

Резюме. В продолжение темы копирования и «интуитивного понимания» действий другого, обслуживаемых зеркальными нейронами, см. соответствующий отрывок из книги их первооткрывателя. Фридрих Энгельс и Людвиг Нуаре были б ему особенно рады — среди прочего там показывается, как именно деятельность древних людей, в т.ч. трудовая, определила слова языка, возникшего несколько позже, и именно в плане членораздельности и акустической формы делений. Также как и идея синкинезии как модуса происхождения языка, опирающаяся на близкий корпус данных. Не менее важен вывод, следующий из этих данных, что «понимание Другого» связано с нервными, а не гуморальными механизмами, с копированием действий и извлечением из них концептов «типичного действия в типичной ситуации», а не влечением (или отвлечением), детерминированным гормональным состоянием, вроде обычных для гг.биологизаторов сентенций, что «чёрный» автоматически интерпретируется как «чужой», «опасный», а женщина — «сексуальный объект». Но этот механизм, обслуживаемый зеркальными нейронами, работает лишь в случае социального равенства, когда источник наблюдаемых действий воспринимается как «такой же как я». Если нет, он отключается, и нам кажется что «их» «нам никак не понять»: и самые опасные стереотипы против женщин, «цветных», других угнетённых групп работают именно на этом поле, утверждая что они «по природе другие«. Тогда внимательность к их поведению и копирование отключаются, как описано в книге — вместе с пониманием и эмпатией.

Джакомо Ризолатти
«Канонические и зеркальные нейроны
Содержание
Анализ функций зоны F5 показал нам, что большинство нейронов этой области коры разряжаются в процессе выполнения определенных действий, таких как хватание, удержание, манипулирование объектами, и что помимо этого часть из них отвечает на зрительные стимулы. Между моторными свойствами этих последних нейронов (например, специфическим типом захвата, который они кодируют) и их перцептивной избирательностью (к форме, размеру, ориентации объекта) существует тесная связь, которая определяет их важную роль в процессе преобразования зрительной информации о воспринимаемом объекте в моторные акты.
Эти нейроны были названы каноническими, потому что в течение долгого времени считалось, что премоторная кора может участвовать в зрительно-моторных преобразованиях. Однако, проведенные в начале 90-х годов эксперименты, в которых обезьян специально не учили выполнять задания, а позволяли им действовать самостоятельно, показали, что это не единственный тип нейронов, имеющих зрительно-моторные свойства1. К удивлению ученых, были обнаружены нейроны, активные и когда животное само выполняло моторный акт (например, брало пищу), и когда оно наблюдало за тем, как это делает экспериментатор. Такие нейроны были найдены в выпуклой части зоны F5 и названы зеркальными нейронами2.
Моторные свойства зеркальных нейронов аналогичны таковым у других нейронов зоны F5 в том, что они избирательно активируются в течение определенного моторного акта, но их зрительные свойства значительно отличаются.
В отличие от канонических нейронов, зеркальные не разряжаются при восприятии пищи или каких-либо иных трехмерных объектов, на их работу не влияет размер зрительного стимула. От чего их активность зависит — так это от наблюдения за определенными действиями, в которые вовлечена соответствующая часть тела (рука или рот), за взаимодействиями с объектами. Интересно, что копируемые моторные акты или нетранзитивные действия (не приложенные к определенному объекту), такие как поднятие или размахивание рукой, не вызывают ответа у таких нейронов. Также необходимо отметить, что активность зеркальных нейронов практически не зависит от положения в пространстве и расстояния от наблюдателя до наблюдаемого действия, хотя в некоторых случаях на нее влияет направление движения или используемая экспериментатором рука (правая или левая).
Если мы будем рассматривать зрительно кодируемый эффективный моторный акт в качестве критерия для классификации, зеркальные нейроны могут быть разделены на отдельные классы, сходные с теми, которые были выделены во второй главе при анализе моторных свойств у нейронов зоны F5. Итак, мы имеем «зеркальные нейроны хватания», «зеркальные нейроны удержания», «зеркальные нейроны манипулирования», а также «зеркальные нейроны размещения» (которые разряжаются, когда экспериментатор кладет объект на подставку) и «зеркальные нейроны взаимодействия с руками» (активные при наблюдении за рукой, движущейся в направлении другой руки, которая держит объект). Эта классификация показывает, что большинство зеркальных нейронов в F5 разряжаются при наблюдении за одним определенным типом действия (например, хватанием). Другие нейроны, похоже, не столь селективны, и разряжаются в ответ на наблюдение за несколькими — двумя или, реже, тремя — типами действий.
На рисунке 4.1 показано, как ведет себя типичный «зеркальный нейрон хватания». В условии А обезьяна наблюдает за тем, как экспериментатор поднимает с подноса кусочек пищи. Нейрон начинает разряжаться, как только рука экспериментатора достигает пищи и начинает принимать форму для ее захвата, и активность нейрона продолжается до конца действия. В условии Б обезьяна сама берет кусочек пищи, и в этом случае активность нейрона также коррелирует с изменением формы руки.
Сравнение разрядки нейрона при наблюдении и выполнении действия позволяет увидеть один из наиболее важных функциональных аспектов зеркальных нейронов: согласованность между кодированием выполнения моторного акта и наблюдением за выполнением его другим.
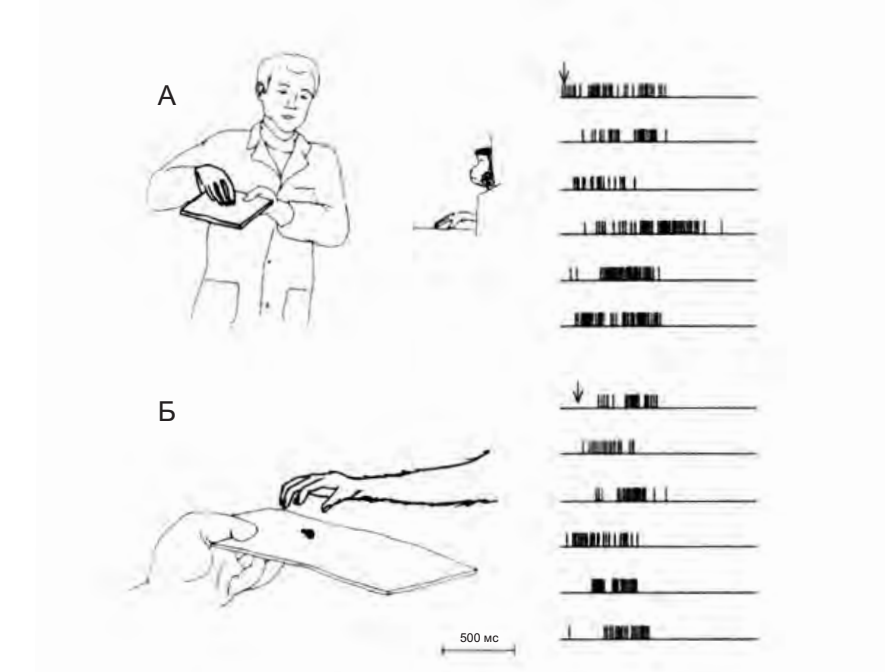
Рисунок 4.1. Зрительные и моторные ответы «зеркального нейрона хватания» (di Pellegrino et al. 1999)
Однако, у различных нейронов степень согласованности различается. Были выделены два основных типа: узко согласованные и широко согласованные. В нейронах с узкой согласованностью активность в ответ на выполняемое действие практически в точности повторяет активность при наблюдении. Пример приведен на рисунке 4.2. В условии А обезьяна наблюдает, как экспериментатор скручивает пальцами изюминку — вращая ее по часовой стрелке и против — как будто желая разорвать ее на две части. Нейрон разряжается только при вращении в одном из направлений. В условии Б экспериментатор и обезьяна держат одну и ту же изюминку, и нейрон разряжается, когда животное крутит ее в одном направлении, а экспериментатор — в другом, как бы пытаясь ее разорвать. В условии В обезьяна берет кусочек пищи с подноса при помощи движения захвата — нейрон не проявляет никакой активности.
В нейронах с широкой согласованностью кодируются наблюдаемое и выполняемое действие, которые явно связаны друг с другом, хотя могут не быть идентичны, и их связь может быть представлена на разном уровне обобщения. На самом деле, некоторые нейроны отвечают только на выполнение одного моторного акта (например, хватания) и на восприятие двух выполняемых актов (хватание и удержание).

Рисунок 4.2. Пример зеркального нейрона с узкой согласованностью. А — обезьяна наблюдает, как экспериментатор крутит изюминку в своих руках движениями, направленными в разные стороны; нейрон активен только при одном из двух направлений движения. Б — экспериментатор вращает кусочек пищи, который обезьяна держит в руке; животное вращает его в противоположном направлении. В — обезьяна берет пищу при помощи точного захвата. Для каждого условия показаны четыре серии записей активности нейрона. Стрелки показывают направление вращения в определенный момент записи (Rizzolatti et al. 1996a)
Другие кодируют только один выполняемый и наблюдаемый акт, но с разной степенью селективности. К примеру, рассмотрим нейрон, представленный на рисунке 4.3. Он разряжается, когда обезьяна наблюдает за экспериментатором, берущим объект при помощи точного захвата или захвата всей кистью, но отвечает на действия обезьяны, только когда она использует точный захват.
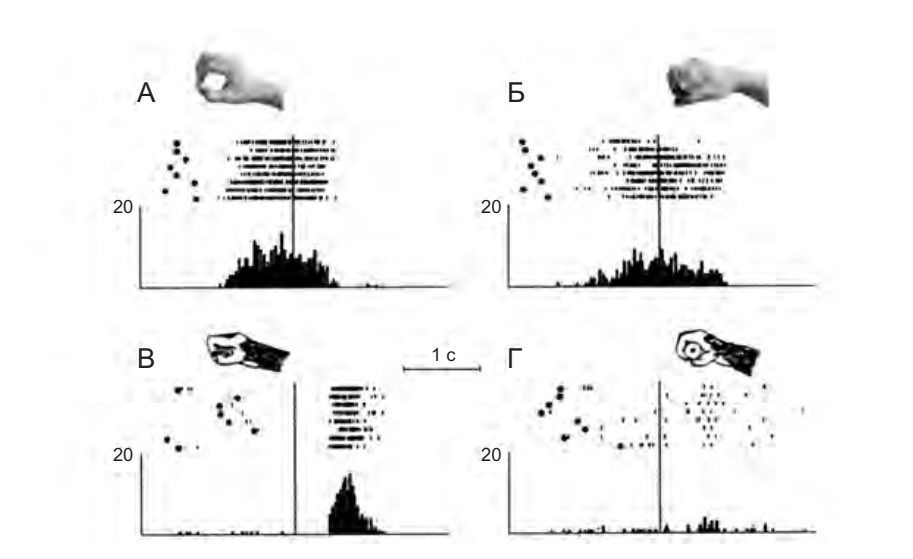
Рисунок 4.3. Пример широко согласованного нейрона. А — экспериментатор берет кусочек пищи, используя точный захват. Б — экспериментатор берет пищу всей кистью. В — обезьяна берет пищу точным захватом. Нейрон обладает высокой избирательностью к активному действию, но не к его наблюдению (Адаптировано из работы Gallese et al. 1996)
Кроме того, существуют нейроны, кодирующие действия зрительно — они разряжаются при выполнении другого действия, связанного с первым. Зеркальные нейроны такого типа разряжаются при наблюдении за тем, как экспериментатор кладет пищу на поднос, а также когда он ее берет. Функциональная роль и теоретическое значение таких нейронов подробно не обсуждались, однако, весьма вероятно, что их поведение есть последовательность организованных в цепь моторных актов. Мы обсудим этот вопрос в следующей части этой главы. С учетом различных типологий, нейроны с широкой степенью согласованности составляют примерно 70 % от общего числа зеркальных нейронов в коре обезьяны.

Коррадо Синигалья
Поедание и общение
До сих пор мы приводили примеры только тех зеркальных нейронов, которые связаны с движениями рук. Практически все первые исследования зеркальных нейронов были сконцентрированы вокруг дорзальной части зоны F5, большинство представленных в которой движений связаны с рукой. В первой главе мы отмечали, как при помощи микростимуляции нейронов электрическим током и регистрации их активности было показано, что движения рта также контролируются вентральной частью F5. Последние исследования показали, что нейроны этой области обладают зрительно-моторными свойствами так же, как и типичные зеркальные нейроны, и отвечают на выполнение движений ртом и наблюдение за движениями рта у других3.
На рисунке 4.4 показано выполнение экспериментатором и обезьяной некоторых действий, которые используются для оценки специфичности моторных и зрительных ответов нейронов животного. Рисунки вверху и в центре иллюстрируют два типичных транзитивных движения (то есть, действия, выполнение которых связано с объектом), которые включают заглатывание жидкости или твердой пищи. На рисунке внизу — нетранзитивное действие (вытягивание губ), которое наряду с чмоканием и оскаливанием зубов входит в репертуар коммуникативной мимики обезьян.
Большая часть зеркальных нейронов (около 85%) отвечают на восприятие действий, таких как хватание кусочка пищи губами, пережевывание или сосание его. Они были названы нейронами глотания. С функциональной точки зрения, эти нейроны сходны с зеркальными нейронами, кодирующими движения руки — они также разряжаются только при взаимодействии соответствующей части тела с объектом, а простое наблюдение покоящегося объекта или совершения нетранзитивного действия не вызывает никакого значимого ответа. Кроме того, большинство таких нейронов являются избирательными к определенному типу действия, и примерно треть из них — узко согласованные с ним (рисунок 4.5).
Зеркальные нейроны, отвечающие на восприятие коммуникативных актов, производимых при помощи рта, ведут себя иначе. На рисунке 4.6 приведены активности двух нейронов: в первом случае экспериментатор чмокает губами (А), вытягивает губы (Б), пьет из шприца (В). Только в условии А нейрон демонстрирует значимую активность. Но тот же самый нейрон становится активным, когда животное хватает пищу при помощи рта, при этом слегка вытягивая губы и язык (Г). Во втором случае экспериментатор вытягивает губы (А), держит пищу между зубами (Б), предлагает пищу обезьяне (В) — в этом случае нейрон также активируется только в условии А, и также разряжается, когда обезьяна выполняет типичное действие, связанное с глотанием — сосание сока из шприца (Г).
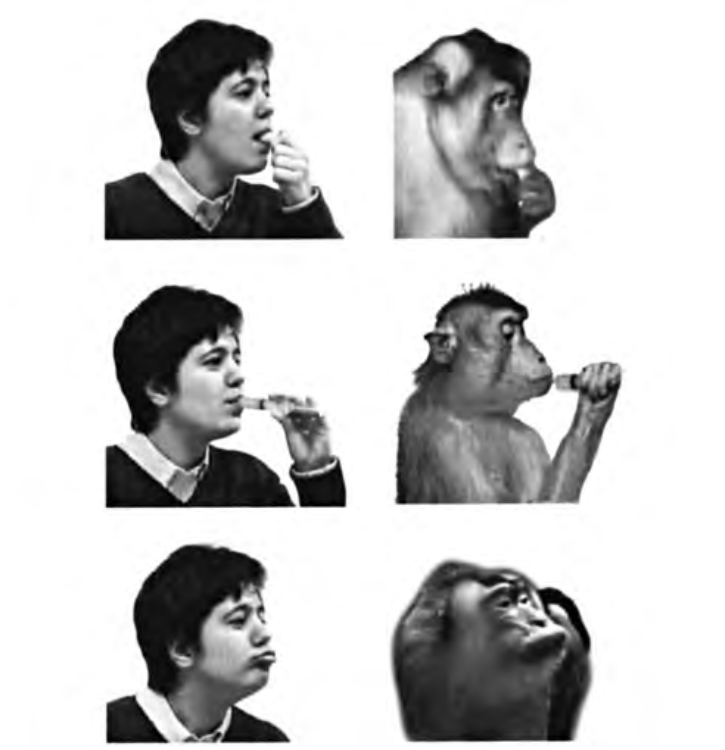
Рисунок 4.4. Примеры транзитивных и нетранзитивных действий экспериментатора и обезьяны, используемых для изучения зеркальных нейронов, кодирующих движения рта. Сверху вниз: захватывание пищи ртом; всасывание апельсинового сока из шприца; вытягивание губ.
Необходимо отметить, что в отличие от других зеркальных нейронов, коммуникативные нейроны отвечают при рассматривании нетранзитивных действий. Можно возразить, что эти ответы не связаны со зрительными стимулами как таковыми, но скорее с интерпретацией их животным как актов глотания. Другими словами, простое действие по вытягиванию языка экспериментатором вызовет моторную репрезентацию лизания у обезьяны. Такое объяснение кажется вполне разумным только потому, что оно придает законченный вид и единство теории о зеркальных нейронах. Однако, к сожалению, оно не объясняет того факта, что наблюдение за действиями глотания едва ли вызывает какую-либо активность у коммуникативных зеркальных нейронов.
Остается прояснить еще один важный вопрос: в отличие от зеркальных нейронов, которые связаны с движениями руки и поеданием пищи, коммуникативные зеркальные нейроны не демонстрируют согласованность между зрительным и моторным ответом.
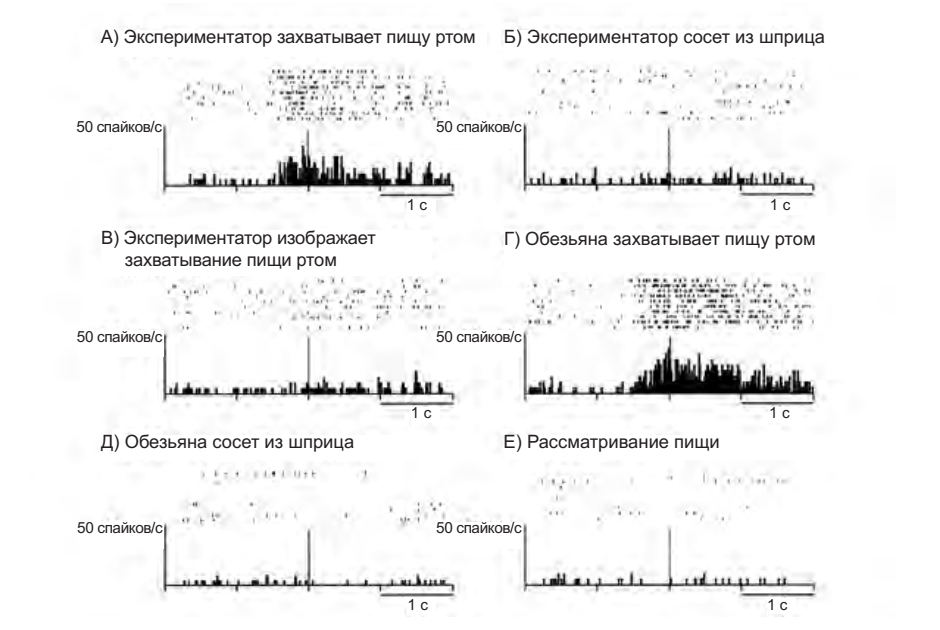
Рисунок 4.5. Пример активности «зеркального нейрона хватания ртом». А — экспериментатор наклоняется к подносу и берет с него пищу зубами. Б — экспериментатор подносит голову к шприцу, закрепленному на подставке, и сосет сок. В — экспериментатор изображает то же действие, что и в условии А, но без пищи. Г — экспериментатор подносит пищу к обезьяне, которая берет ее зубами и съедает. Д — экспериментатор подносит к обезьяне шприц с соком, обезьяна обхватывает губами шприц и сосет сок. Е — экспериментатор насаживает пищу на палочку и перемещает ее в поле зрения животного. На каждом графике представлены серии из десяти проб и усредненные гистограммы. Начало отсчета — момент, в который губы экспериментатора (зрительное кодирование) или обезьяны (моторное кодирование) касаются пищи, или в который пища попадает в поле зрения обезьяны. На рисунке В графики выровнены относительно момента окончания движения. По абсциссе — время в секундах, по ординате — количество спайков за каждые 20 мс
Только первый из них по своей природе коммуникативный, тогда как второй связан с глотанием. Если мы рассмотрим чисто моторные свойства, то в большинстве случаев найдем высокую корреляцию между наблюдаемым и выполняемым действиями: нейрон, который разряжается при наблюдении за вытягиванием губ, активен и при действиях обезьяны, требующих сходных движений (например, сосании сока из шприца), но не отвечает при выполнении других действий. То же самое верно и для чмокания губами. Однако, необходимо помнить, что значения этих действий различаются. Убедить обезьяну выполнять чисто коммуникативные действия в экспериментах с регистрацией активности отдельных нейронов — задача не из легких.
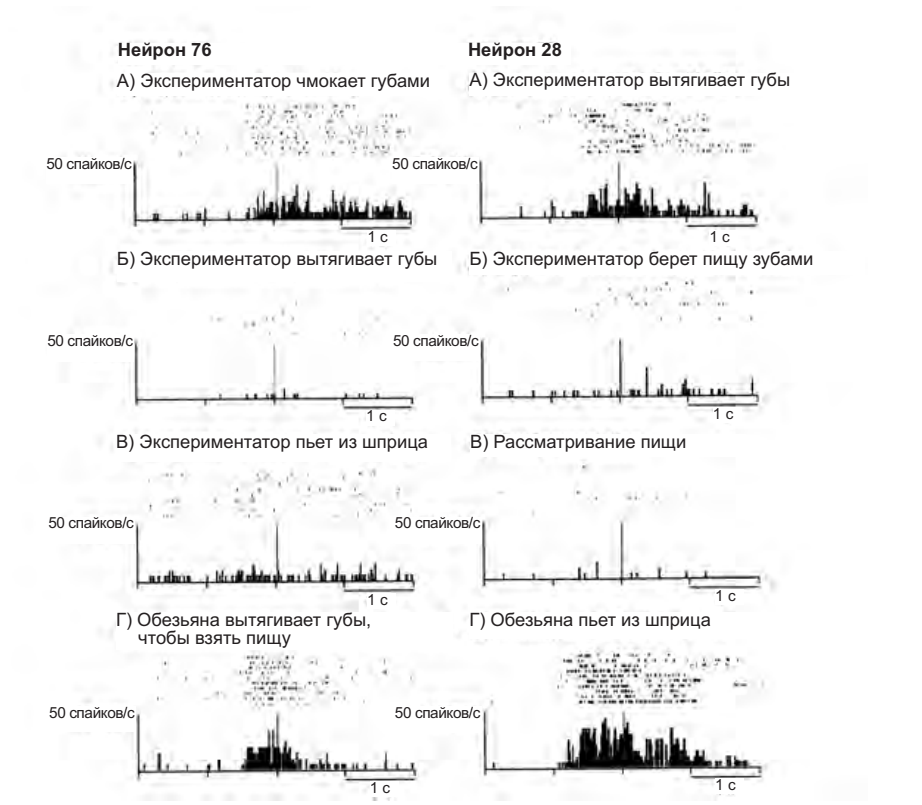
Рисунок 4.6. Примеры двух коммуникативных зеркальных нейронов. Активность в отдельных пробах и результирующие гистограммы выровнены относительно пика действия. Нейрон 76: А — экспериментатор чмокает губами перед обезьяной, Б — экспериментатор вытягивает губы, В — экспериментатор придвигается к закрепленному шприцу с соком и втягивает содержимое, Г — экспериментатор предлагает пищу обезьяне, которая вытягивает губы и хватает ее. Нейрон 28: А — экспериментатор вытягивает губы, глядя на обезьяну, Б — экспериментатор придвигается к пище, закрепленной на подставке, хватает ее зубами и держит, В — экспериментатор помещает палочку с кусочком пищи перед обезьяной, Г — экспериментатор предлагает обезьяне попить из шприца, обезьяна пьет (Ferrari et al. 2003)
Тем не менее, в редких случаях, когда это удавалось, были зарегистрированы четкие нейронные ответы на коммуникативные действия (рисунок 4.7). Эти результаты вселяют в нас надежду, что такая активность свойственна многим, а не только нескольким, нейронам.
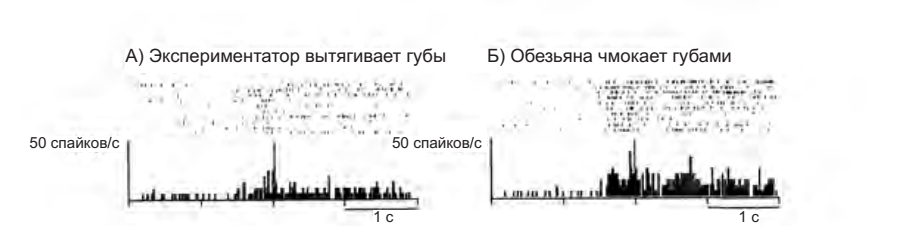
Рисунок 4.7. Пример коммуникативной зеркальной системы, кодирующей вытягивание губ. А — экспериментатор вытягивает губы, глядя на обезьяну. Б — обезьяна отвечает на чмокание экспериментатора чмоканием (Ferrari et al. 2003)
Если оставить в стороне трудности, с которыми мы можем столкнуться в экспериментальных условиях, тот факт, что действия поедания пищи и общения имеют общий нейронный субстрат, кажется нам исключительно интересным, особенно в свете некоторых этологических исследований на приматах4. Коммуникативные действия, такие как чмокание или вытягивание губ, сформировались на основе репертуара движений, изначально связанных с поеданием и грумингом. Хорошо известно, что у приматов груминг является одним из важнейших способов выражения дружелюбного отношения и солидарности с сообществом: он способствует образованию группы, а когда она становится слишком большой — разделению ее на коалиции с целью защиты слабейших членов от агрессии других животных. Когда обезьяна начинает чистить и вычесывать насекомых из шкуры другой обезьяны, она часто сопровождает или предваряет свои первые движения чмоканием. Кормление также предваряется чмоканием, но с другим звуком, словно обезьяны хотят подчеркнуть различия между этими действиями. Таким образом, чмокание в отсутствие груминга оказывается формой ритуализированного моторного акта, который преобразует функции, связанные с объектом, в коммуникативные. То же самое верно и для таких движений, как вытягивание губ или языка. В таком контексте открытие коммуникативных зеркальных нейронов в такой зоне, как F5, вместе с несомненным отсутствием согласования между зрительным и моторным ответами, отражает первичный процесс обработки в коре коммуникативных сигналов, которые еще не полностью отделились от пищевого поведения (то есть, от транзитивных действий по поднесению пищи ко рту и ее заглатыванию).
Связи верхней височной извилины и нижней теменной коры
Как мы видели во второй главе, большая часть зрительной информации, поступающей к каноническим нейронам F5, приходит к ним из передней межтеменной зоны (AIP) [The anterior intraparietal]. Теперь перед нам стоит вопрос о том, какие участки коры поставляют информацию в зеркальные нейроны.
Более пятнадцати лет назад Дэвид И. Перретт и его коллеги5 показали в эксперименте на обезьянах, что передняя часть верхней височной борозды (STS) содержит нейроны, селективно отвечающие на восприятие большого ряда движений тела, выполняемых другим индивидом — некоторые нейроны были активны, когда обезьяна наблюдала за движениями глаз или головы, другие отвечали на движения туловища и ног (хождение), а третьи снова кодировали определенные взаимодействия рук с объектом.
Зрительные свойства нейронов последнего типа кажутся весьма сходными с таковыми у зеркальных нейронов зоны F5. И те, и другие кодируют в значительной мере сходные типы наблюдаемых движений на разном уровне обобщения (например, хватание рукой, использование точного захвата и т. д.) и неактивны при наблюдении за тем, как экспериментатор выполняет нетранзитивные действия или изображает транзитивные действия в отсутствие объектов.
Однако, можно отметить одно существенное расхождение между ними: в отличие от зеркальных нейронов зоны F5, нейроны STS являются чисто зрительными и не принимают участия в кодировании движений. Следовательно, они не обладают свойством совмещения зрительных и моторных функций, что является важной характеристикой зеркальных нейронов.
Нейроны верхней височной борозды чрезвычайно интересны не только из-за их свойств, но также потому, что они помогают нам понять, каким образом возможно возникновение таких сложных по своим функциям клеток как зеркальные нейроны. Они демонстрируют, как происходит кодирование биологических движений, выполняемых другими индивидами, в специфической корковой системе, и как процесс идентификации этих движений начинается в зрительной коре. Следующий шаг кажется довольно простым — предположить, что зрительная информация передается в моторную кору, обеспечивая зрительные свойства моторных нейронов.
Далее встает вопрос о том, как информация передается из STS в F5. С анатомической точки зрения, STS не имеет прямых проекций в вентральную премоторную кору. Однако, она имеет мощные связи с нижней теменной корой и участками префронтальной коры6. Зрительная информация о наблюдаемых действиях может, таким образом, достичь зоны F5 по одному из этих путей. Скорее всего, второй из них не играет важной роли — известно, что связи F5 с префронтальной корой, получающей информацию от STS, не очень сильны. С другой стороны, F5 связана с ростральной частью нижней теменной коры, в которую входят зоны PF и PFG5.
Более того, функции нейронов области PF—PFG говорят о том, что этот комплекс может рассматриваться как мост между зонами STS и F5. В работах Яри Хюваринена и его коллег6, проведенных в 80-е годы, было показано, что нейроны этого комплекса отвечают на сенсорные (соматосенсорные и зрительные) стимулы, и что около трети из них также активируются при выполнении произвольных движений руки и рта. Позднейшие исследования7 показали, что примерно 40 % нейронов, отвечающих на зрительные стимулы, активны во время наблюдения за моторным актом, в который включена рука — например, хватанием, удержанием, дотягиванием, — и что еще более важно, большая часть нейронов (около 70 %) обладает моторными свойствами, отвечая при выполнении обезьяной действий при помощи рук, рта или того и другого. Это теменные зеркальные нейроны.
Как и нейроны зоны F5, теменные зеркальные нейроны не отвечают на простое наблюдение за пассивным субъектом или покоящимся объектом действия, и даже на изображение действия в отсутствие объекта. Половина из них избирательно кодирует только один тип моторного акта, другая половина избирательна к двум видам действий (например, хватание и отпускание). Учитывая связи между наблюдаемым и выполняемым действиями, они работают так же, как и зеркальные нейроны в F5: некоторые из них являются узко согласованными, большинство — широко согласованными.
Функции зеркальных нейронов
Теперь нам предстоит прояснить функциональное значение зеркальных нейронов, зарегистрированных в зонах F5 и PF—PFG. На первый взгляд, мы можем приписать активность этих нейронов при восприятии обезьяной действий, выполняемых другими (в нашем случае, экспериментатором), неспецифическим факторам, таким, как внимание или ожидание пищи, либо подготовке к действию, которая обеспечивает повторение животным увиденных действий настолько быстро, насколько это возможно, чтобы быть готовым к возможному соперничеству с окружающими. Если бы дело обстояло так, то зеркальные нейроны либо были бы лишены всякой специфичности, либо просто представляли бы собой определенную категорию «подготовительных нейронов», которые широко распространены в премоторной коре и которые активируются перед эффективным выполнением действия.
Более внимательный взгляд покажет, что ни одну из этих гипотез нельзя принять. Избирательность ответа и зрительно-моторное согласование, обнаруженные у большинства зеркальных нейронов, не могут быть отнесены на счет поведения, связанного с ожиданием пищи или какой-либо другой формы поощрения. Эксперимент, представленный на рисунке 4.8, подтверждает это. В условиях А и Б обезьяна, у которой регистрировалась активность нейронов, наблюдала за другой обезьяной или экспериментатором, когда они брали пищу при помощи рук. В условии В обезьяна сама выполняла такое действие. Зеркальные нейроны разряжались в условиях А и Б, несмотря на то, что в обоих условиях обезьяна не могла дотянуться до пищи или получить какую-либо другую награду.
Объяснение активности зеркальных нейронов как формы подготовки к действию не является удовлетворительным. На рисунке 4.8 можно увидеть, что в то время как первая обезьяна наблюдает за второй, хватающей пищу, ей нет никакого смысла готовиться к действию — она не может дотянуться до пищи. Кроме того, читатель вспомнит, что ни в одном из вышеописанных экспериментов разрядка зеркальных нейронов при наблюдении за выполнением действия не сопровождалась последующим самостоятельным его выполнением. Мы не должны забывать, что зеркальные нейроны никогда не активировались, если пища предлагалась обезьяне на таком расстоянии, что до нее можно было дотянуться. Если бы ответ нейронов был связан с подготовкой к действию, они бы разряжались перед тем, как обезьяна сама его выполняла.
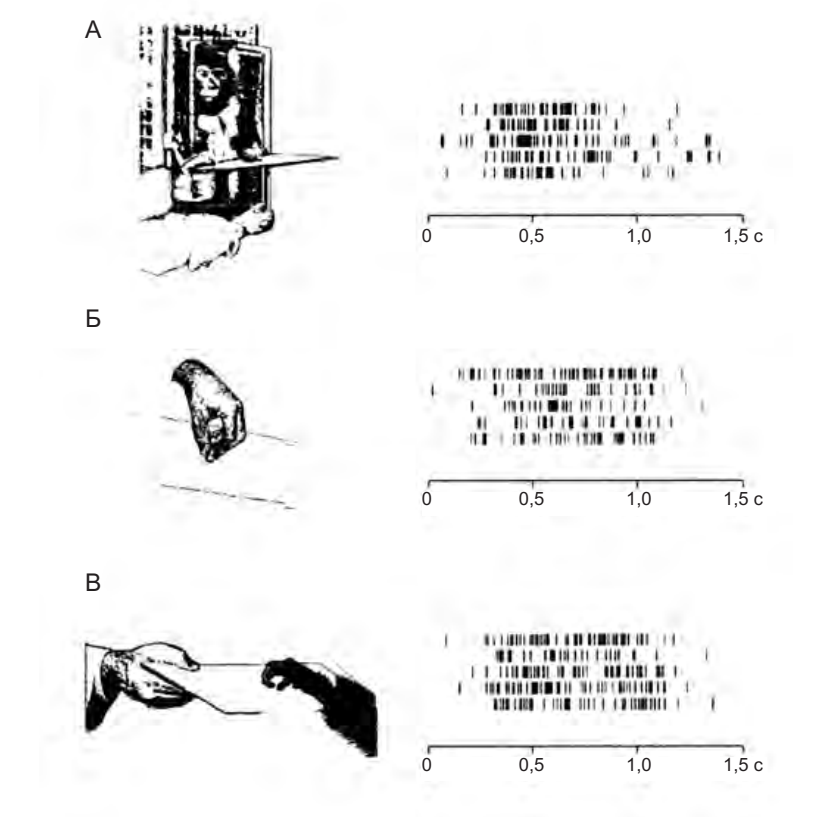
Рисунок 4.8. Активация «зеркального нейрона хватания». А — обезьяна, активность нейронов которой регистрируется, наблюдает за другой обезьяной, берущей пищу при помощи рук. Б — обезьяна смотрит, как экспериментатор берет пищу. В — обезьяна сама выполняет это действие. Для каждого условия показаны по пять записей активности нейрона. Спонтанная активность практически отсутствовала (Rizzolatti et al. 1996a)
Несколько лет назад Марк Жанро в своей статье, посвященной воображаемым действиям, предложил другую (более сложную) интерпретацию функций зеркальных нейронов8. Представьте себе урок музыки и ученика, внимательно следящего за тем, как его преподаватель исполняет сложный пассаж на скрипке, который ему нужно будет повторить после того, как учитель закончит играть. Для этого ученику нужно построить моторный образ быстрых движений пальцев маэстро. Далее, согласно Жанро, те же нейроны, которые отвечают за построение моторного образа, будут активны и во время подготовки с ученика и исполнения пьесы. Другими словами, активация зеркальных нейронов позволяет создавать «внутренние моторные репрезентации» наблюдаемых действий, на которых основана способность к обучению при помощи подражания.
Предположение Жанро чрезвычайно важно и согласуется с результатами экспериментов, которые мы рассматривали ранее. Тесная связь между зрительными и моторными ответами зеркальных нейронов свидетельствует о том, что в то время как индивид наблюдает за выполнением действий другими, в его мозге выполняется потенциальный моторный акт, по всем своим характеристикам сходный с тем, который спонтанно активируется при подготовке и выполнении реального действия. Различие лишь в том, что в одном случае это действие остается на стадии потенциального (в виде «внутренней моторной репрезентации»), а в другом оно переводится в последовательность конкретных движений. Однако, в одном вопросе мы не можем согласиться с Жанро, а именно, мы не верим в то, что основной функцией зеркальных нейронов является обеспечение имитации действий.
Далее мы более глубоко проанализируем широкий ряд феноменов, которые часто рассматриваются как — и часто путаются с — имитацией; также мы оценим, до какой степени способность человека обучаться выполнению действий после наблюдения за тем, как их выполняют другие, зависит от функционирования системы зеркальных нейронов. В любом случае, за последние годы этологи все больше стали утверждаться во мнении, что имитация в правильном смысле этого слова — прерогатива человека и (возможно) человекообразных обезьян, но не макак, на которых проводились описанные нами эксперименты9. Поэтому мы не можем полностью согласиться с интерпретацией Жанро: функции зеркальных нейронов зоны F5 и комплекса PF—PFG имеют более древнее эволюционное происхождение, и на основании приведенных примеров можно сказать, что изначально они были включены в понимание значений «моторных событий» — то есть, действий, производимых другими10.
Используемый нами термин «понимание» не обязательно означает, что наблюдатель (в данном случае, обезьяна) имеет эксплицитное знание или даже осознает то, что наблюдаемое и выполняемое действия идентичны или очень похожи. То, что мы утверждаем, намного проще — мы говорим о способности незамедлительно распознавать определенные типы действий в наблюдаемом «моторном событии», типы действий, которые характеризуются определенным способом взаимодействия с объектами. Это нужно для того, чтобы отличить это действие от других и затем использовать это знание для выбора наиболее подходящего ответа. Таким образом, то, что ранее было сказано относительно канонических нейронов зоны F5 и зрительно-моторных нейронов передней межтеменной коры (AIP), остается верным и в данном случае: зрительный стимул начинает обрабатываться незамедлительно, начиная с соответствующего моторного акта, даже если последний не был реально выполнен. Есть только одно важное различие — в случае с зеркальными нейронами зрительный стимул это не объект или его движения, но движения другого индивида, направленные на хватание, удержание, манипулирование объектом.
Как и в случае с объектами, эти движения приобретают значение для индивида благодаря словарю моторных актов, который регулирует его способность выполнять действия. У обезьяны этот словарь включает захватывание пищи, ее удержание, поднесение ко рту, и так далее.
Вот почему когда она видит, как экспериментатор сгибает свою руку для точного захвата и протягивает ее по направлению к пище, она незамедлительно воспринимает значение этих «моторных событий» и интерпретирует их в терминах целенаправленного действия.
Зрительная репрезентация и моторное понимание действия
Однако, на это имеется и очевидное возражение: как уже обсуждалось ранее, нейроны, избирательно отвечающие на наблюдение за движениями тела других, и в некоторых случаях — за взаимодействием руки и объекта — были обнаружены в передней части верхней височной борозды (STS). Мы отмечали, что эта область связана с зрительной — височной и затылочной — корой и формирует систему, во многом параллельную вентральному потоку (см. рис. 2.7). Какой же смысл тогда говорить о системе зеркальных нейронов, которая обеспечивает формирование в мозге наблюдателя образа действия, производимого другим, и описание его в терминах его собственного моторного акта? Не будет ли намного проще предположить, что понимание действий других основано на чисто зрительных механизмах анализа и синтеза различных элементов, составляющих наблюдаемое действие, без какого-либо моторного вмешательства со стороны наблюдателя?
Перретт и его коллеги показали11, что зрительное кодирование действий в передних областях STS производится на чрезвычайно высоком уровне сложности. Для примера, в этой зоне имеются нейроны, способные объединять получаемую при наблюдении информацию о направлении взгляда и выполняемых движениях. Такие нейроны активируются, только когда обезьяна наблюдает, как экспериментатор берет предмет, на который он при этом смотрит. Если экспериментатор переведет свой взгляд в другую сторону, наблюдение за его дейcтвиями не вызовет никакой значимой активации нейрона. Однако, мы должны выяснить, можно ли эту избирательность — или, в более общем смысле, способность связывать различные воспринимаемые аспекты выполняемого действия — обозначать термином «понимание». Особенности моторной активации нейронов зон F5 и PF—PFG привносят то, что не может быть выведено из чисто зрительных свойств нейронов STS, и без чего взаимосвязь воспринимаемых аспектов действия будет в лучшем случае случайной, ничего не значащей для наблюдателя.
С точки зрения моторных свойств, связь между действием по дотягиванию до предмета и направлением взгляда, в действительности, с не является случайной: еще в колыбели мы научаемся тому, что лучший способ получить объект — уставиться на него. Как и все успешные стратегии, эта становится частью нашего словаря действий, и, таким образом, когда мы видим, как кто-либо воспроизводит это действие, наша моторная система входит в резонанс с ним, благодаря чему мы распознаем интенциональный аспект движений другого и понимаем тип его действий.
Зрительно-моторные свойства зеркальных нейронов позволяют им координировать зрительную информацию с моторным знанием. Активация зеркальных нейронов как моторных во время выполнения действия характеризуется не только тем, что кодируются тип, модальность и временная развертка действия, но также обеспечивается контроль за его выполнением. А контроль за двигательным процессом невозможен без механизма антиципации, и, таким образом, каждый контролируемый процесс определяет корреляцию между соответствующей нейронной активностью и эффектами, которые она порождает. В конкретном случае зон F5 и PF—PFG взаимная координация позволяет сформировать базовое моторное знание о значениях действий, кодируемых различными нейронами — знание, которое может быть использовано как во время выполнения действия, так и при наблюдении за его выполнением другими.
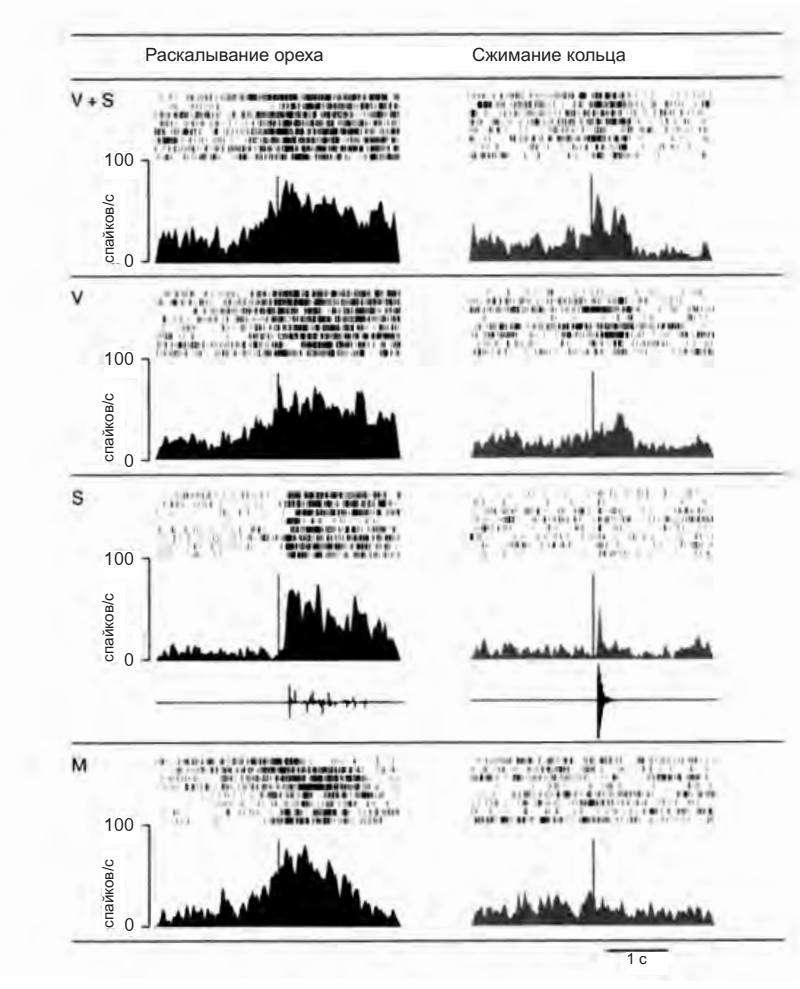
Рисунок 4.10. Два «аудиовизуальных» зеркальных нейрона F5. Вертикальными линиями на гистограммах отмечено начало звука в условиях «зрение и слух» (V + S) и «слух» (S). В условии «зрение» (V) они обозначают тот момент, в который мог бы начаться звук, если бы он предъявлялся. В чисто моторном условии (M) они отмечают момент, когда обезьяна дотрагивается до объекта (Kohler et al. 2003)
Активация той же самой совокупности нейронов говорит о том, что понимание действий других предполагает наличие у наблюдателя того же самого знания о принципах выполнения моторных актов, которое регулирует его собственное выполнение действий.
В некоторых недавних исследованиях была подтверждена гипотеза о том, что моторное знание играет фундаментальную роль в обработке сенсорной информации — до той степени, что без нее вообще было бы сложно говорить о понимании действия12.
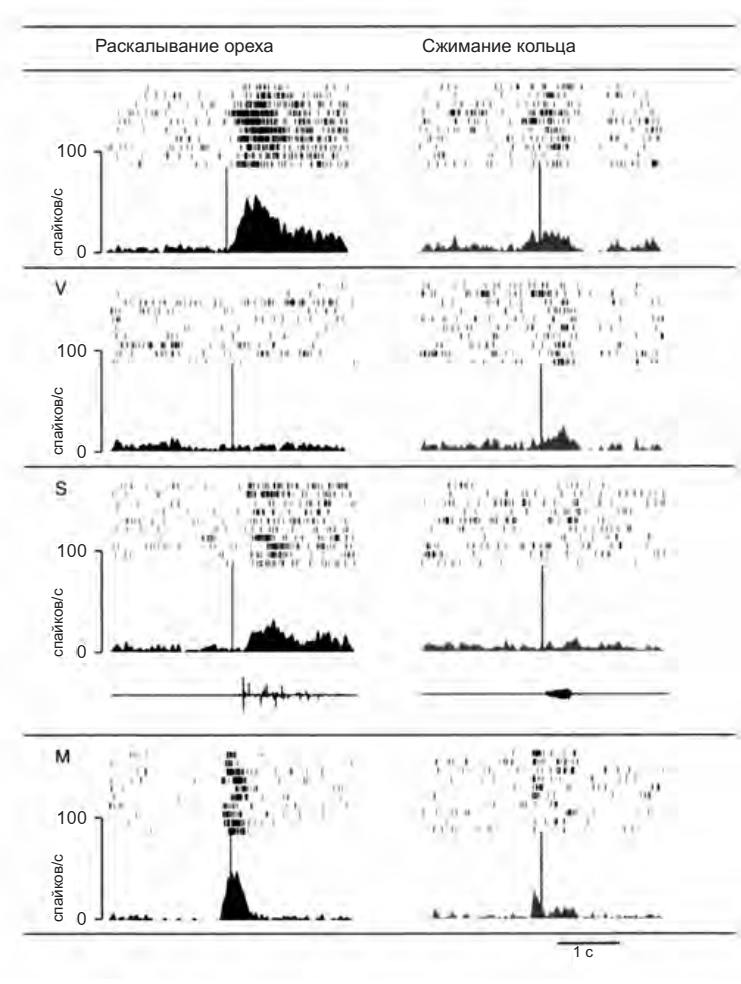
Рисунок 4.10. (продолжение)
Мария Алессандра Умильта и ее коллеги показали, что большинство нейронов зоны F5 отвечают при выполнении действий экспериментатором, независимо от того, что их финальная фаза (то есть, наиболее важный этап — непосредственное взаимодействие руки с объектом) скрыта от обезьяны. Эксперимент представлен на рисунке 4.9 (см. цв. вклейку). Активность отдельных нейронов регистрировалась в четырех различных условиях: А — обезьяна наблюдает за тем, что делает экспериментатор, и полностью видит последнюю часть действия (захват объекта); Б — она видит только начало действия, так как его окончание скрыто экраном; В и Г — условия, аналогичные условиям А и Б, за исключением того, что экспериментатор изображает действие при отсутствии объекта.
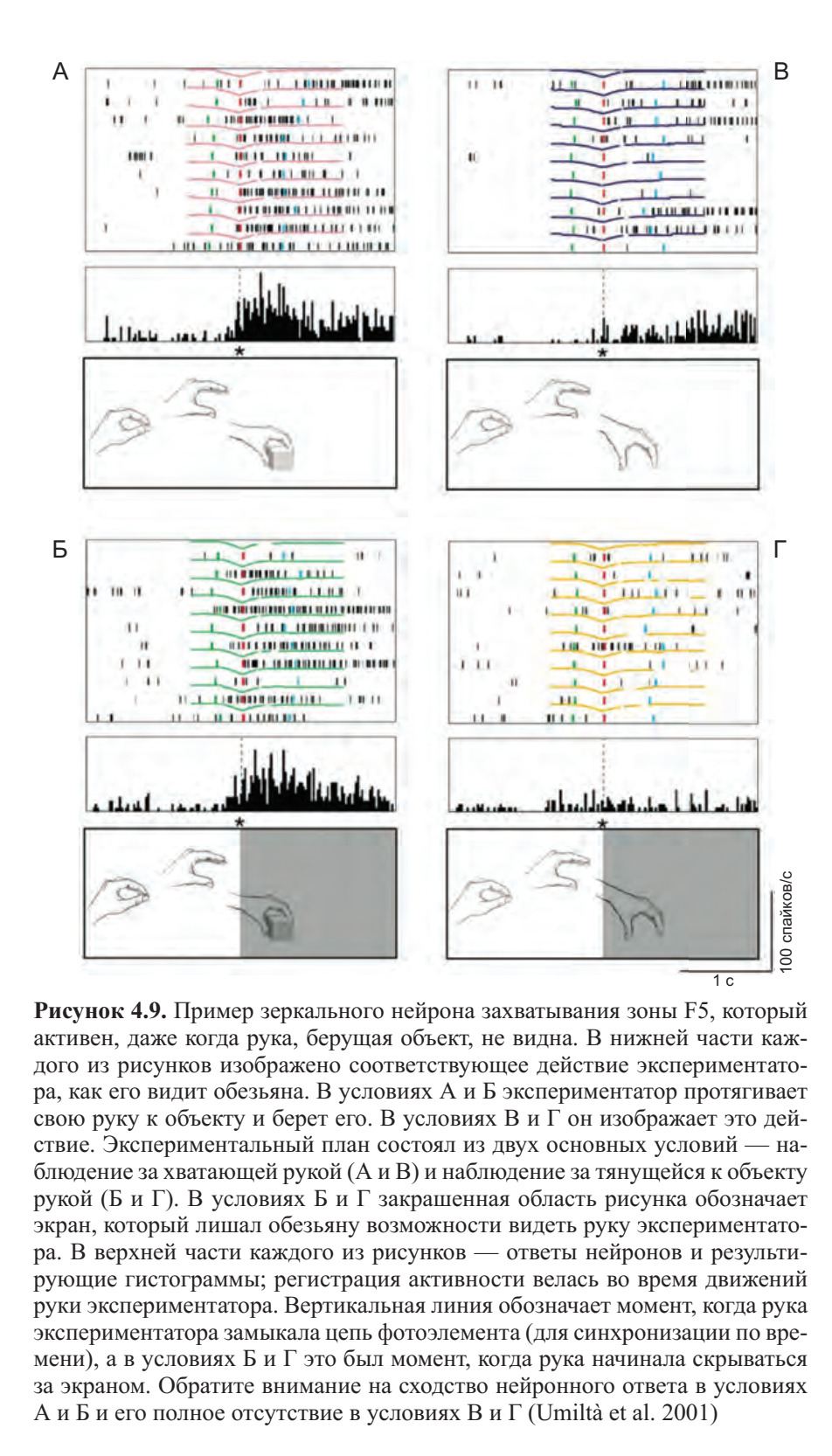 Данные регистрации активности нейронов показали: тот факт, что обезьяна не могла видеть финальную часть действия, никоим образом не изменял паттерн активации нейрона по сравнению с условием, в котором рука экспериментатора была видна на протяжение всего действия. В условии Б обезьяна видела, что объект закрывается экраном. Однако, нельзя говорить, что ответ нейрона есть всего лишь «реминисценция объекта», так как в таком случае разрядка нейрона началась бы, как только объект попадал в поле зрение, а это было не так. Напротив, поведение нейрона говорит о том, что тот же самый моторный акт, только потенциальный, происходит и когда обезьяна наблюдает действие полностью, и когда видит только его начало. В действительности, именно этот потенциальный моторный акт («внутренняя моторная репрезентация») позволяет обезьяне восстановить отсутствующую часть действия и распознать его значение по увиденной части.
Данные регистрации активности нейронов показали: тот факт, что обезьяна не могла видеть финальную часть действия, никоим образом не изменял паттерн активации нейрона по сравнению с условием, в котором рука экспериментатора была видна на протяжение всего действия. В условии Б обезьяна видела, что объект закрывается экраном. Однако, нельзя говорить, что ответ нейрона есть всего лишь «реминисценция объекта», так как в таком случае разрядка нейрона началась бы, как только объект попадал в поле зрение, а это было не так. Напротив, поведение нейрона говорит о том, что тот же самый моторный акт, только потенциальный, происходит и когда обезьяна наблюдает действие полностью, и когда видит только его начало. В действительности, именно этот потенциальный моторный акт («внутренняя моторная репрезентация») позволяет обезьяне восстановить отсутствующую часть действия и распознать его значение по увиденной части.
Исследование, проведенное Эвелин Келер и ее коллегами13, является еще одним доказательством в пользу гипотезы, что активация моторных нейронов отражает значение наблюдаемого действия и не зависит только от его зрительно воспринимаемых характеристик. Авторы выделили особый тип бимодальный зеркальных нейронов в зоне F5 (аудиовизуальные нейроны), которые активируются и при наблюдении обезьяной за действиями экспериментатора, сопровождающимися характерными звуками, и когда животное слышит эти звуки без зрительного восприятия действия.
На рисунке 4.10 представлена активность двух таких нейронов: они не только являются селективными к определенному действию (например, раскалыванию земляного ореха), но также их ответы в различных экспериментальных условиях (зрительное и слуховое восприятие; только зрение; только слух; самостоятельное выполнение действия) совершенно совпадают. Это означает, что вызываемый потенциальный моторный акт всегда один и тот же, тогда как сенсорная информация может меняться в зависимости от конкретной ситуации.
Зрительные характеристики действия оказываются релевантными только до той степени, пока они облегчают понимание действия. Однако, если для этого достаточно других факторов (например, характерного звука), зеркальные нейроны способны кодировать действия экспериментатора даже в отсутствие зрительных сигналов.
Мелодия действия и понимание намерений
Демонстрация того, что зеркальные нейроны зон F5 и PF—PFG обеспечивают понимание действий других, не должна пониматься как единственная функция зеркального механизма. Далее мы приведем целый ряд иллюстраций разнообразных функций зеркальных нейронов и карту корковых зон, включенных в эту систему как у человека, так и у других приматов. Однако, это ни в коей мере не отменяет значимости механизма понимания действий, обеспечиваемого работой нейронов зон F5 и PF—PFG и отмеченного согласованностью их сенсорных и моторных ответов. Это, как мы уже видели, является имплицитной формой понимания их происхождения и значения — оно не связано с конкретной сенсорной модальностью восприятия действия, но ограничено имеющимся словарем моторных актов, который выполняет функцию регуляции и контроля моторного компонента действия.
Следовательно, полная или частичная замена зрительной информации на слуховую не изменяет ход событий. Даже если она будет в высшей степени специфичной и достигнет высокой стадии обработки, как происходит в нейронах верхней височной извилины, все равно в ней будет отсутствовать то моторное значение, которое может быть сформировано только благодаря двусторонним связям между нейронами F5 и системы PF—PFG, где воспринятые при помощи зрения или слуха действия других приобретают специфическое значение или становятся целенаправленными, или интенциональными, моторными актами.
Таким образом, моторное знание о наших собственных действиях есть необходимое и достаточное условие незамедлительного понимания действий других. Как будет показано далее, это знание фундаментально значимо для построения базового понимания целенаправленности, потому что оно обеспечивает не только отдельные моторные акты, которые мы рассматривали до сих пор, такие как хватание, удержание, разрывание, но также и соединение их в более сложные действия.
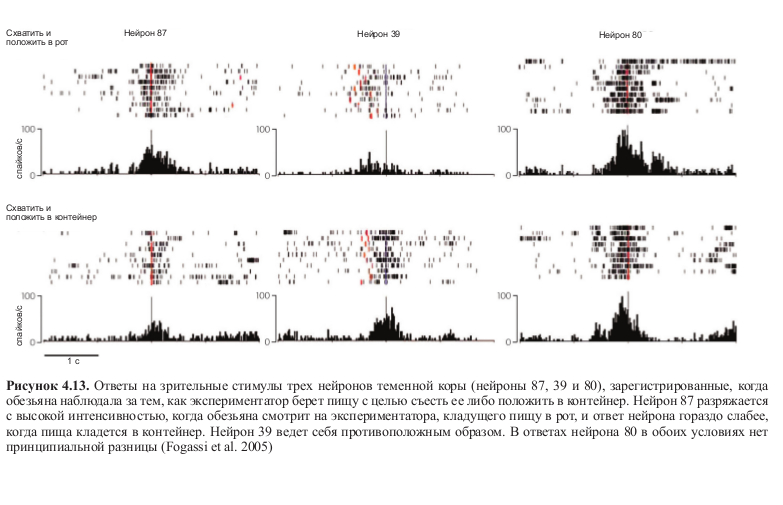
Этот аспект моторной организации был недавно изучен Леонардо Фогасси и его коллегами14, которые регистрировали активность ряда зеркальных нейронов в теменной коре (рис. 4.11 А) при хватании обезьяной объекта. В эксперименте было два условия: в первом обезьяна начинала движение рукой из заранее определенной точки до кусочка пищи, расположенного перед ней, чтобы его схватить и донести до рта. Во втором условии исходное положении руки было таким же, но вместо того, чтобы положить пищу себе в рот, обезьяна клала его в контейнер (рис. 4.11 Б).
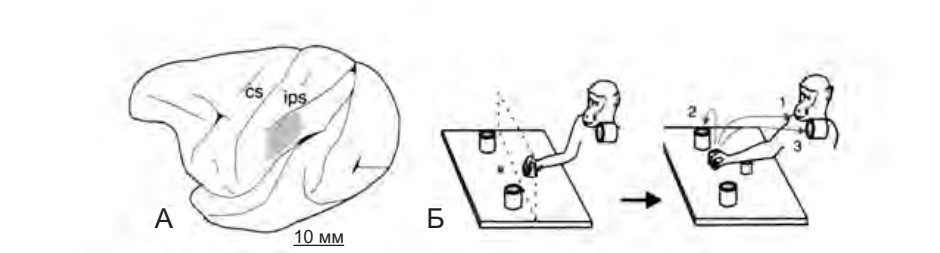
Рисунок 4.11. А — латеральный срез мозга обезьяны. Закрашенный участок — часть нижней теменной коры, активность нейронов которой регистрировалась в эксперименте. Б — условия эксперимента. Контейнер располагался либо рядом с пищей, либо на плече у обезьяны. Так как скорость движения по дотягиванию и захвату пищи определялась последующим моторным актом, при помощи изменении позиции контейнера (и, следовательно, траектории движения) можно было определять, зависит ли избирательность нейронов от особенностей выполнения движения или от конечной цели действия. Результаты эксперимента показали, что в действительности селективность нейронов определяется целью действия (Fogassi et al. 2005).
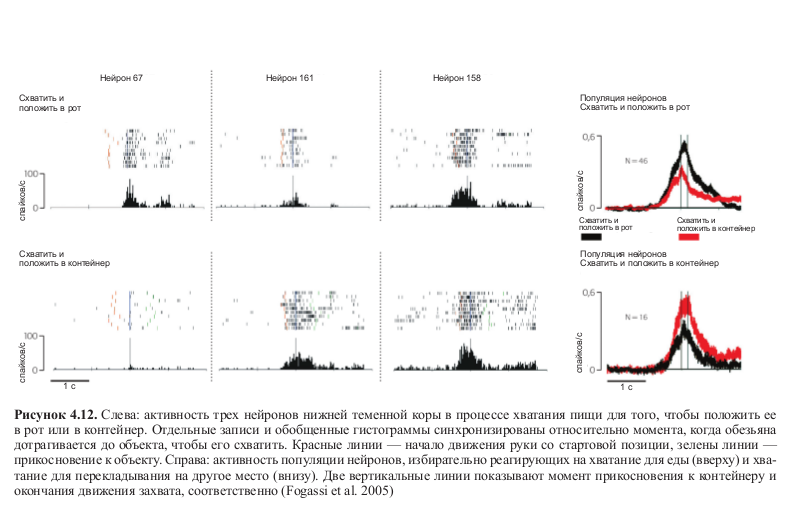
Результаты этих экспериментов показали, что большинство нейронов, активность которых регистрировалась, разряжались по-разному в зависимости от того, какое действие следовало за хватанием пищи — помещение ее в рот или в контейнер (см. табл. 4.1). На рисунке 4.12 (см. цв. вклейку) показана активность трех нейронов: можно увидеть, что два из них разряжаются по-разному в зависимости от того, какое действие следует за хватанием пищи. Третий нейрон кодирует движение хватания независимо от цели действия.
 Была также проведена серия контрольных экспериментов для того, чтобы выяснить, чем еще может быть объяснена избирательность нейронов к определенному действию («положить в рот» и «переместить с одного места на другое») — типом объектов (пища и несъедобные предметы) или силой, с которой обезьяна хватает объект.
Была также проведена серия контрольных экспериментов для того, чтобы выяснить, чем еще может быть объяснена избирательность нейронов к определенному действию («положить в рот» и «переместить с одного места на другое») — типом объектов (пища и несъедобные предметы) или силой, с которой обезьяна хватает объект.
Эти альтернативные интерпретации были отвергнуты по результатам контрольных экспериментов. Кроме того, изучение кинематического рисунка (скорость, ускорение, и т. д.) движений по дотягиванию и хватанию объекта показало, что моторные характеристики выполнения действия не влияют на избирательность нейронов к тем или иным действиям.

Таблица 4.1. Нейроны нижней теменной коры, активность которых изучалась при выполнении действия по захвату пищи для того, чтобы положить ее в рот или в контейнер
Читателю может показаться противоречащим здравому смыслу предположение о том, что моторные нейроны теменной коры настроены на специфические действия. Не слишком ли это расточительно — иметь «нейроны хватания» для каждого конкретного действия? Не было бы более экономным иметь возможность подключать к работе любые из них, когда нам нужно что-нибудь схватить? Ответ на этот вопрос следует искать в фундаментальных свойствах моторной организации: типичным для действий человека и многих животных является плавность действий. «Нейроны хватания» встраиваются в заранее сформированные нейронные цепочки, обеспечивающие выполнение всего действия, таким образом, что каждый нейрон кодирует хватание, но в то же время, будучи включенным в последовательно выполняемый моторный акт, обеспечивает плавность движений.
Но еще более интересным может показаться другой аспект исследования Фогасси и коллег: некоторая форма избирательности, сходная с избирательностью моторной, проявлялась в тот момент, когда обезьяна просто наблюдала за тем, как экспериментатор выполняет ту же последовательность действий. В этом случае нейроны также разряжались по-разному в зависимости от типа действия, в который было встроено кодируемое ими движение. Более того, эти избирательные нейроны также демонстрировали высокую согласованность моторного и зрительного ответов (табл. 4.2, рис. 4.13 и 4.14, см. цв. вклейку).

Таблица 4.2. Нейроны нижней теменной коры, активность которых изучалась при наблюдении за действием по захвату пищи для того, чтобы положить ее в рот или в контейнер
Стоит отметить, что в обоих случаях — когда обезьяна сама выполняет действие и когда она смотрит на выполнение его экспериментатором — нейроны начинают разряжаться, как только рука (обезьяны или экспериментатора) принимает форму, необходимую для захвата пищи или объекта. На самом деле, не так уж и удивительно то, что нейроны начинают кодировать значение целенаправленного действия с самого первого движения. Когда обезьяна протягивает руку по направлению к пище, она уже знает о том, что она собирается делать — донести эту пищу до рта или положить ее в другое место. Даже если намерение животного становится явным только после полного развертывания моторного поведения, оно все равно определяет начальный этап действия. Кроме того, гипотеза об организации моторных актов в специфические моторные цепочки также подтверждается организацией соматосенсорных рецептивных полей нейронов теменной коры. То, что многие теменные нейроны, отвечающие на пассивное сгибание руки, имеют еще и тактильные рецептивные поля, расположенные вокруг рта, свидетельствует о следующем: эти нейроны будут обеспечивать раскрытие рта, если животное будет хватать и поднимать объект15.
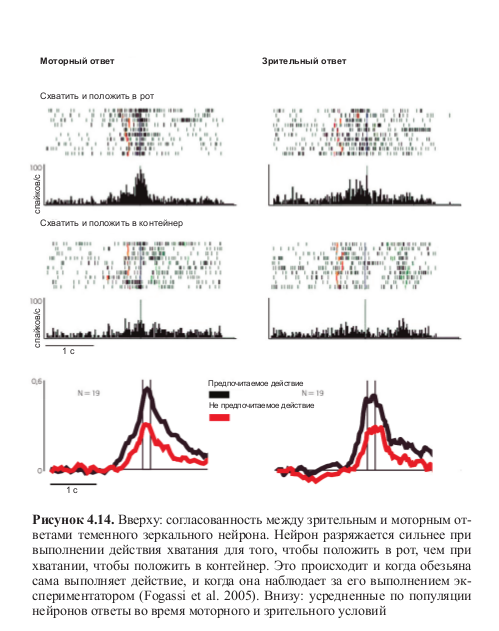 Но эта ситуация меняется, когда обезьяна наблюдает за тем, как экспериментатор берет рукой пищу или другой объект. Однако, тот факт, что зрительный стимул активирует тот же самый нейронный паттерн — то есть, тот же набор потенциальных моторных актов, который отвечает за работу всей моторной цепочки — говорит о том, что обезьяна с самого начала способна ухватить реальную динамику целенаправленных движений, за которыми она наблюдает, и предугадать, чем завершится действие экспериментатора, как только он начнет движение.
Но эта ситуация меняется, когда обезьяна наблюдает за тем, как экспериментатор берет рукой пищу или другой объект. Однако, тот факт, что зрительный стимул активирует тот же самый нейронный паттерн — то есть, тот же набор потенциальных моторных актов, который отвечает за работу всей моторной цепочки — говорит о том, что обезьяна с самого начала способна ухватить реальную динамику целенаправленных движений, за которыми она наблюдает, и предугадать, чем завершится действие экспериментатора, как только он начнет движение.
Правда, в описываемых экспериментах обезьяне были доступны подсказки, позволяющие ей выбрать подходящее значение для действий, которые она видела, но если бы этих подсказок-индикаторов не было, обезьяне бы потребовалась божественная сила! Контейнер был одной из самых важных подсказок: если он стоял на столе, экспериментатор клал пищу в него, если его не было — то себе в рот. Кроме того, подсказки могли взаимодействовать друг с другом: на самом деле, многие нейроны, отвечающие при наблюдении обезьяной за действием по хватанию пищи и поднесению ее ко рту, разряжались, хотя и более слабо, и при наблюдении за действием по перемещению пищи в другое место (однако, так не происходило при работе с объектами, не являющимися пищей).
Это практически выглядело так, как если бы присутствия пищи в поле зрения и возможности протянуть руку и взять ее было бы достаточно для того, чтобы активировать, пусть и слабо, цепочку действий, которая приведет к тому, что пища будет положена в рот — даже если по контексту будет понятно, что результатом действия будет перенесение пищи в другое место. Ответы других нейронов становятся слабее при повторе действия, как если бы активация цепи «хватания для перемещения» постепенно подавляла активацию цепи «хватания для еды».
Все эти аргументы в целом подтверждают важность моторного знания для понимания действий, в то же время расширяя его роли и функции. Такое знание позволяет нам распознавать значения моторных актов, которые мы наблюдаем — и когда они выполняются вне контекста, и когда они являются частью целой цепи действий. В этом последнем случае их значения более не определяются только лишь специфическими связями между объектами, позволяющими отличать одно действие от другого. Хватание теперь не просто хватание, но хватание для того, чтобы съесть или для того, чтобы переместить. Здесь намерение выполнить действие преобладает над выполнением отдельного движения и изменяет его значение в ту или иную сторону.
Если бы моторные цепочки не были организованы так, как продемонстрировали Фогасси и его коллеги, скорее всего, мозг обезьяны практичеcки не мог бы создавать такие инструкции, которые обеспечивали бы плавность движений и их сходство с тем, что Александр Романович Лурия любил называть «кинетическими мелодиями»18. Более того, без зеркальных свойств этих нейронов обезьяна была бы не способна ни мгновенно уловить, так сказать, намерения, которые наполняли эти мелодии содержанием, когда они проигрывались другими, ни смогла бы также предугадать с самых первых движений промежуточный результат действия (то есть, захват пищи рукой) и, что более важно, его конечный итог — хватание для того, чтобы съесть или чтобы переместить.
Чем более точная информация может быть получена из контекста и от самого объекта, тем более избирательной будет активация соответствующей моторной цепи. Однако, даже если сенсорная информация неопределенна, какой она часто и бывает (и не только в экспериментальных условиях), активация одного или нескольких целенаправленных потенциальных моторных актов помогает обезьяне расшифровать намерения экспериментатора.
После этого животное становится способно выбирать ту интенцию, которая ему кажется наиболее соответствующей сценарию действия, вплоть до распознавания самой подходящей, и использует ее, ничего не говоря о том, что эти процессы расшифровки и идентификации были связаны с тем же моторным знанием, которое направляет и приспосабливает выполнение животным той же самой цепочки действий.
5. Зеркальные нейроны у человека
Первые данные
После обнаружения зеркальных нейронов у обезьян логично было сделать следующий шаг и предположить наличие подобной системы и у человека. Часто (и не только в нейрофизиологии) новые открытия способствуют пересмотру и новой интерпретации старых данных, уже имеющихся в литературе, что и случилось с идеей зеркальных нейронов. Доказательства (часто косвенные), подтверждающие наличие механизма, который мы сейчас интерпретируем как зеркальную систему, были найдены при помощи регистрации ЭЭГ (электроэнцефалограммы) в начале 50-х годов, когда были проведены исследования реактивности ритмов мозга при наблюдении за движением.
Хорошо известно, что при помощи ЭЭГ можно зарегистрировать спонтанную электрическую активность мозга. Ритмы ЭЭГ классифицируются по их частоте. У взрослых здоровых людей в спокойном состоянии при закрытых глазах в задних отделах мозга доминирует альфа-ритм (8—12 Гц), а в передних — так называемые ритмы десинхронизации (имеющие высокую частоту и низкую амплитуду). Еще один ритм, известный как мю-ритм и сходный с альфа-ритмом, регистрируется в центральных областях. Альфа-ритм преобладает при бездействии сенсорных систем, в частности, зрительной. Если зрительные условия меняются и закрытые глаза открываются, этот ритм исчезает или значительно ослабляется. Мю-ритм также отчетливо выражен, когда моторная система находится в покое, а активное движение или соматосенсорная стимуляция десинхронизируют его.
В 1954 году Анри Гасто16 и его коллеги провели эксперименты, в которых показали, что мю-ритм десинхронизируется не только при выполнении действий, но и при наблюдении за действиями других. Спустя сорок с лишним лет, вдохновленные открытием зеркальных нейронов, Вилейанур С. Рамачандран и его коллеги, а также Стефани Кочин и ее коллеги17 воспроизвели эти эксперименты, используя усовершенствованные методы. В частности, группа Кочин показала, что наблюдение за движениями ноги или пальцев сопровождается десинхронизацией мю-ритма, и что этого не происходит, если испытуемым демонстрируется движущийся объект. Другими словами, тот же самый ритм, который блокируется или десинхронизируется движением, также подавляется при наблюдении за движением другого человека.
Похожие результаты были получены в серии исследований, проведенных при помощи МЭГ (магнитоэнцефалографии) — метода, который позволяет анализировать электромагнитную активность мозга, используя регистрацию генерируемых мозгом магнитных полей. Результаты этих исследований также показали, что мю-ритм в прецентральной коре подавляется как при манипуляции с объектами, так и при наблюдении действий других18.
Исследования с использованием транскраниальной магнитной стимуляции (ТМС) также обеспечивают очень убедительные доводы в пользу того, что моторная система человека обладает зеркальной функцией. ТМС — это неинвазивный метод, позволяющий стимулировать мозг. Вплотную к голове подносится электромагнитная катушка, образующееся вокруг которой магнитное поле вызывает индуцированные токи определенной интенсивности в моторной коре. Это позволяет регистрировать моторные потенциалы (двигательные вызванные потенциалы, или ДВП) в мышцах контралатеральной конечности.
Так как амплитуда ДВП модулируется контекстом, в котором выполняется действие, этот метод может быть использован для контроля возбудимости моторной системы в различных экспериментальных условиях. Лучано Фадига и его коллеги19 регистрировали ДВП, вызванные стимуляцией левой моторной коры, в различных мышцах правой руки и кисти испытуемых, которых просили смотреть на то, как экспериментатор берет рукой различные объекты и выполняет движения, не имеющие смысла и никак не связанные с объектом. В обоих случаях было обнаружено избирательное увеличение амплитуд ДВП в мышцах при наблюдении за действием. Несмотря на то, что увеличение амплитуд ДВП во время наблюдения за транзитивными дейcтвиями (связанными с объектами) согласуется с данными, полученными на обезьянах, было весьма неожиданным обнаружить подобный эффект и во время наблюдения за нетранзитивными действиями (не направленными на объект), так как у обезьян зеркальные нейроны не отвечали на наблюдение за не связанными с объектами движениями руки.
Это не единственное различие между зеркальными системами человека и обезьяны. При регистрации ДВП в мышцах руки у здоровых испытуемых, наблюдавших за тем, как экспериментатор выполняет типичные движения захвата, было показано, что активация моторной коры по времени очень точно совпадает с временем выполнения наблюдаемого действия. Судя по этим данным, можно предположить, что зеркальные нейроны у человека способны кодировать не только цель действия, но и временные характеристики отдельных входящих в него движений20.
Данные томографических исследований
Важные функциональные следствия описанных выше результатов будут рассмотрены позднее, сейчас же необходимо представить еще один источник данных, благодаря которому были получены подтверждения существования у человека зеркальной системы — это методы картирования мозга.
Несмотря на то, что электрофизиологические методы, такие как ЭЭГ, МЭГ и ТМС, могут быть использованы для регистрации специфичеcкой активации моторной системы человека, вызванной наблюдением за действиями других, они не позволяют точно локализовать корковые зоны и системы, обеспечивающие такую активность. Следовательно, с их помощью невозможно выяснить строение зеркальной системы в целом. Поэтому необходимо обратиться к методам картирования мозга, в частности, позитронно-эмиссионной томографии (ПЭТ) и функциональной магнитно-резонансной томографии (фМРТ), благодаря которым можно регистрировать вызванные выполнением и наблюдением за определенными моторными актами изменения кровотока в тех или иных областях мозга.
Однако, результаты первых таких исследований были далеки от ожидавшихся21. Участникам эксперимента предъявлялись изображения хватательных движений при помощи руки, созданной в виртуальной реальности. Методом ПЭТ не удалось зарегистрировать какую-либо значимую активацию в моторных зонах, которые соответствуют вентральной премоторной коре у обезьян. Эффекты, обнаруженные в описанных ранее электрофизиологических исследованиях, пока не могли быть объяснены.
Позднее этот эксперимент был воспроизведен другими исследователями22, но с одним важным отличием: движения выполнялись настоящей рукой, а не виртуальной, которая явно была нереалистичной. На этот раз результаты ПЭТ подтвердили то, что было найдено у обезьян: существуют зоны в лобной коре, которые активируются при наблюдении за движениями руки другого. Затем и в фМРТ-исследованиях были более точно локализованы зоны, включенные в зеркальную систему: во время наблюдения за действиями других постоянно активируются ростральная (передняя) часть нижнетеменной коры и нижняя часть прецентральной извилины, а также задняя часть нижнелобной извилины. В определенных экспериментальных условиях также активируются более передняя область нижнелобной извилины и дорзальная премоторная кора (рис. 5.1, см. цв. вклейку).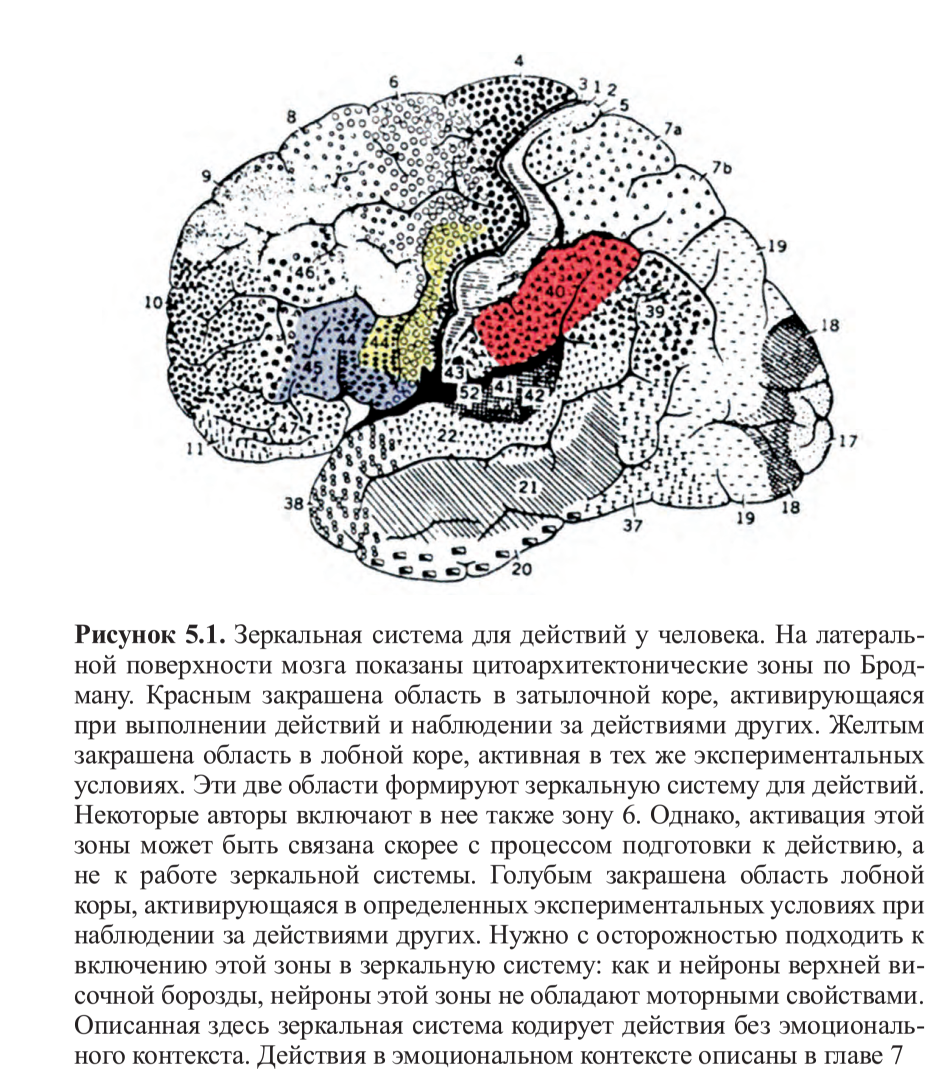 Несмотря на то, что интерпретировать данные о корковой активности, полученные при помощи фМРТ, в терминах цитоархитектонических зон всегда рискованно, очень высока вероятность того, что активирующийся участок нижнетеменной коры соответствует зоне 40 по Бродману — гомологу зоны PF у человека, которая, как мы уже видели ранее, является одной из тех зон, где были найдены зеркальные нейроны у обезьян. Но активацию в нижней части прецентральной извилины и задней части нижнелобной извилины оказалось гораздо сложнее описать цитоархитектонически. Долгие годы считалось, что это две разные зоны, не имеющие никакой функциональной взаимосвязи: задняя часть нижнелобной извилины соответствует зоне 44 по Бродману (то есть, задней части зоны Брока) и отвечает за речь, тогда как нижняя часть прецентральной извилины рассматривается как относящаяся к моторной коре.
Несмотря на то, что интерпретировать данные о корковой активности, полученные при помощи фМРТ, в терминах цитоархитектонических зон всегда рискованно, очень высока вероятность того, что активирующийся участок нижнетеменной коры соответствует зоне 40 по Бродману — гомологу зоны PF у человека, которая, как мы уже видели ранее, является одной из тех зон, где были найдены зеркальные нейроны у обезьян. Но активацию в нижней части прецентральной извилины и задней части нижнелобной извилины оказалось гораздо сложнее описать цитоархитектонически. Долгие годы считалось, что это две разные зоны, не имеющие никакой функциональной взаимосвязи: задняя часть нижнелобной извилины соответствует зоне 44 по Бродману (то есть, задней части зоны Брока) и отвечает за речь, тогда как нижняя часть прецентральной извилины рассматривается как относящаяся к моторной коре.
Нельзя не упомянуть здесь о том, что в самом начале XX века Альфред Уолтер Кэмпбелл, один из отцов цитоархитектоники, обратил внимание на сходство в анатомическом строении задней части нижневисочной извилины и нижней части прецентральной извилины, назвав эту область термином собственного изобретения — «промежуточная прецентральная кора»23. На его наблюдение, однако, никто не обращал внимания в течение многих лет. И только недавние сравнительно-анатомические исследования показали, что зона 44 по Бродману (по крайней мере, ее часть) может рассматриваться как гомолог у человека зоны F5у обезьян24; более того, стало ясно, что в зоне 44 имеются репрезентации руки и рта25.
Может ли теперь, на основании приведенных данных, этой зоне быть приписана ключевая роль в зеркальной нейронной системе человека? Можно ли действительно использовать функциональную гомологичность с зоной F5 у обезьян в качестве ключевого аргумента в пользу того, чтобы интерпретировать полученную в томографических исследованиях активацию как наличие у нейронов задней части нижнелобной извилины свойств, аналогичных зеркальным? Не будет ли проще предположить, что их активация отражает «внутреннюю вербальную репрезентацию»? Мы часто описываем в уме действия, которые наблюдаем — например, мы можем бормотать себе под нос:
«Только вдумайтесь в это — вот, оказывается, как я беру свою чашку кофе!»
Итак, можем ли мы действительно исключать возможность того, что нечто подобное происходит и с испытуемыми в ПЭТ-исследованиях? В конце концов, активируется часть зоны Брока! Джованни Буччино и его коллеги26 попытались ответить на эти возражения, проведя фМРТ-исследование. Они просили нескольких студентов просматривать видеоролики, в которых актеры либо выполняли транзитивные действия, такие как откусывание яблока, поднимание чашки кофе, удар по футбольному мячу, либо изображали их. Наблюдение за транзитивными движениями рта выявило два фокуса активации в лобной коре: один соответствовал задней части нижнелобной извилины, а другой — нижней части прецентральной, и два — в нижнетеменной коре, а наблюдение за транзитивными движениями руки выявило такой же паттерн активации, за исключением того, что активация в нижней части прецентральной извилины была локализована более дорзально, а в ростральной части нижнетеменной коры — более сдвинута назад.
Транзитивные движения ног вызывали фронтальную активацию, расположенную более дорзально по сравнению с активацией на движения руки и рта, и еще больший сдвиг назад фокуса активации в теменной коре. Другими словами, несмотря на значительное перекрытие, зеркальная система, похоже, организована соматотопически и имеет отдельные области для движений рта, рук и ног. При наблюдении за изображением движений был получен аналогичный паттерн активации, ограниченный, однако, только лобной корой (рис. 5.2, см. цв. вклейку).
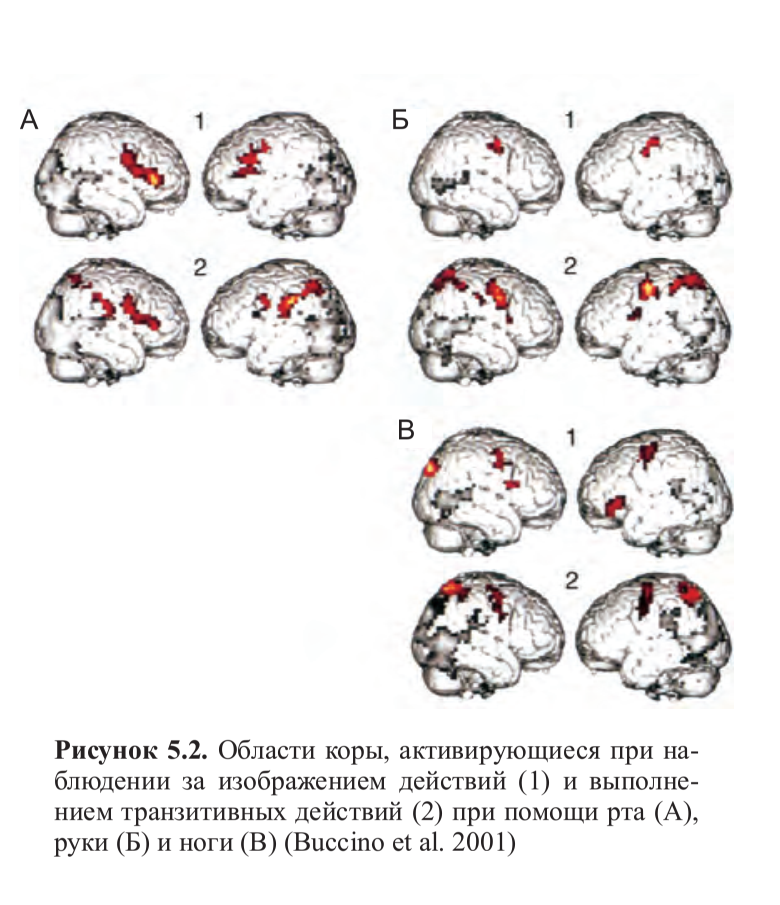 Если бы гипотеза о вербальном опосредовании была верна, зона Брока должна была бы активироваться независимо от типа наблюдаемых действий и использованного органа-эффектора, а также не было бы никакой активации премоторной коры. Однако, описанные выше результаты не подтверждают эту гипотезу, и поэтому, если мы не хотим и далее придерживаться довольно причудливого объяснения ad hoc — настаивая, например, на том, что вербальная репрезентация должна присутствовать при наблюдении за движениями рта и руки, но каким-то волшебным образом должна исчезнуть, когда мы смотрим на движения ног, — мы вынуждены признать, что активация зоны Брока отражает типичное поведение зеркальных нейронов. Более того, эксперимент Буччино и коллег показал, что в зеркальную систему человека помимо зоны Брока включены большие области премоторной и нижнетеменной коры. Это еще одно доказательство в пользу того, что зеркальная система отвечает не только на движения руки и транзитивные действия, но и на изображение действий.
Если бы гипотеза о вербальном опосредовании была верна, зона Брока должна была бы активироваться независимо от типа наблюдаемых действий и использованного органа-эффектора, а также не было бы никакой активации премоторной коры. Однако, описанные выше результаты не подтверждают эту гипотезу, и поэтому, если мы не хотим и далее придерживаться довольно причудливого объяснения ad hoc — настаивая, например, на том, что вербальная репрезентация должна присутствовать при наблюдении за движениями рта и руки, но каким-то волшебным образом должна исчезнуть, когда мы смотрим на движения ног, — мы вынуждены признать, что активация зоны Брока отражает типичное поведение зеркальных нейронов. Более того, эксперимент Буччино и коллег показал, что в зеркальную систему человека помимо зоны Брока включены большие области премоторной и нижнетеменной коры. Это еще одно доказательство в пользу того, что зеркальная система отвечает не только на движения руки и транзитивные действия, но и на изображение действий.
Понимание намерений других при помощи зеркальной системы
Мы увидели, что и электрофизиологические, и томографические исследования показали наличие у человека системы, аналогичной зеркальной системе у обезьян. Но между ними есть и существенные различия: у человека зеркальная система занимает большую площадь коры, чем у обезьян. Однако к этому выводу нужно подходить с особой осторожностью и с учетом различий в экспериментальных методах, использованных в исследованиях с обезьянами и человеком: одно дело — регистрировать активность отдельных нейронов, и совсем другое — анализировать активацию различных корковых зон на основании изменений кровотока. Тем не менее, наиболее важно то, что зеркальная система человека обладает некоторыми свойствами, которых не было найдено у обезьян — к примеру, она кодирует как транзитивные, так и нетранзитивные моторные акты, как цели моторных актов, так и движения, из которых состоит законченное действие, и, наконец, в случае с транзитивными действиями, эффективное взаимодействие с объектом не является обязательным условием, так как для активации зеркальной системы достаточно и изображения действия.
Мы уже упоминали, что эти свойства могут иметь важное функциональное значение, однако, тот факт, что зеркальная система человека может выполнять более широкий спектр задач, чем тот, который наблюдался у обезьян, не должен затмевать основную функцию зеркальной системы, связанную с пониманием значения действий других. На самом деле, эксперименты с ТМС показали, что наблюдения за действиями, выполняемыми другими при помощи рук, приводят к увеличению амплитуд ДВП, регистрируемых в мышцах соответтвующей руки наблюдателя, которая задействована в выполнении тех же дейcтвий. С другой стороны, томографические исследования показали, что активация лобной коры при наблюдении за выполнением другими действий при помощи рук, рта и ног определяет активацию, соответствующую соматотопическим моторным репрезентациям этих частей тела.
Как и у обезьян, у людей наблюдение за выполнением действий другими приводит к незамедлительной активации моторных зон, связанных с организацией и выполнением этих действий, и благодаря этой активации становится возможным расшифровать значение наблюдаемых «моторных событий», то есть, понять их в терминах целенаправленных движений. Это понимание полностью лишено каких-либо рефлексивных, концептуальных или лингвистических опосредствований и основано только на словаре действий и моторном знании, от которого зависит наша способность действовать. Наконец, как и у обезьян, это понимание не ограничено отдельными моторными актами, но может распространяться на целые их цепочки.
Это последнее их свойство очень явно проявляется в фМРТ-исследованиях, проведенных Марко Якобони и его коллегами27. Они показывали добровольцам три различных видеоклипа (рис. 5.3). На первом клипе участники эксперимента видели ряд объектов (чайник, кружку, стакан, тарелку, и т. д.), которые лежали на столе так, как будто кто- то собирался пить чай (а не кофе из нашей привычной чашки!) или только что его пил. Экспериментальное условие, в котором предъявлялось это видео, для краткости было названо «контекст». На втором видеоклипе была показана рука, захватывающая кружку при помощи всей кисти либо точного захвата при отсутствии контекста (условие «действие»). На третьем клипе была та же самая рука, согнутая тем же самым образом, но на этот раз в контексте, который позволил бы предположить, что у действующего есть намерение взять чашку со стола либо для того, чтобы поднести ее к губам, либо чтобы убрать ее со стола после чаепития (условие было названо «намерение»).

Рисунок 5.3. Видеоклипы, использованные в исследовании корковых зон, включенных в понимание намерений других. Клипы, соответствующие разным экспериментальным условиям, расположены в три колонки: контекст, действие, намерение. В условии контекст (первая колонка) участники эксперимента видели стол, накрытый для завтрака (вверху) и тот же стол после завтрака (внизу). В условии действие (вторая колонка) они видели руку, которая брала чашку со стола, используя при этом либо захват всей кистью (вверху), либо точный захват (внизу). Контекста в этом условии не было.
В третьем условии (намерение) были показаны два типа захвата в контексте «до» и «после» завтрака, предполагая, соответственно, намерения действующего «взять кружку, чтобы пить из нее» (вверху) и «взять кружку, чтобы помыть ее» (внизу) (Iacoboni et al. 2005).
Если мы сравним активность мозга, вызванную наблюдением действий в трех различных условиях, с контрольным условием отдых (рис. 5.4, см. цв. вклейку), мы увидим, что в условиях действие и намерение активность увеличивалась в зрительных зонах и в лобнотеменной системе, связанной с кодированием моторных актов, тогда как в условии контекст не было увеличения активности ни в области STS (верхняя височная борозда), отвечающей на движение зрительных стимулов, ни в нижнетеменной коре, хотя активность в премоторных зонах была выражена отчетливо. Это можно объяснить присутствием объектов, которые можно схватить и которые активируют канонические нейроны, отвечающие, как мы видели, на возможности, предоставляемые объектами.
Наибольший интерес представляет сравнение условий намерение и действие, а также намерение и контекст. Как можно увидеть на рисунке 5.5 (см. цв. вклейку), активация дорзальной поверхности задней части правой нижней лобной извилины (рис. 5.5, вверху) была выше в условии намерение, чем в двух других условиях (действие и контекст). Это особенно интересно, потому что фокус активации расположен в лобной части зеркальной системы — это говорит о том, что зеркальные нейроны не только кодируют наблюдаемое действие (в данном случае, хватание чашки при помощи определенного типа захвата), но также и намерение, с которым данное действие выполняется. Это может быть объяснено тем, что в то время как наблюдатель смотрит, как другой выполняет действие, он уже предугадывает возможные следующие действия в цепочке (к примеру, «взять, чтобы выпить чаю» или «взять, чтобы переставить»).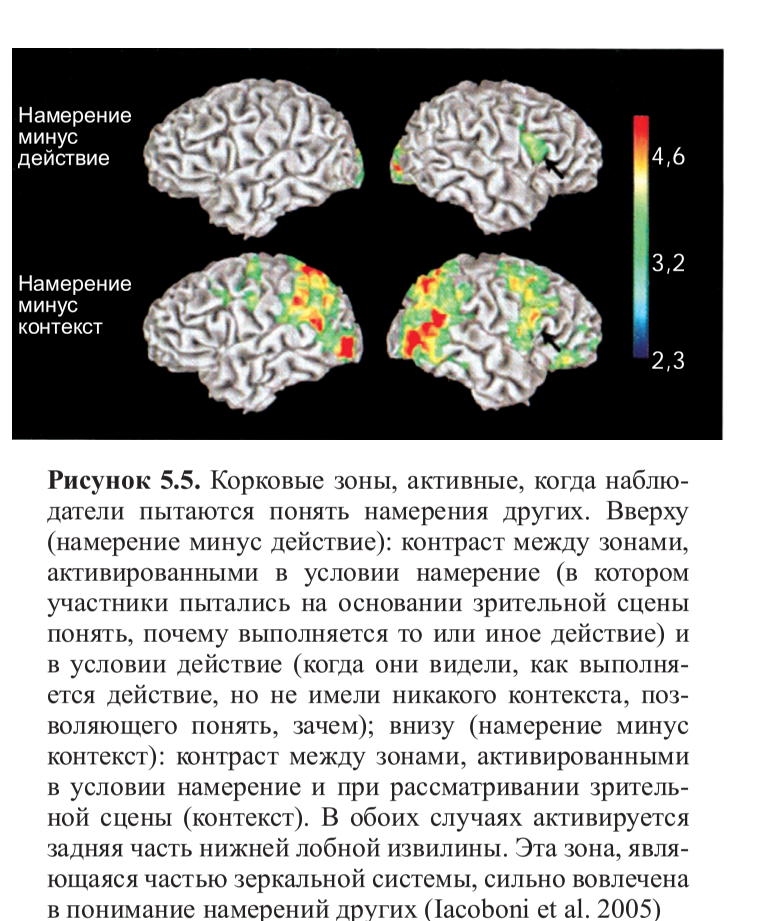 Интересно отметить, что наблюдение за действием поднесения ко рту вызывает более сильную активацию в нейронах зеркальной системы, чем наблюдение за захватом чашки, чтобы убрать ее со стола (рис. 5.6). Это согласуется с данными, полученными Фогасси и коллегами и описанными в предыдущей главе. В их эксперименте было показано, что количество нейронов, кодирующих действие «захвата для еды», больше, чем кодирующих «захват для перекладывания». Кроме того, даже когда сенсорная информация относительно контекста (присутствие контейнера для пищи) предполагает, что наиболее вероятным действием после захвата пищи будет перекладывание ее в контейнер, наблюдение за рукой экспериментатора, берущей пищу, вызывает активность, пусть и достаточно слабую, в той цепочке нейронов, которая отвечает за кодирование «хватания для еды», так как это естественная последовательность действий в репертуаре обезьяны. В исследовании Якобони были получены похожие результаты: активация правой нижнелобной коры была больше при наблюдении за действием «поднести кружку ко рту, чтобы пить из нее», чем в случае «хватания кружки, чтобы помыть ее». Всегда преобладает именно самое естественное двигательное намерение, наиболее глубоко укоренившееся в нашем словаре действий.
Интересно отметить, что наблюдение за действием поднесения ко рту вызывает более сильную активацию в нейронах зеркальной системы, чем наблюдение за захватом чашки, чтобы убрать ее со стола (рис. 5.6). Это согласуется с данными, полученными Фогасси и коллегами и описанными в предыдущей главе. В их эксперименте было показано, что количество нейронов, кодирующих действие «захвата для еды», больше, чем кодирующих «захват для перекладывания». Кроме того, даже когда сенсорная информация относительно контекста (присутствие контейнера для пищи) предполагает, что наиболее вероятным действием после захвата пищи будет перекладывание ее в контейнер, наблюдение за рукой экспериментатора, берущей пищу, вызывает активность, пусть и достаточно слабую, в той цепочке нейронов, которая отвечает за кодирование «хватания для еды», так как это естественная последовательность действий в репертуаре обезьяны. В исследовании Якобони были получены похожие результаты: активация правой нижнелобной коры была больше при наблюдении за действием «поднести кружку ко рту, чтобы пить из нее», чем в случае «хватания кружки, чтобы помыть ее». Всегда преобладает именно самое естественное двигательное намерение, наиболее глубоко укоренившееся в нашем словаре действий.
Рисунок 5.6. Временная развертка активности правой нижнелобной коры демонстрирует увеличение сигнала при сопоставлении условий (контрасты намерение минус действие и намерение минус контекст). Как показано на предыдущем рисунке, это зона, активирующаяся, когда наблюдатель «прочитывает» намерения других. Интересно отметить, что наблюдение действия «взять, чтобы выпить чаю» порождает более сильную активацию зеркальной системы, чем за действием «взять, чтобы помыть» (Iacoboni et al. 2005)
Не стоит и говорить, что, как и в случае с моторными актами, активация зеркальных нейронов — не единственное средство, благодаря которому мы способны понимать намерения, заложенные в действиях других. Каждый день мы приписываем другим — более или менее осознанно — убеждения, желания, ожидания, намерения, и т. д. Наше собственное социальное поведение в основном зависит от нашей способности понимать, что другие имеют в виду, и руководствуясь этим, мы решаем, как нам подстроиться под собеседника.
В настоящее время не найден еще такой нейронный механизм, который объяснил бы чтение мыслей: может случиться так, что он окажется эволюционно связанным с зеркальной системой. Что, однако, имеет значение — то, что зеркальный механизм фиксирует интенциональный аспект действий, общий для наблюдателя и исполнителя. Здесь будет очень к месту цитата из Мерло-Понти:
“Смысл жестов не дан, он понимается, то есть улавливается посредством действия наблюдателя. Вся трудность в том, чтобы правильно понять это действие и не спутать его с познавательной операцией. Общение, или понимание жестов, достигается во взаимности моих интенций и жестов другого, моих жестов и интенций, читающихся в поведении другого. Все происходит так, как если бы интенции другого населяли мое тело, а мои интенции населяли тело другого. Жест, свидетелем которого я являюсь, «очерчивает пунктиром» интенциональный объект. Этот объект становится актуальным и полностью понимается, когда способности моего тела приспосабливаются к нему и его охватывают28.
«Действия наблюдателя» — это потенциальный моторный акт, определяемый активацией зеркальных нейронов, кодирующих сенсорную информацию в моторных терминах, таким образом обеспечивая «реципрокность» действий и намерений, лежащую в основе нашей способности мгновенно понимать, что делают другие. Понимание намерений других не основано, в таком случае, на осмыслении, то есть, на метарепрезентации, но зависит от выбора наиболее подходящей к данной ситуации возможной цепочки действий.
Как только мы видим, как кто-то делает что-либо — отдельное действие или цепочку действий — его движения незамедлительно приобретают для нас смысл, хочет он этого или нет. И, очевидно, верно и обратное: наши действия незамедлительно обретают смысл для тех, кто их видит. Зеркальная система и такая избирательность ответов составляющих ее нейронов способствуют созданию общего пространства действий (shared space of actions), внутри которого каждое действие и цепочка действий, принадлежат они «нам» или «другим», мгновенно регистрируются и распознаются без необходимости какой-либо эксплицитной или намеренной «когнитивной операции».
Различия в словарях
Что произойдет, если окажется, что в нашем словаре действий нет того движения, которое мы наблюдаем? В экспериментах, рассмотренных в предыдущей главе, мы видели, что у обезьян зеркальные нейроны разряжаются не только когда другая обезьяна хватает пищу, но также когда действие выполняет экспериментатор. Это не удивительно, если вспомнить, что хватание присутствует в словаре действий обезьяны (равно как и удержание, разрывание, толкание, бросание, и т. д.). И тем не менее, мы часто видим действия, не являющиеся частью нашего моторного знания — возможно, они не входят в репертуар нашего вида, или же просто мы не способны их выполнять.

Рисунок 5.7. Стимульный материал, использованный в эксперименте, целью которого была регистрация активности мозга при наблюдении за действиями, общими для человека и животных. Кадры из видеоклипов показывают, как человек, обезьяна и собака откусывают кусочки пищи (Buccino et al. 2004a)
В одном недавнем исследовании с использованием фМРТ29 группе добровольцев предъявлялись видеоклипы без звука, в которых представители разных биологических видов (человек, обезьяна, собака) выполняли действия, связанные с употреблением пищи (откусывание) или коммуникативные акты (говорение, чмокание губами, лай).

Рисунок 5.8. Стимульный материал, использованный в эксперименте, в котором сравнивалось восприятие коммуникативных жестов человека и представителей других видов. Кадры из видеоклипов показывают человека, обезьяну и собаку в процессе говорения, чмокания губами и лая, соответственно (Buccino et al. 2004a)
На рисунках 5.7 и 5.8 представлены отдельные кадры из этих клипов. Несмотря на то, что вид человека, откусывающего кусочек пищи, сильно отличается от вида обезьяны или (еще больше) от вида собаки, было обнаружено значительное перекрытие зон активности во всех трех случаях. На самом деле, просмотр всех трех видеоклипов порождал два фокуса активности (передний и задний) в нижнетеменной коре, а также в задней части нижней лобной извилины и примыкающей к ней прецентральной извилины. При сравнении трех условий наблюдалась определенная асимметрия между левым и правым полушариями: активация в левом полушарии практически не зависела от того, кем выполнялось действие — человеком, обезьяной или собакой, тогда как в правом полушарии, напротив, активация была сильнее при наблюдении за выполняющим действия человеком (рис. 5.9, см. цв. вклейку).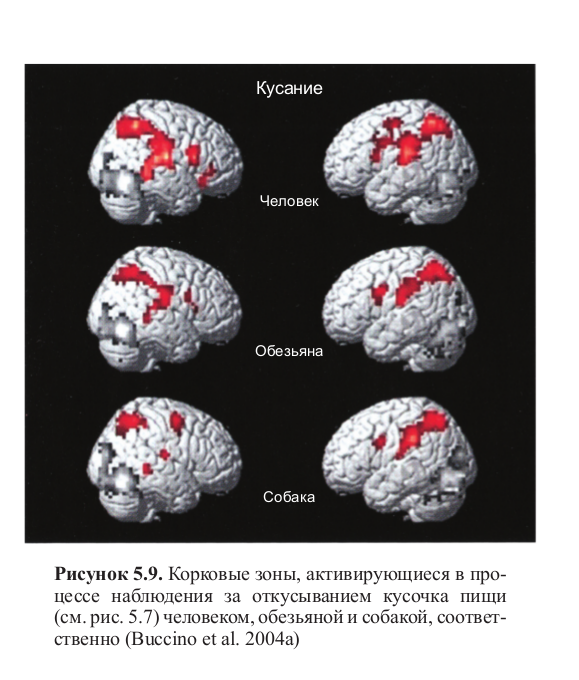
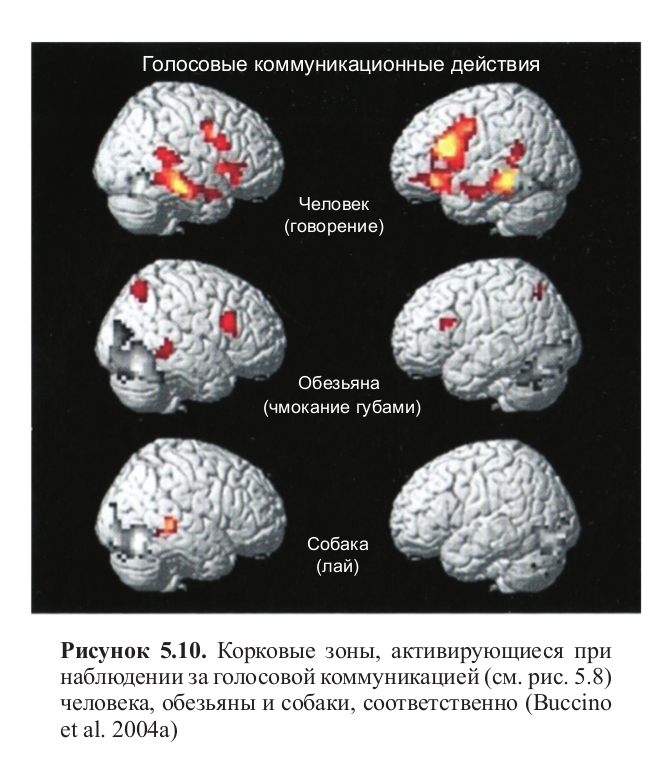 При наблюдении за коммуникативными действиями паттерн активации был совершенно иным. Наблюдение за человеком, двигающим губами так, как будто он разговаривает, вызывало сильную активацию в задней части нижней лобной извилины (область, соответствующая зоне Брока); активация становилась слабее, когда испытуемые наблюдали за чмокающей губами обезьяной, и исчезала совсем, когда они видели лающую собаку (рис. 5.10, см. цв. вклейку).
При наблюдении за коммуникативными действиями паттерн активации был совершенно иным. Наблюдение за человеком, двигающим губами так, как будто он разговаривает, вызывало сильную активацию в задней части нижней лобной извилины (область, соответствующая зоне Брока); активация становилась слабее, когда испытуемые наблюдали за чмокающей губами обезьяной, и исчезала совсем, когда они видели лающую собаку (рис. 5.10, см. цв. вклейку).
В терминах чисто зрительного восприятия различие между коммуникативными актами не окажется большим, чем между действием по откусыванию пищи, выполняемым тремя представителями разных видов, и, следовательно, не сможет объяснить полученные паттерны активации. Отсутствие активности в зеркальной системе при восприятии лающей собаки не может быть просто объяснено типом получаемой зрительной информации. В предыдущей главе мы увидели, что активность зеркальных нейронов не привязана к какой-либо одной модальности — она ограничена словарем действий, регулирующим организацию и выполнение движений. Лай просто не принадлежит к словарю действий человека.
Означает ли это, что мы неспособны понимать движения лающей собаки и отличать их от того, как она откусывает кусочек пищи? Конечно, нет! Просто восприятие этих двух процессов зависит от двух совершенно различных аспектов понимания: первый преимущественно основывается на зрительной информации, а второй — на зрительно-моторной. Когда мы видим лающую собаку, наше понимание ее действий связано преимущественно с активацией зон верхней височной борозды (STS). Эти, а также другие зрительные зоны активируются при наблюдении за коммуникативными действиями других (людей или обезьян), но в этом последнем случае информация, поступающая из STS, запускает потенциальный моторный акт, кодирующийся в зеркальной системе, благодаря чему мы незамедлительно понимаем значение воспринимаемых нами действий30.
Подобные различия были найдены и между индивидами одного вида. Беатрис Кальво-Мерино и ее коллеги31 провели фМРТ-исследование, в котором было показано, что корковая активность может меняться в зависимости от наличия специфических моторных навыков наблюдателей за выполнением этих специфических действий другими. Испытуемыми были классические танцовщики, тренеры капоэйры и люди, никогда не занимавшиеся танцами. Им показывали видеоклипы урока капоэйры и классических балетных па. Зеркальные системы у тренеров активировались сильнее при просмотре записи капоэйры, а зеркальные системы танцовщиков — при просмотре балетных па, в отличие от тренеров и, тем более, новичков.
В дальнейших экспериментах эти авторы попытались понять причину таких различий — было ли это то, что у тренеров капоэйры имелся больший по сравнению с танцовщиками и, разумеется, новичками, зрительный опыт в просматривании шагов этого боевого и танцевального искусства в дополнение к знанию о том, как их нужно выполнять. Некоторые элементы в капоэйре могут быть выполнены как женщинами, так и мужчинами, а другие отличаются у разных полов. Очевидно, что все танцоры, и женщины, и мужчины, должны знать элементы, которые выполняет их партнер. С учетом этого факта, Кальво-Мерино и ее коллеги показали тренерам капоэйры видео, в котором женщины и мужчины выполняли различные элементы. Они обнаружили, что зеркальная система сильнее активировалась в ответ на наблюдение за элементами, выполнявшимися танцором того же пола, что и испытуемый, и, таким образом, в этом случае активация регулировалась только моторным опытом, а не зрительным.
Рассмотренные вместе, эти два эксперимента подтверждают решающую роль, которую моторное знание играет в понимании значений действий других. Мы не хотим сказать, что эти действия не могут быть поняты при помощи других средств — интеллектуальных процессов, основанных на более или менее сложной обработке сенсорной информации в целом и зрительной в частности, но это означает, что между этими двумя модальностями существуют значительные различия. Только в последнем случае наблюдаемое действие влечет за собой включение наблюдателя, как если бы он выполнял его сам, и позволяет мгновенно уловить его значение.
Расширение и достижение этого «как если бы» зависит от двигательного репертуара наблюдателя, либо его собственного, либо определенного биологическим видом. Лео Спербер (сын хорошо известного Дэна Спербера) прекрасно выразил эту идею, сказав однажды:
«Папа, ты знаешь, почему я не хотел бы быть собакой? Я не знал бы, как вилять хвостом!»32
Как только были открыты зеркальные нейроны, возник вопрос, не являются ли они нейронной основой способности к подражанию.
Однако прежде чем рассмотреть эту возможность, мы должны дать точное определение подражания. Всякий, кому доводилось прикоснуться к данной проблематике, знает, что со временем это понятие обросло в разных исследовательских областях (психологии развития, сравнительной психологии, этологии и т. д.) самыми разнообразными, часто противоречащими друг другу значениями. Исходя из целей, преследуемых нами в этой книге, мы сузим спектр возможных определений до двух, невзирая на риск чрезмерного упрощения. В первом из определений, используемом преимущественно в экспериментальной психологии, подражание предстает как способность индивида к повторению действия, которое уже входит в состав его двигательного репертуара, в результате наблюдения за тем, как это действие выполняют другие33; во втором, распространенном главным образом среди этологов, подражание понимается как процесс, посредством которого индивид научается выполнению нового действия через наблюдение, после чего он способен воспроизвести это действие во всех подробностях34.
Оба определения поднимают, хотя и по-разному, целый ряд вопросов, ответ на которые должна давать любая теория подражания, вне зависимости от того, какое из определений будет принято в ней за основу. Прежде всего, это вопросы, связанные с так называемой проблемой соответствия: как мы делаем что-то, что видели в исполнении других? Иными словами, как мы можем выполнить действие, которое осуществил на наших глазах кто-то другой, на основе одного только наблюдения? Параметры кодирования в зрительной системе отличаются от параметров кодирования в моторной. Какие же корковые процессы вступают здесь в действие и какие сенсомоторные преобразования необходимы? Обратившись к процессу научения, мы тут же сталкиваемся с новыми затруднениями: вдобавок к проблеме соответствия возникает проблема передачи умений и двигательных возможностей, которые во всей своей сложности могут быть не представлены в нашем словаре двигательных актов. Как мы добавляем к своему поведенческому репертуару новые действия? Как переводим зрительно воспринимаемую последовательность движений, каждое из которых, взятое по отдельности, может быть лишено смысла, в потенциальное действие, вполне для нас осмысленное?
Начнем с определения подражания как способности индивида повторить наблюдаемое действие. Для объяснения этой способности предложено две основных теоретических модели. Первая основана на четком разделении сенсорных и моторных кодов: подражание, согласно этой модели, возможно благодаря ассоциативным процессам, связывающим элементы, между которыми a priori нет ничего общего35.
Во второй, напротив, допускается, что у действия наблюдаемого и действия выполняемого должен быть общий нервный код, и это обязательное условие для подражания.
В последние годы на передний план вышла вторая модель, и произошло это прежде всего благодаря работам Вольфганга Принца и его сотрудников. В основе этих работ лежит понятие «идеомоторного действия», разрабатывавшееся Германом Лотце, а вслед за ним — Уильямом Джемсом36, и распространенное на подражание с опорой на принцип «идеомоторной совместимости», введенный американским психологом Энтони Г. Гринвальдом37. В соответствии с этим принципом, чем больше сходство наблюдаемого акта с тем, который присутствует в поведенческом репертуаре наблюдателя, тем сильнее тенденция к его выполнению: следовательно, у восприятия и исполнения должна быть «общая репрезентативная основа», на которую оказывает влияние понимание наблюдателем цели движений наблюдаемого38.
Открытие зеркальных нейронов влечет за собой возможность переформулировать принцип идеомоторной совместимости: общую репрезентативную основу следует рассматривать не как абстрактную и амодальную, но как механизм, преобразующий зрительную информацию напрямую в возможные двигательные акты. Это подтверждается томографическими исследованиями мозга, среди которых наиболее показательные результаты были получены в работах Марко Якобони и его коллег39.
В основу этих экспериментов была положена следующая экспериментальная процедура: на экране компьютера предъявлялись фиксационная точка и видеофрагменты, в которых демонстрировались человеческая рука с поднимающимся указательным или средним пальцем (А), та же рука, на сей раз неподвижная, однако с помеченным крестиком указательным или средним пальцем (Б), или серый фон с изображенным на нем крестиком (В). Испытуемых просили либо просто смотреть на стимулы, либо, посмотрев на них, поднять палец, движение которого они видели в видеофрагменте («подражание»), или же поднять палец, который был помечен крестиком. В последнем условии их просили поднять указательный палец, если крестик появлялся слева от точки фиксации, и средний — если крестик появлялся справа от нее. Было показано, что при подражании активируются задняя часть нижнелобной извилины левого полушария (так называемый лобный полюс зеркальной системы) и участок верхней височной борозды (STS) правого полушария, и что эта активация сильнее, чем в случае двигательных актов, не являющихся подражательными. Выявленные различия в активации указывают на то, что зеркальная система участвует в подражании действиям, уже присутствующим в поведенческом репертуаре наблюдателя, обеспечивая, по всей видимости, непосредственную двигательную реализацию наблюдаемого действия.
Сходные результаты были получены Нобуяки Нишитани и Риттой Хари в эксперименте с регистрацией магнитоэнцефалограммы (МЭГ)40. В этом исследовании испытуемым давали инструкцию взять предмет (А), пронаблюдать, как это действие выполняет экспериментатор (Б), и, пронаблюдав действие в исполнении экспериментатора, повторить его (В). Как хорошо известно, МЭГ уступает функциональной магнитно-резонансной томографии с точки зрения пространственного разрешения, однако этот недостаток компенсируется отличным временным разрешением, позволяющим подвергнуть исследованию динамику изучаемого процесса.
В результате было показано, что в чисто двигательном условии (А) перед прикосновением руки к предмету активируются нижние отделы лобной коры левого полушария (зона 44 — лобный полюс зеркальной системы), а 100—200 мс спустя наблюдается активация прецентральной моторной зоны левого полушария. В условиях с наблюдением (Б) и подражанием (В) последовательность была сходной, но начиналась с активации затылочной коры левого полушария. Наиболее сильная активация была получена в условии с подражанием. Эти результаты однозначно указывают на то, что зона 44 левого полушария играет существенную роль в подражании наблюдаемому действию.
Важно помнить, что данные, получаемые с использованием методов регистрации активности мозга, носят корреляционный характер: они говорят нам лишь о том, что определенные зоны мозга активируются при выполнении определенных задач, но ничего не говорят о том, какую роль активированная зона играет в осуществлении исследуемой функции. Эту информацию дает повторяемая транскраниальная магнитная стимуляция (пТМС), поскольку посредством длительной стимуляции можно вызвать временное снижение функций той или иной зоны мозга. Недавно с использованием этого метода было доказано, что зеркальные нейроны играют в подражании ключевую роль41.
Группа добровольцев подвергалась стимуляции задней части лобной извилины левого полушария (зоны Брока), в то время как они нажимали на кнопки на клавиатуре, либо подражая аналогичному действию другого человека, либо реагируя на указатель в виде луча красного цвета, направленного на клавиатуру и подсказывающего, какую кнопку нажать. Согласно полученным данным, после ТМС испытуемые хуже решали задачу подражания, тогда как успешность решения зрительно-двигательной задачи не снижалась. Интересно, что с точки зрения двигательного состава подражательная и неподражательная задачи были при этом идентичны.
Приведенные выше данные свидетельствуют о том, что система зеркальных нейронов на самом деле играет важнейшую роль в процессах подражания, перекодируя наблюдаемые действия в двигательные коды и тем самым давая нам возможность их повторить. Здесь следует также отметить, что в эксперименте, проведенном Якобони и его коллегами, область верхневисочной борозды активировалась во время подражания несколько сильнее, чем во время наблюдения.
Некоторое время спустя авторы провели исследование, в котором испытуемые должны были либо просто наблюдать за осуществлением определенного движения, которое экспериментатор выполнял правой или левой рукой, либо повторять это движение после наблюдения, используя при этом только правую руку; согласно полученным результатам, активация в районе верхневисочной борозды не только варьирует в зависимости от того, наблюдает ли испытуемый за действием или должен его повторить, но и зависит от используемой руки. Когда требовалось только наблюдать, наиболее сильную активацию вызывали движения той руки экспериментатора, которая анатомически соответствовала руке, используемой испытуемым (т. е. правой); а при подражании, напротив, наиболее сильную активацию вызывали движения той руки экспериментатора, которая пространственно соответствовала руке, используемой испытуемым (т. е. левой).
Иными словами, когда задача ограничивалась наблюдением, доминировало анатомическое взаимосоответствие (правая рука — правая рука), а когда от участников эксперимента требовалось повторить действия экспериментатора, взаимосоответствие становилось пространственным (моя правая рука, твоя левая рука)42. Не исключено, что эту инверсию в активации верхневисочной борозды в процессе подражания следует рассматривать как следствие влияния фронтопариетальных зеркальных нейронов, способствующих отбору двигательных прототипов, пространственно согласующихся с наблюдаемыми. Попробуйте сказать своему приятелю, что у него испачкана щека, и укажите на свою собственную правую щеку правой же рукой; вот увидите, он примется тереть левую щеку левой рукой!
Подражание и научение
В предыдущем разделе мы обсудили подражание в первом значении этого слова; теперь следует рассмотреть, что происходит, когда речь идет не просто о повторении акта, уже присутствующего в двигательном репертуаре наблюдателя, но об освоении новой двигательной схемы. Не задействованы ли зеркальные нейроны и здесь?
В последние годы был предложен целый ряд моделей, объясняющих механизм этой формы подражания; среди них особо следует выделить модель Ричарда Бирна, этолога из шотландского университета Св. Андрея. Согласно модели Бирна, научение через подражание является результатом сопряжения двух отдельных процессов: первый из них представляет собой расчленение действия, подлежащего повторению, на составные части, или, иными словами, преобразование непрерывного потока наблюдаемых движений в набор действий, наличествующих в двигательном репертуаре наблюдателя; второй процесс состоит в упорядочении этих перекодированных двигательных актов в последовательность, которая соответствует действию, воспроизводящему только что продемонстрированное43. Сходный процесс может лежать и в основе освоения двигательных схем, не раскладывающихся в последовательность — как, например, в случае исполнения аккордов на фортепиано или на гитаре.
Эту форму научения через подражания изучала Пармская лаборатория в сотрудничестве с исследователями из Юлихского научно-исследовательского центра. Группе людей, никогда не игравших на гитаре, показывали видеозапись руки наставника, берущего определенные аккорды; после краткой паузы испытуемых просили воспроизвести показанные аккорды44. Методика включала также три контрольных условия: в первом испытуемые после просмотра видеофрагмента прикасались к грифу гитары, однако получали специальную инструкцию не брать никаких аккордов; во втором их просили следить за грифом гитары, который двигался туда-сюда, а потом смотреть, какой аккорд берет наставник; наконец, в третьем им давалась возможность взять любой произвольный аккорд (рис. 6.1, см. цв. вклейку). Система зеркальных нейронов активировалась, когда испытуемые смотрели на взятый наставником аккорд с целью подражания; кроме того, она активировалась, хотя и в меньшей степени, в двух контрольных условиях: когда испытуемые просто наблюдали, как наставник берет аккорд, или, посмотрев на руку наставника, прикасались к грифу гитары, но сами при этом не брали аккорда (рис. 6.2, см. цв. вклейку). Наиболее же интересным открытием стала значительная и весьма обширная активация участка лобной коры, соответствующего зоне 46 по Бродману, во время паузы, предшествовавшей подражанию (рис. 6.3, см. цв. вклейку). Во время выполнения двигательного акта, вне зависимости от того, требовала ли задача подражания или нет, активировались моторные зоны коры.
Система зеркальных нейронов активировалась, когда испытуемые смотрели на взятый наставником аккорд с целью подражания; кроме того, она активировалась, хотя и в меньшей степени, в двух контрольных условиях: когда испытуемые просто наблюдали, как наставник берет аккорд, или, посмотрев на руку наставника, прикасались к грифу гитары, но сами при этом не брали аккорда (рис. 6.2, см. цв. вклейку). Наиболее же интересным открытием стала значительная и весьма обширная активация участка лобной коры, соответствующего зоне 46 по Бродману, во время паузы, предшествовавшей подражанию (рис. 6.3, см. цв. вклейку). Во время выполнения двигательного акта, вне зависимости от того, требовала ли задача подражания или нет, активировались моторные зоны коры.
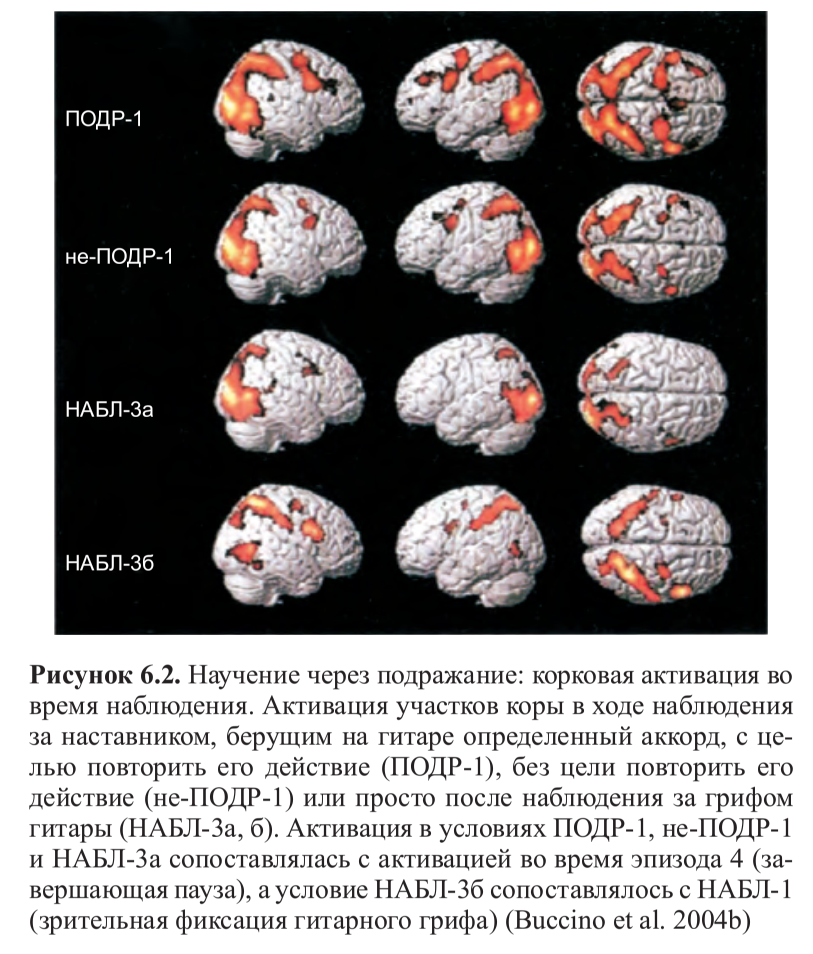
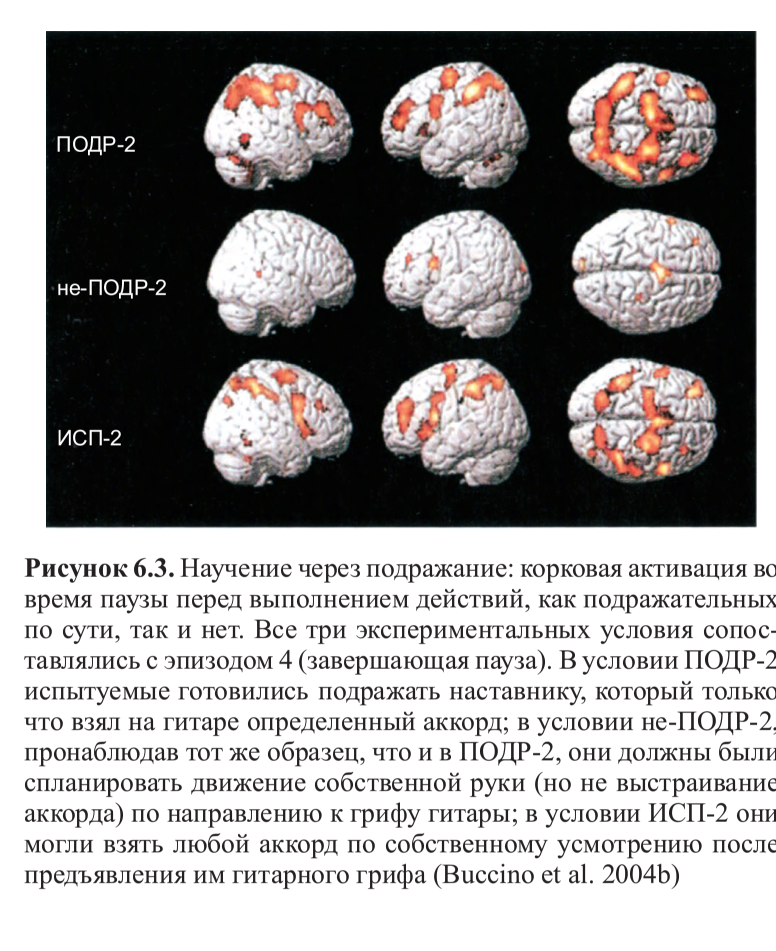 Эти данные показывают, что преобразование зрительной информации в соответствующую двигательную реакцию осуществляется системой моторных нейронов; точнее сказать, зеркальные нейроны, локализованные в нижней теменной и лобной коре, переводят элементарные акты, из которых состоит наблюдаемое действие (в данном конкретном случае — положение, которое необходимо придать пальцам, чтобы взять аккорд), на язык движений.
Эти данные показывают, что преобразование зрительной информации в соответствующую двигательную реакцию осуществляется системой моторных нейронов; точнее сказать, зеркальные нейроны, локализованные в нижней теменной и лобной коре, переводят элементарные акты, из которых состоит наблюдаемое действие (в данном конкретном случае — положение, которое необходимо придать пальцам, чтобы взять аккорд), на язык движений.
Согласно модели Бирна, это условие необходимое, но недостаточное для научения через подражание. Однако ответы нейронов, записанные во время паузы перед повторением действия наставника и перед исполнением аккорда по собственному усмотрению, указывают на то, что система зеркальных нейронов активируется, так сказать, под контролем определенных областей лобной коры — в частности, зоны 46 по Бродману. Прежде отдельные авторы45 соотносили зону 46 с функциями, преимущественно связанными с рабочей памятью, однако результаты этих и других подобных экспериментов заставляют предположить, что эта зона может участвовать и в обеспечении других функций. Помимо рабочей памяти, зона 46, судя по всему, отвечает за рекомбинацию отдельных двигательных актов и формирование новой схемы действия, максимально приближенной к той, которую демонстрирует наставник46.
Проведенный нами анализ показывает, что обе формы подражания зависят от активации корковых зон, обладающих зеркальными свойствами, а следовательно, за ними стоит механизм, осуществляющий сопоставление зрительной информации, получаемой посредством наблюдения за двигательными актами других исполнителей, с соответствующими моторными репрезентациями. Известно, что у людей, в отличие от обезьян, зеркальные нейроны кодируют как транзитивные, так и нетранзитивные двигательные акты и осуществляют четкое отслеживание временных аспектов наблюдаемого действия. Значит, вполне допустимо предположить, что человек, с его из ряда вон выходящим двигательным репертуаром, по своей способности к подражанию и, прежде всего, к научению через подражание, превосходит обезьяну.
И все-таки одного только богатства двигательного репертуара недостаточно для развития способности к научению, равно как мало для этого и системы зеркальных нейронов, которая, вне всякого сомнения, является необходимым условием подражания, но, вместе с тем, не может считаться достаточным его условием. Это касается не только способности к научению через подражание, которая, как мы увидели, требует вмешательства корковых зон, не относящихся к зеркальной системе, но также и способности к повторению действий, выполняемых другими индивидами и входящих в состав нашего двигательного репертуара.
Подражание требует системы, которая управляла бы зеркальными нейронами, осуществляя при этом две функции — облегчения (фасилитации) и торможения (ингибиции). Эта система должна облегчать переход от потенциального действия, закодированного зеркальными нейронами, к непосредственному выполнению моторного акта, когда наблюдатель захочет его осуществить; в то же время, она должна быть способна и оттормозить это действие. Иначе наша зеркальная система работала бы в циклическом режиме: мы бы немедленно бросались повторять любое увиденное нами действие. До чего же хорошо, что это не так!
Существование механизма, управляющего зеркальными нейронами, подтверждается и огромным количеством экспериментальных данных, по преимуществу клинических. Известно, что больным с обширными поражениями лобной коры трудно перестать повторять действие, которое выполнил на их глазах кто-то другой — прежде всего, лечащий врач (т. е. для них характерно подражательное поведение). У больных с более серьезными нарушениями этих управляющих механизмов может наблюдаться еще одна форма патологического поведения — эхопраксия. Больные, страдающие этим расстройством, характеризуются навязчивым стремлением тотчас же повторять, словно бы рефлекторно, даже самые причудливые действия окружающих. Таким образом, поражения лобной коры выводят из строя тормозный механизм, блокирующий преобразование потенциальных действий, закодированных фронтопариетальной системой, в подражательные акты. Блокирование определяется торможением передних медиальных областей, в частности, предлежащих дополнительной моторной области (пре-SMA): судя по всему, эти области оказывают стимулирующее воздействие на фронтопариетальную систему.
Вероятно, именно эти медиальные области, при условии сохранности лобной коры, отвечают за осуществление подражательных актов, когда индивид сочтет их полезными или уместными. И хотя непосредственного доказательства активации этих зон в момент принятия решения о подражании на данный момент не получено, электрофизиологические исследования показывают, что медиальные отделы коры активируются за 800 миллисекунд до начала действия, из чего можно предположить, что их активация отражает общее решение приступить к действию.
Наконец, отношения между зеркальными механизмами и системами управления проливают свет еще на некоторые вопросы, связанные с ранним подражанием у новорожденных и с псевдоимитацией у взрослых. Наблюдение за младенцами показывает (хотя и небесспорно), что уже в первые часы после рождения они способны повторять за родителями определенные движения рта и губ (например, высовывание языка), хотя еще не видели собственного лица47. Как заметил Эндрю Мельцофф, «в колыбелях зеркал не бывает» — и тем не менее, младенцы, судя по всему, наделены способностью к этой форме подражания. Этот факт можно объяснить тем, что у них уже есть система зеркальных нейронов, пусть весьма рудиментарная, в то время как управляющие механизмы еще слабы, на что указывает также низкая степень миелинизации и, как следствие, ограниченные функциональные возможности лобных долей.
Что касается взрослых, в 1872 году Чарльз Дарвин привлек внимание ученых к определенным формам поведения, свойственным для моторного резонанса:
По уверению одного лица, на которое я могу положиться, нередко можно услышать, как многие зрители начинают откашливаться всякий раз, когда выступающий перед ними певец внезапно охрипнет. […] Я слыхал также, что на состязаниях в прыжках многие из зрителей […] начинают двигать ногами в момент, когда прыгун делает прыжок48.
Если мы хотим проверить, насколько верно это замечание сэра Чарльза, все, что нам нужно — это отправиться на футбольный матч! В любом случае, весьма вероятно, что это двигательное «раскрепощение» определяется ослаблением управляющих механизмов, связанных также и с феноменами эмоционального сопереживания, и в результате него проявляются определенные движения и даже действия, которые иначе остались бы в системе зеркальных нейронов на стадии потенциальных.
Жестовая коммуникация
Выше мы рассмотрели механизм, лежащий в основе нашего непосредственного понимания действий других людей. В результате проведенного анализа нам удалось прояснить нервную основу разных типов подражания. Кроме того, как будет показано в данном разделе, тем самым мы сможем больше узнать о нервных основах некоторых форм коммуникации, благодаря чему нам удастся хотя бы в общих чертах воссоздать один из возможных вариантов процесса происхождения человеческого языка49.
Это отнюдь не означает, что самого по себе наличия системы зеркальных нейронов, присущей и обезьянам, достаточно для того, чтобы объяснить появление преднамеренного и тем более речевого коммуникативного поведения. Это следует из нашего обсуждения подражания: одно дело — понять, и совсем другое — повторить только что увиденное действие. И тем не менее, хотя подражание требует активации не только системы зеркальных нейронов, но и других зон коры, нам было бы крайне трудно повторить по образцу какое бы то ни было действие, если бы мы были лишены специального механизма, кодирующего сенсорную и моторную информацию о действии или о последовательности действий в едином формате. Но ведь то же самое верно и в отношении любой из естественных форм коммуникации? Вне зависимости от того, будет ли это вербальная или невербальная коммуникация, разве не должна она удовлетворять требованию взаимосоответствия, согласно которому
«передатчик и приемник должны быть объединены общим пониманием значений»? Разве была бы возможна коммуникация, если бы то, что важно для передатчика, было бы лишено смысла для приемника, если бы «процессы порождения и восприятия» не были бы «некоторым образом связаны друг с другом», а «их репрезентация» не была бы «до определенного уровня одной и той же»50?
В практическом смысле и вне зависимости от конкретных нейронных механизмов, зеркальная система лежит в основе общего пространства действия: если мы видим, как кто-то берет в руку пищу или кофейную чашку, нам сразу становится ясно, что этот человек делает. Нравится нам это или нет, но уже первые намеки на движение руки что-то нам «коммуницируют», и это что-то — значение действия: то самое значение, которое является общим для нас с человеком, выполняющим действие, благодаря активации нашей моторной коры. Разумеется, термин «коммуникация» используется здесь в максимально широком смысле слова, и нельзя отрицать ту огромную пропасть, которая разделяет узнавание двигательного акта (например, захвата рукой) и понимание жеста (неважно, ручного, мимического или вербального), реализующего недвусмысленно коммуникативное намерение. Но хотя эта пропасть и огромна, ее ни в коем случае не следует считать непреодолимой.
6. Подражание и язык
Предположим, к примеру, что действие, выполнение которого вы наблюдаете, представляет для вас особый интерес. В этом случае, когда вы видите, как у другого человека двигается рука, ваша рука тоже начинает сходным образом двигаться, хотя сами вы можете этого и не осознавать; но это едва заметное движение не ускользнет от внимания другого и может повлиять на его поведение. Зеркальный механизм, благодаря которому вы понимаете действие другого с самых первых движений, обеспечивает также и понимание вами последствий, которые влечет за собой ваша непроизвольная реакция, обеспечивая тем самым отношения реципрокного взаимодействия между вашей рукой и рукой другого.
Это взаимодействие не слишком сильно отличается от «жестового разговора», который, согласно Миду, характерен для подготовительных фаз многих форм поведения животных, от борьбы и до ухаживания, от заботы о молодняке и до игры:
Даже у животных, находящихся на более низких ступенях развития, чем человек, есть […] разновидности поведения, которые по природе своей могут быть отнесены к жестам. К ним относятся начальные фазы действий, вызывающих инстинктивные реакции со стороны других особей. Эти начальные фазы поведенческих актов вызывают реакции, которые ведут к поднастройке начатых актов, а эта поднастройка, в свою очередь, ведет к новым начальным реакциям, вновь влекущим за собой поднастройку51.
Эта взаимная поднастройка наделяет поведенческие акты животных социальным значением, тем самым выстраивая формы взаимопонимания, которые во многих отношениях можно считать предшественниками истинно намеренной коммуникации. Последняя, однако же, требует способности к управлению системой зеркальных нейронов и к включению знаний о влиянии, которое жесты оказывают на поведение другого, в моторную память, чтобы впоследствии их можно было опознавать в поведении других. Более того, «разговор» не может быть просто набором транзитивных жестов, он должен предполагать доступ к двигательному репертуару, который кодирует нетранзитивные, пантомимические и чисто коммуникативные акты.
Мы уже видели, что многие коммуникативные жесты из числа наиболее типичных для обезьян — такие как чмокание губами или их выпячивание — появились в результате ритуализации актов, связанных с поеданием (либо пищи, либо паразитов, добытых из шерсти сородичей во время груминга), и используются для выстраивания отношений и закрепления связей с членами племени; мы знаем также, что определенные зеркальные нейроны, кодирующие движения рта, активируются и во время еды, и во время наблюдения за коммуникативными актами, связанными с движениями губ (орофациальными).
В множестве работ рассматриваются ручные жесты горилл и шимпанзе, несущие более или менее явную коммуникативную функцию, вне зависимости от того, живут ли обезьяны в естественной среде обитания или содержатся в неволе52. Что касается людей, известный русский [советский, в «исторической России» он стал бы не великим психологом, а максимум преуспевающим юристом, в лучшем случае — театральным критиком в свободное время. Прим.публикатора] психолог Лев Выготский выдвинул предположение, что в основе большинства нетранзитивных актов, осуществляемых детьми, лежат транзитивные акты. В частности, он заметил, что если предмет можно достать рукой, ребенок его хватает, а если отодвинуть предмет подальше, все равно вытягивает руку, как если бы пытаясь дотянуться до этого предмета.
Если мать ребенка реагирует на эти попытки достать предмет и приходит на помощь, ребенок впоследствии повторяет жест дотягивания для указания на предметы, которые ему хочется получить53. В то же время, исследования, которые мы обсуждали выше, со всей очевидностью указывают на то, что система зеркальных нейронов у человека активируется также при наблюдении за пантомимическими движениями рук, нетранзитивными актами и результативными орофациальными коммуникативными жестами.
А может быть, поступательная эволюция системы зеркальных нейронов, исходно сформировавшейся для распознавания транзитивных актов (таких как хватание, удерживание, дотягивание и т. п.), породила нервный субстрат, который лег в основу первых форм коммуникации между индивидами? Возможно, отвечающая за управление речевыми процессами человека и порождение речи система, локализованная на боковых поверхностях больших полушарий головного мозга, сложилась в эволюции на основе этой системы с анатомически сходной локализацией?
Можно было бы возразить, что эта гипотеза притянута за уши, и что наверняка есть другие, более экономные объяснения первых шагов человечества в направлении языка и речи. Например, известно, что многие приматы издают разнообразные звуки, начиная от контактных криков (указывающих, где находится особь, и позволяющих большим группам животных скоординировать свои перемещения) и заканчивая криками, сигнализирующими об отыскании пищи или предупреждающими о появлении хищника.
Дороти Чини и Роберт Сейфарт провели серию остроумных экспериментов, в которых показали, что восточноафриканские зеленые мартышки (Cercopithecus aethiops) издают разные крики в зависимости от того, какое место в иерархии занимает приближающаяся особь, и в зависимости от типа выполняемого ими действия (например, следят ли они за находящимся на некотором расстоянии конкурирующим племенем или перемещаются по открытой местности); кроме того, они проводят различение между разными типами хищников (птицы, кошки, змеи и т. д.) и, предупреждая сородичей об их приближении, используют для каждого типа определенный крик54.
Так почему бы нам вслед за Стивеном Пинкером не допустить, что «первые шаги» в эволюции человеческого языка были сделаны тогда, когда «набор квазиреференциальных криков подобного рода оказался под контролем коры больших полушарий головного мозга», благодаря чему первым делом появляется возможность использовать сочетания последовательностей криков для обозначения «сложных событий», а затем — применение этой
«способности анализировать сочетания криков […] к частям каждого отдельного крика»55.
Есть по меньшей мере две причины усомниться в правомерноcти этой эволюционной гипотезы. Первая по природе своей функциональна: звуки, издаваемые обезьянами, относятся исключительно к эмоциональному поведению; более того, даже если эти звуки в некотором смысле указывают на конкретный объект, как крики зеленых мартышек, они привязаны к определенной функции (скажем, к такой, как предостережение) и не могут использоваться в других целях. Иными словами, один и тот же сигнал может нести в себе не более одного сообщения (например, «иди сюда» или «убегай», в зависимости от обстоятельств).
Следовательно, ни один из этих сигналов не может повлечь за собой какой-либо иной формы поведения, кроме той, для которой он был порожден. В коммуникативных системах, не связанных с эмоциями, дела обстоят иначе: один и тот же знак может нести в себе множество значений, актуализирующихся в зависимости от контекста, в котором этот знак используется. В человеческом языке, к примеру, одно и то же слово (скажем, «огонь») может означать, что начался пожар (сообщение «убегай!»), а может сигнализировать, что удалось разжечь костер, на котором можно приготовить пищу (сообщение «иди сюда!»). Именно недостаток подобной гибкости делает систему криков зеленой мартышки непригодной в качестве основы для эволюции языка56.
Вторая причина носит преимущественно анатомический характер: мозговые системы, задействованные в порождении криков у обезьян, кардинально отличаются от речевых зон человека. Если к первым относятся прежде всего поясная кора, промежуточный мозг и стволовые структуры57, то вторые группируются вокруг латеральной (Сильвиевой) борозды и задней части нижнелобной извилины. Разве разумно было бы полагать, что в ходе эволюции приматов система голосовой коммуникации перепрыгнула из нижележащих отделов мозга на латеральную поверхность коры? Глубинные структуры человеческого мозга тоже содержат систему голосовых сигналов, но она связана с эмоциональными голосовыми реакциями (разнообразными восклицаниями, возгласами и т. п.) и едва ли имеет отношение к настоящему языку. Следовательно, куда логичнее было бы исследовать зоны коры головного мозга обезьян, гомологичные тем зонам, которые управляют речевыми процессами у человека, обращая особое внимание на функциональные свойства теменно-височных и нижних лобных участков коры.
[Есть и третья причина — самые совершенные из систем коммуникации животных, дифференцированные сигналы, имеющие внешнего референта, исследование которых и началось работами Сифарта-Чини, а) инстинктивной природы и б) закрытые. Как бы ни был богат индивидуальный опыт животного, как бы ни важны были выводы его рассудочной деятельности для сородичей, он (или она) не могут передать их другим средствами данной системы коммуникации. См. характерный пример с мартышкой Диана C.diana. Ею передаются лишь значения, жёстко закреплённые в видовой норме, даже когда последовательности сигналов имеют иное значение, чем каждая из составляющих, как у больших белоносых мартышек C.nicticans.
Человеческий же язык — открытая система; комбинируя друг с другом знаки и удлиняя последовательности, она позволяет общими средствами передать индивидуальное знание и индивидуальное чувство всем прочим членам сообщества, включая неизвестных лично тебе. Причём реципиенту «не нужно догадываться» о содержании сообщения, всю работу по его кодированию произвёл передатчик, другой особи достаточно воспринять. Последнее и есть признак истинной коммуникации по Гюнтеру Тенброку, в отличие от более примитивной ситуации, когда работа по декодированию лежит на приёмнике сообщения и не может быть выполнена безотносительно изучения им обстоятельств контекста.
Именно таковы жесты и вокализации обоих видов шимпанзе, скорей всего и других антропоидов: дифференцированные системы сигналов инстинктивной природы «типа мартышек» у них уже исчезли, открытые системы вроде человеческого языка ещё не появились, почему все жесты, позы и крики представляют собой идеальный континуум, разные части которого могут использоваться только как сигналы ad hoc, непоонятные реципиенту вне отслеживания им происходящего наряду с передатчиком. Прим.публикатора]
Известно, что зона Брока, одна из классических речевых зон, задействована не только в речевых, но и в других двигательных актах и активируется при движениях губ (орофациальные движения), рук (брахиомануальные движения) и гортани (ороларингальные движения), а ее организация сходна с организацией гомологичной области у обезьян — F5. Более того, зона Брока, как и F5, входит в состав зеркальной системы, основная функция которой как у человека, так и у обезьян состоит в установлении связи между пониманием действия и выполнением действия.
Отсюда следует, что ответ на вопрос о происхождении языка следует искать не в примитивных формах голосовой коммуникации, а в эволюции жестовой коммуникации, находящейся под контролем латеральных зон коры. Поскольку данные в поддержку гомологии зоны Брока у человека и F5 у обезьян относятся к области анатомии и цитоархитектоники (и сами по себе в обоих случаях никак не связаны с открытием моторных нейронов), тот факт, что та и другая зоны объединены общим механизмом (и что у человека этот механизм обладает новыми свойствами, связанными с освоением языка), позволяет выдвинуть предположение, что поступательное развитие системы зеркальных нейронов сыграло ключевую роль в появлении и эволюции способности человека к коммуникации, сначала с использованием жестов, а потом и устной речи.
Рот, рука, голос
Идея о происхождении языка от жестов отнюдь не нова: Этьен Бонно де Кондильяк рассказывает притчу в библейском стиле о двух маленьких детях, мальчике и девочке, которые, после того как они пережили Всемирный потоп, потерялись в пустыне и, совершенно не используя язык «знаков», начали общаться при помощи «жестов»58.
Подобные представления могут быть найдены и в рассуждениях психолога Вильгельма Вундта относительно «естеcтвенного пути» развития языка жестов, который, будучи «несовершенным» и в основном связанным с «мимической репрезентацией», обеспечивает «раннюю форму дискурса», в который только спустя определенное время была интегрирована голосовая форма общения59. Однако, наиболее существенные данные в пользу этого подхода появились в последние пару десятков лет в областях палеонтологии, этологии, нейрофизиологии и сравнительной анатомии; он приобрел огромное количество поклонников, особенно среди тех, кто поддерживает так называемые сенсомоторные теории происхождения и восприятия речи60.
Один из таких ученых — Питер МакНейлидж. Он утверждает, что чередование открывания и закрывания рта представляет собой типичный для человеческой речи «силлабический строй», а его модуляции определяют различное «содержание» (гласные и согласные). Такое чередование является, согласно этому автору, эволюционным приобретением артикуляционной системы человека, ведущим свое происхождение от типичного циклического движения челюстей для пережевывания и заглатывания пищи. По его мнению, эта гипотеза может быть подтверждена тем фактом, что у приматов наибольшие изменения происходят в той зоне лобной коры, которая является гомологом зоны Брока и отвечает в первую очередь за контроль жевательных движений61.
Обнаружение в вентральной премоторной коре обезьян зеркальных нейронов, кодирующих движения рта, может служить доказательством теории МакНейлиджа, в которой, однако, недооценивается роль жестов рук (брахиомануальных движений) в эволюции языка. Именно вследствие таких особенностей строения и функционирования зон F5 и Брока, которые характеризуются брахиомануальными моторными репрезентациями (а не только орофациальными и ороларингальными), можно предположить, что истоки межиндивидуальной коммуникации стоит искать не только в двигательной модальности, но в увеличении интеграции мимических, ручных и, не в последнюю очередь, вокальных жестов, чему также способствует возникновение соответствующих зеркальных систем.
Перефразруя Майкла Корбаллиса, можно сказать, что истоки языка следует искать не только во рту, но и в руках, а их взаимодействие дает начало развитию речи62. Невозможно отрицать, что коммуникативные возможности были бы резко ограничены, если бы орофациальная система не поддерживалась брахиомануальной. Попробуем осознать, насколько в коммуникации важны движения рук: они позволяют нам вводить в диалог «третью сторону», указывая на него и описывая некоторые его характеристики. Первая открытая система коммуникации, которая позволяла выражать новые значения при помощи использования всех возможных комбинаций отдельных движений (чего не происходит при орофациальной коммуникации у приматов), несомненно, больше обязана использованию рук, чем рта.
Как недавно заметил Майкл Арбиб, способность человекообразных сначала подражать, а затем и воспроизводить движения из своего моторного репертуара, а также выполнять движения рук, являющиеся «прото-знаками», которые обеспечивают точность и надежность при коммуникации, должна была сыграть определяющую роль в эволюции этой коммуникативной системы63. Это неизбежно приводит к радикальным преобразованиям на уровне коры, в частности, моторной. Вполне вероятно, что наши человекообразные предки, жившие 20 миллионов лет назад, обладали уже зеркальной нейронной системой, позволявшей им выполнять и распознавать простые моторные акты, такие как хватание, удержание, и так далее, и что наши общие предки с шимпанзе около 5—6 миллионов лет назад также имели зеркальные нейроны, обеспечивавшие простейшие формы подражания.
К сожалению, от этого периода между разделением эволюционных ветвей шимпанзе и человека и появлением первых австралопитеков сохранилось очень мало ископаемых свидетельств, но анализ эволюции головного мозга, проведенный на основании исследования черепных коробок, принадлежавших представителям Homo habilis, человека умелого, жившим около 2 миллионов лет назад, показал, что на этом этапе эволюции особенно сильно развивались лобные и височно-теменные отделы64. На основании этого можно предположить, что переход от австралопитека к Homo habilis совпал с переходом к более дифференцированной зеркальной системе, обеспечивавшей нейронный субстрат для формирования «мимической культуры», пик которой, согласно Мерлину Дональду, совпал с появлением Homo erectus, человека прямоходящего, который ходил по Земле в период 1,5 миллиона — 300 тысяч лет назад65.
Также возможно предположить, что зеркальные нейроны развивались и далее, во время перехода 250 тысяч лет назад от Homo erectus к Homo sapiens, в ответ на увеличение как моторного репертуара, так и новообразованной способности к целенаправленной коммуникации путем жестов рук, которая постепенно становилась все более развитой и часто сопровождалась вокализациями. Вероятнее всего, развитие брахиомануальной системы коммуникации повлияло на степень важности вокализаций и, преимущественно, на их контроль. Мы не хотим сказать, что они не сопровождали орофациальную коммуникацию, а как раз наоборот.
Однако, в этом случае добавление голоса к коммуникативному акту выполняло функцию передачи эмоционального значения и служило усилением важности сообщения (например, выражая радость или гнев). Для их выполнения не требовалось соблюдения какой-либо определенной формы, чем и объясняется то, что такого рода вокализации оставались под контролем древних систем, локализованных преимущественно в подкорковых центрах. Положение дел должно было радикально измениться, когда в целенаправленной коммуникации звуки начали использоваться совместно с мимикой и жестами, «прото-знаками»: как ранее произошло в случае с жестами рук, точность, описательность, надежность выражения теперь требовались и от вокализаций, равно как и способность распознавания этих навыков и выучивания их путем имитации. Если бы это было невозможно, вряд ли голосовая коммуникация смогла бы функционировать параллельно с жестовой, а в итоге и превзойти ее по значимости.
Однако, для этого также понадобилось включение новых корковых зон, чтобы контролировать произносимые звуки — возможно, именно так и была сформирована нынешняя зона Брока, обладающая зеркальными свойствами, репрезентациями движений гортани, а также очень тесно связанная с прилежащей первичной моторной корой. В начале 1930-х годов cэр Ричард Пэджет предпринял попытку продемонстрировать, насколько вокальные «прото-знаки», равно как и их жестовые аналоги, были связаны с мимикой, и что это позволяло предположить у них коммуникативную функцию. Таким образом, образовывался вербальный «прото-язык» с несколько примитивным словарем и чрезвычайно простым синтаксисом66. Анализируя ряд корней слов из совершенно различных языков — от полинезийского и китайского до некоторых семитских и индо-европейских — Пэджет обнаружил определенные параллели между звучанием слов и их значением, что, по его мнению, было следствием того факта, что движения рта, губ, и, особенно, языка представляли собой подражание жестам рук и тела, сопровождаемых издаванием специфических звуков.
Такая связь звуков и жестов не только сохранилась в эволюции, но и положила начало речи как таковой. Далее, он утверждал, что, например, использование гласных, таких как «а» или «и», не столько зависело от качества звука, сколько обусловливалось отсылкой к чему-то большому или маленькому, соответственно, так как форма рта при издавании этих звуков воспроизводила форму рук, принимаемую для захватывания объектов. То же верно и для других звуков, например, «м», который обозначает неподвижное состояние, или звука «тр», обозначающего бег или ходьбу.
Конечно, это все — только спекулятивная теория, зачастую основанная на непроверенных предположениях, но, как выяснилось благодаря ряду ученых, в числе которых американский лингвист Моррис Суодеш67, она гораздо менее наивна, чем считалось на протяжении многих лет. Будучи таковой, эта теория является удачной попыткой объяснения, как доступная для зрительного восприятия брахиомануальная система жестов была интегрирована и в дальнейшем вытеснена закрытой от непосредственного восприятия системой ороларингальных жестов без утраты возможности передачи значений и, следовательно, коммуникации. Более того, с нейрофизиологической точки зрения это означало бы, что две эти жестовые системы тесно связаны на корковом уровне и частично имеют общий нейронный субстрат. Ряд недавних исследований показал, что именно так все и происходит.
Эксперименты с использованием ТМС показали, как возбудимость в зоне моторных репрезентаций правой руки увеличивается при чтении или говорении68. Этот эффект ограничен только репрезентацией правой руки и не включает моторную зону, кодирующую движения ноги. Нужно отметить, что возбудимость моторной коры, кодирующей движения руки, не может быть приписана вербальной артикуляции, так как эта функция является общей для обоих полушарий, а наблюдаемый эффект присутствовал только в левом полушарии. Следовательно, зарегистрированная фасилитация должна быть вызвана совместной активацией моторной зоны для правой руки и речевой системы.
Маурицио Гентилуччи69 и его коллеги пришли к подобному выводу, используя совершенно другой подход. Они просили участников эксперимента хватать два объекта разного размера при помощи рта, одновременно разжимая правую руку. Их результаты показали, что наибольшая ширина раскрытия пальцев наблюдалась при захватывании ртом большего из объектов. Еще более явно тесную связь между движениями руки и гортани показывают результаты другого их эксперимента, в котором испытуемым предъявлялись трехмерные объекты, один большой и один маленький, на поверхности которых было изображено два символа или последовательность символов.
Испытуемых просили хватать объекты, но когда они видели символы, им также нужно было открывать рот. Регистрация движений руки, кисти руки и рта показала, что несмотря на полученную инструкцию открывать рот каждый раз одинаково во всех экспериментальных условиях, испытуемые шире раскрывали рот, когда брали в руку больший из двух объектов. В контрольной серии было ясно показано, что этот эффект был специфичным для движений рта и кисти контралатеральной руки, и что одновременное вытягивание предплечья контралатеральной руки никак не влияло на выполнение задания.
Этими авторами был также проведен еще один эксперимент с использованием того же метода, но отличие было в том, что испытуемых просили произносить слоги (такие как «гу» или «га») вместо того, чтобы просто открывать рот. Слоги были написаны в тех же местах, что и наборы символов в предыдущем эксперименте. Было обнаружено, что наибольшее раскрытие губ и мощность голоса при произнесении слогов соответствовала условию с хватанием большего объекта.
Как показано выше, и простые движения рта, и комплексные ороларингальные синергии, необходимые для произнесения слогов, связаны с жестами руки. Более того, выполнение и моторного акта, требующего большого раскрытия ладони, и ороларингального движения, требующего большого раскрытия рта, основаны на общей нейронной организации, представляющей собой рудимент той стадии эволюции языка, на которой звук начал передавать значение благодаря способности рта и ороларингальных систем к артикуляции и выражению семантических значений, аналогичных тем, которые кодирует система жестов руки.
Гентилуччи и его коллеги также показали, что хватательные движения руки влияют на произношение слогов, даже если они не выполняются человеком, а только наблюдаются у другого исполнителя. В очередном эксперименте участников просили произносить слоги «ба» или «га» и одновременно наблюдать за тем, как другой человек берет объекты разного размера70. На кинематику движений губ и ширину их раскрытия, а также спектр и амплитуду голоса влияли хватательные движения наблюдаемого человека: в частности, максимальные амплитуда голоса и ширина раскрытия рта были больше при наблюдении действий с наибольшими объектами. Здесь необходимо отметить, что существование связи между жестовыми и голосовыми системами подтверждается рядом клинических исследований.
К примеру, было описано, как указывание на объекты на экране при помощи правой руки облегчает выполнение задания на называние объектов у афазиков71, а также, что использование жестов руки может помогать пациентам с повреждениями мозга восстанавливать речевую функцию72. [см. также феномены, которыми обосновывается концепция синкинезии. Прим.публикатора]
Однако, давайте ненадолго вернемся к рассмотрению сценария эволюции. Кажется весьма вероятным то, что эволюционная необходимость в формировании более сложных форм коммуникации подтолкнула развитие высокодифференцированного нейронного механизма контроля за произношением, который затем не только контролировал произнесение специфических звуков, но также и способствовал созданию постоянно расширяющейся (на самом деле, практически бесконечной) совокупности возможных комбинаций звуков, которая привела в итоге к отделению системы голосовой коммуникации от жестовой. Как и когда голосовая система приобрела окончательную независимость, оставив жестовой системе роль только дополнительного к звуковой коммуникации средства, еще является предметом обсуждения.
Это могло произойти совсем недавно в связи с появлением типичного для Homo sapiens голосового тракта73, хотя некоторые авторы предлагают альтернативные решения этого вопроса74. В любом случае, разумно предположить, что этот процесс (или какой-либо другой аналогичный) сопровождался радикальными изменениями в коре, захватывавшими, в частности, моторные центры, связанные с восприятием и продуцированием речевого матерала. Однако, такие изменения были бы совершенно бесполезны без одновременной эволюции механизма, который был бы способен удовлетворять тому, что мы назвали обязательным условием для любых форм коммуникации — другими словами, требованию взаимосоответствия, обеспечивающему то, что и передающий, и получающий сообщение имеют общее понимание того, что происходит.
Элвин Либерман, который более других исследователей внес вклад в изучение условий, необходимых для любого вида коммуникации, показал, что в языковой коммуникации важны не столько звуки, сколько артикуляционные движения, благодаря которым они генерируются, так как последние обеспечивают фонетическое единообразие первых. Вот почему мы способны мгновенно воспринимать различия между слогом «ка» и звуком кашля75. Если мы принимаем интерпретацию Либермана (а большая часть результатов, полученных им за тридцать лет исследований в этой области, ее подтверждает), необходимо осознавать, что переход к автономной голосовой системе должен означать, что моторные нейроны, отвечающие за контроль движений гортани, приобрели способность к активации в ответ на восприятие звуков, производимых аналогичными движениями у других. Иными словами, зеркальная нейронная система претерпела дальнейшую реорганизацию для того, чтобы обеспечивать преобразование звуков речи в моторные репрезентации соответствующих артикуляционных движений. Открытие еще одной зеркальной системы — зеркальной системы для речи — является доказательством того, что такая реорганизация действительно имела место76.
Эксперимент, который провели Лучано Фадига и его коллеги, предоставил первые доказательства существования такой зеркальной системы. Исследователи просили своих испытуемых внимательно слушать предъявляемые им вербальные и невербальные стимулы (слова, псевдослова и звуки, состоящие из двух тонов). При этом регистрировались ДВП с мышц их языка. В словах и псевдословах встречались удвоенные звуки «ф» или «р». Звук «ф» является лабиодентальным фрикативным, то есть, при произнесении требует только незначительных движений языка, тогда как звук «р» — лингвопалатальный фрикативный и требует существенного движения языка. Экспериментаторы отметили, что когда испытуемые слышали слова или псевдослова, содержащие звук «рр», амплитуды ДВП значительно увеличивались по сравнению с теми условиями, когда они слушали битональные звуки и слова и псевдослова, содержащие «фф» (рис. 6.4)77.
Томаш М. Пауш78 и его коллеги получили похожие результаты в ТМС-эксперименте с регистрацией ДВП от губ (от круговой мышцы рта — orbicularis oris), но только в ответ на стимуляцию левого полушария, тогда как стимуляция правого полушария не приводила ни к каким изменениям ДВП ни в одном из экспериментальных условий. Обобщая вышесказанное, результаты нейрофизиологических исследований, как этих, так и описанных ранее, говорят о том, что долгий процесс эволюции языка был отмечен рядом важных событий (интеграцией систем движений рта и рук, формированием репертуара главным образом подражательных жестовых «прото-знаков», возникновением бимодального «прото-языка», состоящего из жестов и звуков, и, наконец, появления преимущественно голосовой системы коммуникации), каждое из которых связано с определенной фазой развития механизма, такого как зеркальная система, который изначально был предназначен для распознавания действий других и был лишен функции целенаправленной коммуникации.
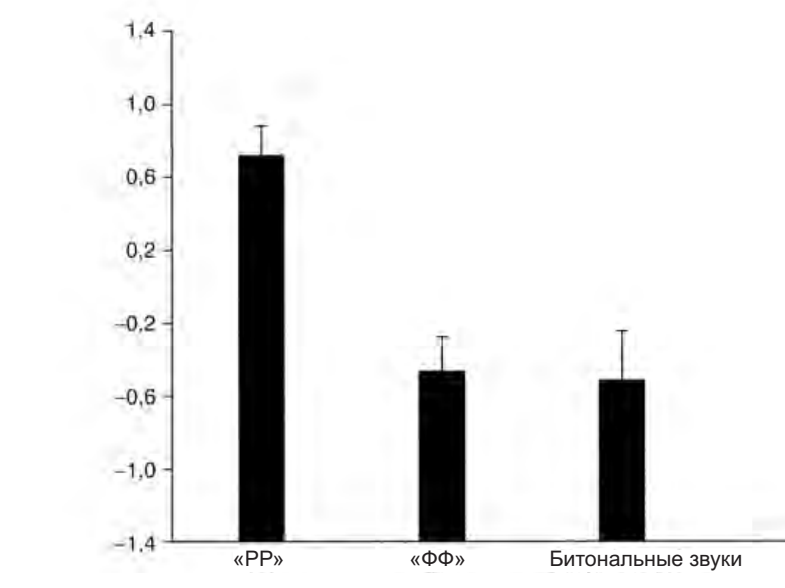
Рисунок 6.4. Двигательные вызванные потенциалы (ДВП), зарегистрированные от мышц языка при прослушивании вербального материала и битональных звуков. На графиках показаны результаты всех испытуемых. «РР» — вербальные стимулы, содержащие лингвопалатальные фрикативы, «ФФ» — вербальные стимулы, содержащие лабиодентальные фрикативы (Модифицировано из работы Fadiga et al. 2002)
Разумеется, это только один из многих возможных сценариев: учитывая чрезвычайную сложность факторов, повлиявших на формирование и развитие языковой способности, большое количество исследований еще предстоит провести. И, все же, мы уверены, что изучение свойств зеркальных нейронов и различных систем, в которые они включены, позволит определить, какие нейронные структуры, снова говоря словами Пинкера,
«могли бы дать эволюции строительный материал для создания систем, обеспечивающих речь человека»
Джакомо Ризолатти, Коррадо Синигалья. Зеркала в мозге. О механизмах совместного действия и сопереживания.
Примечания
1 di Pellegrino et al. (1992).
2Rizzolatti et al. (1996 a); Gallese et al. (1996).
3 Ferrari et al. (2003).
4 См. Van Hoof (1962; 1967); но также Maestripieri (1996).
5 Selzer and Pandya (1994). См. Petrides and Pandya (1984); Matelli et al. (1986). Более подробно обсуждение этого вопроса читатель может найти в первой главе данной книги.
6 Leinonen et al. (1979); Leinonen and Nyman (1979); Hyvärinen (1981). См.
также Graziano and Gross (1995).
7Fogassi et al. (1998); Gallese et al. (2002).
8 Jeannerod (1994).
9См. Byrne (1995); Tomasello and Call (1997); Visalberghi and Fragaszy (1990; 2002) и др.
10di Pellegrino et al. (1992).
11Jellema et al. (2000; 2002).
12 Umiltà et al. (2001).
13Kohler et al. (2002); также см. Keynes et al. (2003).
14 Fogassi et al. (2005).
15Другие примеры см. в работе Yokochi et al. (2003).
16Gastaut and Bert (1954); Cohen-Seat et al. (1954).
17Altschuler et al. (1997; 2000); Cochin et al. (1998; 1999).
18 Hari et al. (1998).
19Fadiga et al. (1995). См. также Maeda et al. (2002).
20Gangitano et al. (2001).
21Decety et al. (1994).
22Rizzolatti et al. (1996b). См. также Grafton et al. (1996); Grézes et al. (1998;
2001).
23Campbell (1905). Petrides and Pandya (1997).
24 Krams et al. (1998); Binkofski et al. (1999); Ehrsson et al. (2000).
25См. Grézes and Decety (2001); Heyes (2001).
26Buccino et al. (2001).
27Iacoboni et al. (2005).
28 Merleau-Ponty (1945, p. 215). Цит. по: Мерло-Понти (1999, с. 241—242).
29 Buccino et al. (2004a).
30 Подробный обзор этих исследований см. в работе Allison et al. (2000).
31Calvo-Merino et al. (2005).
32Дэн Спербер, из личного общения.
33 См., напр., Bekkering and Wohlschlaeger (2002), Wohlschlaeger et al. (2003).
34 См., напр., Byrne (1995), Tomasello and Call (1997); Visalberghi and Fragaszy
(2002).
35См., напр., Welford (1968), Massaro (1990).
36Lotze (1852); James (1890). Джемс в XXVI главе своих «Принципов психологии», где неоднократно цитируется «Медицинская психология» Лотце, привлекает внимание читателя к тому факту, что большинство наших повседневных действий не требуют никаких «решений по поводу этих действий», особого приказа, вслед за которым «непосредственно, без особого решения воли» появляется идея или представление — что гарантирует отсутствие «в нашем сознании противодействующего представления»: «Мы знаем, что такое встать с постели в морозное утро в нетопленой комнате: сама натура наша возмущается против такого мучительного испытания. Многие, вероятно, лежат каждое утро целый час в постели, прежде чем заставить себя подняться. Мы думаем лежа, как мы поздно встаем, как от этого пострадают обязанности, которые мы должны выполнить в течение дня; мы говорим себе: Это черт знает что такое! Должен же я наконец встать!» — и т. д. Но теплая постель слишком привлекает нас, и мы снова оттягиваем наступление неприятного мгновения. Как же мы все-таки встаем при таких условиях? Если мне позволено судить о других по личному опыту, то я скажу, что по большей части мы поднимаемся в подобных случаях без всякой внутренней борьбы, не прибегая ни к каким решениям воли. Мы вдруг обнаруживаем, что уже поднялись с постели; забыв о тепле и холоде, мы в полудремоте вызываем в своем воображении различные представления, имеющие какое-нибудь отношение к наступающему дню; вдруг среди них мелькнула мысль: «Баста, довольно лежать!» Никакого противодействующего соображения при этом не возникло — и тотчас же мы совершаем соответствующие нашей мысли движения. Живо сознавая противоположность ощущений тепла и холода, мы тем самым вызывали в себе нерешительность, которая парализовала наши действия, и стремление подняться с постели оставалось в нас простым желанием, не переходя в хотение. Как только задерживающая действие идея была устранена, первоначальная идея (о необходимости вставать) тотчас же вызвала соответствующие движения.» (James 1890, p. 793; Цит. по: Джемс (1991)).
37Greenwald (1970).
38Bekkering et al. (2000); Bekkering (2002).
39Iacoboni et al. (1999; 2001).
40Nishitani and Hari (2000); см. также Nishitani and Hari (2002).
41Heiser et al. (2003).
42Iacoboni et al. (2001); описание аналогичных экспериментов см. в работах
Koski et al. (2002; 2003).
43Byrne and Russon (1998); Byrne (2002; 2003).
44Buccino et al. (2004b).
45См., напр., Fuster and Alexander (1971), Funahashi et al. (1990).
46 Сходную интерпретацию можно найти в работах: Passingham et al. (2000),
Rowe et al. (2000).
47Meltzoff and Moore (1977).
48Darwin (1872, p. 143). Цит. по: Дарвин (2001, с. 33)
49Rizzolatti and Arbib (1998); Fogassi and Ferrari (2005).
50Liberman (1993); см. также Liberman and Whalen (2000).
51Mead (1910, p. 124).
52Vygotskij (1934). См. Выготский (2005).
53De Waal (1982); Tanner and Byrne (1996); Tomasello et al. (1997).
54Cheney and Seyfarth (1990).
55Pinker (1994, p. 352). Необходимо отметить, что Пинкер и сам не без иронии признавал, что «эта идея имеет не больше подтверждений, чем теория «диндон» (или чем предположение Лили Томлин, что первыми словами, сказанными человеком, была фраза: «Какая волосатая спина!»)» (там же).
56Hauser et al. (2002).
57 Juergens (2002).
58Condillac (1746, p. 175—176). Цит. по: Кондильяк (1980, с. 185)
59 Wundt (1916, p. 128). См.: Вундт (2010).
60См., напр., Armstrong et al. (1995); Armstrong (1999); Corballis (1992; 2002;
2003).
61MacNeilage (1998).
62Corballis (2002, p. 153).
63Arbib (2002; 2005).
64Tobias (1987); Holloway (1983; 1985); Falk (1983).
65Donald (1991).
66 Paget (1930). См. упоминание прото-языка в работе Bickerton (1995).
67Swadesh (1972).
68Meister et al. (2003); см. также Tokimura et al. (1996); Seyal et al. (1999).
69Gentilucci et al. (2001).
70Gentilucci (2003).
71Hanlon et al. (1990).
72 Hadar et al. (1998).
73Lieberman (1975).
74См., напр., работу Gibson and Jessee (1999).
75Liberman and Whalen (2000).
76Rizzolatti and Buccino (2005).
77Fadiga et al. (2002).
78Watkins et al. (2003).
Рекомендуем прочесть
 Зеркальные нейроны и интуиция (отрывок из книги)
Зеркальные нейроны и интуиция (отрывок из книги) «В начале было дело…»
«В начале было дело…» Синкинезия как модус происхождения языка
Синкинезия как модус происхождения языка Что такое эмпатия: когнитивные теории и модели
Что такое эмпатия: когнитивные теории и модели Про несохранение детских воспоминаний
Про несохранение детских воспоминаний «Животное» и «человеческое», расчёт и равенство
«Животное» и «человеческое», расчёт и равенство Нейробиология системы внутреннего представления собственного тела: введение в проблему и прикладные аспекты
Нейробиология системы внутреннего представления собственного тела: введение в проблему и прикладные аспекты![[Наркотическая] зависимость: миф о болезни мозга](http://www.socialcompas.com/wp-content/uploads/2017/02/cover1__w600-150x150.jpg) [Наркотическая] зависимость: миф о болезни мозга
[Наркотическая] зависимость: миф о болезни мозга
