кивал и кланялся. Если его предложение милостиво принималось, самка засовывала свой острый клюв глубоко в глотку кавалера. Таким образом, влюбленные крачки делятся супом из полупереваренной пищи. Потом они несколько раз почистили друг о друга клювы и закивали белыми головками».
И здесь, без сомнения, самец демонстрировал свою способность быть заботливым отцом семейства.

Обыкновенные глупые крачки Anous stolidus
Мы привели несколько примеров из жизни птиц, за которыми проведены достаточно детальные наблюдения. Вероятно, ритуал кормления свойствен и многим другим видам. Самцы этих видов в большей мере, чем самки, заботятся о прокормлении птенцов. Это, возможно, связано с их колониальностью. И мы напрасно стали бы искать подобную форму ритуального поведения среди птиц полигамов, которым она вообще не известна, или даже у многих моногамов, которым она не нужна, так как самец не принимает или в малой мере принимает участие в выкармливании птенцов.
Брачные ритуалы у мухоловок — пеструшек
Известны случаи, когда мы можем увидеть функциональность брачного поведения в процессе самих демонстраций. Это, например, ритуальное поведение, «сватовство» при встрече самца и самки мухоловок-пеструшек (Благосклонов, 1975в). Самцы этих птиц прилетают с юга на неделю раньше самок, находят место для гнезда и начинают петь. Привлекается самка. Ритуал составления пары довольно сложен. Самка приближается к синичнику, самец,
цыкая, влетает в леток и «ылетает, как бы приглашая самку осмотреть синичник (рис. 4). Это негромкое цыканье мы слышали у пеструшек только в такой ситуации. Самка не приближается к синичнику, наблюдает издали за самцом, а затем улетает. Самец не следует за ней, он садится рядом с гнездовьем, например на крышу синичника, и поет в быстром темпе. Только через несколько минут самка возвращается, осматривает синичник, и, если он хорош, пара составляется. Таким образом, самка выбирает гнездовье, и отбор самцов идет по их способности находить хорошее место для гнезда и степени «привязанности» к нему.

Рис. 4. Некоторые позы демонстративного поведения птиц в брачный период 1 — Мухоловка-пеструшка: а) самец, вцепившись в леток гнездовья, показывает самке свое дупло; б) самка подлетает к синичнику для осмотра дупла; в) самка просит у самца корм; 2 — самец горихвостки разными способами показывает самке свое гнездовье (Blume, 1973)
В течение нескольких дней я наблюдал за интенсивно поющим, явно холостым самцом мухоловки-пеструшки в то время, когда в нескольких гнездах этого вида уже появились птенцы. Время от времени самец подлетал к дуплу в развилке сосны, но никогда не влезал в него. При обследовании дупла оказалось, что он и не мог этого сделать — оно было таким маленьким, что в нем едва помещался мой палец. Вероятно, к самцу подлетали самки, но ни одна из них не могла составить ему пару, так как гнездиться было негде. То есть самец должен остаться холостым, если вся процедура образования пары не может быть проведена, а его психическая неполноценность элиминирует в процессе отбора.
«Поцелуй» птиц
«Целуются ли птицы»? — задает риторический вопрос Р. Шовен (1965) и заключает, что «в это легко поверить». Среди целующихся пар он называет кайр, чистиков, чомг, буревестников, гра чей, олуш (все колониальные). Птицы «целуются» во время ухаживания самца за самкой (рис. 5), при встречах партнеров, во время насиживания кладки, но, по-видимому, не позднее этого периода размножения.
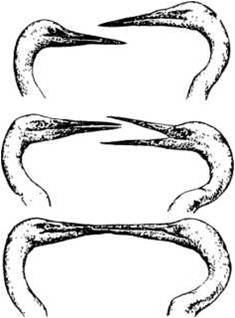
Рнс. 5. Белые цапли обмениваются церемониальным «поцелуем», который, по Н. Тинбергену (1969), устраняет «недоверие» между самцом и самкой. Автор этой книги считает, что «поцелуй» — это только ритуал передачи корма самке самцом
Партнеры составившейся пары мухоловки-пеструшки «целуются» так: они медленно, маленькими шажками приближаются по ветке один к другому с вытянутыми шеями, наклоняют головы в разные стороны и соприкасаются клювами, которые при этом приоткрыты и совершают быстрые днижения челюстями. Самец активнее, и его клюв дольше находится в более широко открытом клюве самки. Иногда птицы изменяют наклон головы на противоположный.
Создается впечатление, что птицы именно целуются. Так обычно и называют этот акт у голубей, попугаев и других птиц, которые совершают его обязательно и часто при составлении пары (возможно, что именно из-за этого возникло неверное представление, что голуби — пример нежной любви и супружеской верности). Однако, если наблюдая «целующихся птиц» представить себе, что самец передает самке из клюва в клюв какой-то очень маленький предмет, впечатление, что птицы «целуются», исчезает. На самом деле нет никакого и самого маленького предмета. Есть только ритуал передачи корма самке самцом.
Способность самца кормить самку — запрограммированный, но не всегда проявляющийся рефлекс. Пеструшки «целуются», когда насиживающая самка, сойдя с гнезда, встречается около него с самцом. Самец как бы напоминает самке о своей способности к кормлению. Если нет нужды в нем, самка отвечает самцу «поцелуем», т. е. тоже совершает ритуал получения корма. Но при большой кладке (а мы искусственно увеличивали ее до 12 яиц) самка не может отлучиться от гнезда при насиживании (см. гл. 5). В этом случае она при встрече с самцом у гнезда подобно птенцу трясет крыльями, прося корм. Это и есть сигнал к тому, что от ритуала самцу нужно переходить к обеспечению самки реальным кормом. В наших экспериментах самцы кормили самок до 6,6 раз в час (при кладке 12 яиц).
Ритуальные демонстрации поганок
У некоторых птиц брачные игры, ритуалы, демонстрации выполняют оба партнера, как бы испытывая друг друга («взаимное ухаживание», по терминологии Тинбергена). Среди таких птиц особенно типично брачное поведение поганок. Кульминационный момент ритуальной игры двух поганок (известнее других брачное поведение чомг) состоит в том, что после одинаковых и синхронных движений обе птицы ныряют и выныривают с пучками растений в клюве (рис. 6).
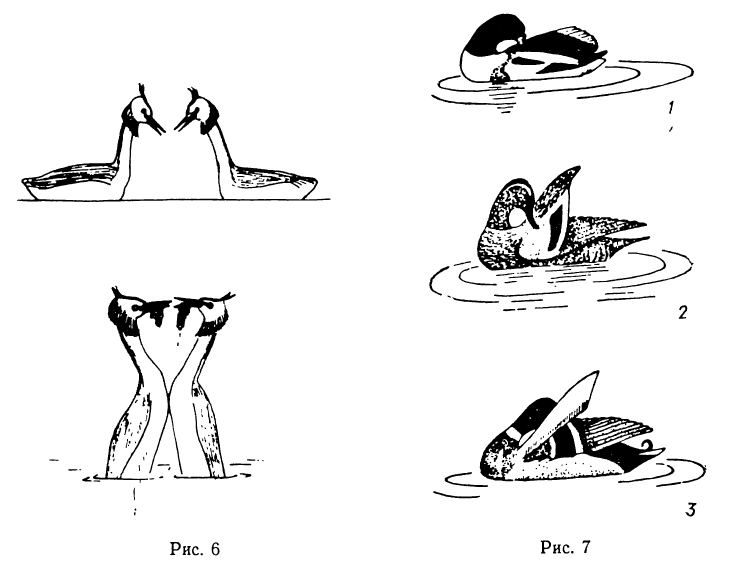
Рис. 6. Позы брачного поведения чомг. Обе птицы, выныривая, демонстрируют вытащенные из воды пучки травы Рис. 7. Позы демонстративного поведения у селезней речных уток, имеющих на крыльях «зеркальце»: 1 — пеганка энергично теребит основание крыла; 2 — чирок-трескунок легкими движениями клюва демонстрирует сине-голубое «зеркальце»; 3 — селезень кряквы приподнимает крылья, демонстрируя яркое «зеркальце» (Тинберген, 1969)
Пара красношейных поганок с травой в клювах проделывает сложный ритуал, напоминающий танец (Storer, 1969). Именно демонстрация пучка травы бывает заключительной в ритуале поганок. В сложных движениях партнеры допускают ошибки, неточно копируют движения друг друга, что оставляется без внимания другим партнером. Но пучок травы должен быть обязательно. Какую же связь с жизненно важным поведением мы можем предположить в этом случае? Гнездо поганок громоздко, лежит между стеблями растений на дне водоема на глубине обычно до метра. В процессе насиживания оно быстро уплотняется, оседает. Поганки часто сменяют друг друга при насиживании, и почти каждый раз птица приносит в клюве пучок травы, который укладывает вокруг себя.
У черношейной поганки, например, смена птиц на гнезде происходит каждые 2—4 ч, и всегда с достройкой гнезда сменяющим партнером (Бородулина, устн. сообщ.; Borodulina, 1970). Если птица не будет достраивать гнездо, первоначальный лоток гнезда за время насиживания и утаптывания птенцами постепенно опустится и кладка непременно окажется в воде. Нам известно также, что две чомги при брачных играх всегда демонстрируют друг другу собранные со дна пучки травы, именно такие, какие идут на достройку гнезда. Известно (по Тинбергену) и следующее: если один из партнеров этого не сделает, то пара не составится. Доказательство связи двух явлений непрямое, но достаточно убедительное. Понятна и однородность демонстраций двух партнеров: функция достраивания гнезда в равной мере важна для обоих.Н. Тинберген считает, что многие ритуальные позы уток произошли от движения комфорта — чистки оперения. Нас интересует другое: почему именно эти, а не какие-либо другие движения стали основой брачного ритуала. «Вопрос о специфике «брачных церемоний» у различных видов гусей и уток остается все еще недостаточно выясненным, но несомненно, что поведение уток в период размножения довольно однообразно, — замечает Г. П. Дементьев (1940), — последнее справедливо и совершенно естественно, так как главная задача «брачных церемоний» уток одна и та же: установить видовую принадлежность партнера». Вследствие очень сходных морфологических адаптаций к обитанию в водной среде при сохранении способности к полету внешние формы уток очень схожи. Различение ‘видов построено на различиях в окраске, так как возможность изменить типичный профиль плавающей утки весьма ограничена.
Встреча самца и самки начинается с того, что утки по весенней воде выплывают на открытые плёсы, а селезни совершают облет вдоль берегов и подсаживаются на воду, заметив самку. На этом основана и весенняя охота на селезней с подсадными утками или чучелами. Определение видовой принадлежности встречающихся особей происходит позднее, когда расстояния между ними невелики. Сам ритуал опознавания вида у уток может включать до десятка различных положений, движений, демонстраций (Johnsgard, 1973). Однако есть ключевая демонстрация, характерная для каждого вида. К. Лоренц считает, что у речных уток (т. е. уток, имеющих на крыле зеркальце) она одинакова для всех видов. Самец становится боком к самке, слегка распускает крыло с обращенной к ней стороны, что увеличивает размеры зеркальца, и, изгибая шею, клювом многократно касается или почти касается зеркальца (рис. 7). Ритуальность всех движений направлена на большую демонстративность, броскость видовых знаков различия: раскрывание крыла именно в той части, где находится зеркальце; показывающие движения головой.
Принципиально иные опознавательные видовые признаки у ряда других уток. Опознавательный признак шилохвости — остроконечный хвост, именно его демонстрирует селезень этого вида при встрече с самкой (например, погрузившись наполовину под воду и поставив хвост прямо вверх; рис. 8,9). «У уток, которые имеют яркую опознавательную окраску на груди, шее, голове, наиболее типичное движение — закидывание головы на спину клювом вверх или вытягивание шеи по поверхности воды» (Johnsgard, 1973). Только такого ‘рода опознавательная окраска свойственна нырковым уткам. Мы объясняем это тем, что плотность нырков выше, чем у речных уток, и при их более глубоком погружении большая часть крыла оказывается под водой. Всегда над водой у плавающей нырковой утки голова. Именно она у них ярко окрашена, имеет пятна, хохол. В данном случае мы можем ждать от самца «игры головой», как и отмечено у нырковых уток.
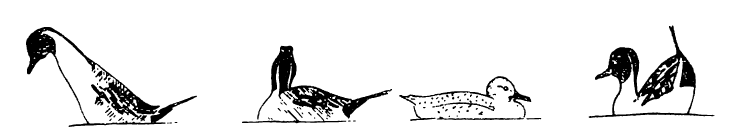
Рис. 8. Поза демонстративного брачного поведения у шилохвости
У гоголя во время ухаживания за самкой голова все время в движении (Blume, 1973), самец располагается боком к самке: именно сбоку непропорционально большой темной головы селезня расположено яркое белое пятно (рис. 10). Вот запись наблюдений за двумя самцами гоголя, токовавшими вокруг своих самочек на пруду Новодевичьего монастыря в Москве 1 мая 1977 г. Самец поворачивается боком к самке, вытягивает шею и погружает ее в воду почти всю, сам погружается при этом так, что над водой остаются небольшая часть спины и вся голова, низ клюва касается поверхности воды. Такое погружение осуществляется, как показано на примере поганок, не только «выдохом» части воздуха из воздушных мешков, но и прижиманием перьев к телу для уменьшения воздушной прослойки (Бородулина, 1966). В вытянутом полупогруженном положении птица находится иногда несколько минут. При этом самец либо медленно передвигается головой вперед, чуть боком под углом к оси тела, либо остается на месте. Погружение может быть большим или меньшим, но положение головы у самой поверхности относительно неизменно. Время от времени птица откидывает голову назад, на спину, зобом вверх. Это длится всего несколько секунд. Очевидно, при этом важно само движение. После этого самец принимает обычное положение тела плывущего нырка, и игра начинается через некоторый промежуток времени или сразу в том же порядке. При нырянии самки самец сейчас же ныряет вслед за ней.

Итак, в брачных демонстрациях уток, не имеющих зеркальца на крыле, нет ничего хоть сколько-нибудь напоминающего движения комфорта — чистку клювом пера. И у всех видов демонстрируются наиболее яркие участки оперения. Все направлено к тому, чтобы точно определить вид партнера. И все же именно среди уток очень часты случаи межвидовой и даже межродовой гибридизации. Более всего известно гибридов кряковой утки (Anas platyrhynchos) с чирком-свистунком (А. сгесса), с шилохвостью {A. acuta), со свиязью (Л. penelope) и даже с красноносым нырком (Netta rufina). Встречаются также гибриды чирка-свистунка со свиязью и шилохвостью; нырков красноголового (Aythya ferina) и белоглазого (А. пугаса) и другие (Дементьев, 1939).

Рис. 10. Брачные демонстрации у нырковой утки — гоголя:
1 — селезень гоголя. На большой черной голове белое опознавательное пятно. Фото А. Пудова; 2 — токующие гоголи. Фото Ю. Шибаева; 3 — схема брачных демонстративных движений селезня гоголя. Во время ухаживания за самкой селезень движениями большой черной головы демонстрирует на ней белое опознавательное пятно (Blume, 1973)
При обычности гибридов между разными видами уток в природе гибриды между видами гусей чрезвычайно редки (Дементьев, 1940) [сейчас известно, что при подходящихся условиях они появляются не реже утиных, скажем, серых гусей и канадских казарок в Европе]. Это объясняется, по всей вероятности, не тем обстоятельством, что морфологические различия в группе гусей более значительны, но скорее всего особенностями биологии, связанными с размножением. Для уток разных видов характерны большее совпадение циклов деятельности, меньшая привязанность к гнездовой территории, отсутствие постоянных гнездовых пар и участия селезней в заботе о потомстве. У гусей (при отсутствии резкого полового диморфизма) самцы, как известно, спариваются лишь с одной самкой и принимают участие в заботе о выводке. Весьма существенна большая стайность гусей, особенно при перелетах, так как гуси спариваются еще до прилета на места гнездования. В условиях стаи нахождение партнера того же вида не может встретить затруднений. Из этих соображений, высказанных Г. П. Дементьевым, мы можем сделать вывод, что для гусей и не нужен ритуал опознавания вида, но он совершенно необходим для уток.
Брачные демонстрации самца трёхиглой колюшки
Ритуальное поведение ярко выражено у птиц, но не им одним оно присуще. Н. Тинберген (1974; Tinbergen, 1976) подробно описал брачные демонстрации трехиглой колюшки:
«У самца трехиглой колюшки с сильно стимулированным сексуальным побуждением мы наблюдаем элементы родительского поведения, хорошо нам известные по более позднему этапу цикла размножения этой рыбки, — движения «обмахивания», которые обеспечивают аэрацию икры. Обмахивание — столь же обязательный компонент родительского поведения колюшки, как и насиживание у птиц. Когда на территории колюшки появляется самка, самец начинает интенсивно ухаживать за ней, но время от времени возвращается к гнезду и производит обмахивание, хотя там нет икры, которая нуждалась бы в аэрации». Во всем этом Н. Тинберген видел только смещенное, необоснованное поведение, «странное сочетание двух форм поведения».
Но ведь это движение — важнейшее, определяющее нормальное развитие потомства (Драпкин, 1954). Если этот поведенческий признак выражен слабо или совсем не выражен у данного самца, он, очевидно, не сможет его демонстрировать перед самкой. Поскольку брачный ритуал не соблюден, самочка не получит побуждения, достаточного для спаривания, и икра не будет отложена. Это целесообразно, так как при спаривании с самцом, который «не умеет» аэрировать гнездо, икра, вероятно, погибает. Положительная реакция самки должна быть прежде всего на «проверяемые» ритуалом способности самца».
P.S. Я добавлю: сравнительная этология, уже в силу «морфологического подхода к поведению», определяет единицы последнего (демонстрации) структурно, а не функционально: формой каждой из единиц, выделенных наблюдателем в предположении, что они реально существуют в «мире» (die Umwelt) самих животных и существенны (т.е. также выделяются) ими самими. О чём см. «Три антропоморфизма».
Что для животных, что для наблюдателя существенны 3 аспекта формы: её специфичность, выделенность облика или образа, репрезентируемого партнёру, и типологическая определённость формы. Степень развитости всех 3-х аспектов говорит о реальности соответствующих демонстраций в видовом репертуаре, показывая, что их формы – действительно отдельности, дискретизируемые объективно (самими особями[3]), а не просто удобные наблюдателю единицы классификации.
Соответственно, каждая из демонстраций — территориальной, брачной или иной мотивации – суть устойчивая комбинация выразительных телодвижений, происходящих из разных источников (обычно повседневной активности – действий кормления, чистки и т.д.), но жёстко скоррелированных по времени исполнения. См. как это проявляется в единицах ритуализованной угрозы БПД (источники – «От стимула к символу»). Вот репертуар территориальной агрессии большого пёстрого дятла Dendrocopos major: демонстрации как специфические корреляции выразительных движений, инвариантные относительно изменений контекста.
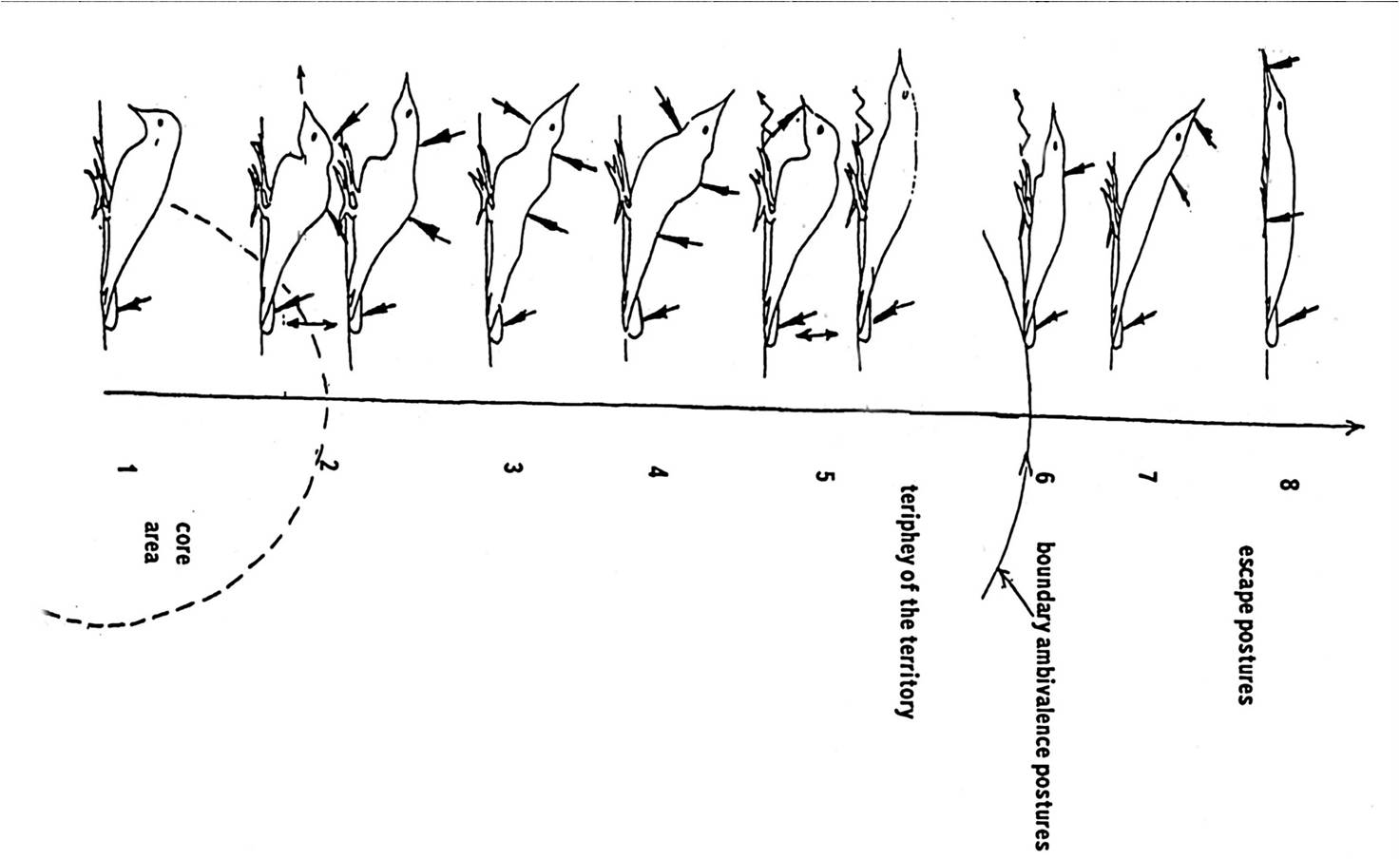
А. Демонстрации территориальной агрессии №№1-8, выделенные как устойчивые комбинации выразительных движений, исходно моторно не связанных друг с другом (стрелки). Прочие комбинации движений, постоянно возникающие на волне агрессивного возбуждения животного, не обладают ни высокой скоррелированностью предъявления разнородных движений в составе комбинации (Б), ни сравнимой устойчивостью корреляций относительно изменений контекста (В). Обозначения. Демонстрации «расставлены» на градиенте от центра к границе территории в соответствии с тем, где данные демонстрации чаще всего «открывают» конфликт: сигналы угрозы №№1-4 – в центре собственной территории, амбивалентные демонстрации №№5-6 – в районе границы, позы подчинения №№5-6 – на чужой территории.
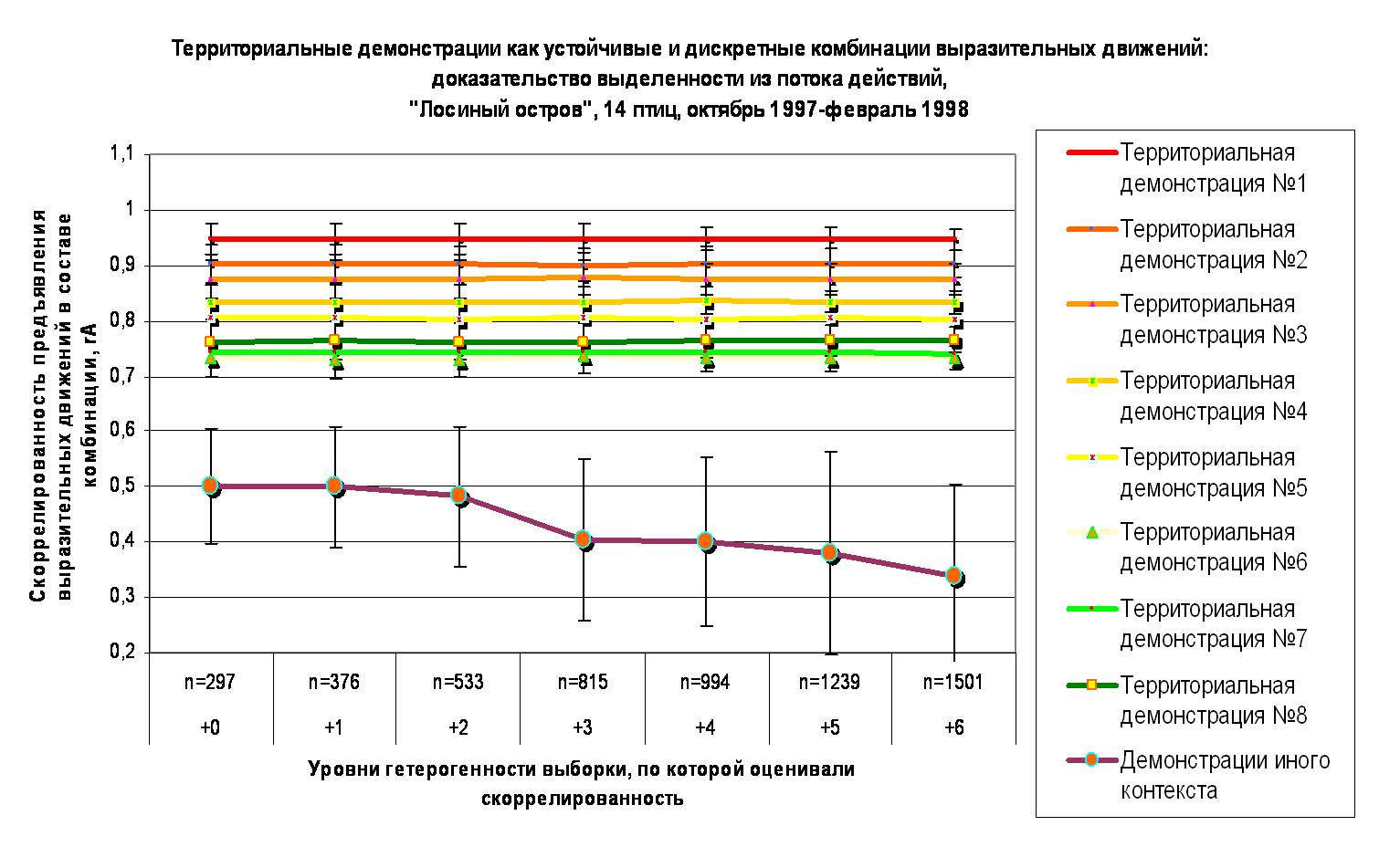
Б. Скоррелированность предъявления ЭДА как устойчивая характеристика демонстраций, отделяющая их от экспрессивных реакций животного, «демонстративных», но не обладающих устойчивой формой. Ось абсцисс — уровни гетерогенности выборки: 0. Основная выборка – 14 особей, наблюдения с октября 1997 по февраль 1998 года, +1 – + Разные уровни возбуждения особей, +2 — +Изменения социального статуса тех же индивидов, +3 — +Новые взаимодействия особей в другие недели и месяцы наблюдений, +4 — +Разные годы жизни особей в данной группировке, отличающиеся урожаем семян и связанной с ним плотностью, +5 — +Птицы из разных группировок, отличающихся степенью постоянства данного «узла» в сети поселений вида, +6 – +Птицы разных географических популяций и/или подвидов. Ось ординат – скоррелированность предъявления ЭДА в составе комбинации, коэффициент ассоциации Пирсона (rA).
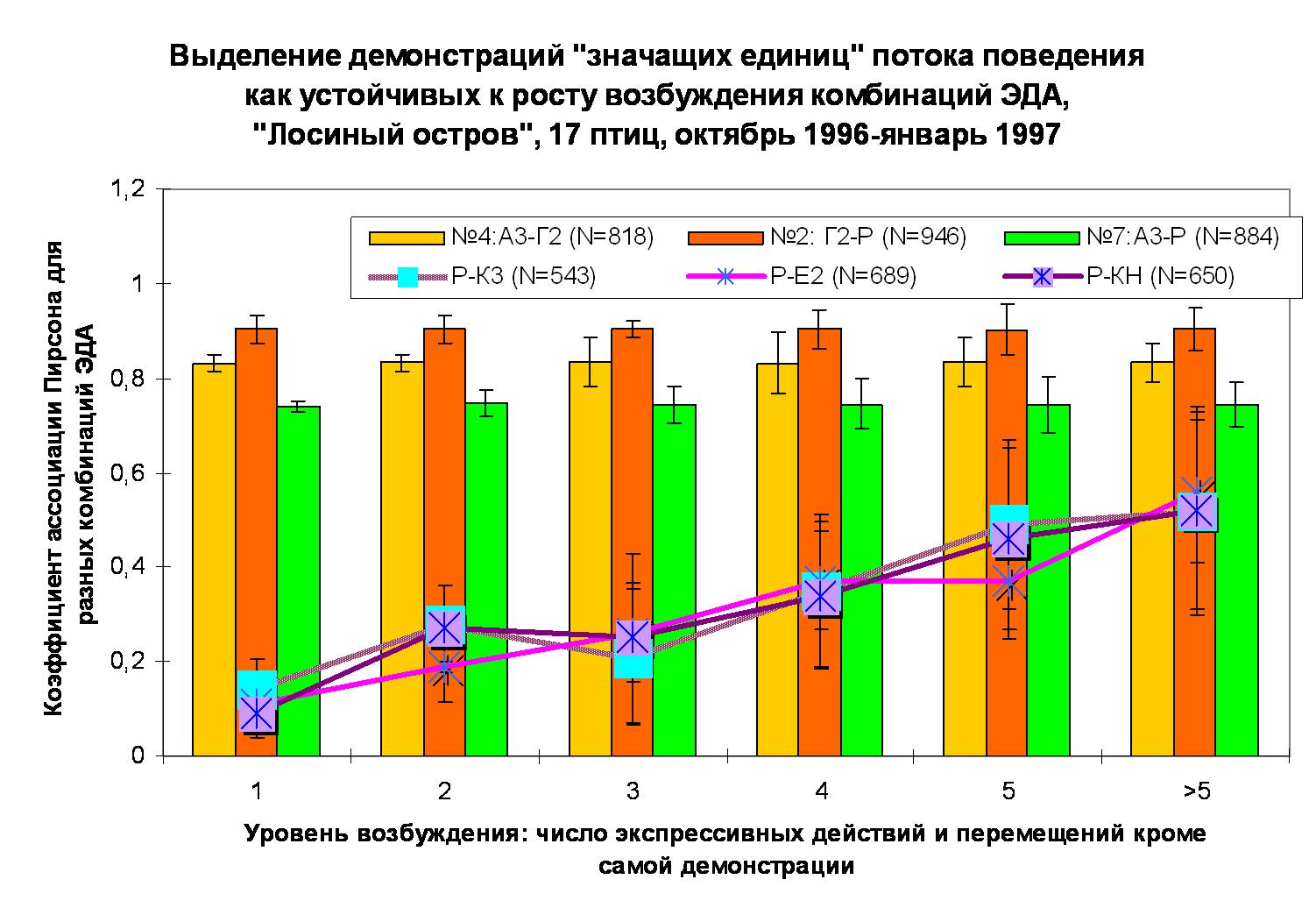
В. Устойчивость корреляций ЭДА, образующих территориальные демонстрации №№1-8, к изменениям контекста, в противовес неустойчивости демонстраций иного контекста и экспрессивных реакций. Ось абсцисс – уровень возбуждения животного, мера напряжённости ситуации взаимодействия, ось ординат – скоррелированность предъявления движений в составе комбинации, коэффициент ассоциации Пирсона (rA).
Примечание. Высокая скоррелированность предъявления движений, моторно не связанных друг с другом до объединения их в общую комбинацию, обеспечивает специфический образ последней, позволяет с высокой точностью распознавать его в процессе взаимодействия и дифференцировать от образов других демонстраций. Устойчивость корреляций к изменениям контекста обеспечивает инвариантность воспроизведения формы сигнала разными особями, в разном мотивационном состоянии, в противодействии разным противникам и пр.
Поэтому каждая демонстрация
а) выглядит неестественной, её движения не выполнить ловко, почему её исполнение рискованно тем более, чем жёстче сопряжены разнородные телодвижения во времени (т.е. чем одновременнее они выполняются), см. «Ещё про знаки и символы в коммуникации животных»;
б) все элементарные телодвижения из состава комбинаций на равных вносят вклад в её облик, отчего последний уникален, специфичен и произволен, т.е. только по нему не «читаются» «значения», контекст появления или последствия исполнения демонстрации, какими бы они ни были. В т.ч. «не предсказывается» мотивация, могущая проявиться в отдельных движениях, как агрессия в вертикальной постановке тела и «распушении головы» воронов (реально же, как показывает Благосклонов, проявляется установившееся отношение доминирование одного и подчинения другого самца в стае). См. раздел про «двойное членение» и произвольность знака в «От стимула к символу»
Модель появления ритуализированных демонстраций К.Н.Благосклонова предсказывает, что если k демонстраций образованы комбинациями из n1, n2, n3… nk элементарных телодвижений, то одно из них (именно то, о котором идёт речь в модели, отражающее способность партнёров устойчиво «сцепляться» в данном взаимодействии, выполняя скоординированно те совместные действия что критически важны для биологически адекватного результата всего общения) будет общим для всех k единиц.
Причём общим оказывается то телодвижение, которое отражает способность партнёров «сцепляться» устойчиво в данном взаимодействии, выполняя скоординированно те совместные действия, что критически важны доля биологически адекватного результата всего общения. В случае демонстраций территориальной агрессии больших пёстрых дятлов на рис. выше это развёртывание крайних рулевых, на которые птица не опирается, белый фон которых делает этот акт крайне заметным на фоне темного хвоста.
«Отражает» в очень разном диапазоне, от «прямо показывает» до «символически маркирует», т. е. может пробегать весь диапазон ритуализации, и опыты К.Н. Благосклонова показывают, что «бег» управляется экстремальностью среды, скажем трудностью с пищей и, добавлю, социальным стрессом, скажем, напряжённостью конкуренции за территории у больших пёстрых дятлов. Т.е. именно той дихотомией состояний экологической vs социальной среды, градиент которой способствует дифференциации альтернативных стратегий внутри популяции.
Соответственно, эти выделенные телодвижения обязательно входят во все демонстрации «ряда» (каждый из которых характеризуется типом «конфликта интересов», разрешение которого определяет тип компаньонов и организацию взаимодействий в данном контексте (в т.ч. в пространстве, скажем участка, или во времени — по числу дней от начала весеннего возбуждения). Часто он также связан с определённым внутренним состоянием участников последних, т. е. со специфической мотивацией. Иными словами, часто случается устойчивое соответствие демонстрации, мотивации и эффекта (хотя временами и нарушается), соответствие демонстрации и проблемной ситуации взаимодействия обязательно.
Кроме выдвижения рулевых, таковы таковы движения закатывания глазного яблока, связанные с ритуализированной угрозой у лорикетов р.Trichoglossus, «поцелуй» и «передача гнездового материала», разобранные самим К.Н.Благосклоновым. Будучи лишь одним из телодвижений в комбинации, формирующей единицу поведения, «выделенные» телодвижения не влияют на произвольность знака, относящуюся к форме демонстраций как целого, но показывают биологическое значение взаимодействий в тех контекстах, которые обслуживаются наборами из k демонстраций.
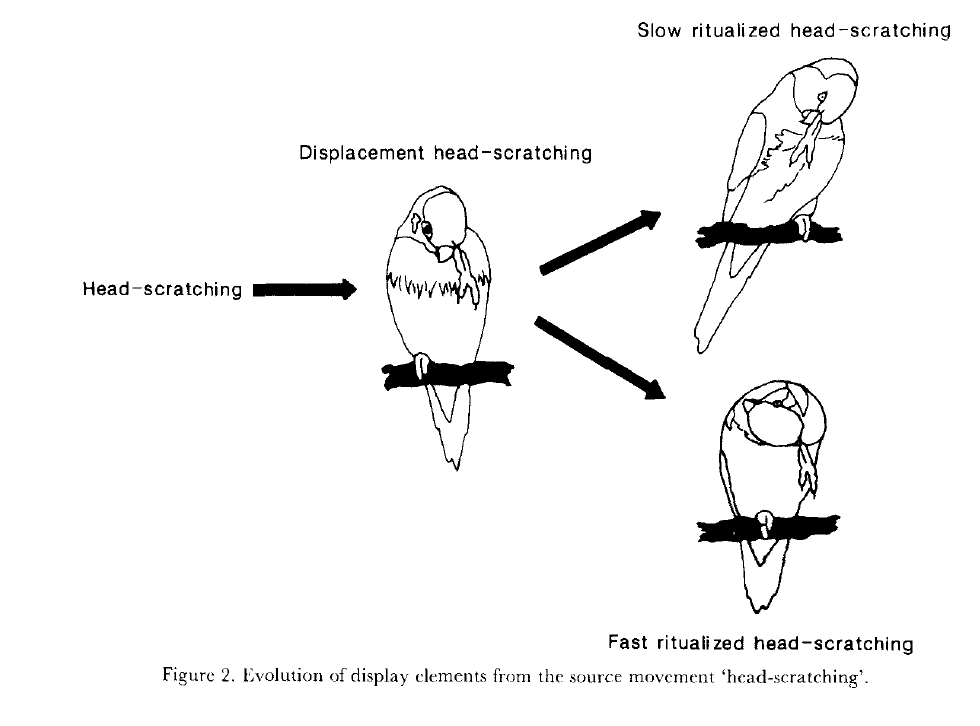
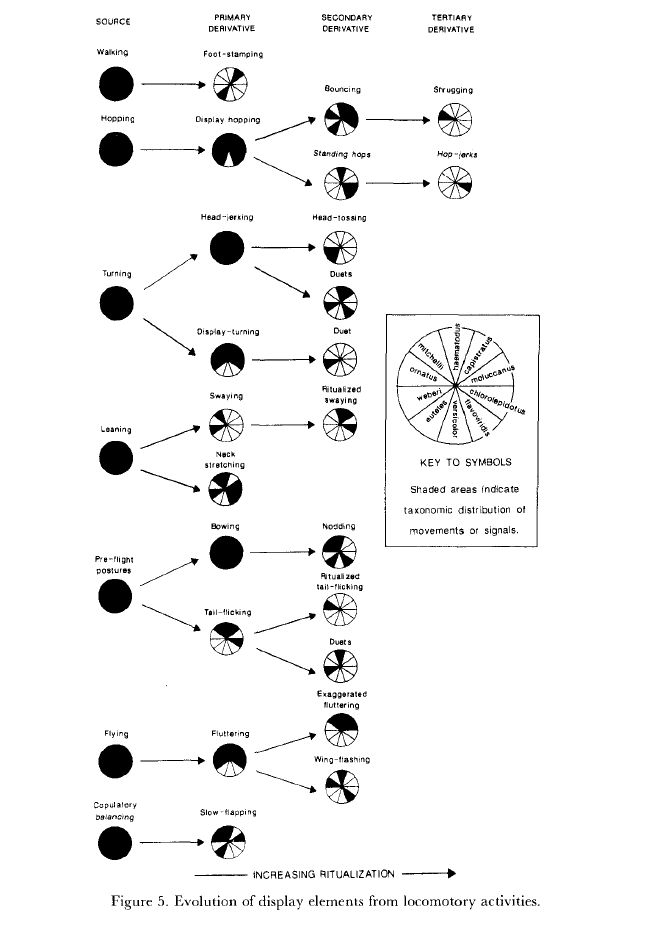
Fig.2-5. Общее направление ритуализации элементарных телодвижений в агонистических демонстрациях лорикетов. Источник: Serpell, 1989
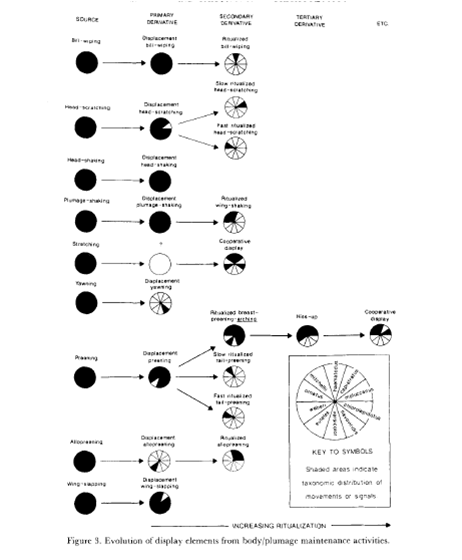
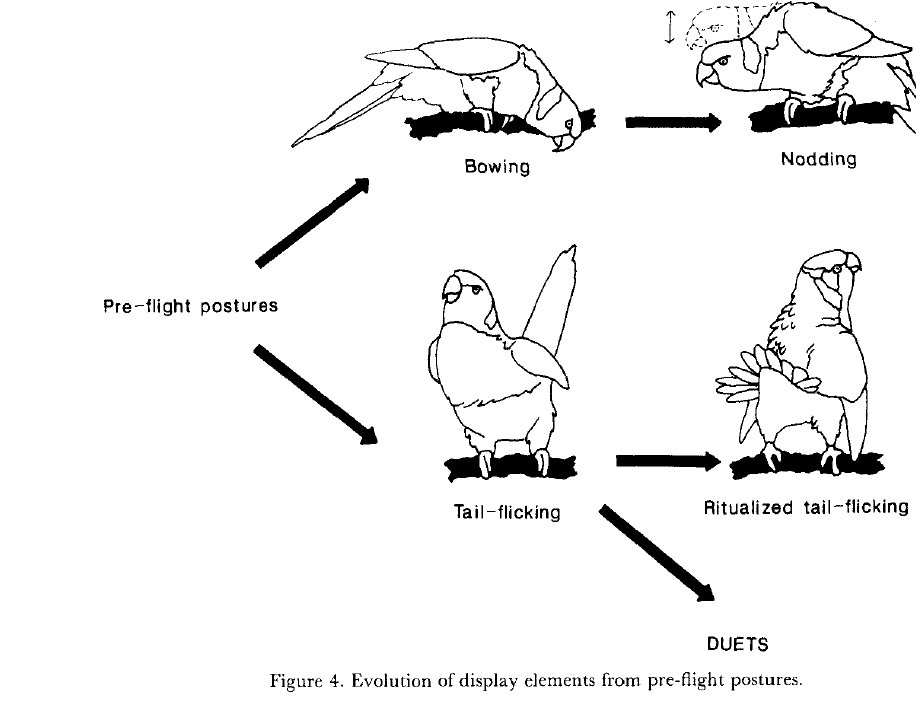
Fig.2-5. Общее направление ритуализации элементарных телодвижений в агонистических демонстрациях лорикетов. Источник: Serpell, 1989
Т.е. между классической (тинбергеновской) ритуализацией и «благосклоновской» — своего рода разделение труда. Вторая начинает дело, ритуализируя движения, обеспечивающие эффективную «связь» компаньонов во взаимодействиях, ключевых для данного вида, превращает прямое действие в пантомиму, чем даёт предпосылки использования его не только как действия, но также барьера для непосредственного сближения участников, важного для снятия стресса, и скоординированности их действий в адрес друг друга, вроде клевков и копуляций. Первая «продолжает работу» на другом уровне – формы демонстрации в целом, а не отдельного телодвижения, увеличивая сопряжённость исполнения всех движений координации. Так форма каждой демонстрации из k фигурирующих в данном контексте делается уникальной, неповторимой, а «пантомима» обращается в знак.
А сопряжение «тинбергеновской» концепции с «благосклоновской» разумно изменяет представления о характере отбора на ритуализацию. Классические этологи видели его движущим – более «вычурные» и «неестественные» телодвижения, более экспрессивные их комбинации «лучше». На деле же этот отбор стабилизрующий – он поддерживает не более ритуализованные, а «типичные» формы демонстраций, устойчиво воспроизводимые отправителем и устойчиво распознаваемые реципиентам вопреки множеству «помех», созданных конкурентным характером взаимодействия обоих и растущих по ходу взаимодействия. И «ритуализация по Благосклонову» объясняет, почему так получается – отбор на большую «сцепленность» партнёров в ключевом акте территориального или брачного взаимодействия воленс-ноленс типизирует все прочие демонстрации, обмен которыми предваряет появление этого акта и устойчиво ведёт к нему, несмотря на «помехи».
Примечания
[1]Описана в статье : Благосклонов К.Н., 1988. О биологическом значении брачного демонстративного поведения птиц // Журн. общей биологии. Т.49. №3. С.409-417: и в книге 1991 г. «Гнездование и привлечение птиц в сады и парки» (М.: изд-во МГУ. 271 с.). Излагается по последней.
[2]Два слова о компаньонах — понятии, некогда бывшем ключевым, но в процессе развития этологии вполне забытом уже к 1960-м гг., хотя чрезвычайно важном. Все особи-элементы социальной системы являются социальными компаньонами того или иного рода (der Kumpan) К.Лоренца. Соответствующее немецкое слово имеет значения 1. соучастник, собутыльник, 2. сообщник. В обоих подчёркивается принадлежность к группе, маркируемой участием в определённой деятельности, и интенсивное общение при осуществлении данной деятельности, что и составляет смысл этологического понятия «Кумпан».
Компаньоны — это кормящие родители и выпрашивающие птенцы, резидент, охраняющий собственную территорию и вторгающийся туда захватчик, самец, привлекающий самку пением, и самки, перемещающиеся по территории между участками самцов в поисках оптимального партнёра. «Типичный» набор социальных компаньонов составляет der Kumpan особи или вида (Lorenz, 1935). Компаньоны «одного круга» постоянно воспроизводят «типичные взаимодействия в типичных обстоятельствах» в связи с распределением социальных ресурсов разного «качества» в анизотропном пространстве группировки — территорий, брачных партнеров, безопасных мест кормления и т.п.
Если индивид оказался в некой специфической ситуации взаимодействия с другими особями, или уже был вовлечён в социальное общение, он «автоматически» функционирует как «узел» (приёмник и передатчик сигнала) в системе обмена сигналами и/или стимулами с положительной обратной связью, на основе которой формируется коммуникативная сеть. Совокупности определённого типа «узлов» – то есть особей, связанных однородным процессом коммуникации в системное единство (ток, колонию, выводок, группировку территориальных самцов и пр.), были названы Лоренцем der Kumpan (компаньонство).
Увы, «не знаю, как перевести» понятие Kumpan – слово «компания» уже имеет иной смысл товарищества, вместо смысла коммерческого предприятия и конкуренции в Kumpan (в петровскую эпоху была «кумпания» именно с этим смыслом). «Сообщество» преоккупировано для обозначения связей, объединяющих животных в систему-социум, а не взаимодействий, которыми устанавливаются и поддерживаются эти связи. Иногда круг социальных компаньонов расширяется и дифференцируется: к выпрашивающим птенцам и кормящей самке добавляется самец, поющий и тревожащийся у гнезда, к резиденту и захватчику – проходящие особи, лениво атакуемые при непосредственном столкновении, и «активные вселенцы», чьи претензии занять территории пересекаются направленно, активно, часто несколькими соседями одновременно.
Компаньоны одного круга «равноправны» как участники общей системы взаимодействий, обменивающиеся стимулами и информацией по неким общим «правилам игры», и конкурирующие за одни и те же «возможности» улучшения социального статуса. Последние предусмотрены в структуре системы, сопрягающей поведение независимых индивидов, так что получается кооперативный эффект – рост устойчивости видоспецифической социальной системы достигается одновременно с эффективным разрешением взаимодействий и дифференциацией ролей особей в данной системе (а не противостоит ему). По отношению друг к другу компаньоны являются конкурентами за социальный ресурс, предоставленный дифференцированными ролями системы: выигрыш взаимодействий позволяет эксплуатировать этот ресурс, удержаться в соответствующей роли вопреки притязаниям других особей и впоследствии улучшить её. Каждый компаньон в процессе взаимодействия стремится максимизировать собственный выигрыш, управляя развитием процесса в собственных интересах, «перенаправив» развитие последнего к исходам, приемлемым для данной особи, а не соперника.
Для этого используются специфические управляющие воздействия на поведение компаньонов и течение процесса в целом и неспецифические воздействия, лишь возбуждающие участников и толкающие продолжать взаимодействие дальше, несмотря на ошибки и «сбои» прошлой коммуникации. Первые – это знаки и стимулы, реализованных на одном и том же «материальном носителе» – демонстрациях). Вторые — экспрессивные реакции, движения намерения, смещённые и переадресованные действия и т.п. преддемонстрации («сырьё», из которого в процессе «ритуализации по Тинбергену» (или «по Благосклонову», но другим путём) образуются «настоящие» демонстрации, Tinbergen, 1962, 1965).
[3]С одной стороны, это происходит при продуцировании животными «должных» форм демонстраций в разных (обычно «должных») ситуациях взаимодействия, с другой – при распознавании их реципиентом, в ходе в3осприятия и отреагирования.
Рекомендуем прочесть
 Как Европа открыла для себя Московскую Русь, а потом закрыла
Как Европа открыла для себя Московскую Русь, а потом закрыла COVID-19 вскрывает правду
COVID-19 вскрывает правду «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве «Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)
«Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)- Социальная сегрегация в городах
 «Жесты», вόроны и шалашники
«Жесты», вόроны и шалашники- Банковское дело и эволюция целей банковского регулирования
 Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей
