")

Здесь и далее иллюстрации из лекций О.В.Смирновой. Лекция 7. Потенциальная растительность и потенциальный экосистемный покров
В продолжение темы единой теории охраны природы и природопользования
О. В. Смирнова, Л. Б. Заугольнова, В. Н. Коротков
Резюме. Дан синтез современных представлений, посвященных проблемам сохранения и восстановления биоразнообразия лесного покрова. Теоретическую основу реконструкции потенциального лесного покрова составляют концепции синэкологии: мозаично-циклической организации экосистем, иерархического континуума и ключевых видов. Представлена модельная реконструкция лесного покрова Восточной Европы с конца плейстоцена по настоящее время. Обоснована необходимость разработки программ по восстановлению потенциальной биоты особо охраняемых природных территорий и организации экосистемного природопользования в связи с невозможностью полной реализации экосистемных функций как эксплуатируемых, так и охраняемых лесов.
В научной и политической среде мирового сообщества оформилось представление, что устойчивое существование цивилизации невозможно без разработок комплексных программ, направленных на оптимизацию экосистемных функций живого покрова Земли. Особенно актуальна эта проблема в лесной экологии, поскольку именно природные леса в доагрикультурных ландшафтах выполняли основные экосистемные функции: оптимизации климата и гидрологического режима, почвенного плодородия, максимально возможной продуктивности и биоразнообразия (MEA – Millenium…, 2005; Экономика экосистем…, 2010). Однако все возрастающая интенсивность лесопользования приводит к нежелательным изменениям климата и водного режима, деградации почв, падению продуктивности и биологического разнообразия (Schwartz et al., 2000; Hooper et al., 2005; Gamfeldt et al., 2008; Павлов, Букварева, 2011; Сiccarese et al., 2012).
Ситуация требует переориентации систем лесопользования на возможно полную реализацию экосистемных функций. В качестве эталонных объектов, вполне реализующих основные экосистемные функции лесов, рассматривают леса особо охраняемых природных территорий (ООПТ): заповедников, национальных парков и пр. При этом многие полагают, что уровень биологического разнообразия выступает как индикатор степени реализации остальных экосистемных функций (Hector, Bagchi, 2007; Feld, 2009).
 Теоретическая сложность решения проблемы переориентации лесопользования на поддержание экосистемных функций состоит в том, что в современном лесном покрове нет лесов, которые в полной мере можно было бы считать природными и, следовательно, рассматривать как эталонные объекты. Многолетние исследования динамики биологического разнообразия в ООПТ лесного пояса Российской Федерации показали, что режим полной охраны ведет к сокращению видового разнообразия и к существенным изменениям состава и структуры лесов, стимулирует деградационные процессы. Эти изменения, в свою очередь, обусловливают неблагоприятные изменения гидрологического режима и локального климата (Смирнова и др., 1997; Сукцессионные процессы…, 1999; Оценка…, 2000; Мониторинг…, 2008).
Теоретическая сложность решения проблемы переориентации лесопользования на поддержание экосистемных функций состоит в том, что в современном лесном покрове нет лесов, которые в полной мере можно было бы считать природными и, следовательно, рассматривать как эталонные объекты. Многолетние исследования динамики биологического разнообразия в ООПТ лесного пояса Российской Федерации показали, что режим полной охраны ведет к сокращению видового разнообразия и к существенным изменениям состава и структуры лесов, стимулирует деградационные процессы. Эти изменения, в свою очередь, обусловливают неблагоприятные изменения гидрологического режима и локального климата (Смирнова и др., 1997; Сукцессионные процессы…, 1999; Оценка…, 2000; Мониторинг…, 2008).
Как показывает литература по исторической экологии (The Ecological History…, 1998; Vera, 2000; Восточноевропейские леса…, 2004; Vera et al., 2006; Бобровский, 2010; Кюстер, 2012), основная причина падения видового разнообразия в ООПТ – организация их на территориях, испытавших длительные антропогенные преобразования. Лесные земли ООПТ были преобразованы во второй половине – конце голоцена в результате многовекового традиционного природопользования: охоты, выпаса скота в лесах, подсечно-огневого, переложного и пахотного земледелия, рубок, сбора подстилки и пр. В результате были уничтожены виды животных, оказывавших мощное воздействие на лесной покров (зубры, туры, тарпаны, бобры и пр.), и существенно изменены ареалы видов деревьев, которые определяли организацию и динамику природного лесного покрова. Практически все его параметры с позднего голоцена определял человек (Vera, 2000; Смирнова и др., 2006; Бобровский, 2010).
Изъятие территорий традиционного природопользования из хозяйственной деятельности при организации ООПТ не сопровождалось восстановлением природного разнообразия видов растений и животных, а при определении размеров этих территорий не учитывали особенности их популяционной биологии. В результате господствующие позиции в лесном покрове ООПТ стали занимать позднесукцессионные виды деревьев, что сопровождается падением биоразнообразия и ведет к невозможности в полной мере реализовать экосистемные функции.
Существующее на настоящий момент положение, а именно невозможность в полной мере реализовать экосистемные функции как эксплуатируемых, так и охраняемых лесов требует создания программ экосистемного природопользования, сочетающего элементы устойчивого использования ресурсов и поддержания экосистемных функций. При этом на современном уровне знаний восстановление потенциального биоразнообразия, которое может быть достигнуто в современном климате при условии реинтродукции природных обитателей, представляется вполне возможным.
 Необходимость создания таких программ обусловлена:
Необходимость создания таких программ обусловлена:
1) прогрессирующим сокращением биологического разнообразия в связи с прямым уничтожением видов в результате природопользования и антропогенно обусловленной фрагментацией ареалов видов, сохранившихся до настоящего времени;
2) невозможностью поддержания устойчивой структуры популяций видов на основе охраняемых природных территорий в связи с их островным характером и малой дальностью распространения потомства у большинства видов растений и животных;
3) падением биологической продуктивности и сокращением ресурсов местообитаний на хозяйственно используемых территориях в связи с заменой природного разнообразия монокультурами или антропогенно созданными сочетаниями видов;
4) сокращением биологического разнообразия на большей части заповедных территорий в результате введения режима абсолютной охраны.
Восстановление остальных экосистемных функций требует дальнейших теоретических разработок структурно-функциональной организации и динамики экосистем и их ландшафтных комплексов на локальном и региональном уровнях.
Развитие концепции сохранения биоразнообразия на основе современных экологических представлений
Содержание
Представление о современном и потенциальном лесном покрове. Решение проблем оценки, поддержания и восстановления максимального биоразнообразия напрямую связано с развитием представлений теоретической экологии о потенциальной растительности и животном мире и о роли ключевых видов в поддержании лесного покрова (Tuxen, 1956; The Mosaic-Cycle Concept…, 1991; Brzeziecki et al., 1993; Смирнова, 1998; Восточноевропейские леса…, 2004; Мониторинг…, 2008; Методические подходы…, 2010).
Концепция сохранения биоразнообразия основывается:
1) на модельной реконструкции потенциального лесного покрова Северной Евразии в разные этапы его развития, начиная с конца плейстоцена;
2) на выявлении специфики антропогенных воздействий на лесной покров Северной Евразии на разных этапах голоцена.
Необходимость развития представлений о потенциальном растительном покрове возникла у исследователей XX в. в связи с осознанием существенного антропогенного преобразования современного растительного покрова. В качестве альтернативы понятия “современный растительный покров” было предложено понятие “потенциальный растительный покров” (Tuxen, 1956), т.е. такой покров, который мог бы сформироваться после прекращения антропогенных воздействий и восстановления в течение времени, достаточного для достижения всеми видами растений всех мест их возможного обитания и формирования устойчивых потоков поколений.
В потенциальном растительном покрове должны устойчиво существовать все представители потенциальной флоры, локальной или региональной (Tuxen, 1956; Ellenberg, 1988; Brzeziecki et al., 1993; Смирнова, 2010).
 В начале развития представлений о потенциальном растительном покрове исследователи предполагали возможность его восстановления на заповедных территориях независимо от других подсистем биоты (в первую очередь от животного населения) (Соколов и др., 1997). Однако длительные наблюдения в заповедниках и накопление данных о направлениях смен показали, что восстановление потенциальной растительности невозможно без адекватного восстановления животного населения и представителей других царств, формирующих биоту. При этом на конкретных примерах было показано, что восстановление биоразнообразия приводит к изменениям почвенного покрова, гидрологического режима и локального климата (Краснитский, 1983; Злотин, Ясный, 1992; Смирнова, 1998; Оценка…, 2000; Vera, 2000; Восточноевропейские леса…, 2004).
В начале развития представлений о потенциальном растительном покрове исследователи предполагали возможность его восстановления на заповедных территориях независимо от других подсистем биоты (в первую очередь от животного населения) (Соколов и др., 1997). Однако длительные наблюдения в заповедниках и накопление данных о направлениях смен показали, что восстановление потенциальной растительности невозможно без адекватного восстановления животного населения и представителей других царств, формирующих биоту. При этом на конкретных примерах было показано, что восстановление биоразнообразия приводит к изменениям почвенного покрова, гидрологического режима и локального климата (Краснитский, 1983; Злотин, Ясный, 1992; Смирнова, 1998; Оценка…, 2000; Vera, 2000; Восточноевропейские леса…, 2004).
Стало очевидно, что потенциальный растительный покров есть подсистема потенциального лесного покрова в целом; структура и функционирование последнего обусловлено взаимодействиями как биотических, так и абиотических подсистем. Следовательно, антропогенные преобразования любой из подсистем (например, уничтожение диких стадных копытных и/или бобров в лесном поясе Северной Евразии и Северной Америки) обязательно должны были повлиять на все остальные подсистемы. В связи с этим модельная реконструкция потенциального растительного покрова может быть осуществлена только в рамках такой же реконструкции лесного покрова в целом. В результате сложность формирования представлений значительно возрастает, так как в настоящее время отсутствует достаточная информация о специфике преобразований многих биотических и абиотических подсистем (Реймерс, 1992).
Наиболее полные сведения есть только о состоянии растительности и животного мира некоторых регионов. Так, подробное исследование изменений растительного покрова и животного мира Европы показало, что с конца плейстоцена до середины голоцена человек приобрел статус наиболее мощного (по сравнению с остальными членами биоты) средопреобразователя, постепенно заменив мощных средопреобразователей среди животных и растений. В результате часть экосистем, организованных и поддерживавшихся средопреобразующей деятельностью природных средопреобразователей, стала напрямую зависеть от хозяйственной деятельности человека. Вследствие этого значительная часть видов растений и животных в современном покрове существует только в местообитаниях, формируемых после рубок, распашек, выжигания, выпаса скота и пр., и исчезает после введения заповедного режима.
При этом в пределах лесного пояса Восточной Европы растительный покров, восстанавливающийся в ООПТ после прекращения антропогенной деятельности и при отсутствии природных катастроф, содержит не более 30–40% локальной или региональной флоры, соответственно (Смирнова и др., 1997; Оценка…, 2000; Vera, 2000; Восточноевропейские леса…, 2004).
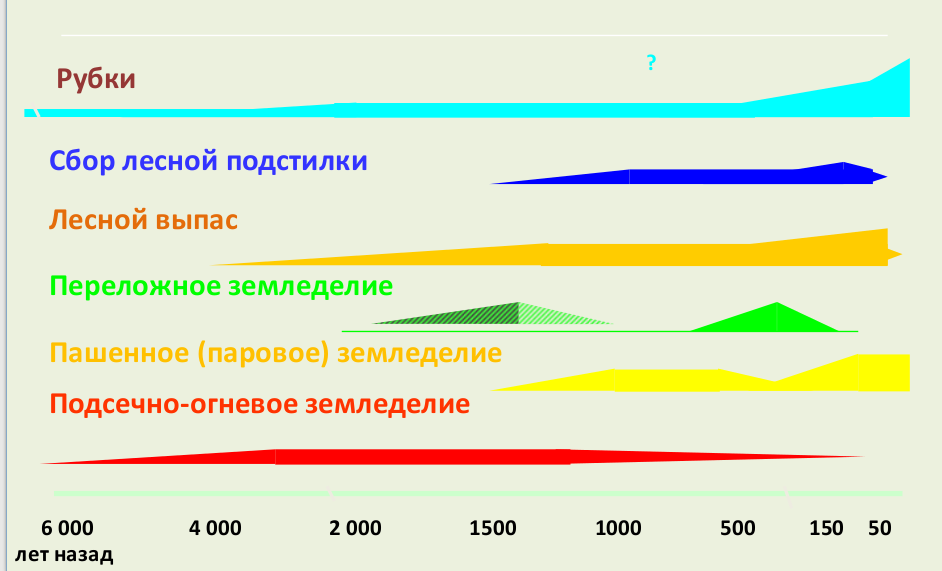 Причины этого явления становятся понятны, если создать модельную реконструкцию доантропогенного лесного покрова. Следует отметить, что такая реконструкция затруднена из-за практически полного отсутствия представлений о потенциальном экотопе и потенциальном почвенном покрове. В связи с этим на современном уровне знаний модельные реконструкции следует рассматривать как предварительные. Для таких реконструкций используют палеонтологические и археологические данные, архивные документы, исторические и современные сведения о распространении, о биологии и экологии сохранившихся до настоящего времени или недавно исчезнувших мощных средопреобразователях среди растений и животных и сопутствующих им видах разных трофических групп.
Причины этого явления становятся понятны, если создать модельную реконструкцию доантропогенного лесного покрова. Следует отметить, что такая реконструкция затруднена из-за практически полного отсутствия представлений о потенциальном экотопе и потенциальном почвенном покрове. В связи с этим на современном уровне знаний модельные реконструкции следует рассматривать как предварительные. Для таких реконструкций используют палеонтологические и археологические данные, архивные документы, исторические и современные сведения о распространении, о биологии и экологии сохранившихся до настоящего времени или недавно исчезнувших мощных средопреобразователях среди растений и животных и сопутствующих им видах разных трофических групп.
Теоретические основы модельной реконструкции биоразнообразия. Теоретической основой реконструкции мы считаем следующие концепции синэкологии (экологии экосистем): мозаично-циклической организации экосистем, иерархического континуума и ключевых видов (The Mosaic-Cycle Concept…, 1991; Collins et al., 1993; Завьялов, 2008; Crain et al., 2008). С позиций концепции иерархического континуума лесной покров можно трактовать как иерархию элементарных популяций видов, формирующих этот покров (Сукцессионные процессы…, 1999; Методологические подходы…, 2010). Хорологическая и хронологическая континуальность лесного покрова связана с совмещением на одной территории элементарных популяций, имеющих разные собственные времена, пространства и уровень плотности. Из-за различий значений этих параметров на конкретном участке формируется разномасштабная мозаика популяций видов разных трофических групп. Механизмом, интегрирующим эту разномасштабную мозаику и приводящим к формированию экосистем как множеств совместно обитающих элементарных популяций, выступает популяционная жизнь наиболее мощных (для данной территории) средопреобразователей.
 В мировой литературе есть несколько синонимичных по сути терминов, обозначающих мощных средопреобразователей. Это эдификаторы (aedificators), экосистемные инженеры (ecosystem engineers) и ключевые виды (keystone species) (The Mosaic-Cycle Concept…, 1991; Завьялов, 2008; Crain et al., 2008; Бобровский, 2010). Исходным можно считать понятие “эдификатор” (aedificator – строитель сообщества) (BraunBlaunquet, Pavillard, 1925), введенное в отечественную литературу В.Н. Сукачевым (1928). По его представлениям, эдификатор – это вид, чье присутствие в биогеоценозе меняет экологические режимы: освещенность, влажность, температуру, химический состав почвы, воды и воздуха.
В мировой литературе есть несколько синонимичных по сути терминов, обозначающих мощных средопреобразователей. Это эдификаторы (aedificators), экосистемные инженеры (ecosystem engineers) и ключевые виды (keystone species) (The Mosaic-Cycle Concept…, 1991; Завьялов, 2008; Crain et al., 2008; Бобровский, 2010). Исходным можно считать понятие “эдификатор” (aedificator – строитель сообщества) (BraunBlaunquet, Pavillard, 1925), введенное в отечественную литературу В.Н. Сукачевым (1928). По его представлениям, эдификатор – это вид, чье присутствие в биогеоценозе меняет экологические режимы: освещенность, влажность, температуру, химический состав почвы, воды и воздуха.
В.Н. Сукачев использовал понятие “эдификатор” применительно к растениям, но в дальнейшем аналогичное понятие было охарактеризовано с помощью терминов “экосистемные инженеры” и “ключевые виды”. В статье мы используем термин “ключевые виды”, поскольку в мировой литературе его применяют при описании как растений, так и животных (The Mosaic-Cycle Concept…, 1991; McCarthy, 2001).
 Для модельных реконструкций потенциального покрова принципиально важно, что ключевые виды в процессе спонтанной смены поколений наиболее значимо (по сравнению с другими видами того же трофического уровня) преобразуют как местообитания элементарных популяций в целом, так и фрагменты этих местообитаний. Это ведет к изменению гидрологического, температурного, светового режимов; микро-, мезорельефа; строения почвенного покрова и пр. Иерархически организованная гетерогенность местообитания элементарной популяции ключевого вида определяет возможность совместного существования в нем экологически и биологически различных подчиненных видов разных трофических групп и в итоге – высокий уровень биоразнообразия.
Для модельных реконструкций потенциального покрова принципиально важно, что ключевые виды в процессе спонтанной смены поколений наиболее значимо (по сравнению с другими видами того же трофического уровня) преобразуют как местообитания элементарных популяций в целом, так и фрагменты этих местообитаний. Это ведет к изменению гидрологического, температурного, светового режимов; микро-, мезорельефа; строения почвенного покрова и пр. Иерархически организованная гетерогенность местообитания элементарной популяции ключевого вида определяет возможность совместного существования в нем экологически и биологически различных подчиненных видов разных трофических групп и в итоге – высокий уровень биоразнообразия.
Несомненно, что все анализируемые виды по размерам их элементарных популяций и интенсивности преобразований среды можно выстроить в континуальные ряды. На современном уровне знаний для выяснения общих принципов организации экосистем и их комплексов можно использовать сведения о наиболее мощных ключевых видах анализируемой территории. Однако для выявления потенциального состава биоты необходимо исследовать или реконструировать состав подчиненных видов, возможность существования которых обусловлена средопреобразующей деятельностью ключевых видов.
В Северной Евразии и Северной Америке с конца плейстоцена описаны наиболее значимые для существования лесного покрова ключевые виды трех функциональных групп:
1) крупные стадные копытные – зубры, туры, тарпаны и др.;
2) бобры;
3) хвойные и широколиственные деревья (ель, пихта, дуб, липа, вяз, клен и др.).
 Наблюдения за представителями этих групп, сохранившихся в ООПТ, позволили охарактеризовать их средопреобразующую деятельность. Элементарные популяции зубров, туров, тарпанов и других крупных стадных копытных занимали бассейны малых рек площадью в сотни кв. км, используя как водораздельные, так и долинные ландшафты. В результате их жизнедеятельности лесной покров был пронизан луговыми и луговостепными полянами, по берегам водоемов формировались водопойные поляны; вся территория была пронизана сетью троп, которые служили каналами миграции для многих подчиненных видов. Тщательные наблюдения за зубрами на протяжении многих десятилетий в разных заповедниках позволили предложить вербальную реконструкцию лесо-лугово-степной общности сообществ, способной поддерживать огромное флористическое и фаунистическое разнообразие, в том числе и много видов деревьев и кустарников, в современном покрове способных существовать на опушках, создаваемых человеком (Vera, 2000; Восточноевропейские леса…, 2004).
Наблюдения за представителями этих групп, сохранившихся в ООПТ, позволили охарактеризовать их средопреобразующую деятельность. Элементарные популяции зубров, туров, тарпанов и других крупных стадных копытных занимали бассейны малых рек площадью в сотни кв. км, используя как водораздельные, так и долинные ландшафты. В результате их жизнедеятельности лесной покров был пронизан луговыми и луговостепными полянами, по берегам водоемов формировались водопойные поляны; вся территория была пронизана сетью троп, которые служили каналами миграции для многих подчиненных видов. Тщательные наблюдения за зубрами на протяжении многих десятилетий в разных заповедниках позволили предложить вербальную реконструкцию лесо-лугово-степной общности сообществ, способной поддерживать огромное флористическое и фаунистическое разнообразие, в том числе и много видов деревьев и кустарников, в современном покрове способных существовать на опушках, создаваемых человеком (Vera, 2000; Восточноевропейские леса…, 2004).
Элементарные популяции бобров занимали долины малых рек площадью в десятки кв. км. В результате их жизнедеятельности долины малых рек и ручьев были преобразованы в совокупности прудов, каналов, низинных болот, заболоченных лесов, приречных луговых полян; появлялись новые микроместообитания: норы, хатки, тропы и пр. Это разнообразие местообитаний (от микро до мезо-), формирующихся в пределах элементарной популяции бобра европейского, поддерживает огромное разнообразие животных, растений и микроорганизмов. Многолетние наблюдения за процессом восстановления природного ареала бобра европейского на территории Северной Евразии существенно дополнили вербальную модель лесо-лугово-степной общности сообществ, включив в нее долинные комплексы “бобровых ландшафтов” (Смирнова, 1998; Восточноевропейские леса…, 2004; Завьялов, 2008).
 Элементарные популяции лиственных и хвойных деревьев разных видов занимали разные позиции в бассейнах малых рек и разные местообитания, создаваемые ключевыми видами животных. Размеры площадей элементарных популяций деревьев разных видов не превышали 1 км2 . Изучение особенностей популяционной биологии деревьев разных видов в ООПТ позволило выявить общие черты их средопреобразующей деятельности. В процессе жизни и смерти от старости или повреждения их насекомыми и/или грибами они создают разнообразные местообитания. В подкроновом пространстве взрослых деревьев в результате затенения, опада, изменения режима влажности и пр. формируются фитогенные поля, которые представляют собой специфические микроместообитания для многих подчиненных видов в наземном слое и в почве.
Элементарные популяции лиственных и хвойных деревьев разных видов занимали разные позиции в бассейнах малых рек и разные местообитания, создаваемые ключевыми видами животных. Размеры площадей элементарных популяций деревьев разных видов не превышали 1 км2 . Изучение особенностей популяционной биологии деревьев разных видов в ООПТ позволило выявить общие черты их средопреобразующей деятельности. В процессе жизни и смерти от старости или повреждения их насекомыми и/или грибами они создают разнообразные местообитания. В подкроновом пространстве взрослых деревьев в результате затенения, опада, изменения режима влажности и пр. формируются фитогенные поля, которые представляют собой специфические микроместообитания для многих подчиненных видов в наземном слое и в почве.
Смерть крупных деревьев часто сопровождается педотурбациями, и формируются совокупности микроместообитаний – ветровально-почвенные комплексы (ВПК). Постоянное отмирание одного или небольших групп деревьев приводит к формированию “окон” (gaps) в пологе леса – специфических местообитаний, где развиваются подрост деревьев, кустарников, крупные травы и пр. Постоянное формирование “окон” в разновозрастных лесах создает мозаику скоплений деревьев разных видов и разных этапов развития (возрастные парцеллы).
С позиций концепции иерархического континуума минимальную площадь выявления природных экосистем на разных этапах развития лесного покрова условно можно определить, исходя из размеров элементарной популяции наиболее мощного ключевого вида; в доисторических лесах такими видами были крупные стадные животные-фитофаги. Границы между разными экосистемами с этих позиций условно можно провести на основе смены состава ключевых видов в пределах одной и той же функциональной группы. Следствием понимания значимости популяционных мозаик ключевых видов в жизни природных объектов стало создание концепции мозаично-циклической организации экосистем (The Mosaic-Cycle Concept…, 1991). Суть ее в следующем: устойчивое существование экосистемы достигается за счет комплементарности иерархически организованных популяционных мозаик ключевых и подчиненных видов разных функциональных групп и трофических уровней.
Использование основных концепций синэкологии позволяет выявить принципиальные отличия условно конструируемых доисторических и современных экосистем лесного пояса. Доисторические экосистемы представляли собой иерархически организованные множества элементарных популяций ключевых видов разных функциональных групп и сопутствующих им элементарных популяций подчиненных видов разных трофических групп. Возможность устойчивого совместного существования этих множеств видов была основана на принципе комплементарности – пространственно-временном разделении ресурсов.
Минимальная площадь доисторических экосистем лесного пояса Северной Евразии и Северной Америки включала водораздельные и долинные ландшафты бассейнов малых рек или ручьев и предположительно составляла сотни км2. В континуальном лесном покрове границы между ними можно было провести лишь условно, по изменению видового состава ключевых видов.
Минимальная площадь современных экосистем, лишенных ключевых видов животных и части видов деревьев, занимает небольшие части водоразделов или долин в пределах бассейнов малых рек или ручьев. Границы между ними, как правило, обусловлены предшествующими антропогенными воздействиями. Принцип комплементарности не реализуется полностью, что обусловлено неполночленностью групп ключевых видов.
Модельная реконструкция лесного покрова Восточной Европы
Обобщение палеонтологических, археологических и исторических материалов на основе современных представлений теоретической синэкологии позволило создать серию реконструкций лесного покрова с конца плейстоцена до современности. В качестве модельной территории выбрана Восточная Европа (Восточноевропейские леса…, 2004; Смирнова и др., 2006).
Конец плейстоцена (поздний вюрм) – древний голоцен (поздний палеолит) – (40–10 тыс. л.н.). На территории Восточной Европы, как и на территории Северной Евразии и Северной Америки, господствовали травяные (пастбищные) сообщества с фрагментами лесов, где ключевыми видами были представители мамонтового комплекса, в первую очередь – мамонт, наиболее крупный из них. Средопреобразующее воздействие этих животных приводило к подавлению древесной растительности и к формированию травяных сообществ с высокой продуктивностью (Пучков, 1992; Кузьмина, 1997; Смирнова и др., 2006). Радиоуглеродное датирование и анализ ареалов этих видов показывает, что как в периоды потеплений, так и в периоды похолоданий устойчиво существовали пастбищные сообщества с характерной для них смешанной флорой и фауной (Восточноевропейские леса…, 2004).
Учитывая особенности фауны и флоры, растительный покров рассматриваемого периода можно охарактеризовать как мозаичный лесолугово-степной. Причем господство пастбищных сообществ было обусловлено не только неблагоприятными для древесных видов климатическими условиями, но и, главным образом, подавлением древесной растительности наиболее крупными представителями мамонтовой фауны.
 В конце рассматриваемого периода в результате охотничьей деятельности численность ключевых видов мамонтового комплекса сократилась ниже критического уровня. Вымирание одних и сокращение ареалов и численности других ключевых видов обусловило начало экспансии древесной растительности и сокращения площадей травяных сообществ, чему благоприятствовало незначительное потепление климата (Климанов, 1989).
В конце рассматриваемого периода в результате охотничьей деятельности численность ключевых видов мамонтового комплекса сократилась ниже критического уровня. Вымирание одних и сокращение ареалов и численности других ключевых видов обусловило начало экспансии древесной растительности и сокращения площадей травяных сообществ, чему благоприятствовало незначительное потепление климата (Климанов, 1989).
Пионерами залесения были виды деревьев r-стратегии с легколетучими семенами: ивы, березы, а также осина, сосна обыкновенная, лиственница сибирская (Нейштадт, 1957; Хотинский, 1989). Таким образом, с конца позднего вюрма начинается процесс смены ключевых видов: гиганты и крупные виды фитофагов мамонтового комплекса уступают свои позиции древесным видам, что заложило основу формирования современного лесного пояса (Антипина, 1994).
 Ранний голоцен – мезолит (10–7 тыс. л.н.) и средний голоцен – неолит и бронза (7–2.5 тыс. л.н.). Начало этого периода характеризуется заметным потеплением климата на фоне периодических похолоданий и потеплений (Климанов, 1989). В процессе трансформации пастбищных экосистем в детритные вслед за пионерными видами, характеризующимися самыми большими по сравнению с другими видами деревьев скоростями расселения (Удра, 1988), широко распространились К-стратеги – виды темнохвойных и широколиственных деревьев. Изменение соотношения травяных и древесных сообществ в пользу древесных положило начало формированию целостного лесного пояса на большей части Восточной Европы за исключением самых северных территорий. В свою очередь, этот процесс способствовал повышению численности лесных животных: белки, бобра, бурого медведя, кабана, лося и других видов копытных, для которых расширение площадей лесов было благоприятным.
Ранний голоцен – мезолит (10–7 тыс. л.н.) и средний голоцен – неолит и бронза (7–2.5 тыс. л.н.). Начало этого периода характеризуется заметным потеплением климата на фоне периодических похолоданий и потеплений (Климанов, 1989). В процессе трансформации пастбищных экосистем в детритные вслед за пионерными видами, характеризующимися самыми большими по сравнению с другими видами деревьев скоростями расселения (Удра, 1988), широко распространились К-стратеги – виды темнохвойных и широколиственных деревьев. Изменение соотношения травяных и древесных сообществ в пользу древесных положило начало формированию целостного лесного пояса на большей части Восточной Европы за исключением самых северных территорий. В свою очередь, этот процесс способствовал повышению численности лесных животных: белки, бобра, бурого медведя, кабана, лося и других видов копытных, для которых расширение площадей лесов было благоприятным.
В раннем голоцене, как и в предшествующий период, практически на всей территории распространения лесов отмечено совместное обитание видов темнохвойных (ель и пихта) и широколиственных (дуб, лип, вяз и др.) деревьев. Позже, в среднем голоцене, практически на всей территории Восточной Европы формируется своеобразный лесо-лугово-степной (хвойно-широколиственный) пояс, в котором постепенно уменьшается доля широколиственных видов деревьев и неморальных трав с юга лесного пояса на север и темнохвойных видов деревьев и бореальных трав и кустарничков в противоположном направлении.
 В то же время в южной полосе лесного пояса до начала железного века, а местами и до средневековья, продолжали обитать различные виды крупных стадных копытных животных. Это позволяет реконструировать растительный покров данного времени как мозаику лесных участков, чередующихся с луговыми или степными участками зоогенного происхождения (Восточноевропейские леса…, 2004; Смирнова и др., 2006).
В то же время в южной полосе лесного пояса до начала железного века, а местами и до средневековья, продолжали обитать различные виды крупных стадных копытных животных. Это позволяет реконструировать растительный покров данного времени как мозаику лесных участков, чередующихся с луговыми или степными участками зоогенного происхождения (Восточноевропейские леса…, 2004; Смирнова и др., 2006).
Наличие травяных (луговых и степных), лесных сообществ и экотонов в пределах доисторических экосистем определяло возможность приживания и устойчивого существования в растительном покрове светолюбивых видов деревьев, кустарников и трав. Среди светолюбивых видов деревьев, не возобновляющихся в современных лесах, особенно значимы такие мощные средопреобразователи, как дуб черешчатый, сосна обыкновенная, лиственница сибирская. На большей части лесного пояса в этот период были найдены костные остатки бобра, что свидетельствует о широком распространении водных и прибрежно-водных сообществ, формирующихся вследствие строительной деятельности этих животных. Значительно более широкое распространение гидроморфных ландшафтов и, вследствие этого, значительно более высокий уровень влажности почв и воздуха в лесном поясе были серьезным препятствием к распространению пирогенных ландшафтов (Тюрюканов, 1990). Видимо, это было одной из существенных причин далекого проникновения на юг Восточной Европы видов темнохвойных деревьев (ели и пихты), и, следовательно, поддержания единого лесо-лугово-степного (хвойно-широколиственного) пояса.
С середины среднего голоцена оформляется мощнейший фактор воздействия на лесной покров: хозяйство производящего типа (земледелие, скотоводство, выплавка металлов). Оно начало распространяться в Восточной Европе 6.0 – 5.5 тыс. л. н. и охватило большую часть территории на протяжении бронзового века (4.6 – 3.2 тыс. л. н.). В это время в остеологическом материале сильно сократилась доля костей диких копытных (зубра, тура, тарпана и др.) и увеличилась доля костей домашнего скота (Цалкин, 1966), а в спорово-пыльцевых спектрах появилась пыльца культурных злаков (Крупенина, 1973; Серебрянная, 1980).
Хозяйство производящего типа принципиально изменило структуру лесного покрова. В первую очередь на таких территориях исчезают крупные стадные копытные и бобры, уже не столько в силу непосредственного преследования, сколько из-за уничтожения необходимых для их жизни местообитаний. В результате доля природных пастбищных сообществ сильно сокращалась, а доля детритных возрастала. Все более ограниченной становилась трофическая и топическая деятельность крупных стадных копытных и бобров, в значительной степени определявших возможность существования светолюбивых видов деревьев (в первую очередь дуба, сосны, лиственницы) и кустарников, а также всех светолюбивых видов растений иных жизненных форм и многих видов животных открытых местообитаний. Их существование прямо (посадки деревьев) или косвенно (распашки, выжигания, рубки с нарушением напочвенного покрова, выпас в лесу, сбор подстилки, осушение, сооружение искусственных водоемов и пр.) стало определяться хозяйственной деятельностью.
Только теневыносливые виды широколиственных и темнохвойных деревьев, кустарников, кустарничков и трав оказались способны в таких условиях поддерживать устойчивый поток поколений без антропогенного содействия. Сформировались сообщества “теневых” хвойно-широколиственных лесов. К концу среднего голоцена выжигание лесов в цикле подсечно-огневого земледелия сильно отодвинуло на север южную границу лесного пояса. Распространение кочевого скотоводства на юге Восточной Европы привело к формированию в конце среднего голоцена степной и полупустынно-степной зон, аридизации климата и уменьшению смешанности флоры и фауны юга Восточной Европы.
Эти события были крупным шагом к формированию современной зональности, в том числе таежной (бореальной) полосы, и они, вероятно, оказали существенное влияние на изменения макроклимата Евразии в целом. Возможно, они стали одной из причин нарастания нестабильности климата второй половины голоцена (Климанов, 1989; Хотинский, 1989).
Поздний голоцен в целом (2500–0 л. н.) Железный век и раннее средневековье (2.5–0.5 тыс. л. н.). Анализ палеоботанических данных показывает, что в начале позднего голоцена, в раннежелезном веке (2.5 – 1.6 тыс. л. н.), ареалы видов деревьев не претерпели существенных изменений. Так же, как и в среднем голоцене, виды широколиственных и темнохвойных деревьев встречались вместе на большей части лесного пояса. Однако оценка количественного участия видов в спорово-пыльцевых спектрах показывает сокращение доли темнохвойных деревьев на юге, а широколиственных – на севере (Нейштадт, 1957; Хотинский, 1989). Этот процесс был обусловлен расширением степной зоны на север и усилением сухости почв и воздуха вследствие антропогенной аридизации юга Восточной Европы. В это же время главным образом в результате подсечно-огневого земледелия существенно отступили на юг северные границы ареалов широколиственных видов деревьев, что положило начало формированию современной тайги. Одновременно на массивах песчаных почв лесного пояса были сформированы специфические пирогенные леса с господством сосны обыкновенной. Подсечно-огневое, а затем переложное и пахотное земледелие, выпас в лесу, сбор подстилки и валежа и прочие традиционные методы лесопользования привели к деградации почвенного покрова на больших площадях (Бобровский, 2010).
Выжигание лесов на их северной границе в цикле домашнего оленеводства привело в позднем голоцене к развитию зоны тундр на месте северной тайги. На севере лесного пояса антропогенные факторы и изменения климата в позднем голоцене действовали однонаправленно. Они приводили к разрушению лесного пояса и способствовали отступлению на юг северотаежных видов и их комплексов. Этот процесс можно назвать “бореализацией” лесного пояса.
Таким образом, изменение в позднем голоцене дендрофлоры и фауны, начатое еще в среднем голоцене, привело к расчленению единого лесолугово-степного (хвойно-широколиственного) пояса на две принципиально отличные группы сообществ (Смирнова, 1998):
1) способные поддерживать себя;
2) требующие для своего поддержания постоянных антропогенных воздействий (пойменные и суходольные луга, луговые степи, леса из пионерных видов деревьев: березы, сосны обыкновенной, лиственницы). Благодаря антропогенной деятельности поддерживалась мозаика лесного покрова, в котором существовали как светолюбивые виды деревьев, так и виды травяных сообществ.
Одновременно был сделан завершающий шаг в формировании зональности: антропогенно обусловленное расчленение лесного пояса Восточной Европы на полосы темнохвойных (бореальных), хвойно-широколиственных (бореально-неморальных) и широколиственных (неморальных) лесов.
Параллельно с изменениями растительности в течение голоцена изменяется и животный мир:
1) вымирает полностью или в пределах Палеарктики значительная часть животных ключевых видов;
2) сокращаются ареалы многих видов животных, в первую очередь наиболее крупных и активно промышляемых;
3) исходно единая фауна по мере расчленения растительности лесного пояса расчленяется на фауны тундр, лесов и степей (Смирнова и др., 2006).
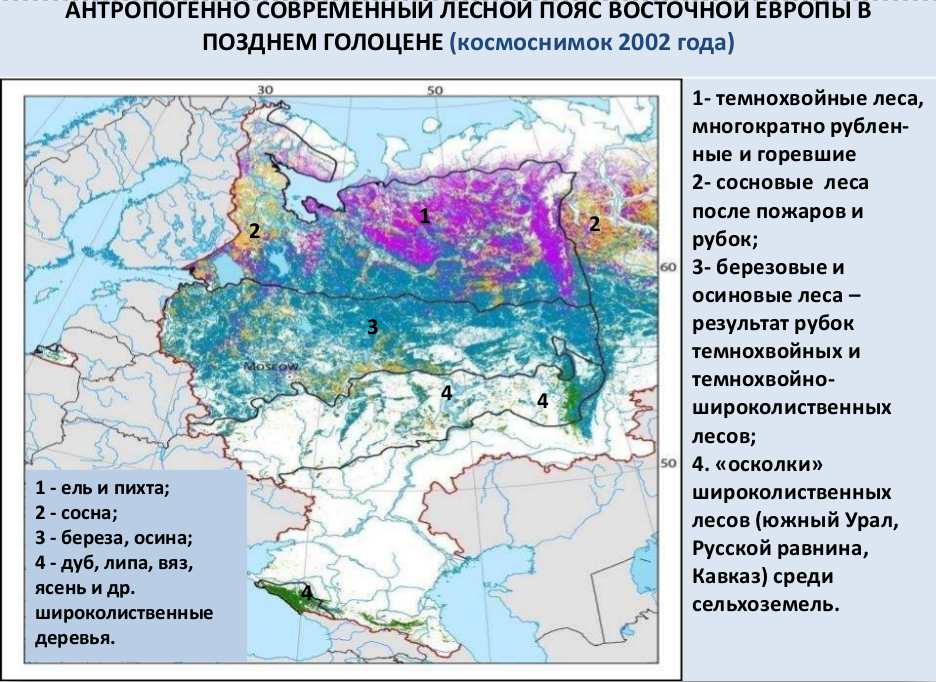
Современность (0.5–0 тыс. л .н.). В последние столетия, особенно в XIX и XX вв., произошли дальнейшие преобразования лесного покрова Восточной Европы в связи с изменениями способов природопользования и все усиливающейся индустриализацией хозяйства в целом. Полосы неморальных (широколиственных) и неморально-бореальных (хвойно-широколиственных) лесов в связи с интенсивным земледельческим освоением и с высокой скоростью оборотов рубок практически превратились в единую полосу, где мелколиственные леса на суглинистых субстратах и посадки сосны на песчаных субстратах чередуются с сельскохозяйственными землями. В то же время леса с господством зональных ключевых видов занимают небольшие площади (Восточноевропейские леса…, 2004).
В полосе бореальных (таежных) лесов в связи с многократными (антропогенно инициированными) пожарами, сокращением сельхозпользования из-за его неэффективности и падением численности населения сформировалась мозаика пирогенных и пасквальных сообществ разного времени возникновения: от молодых сосняков и березняков до старых и разновозрастных ельников и пихто-ельников. При этом значительная часть травяных сообществ, поддерживаемых в таежных лесах человеком с середины – конца среднего голоцена, исчезла в последние столетия в связи с катастрофическим сокращением численности населения.
В настоящее время значительная часть таежных лесов находится на начальных этапах сукцессий, и их древесные синузии представлены или раннесукцессионными (пионерными) видами деревьев (сосна, береза), или первым поколением позднесукцессионных видов (ель, реже пихта) и отличаются минимальным видовым и структурным разнообразием. К ним относятся ельники и пихто-ельники зеленомошные, зеленомошно-черничные и кисличные. Эти наиболее широко распространенные типы восточноевропейских темнохвойных лесов (Растительный покров СССР, 1956), сформированные и поддерживаемые антропогенной деятельностью (Восточноевропейские леса…, 2004), часто рассматриваются как природные образования.
Одновременно с антропогенным упрощением состава лесов к XVI–XIX вв. в дикой природе практически исчезают такие виды, как тур, зубр, тарпан, максимально сокращаются ареалы всех крупных млекопитающих, бобра и ряда других видов животных.
Таким образом, проведенные модельные палеореконструкции и анализ литературы по природопользованию позволяют утверждать, что основным фактором формирования и развития лесного покрова Восточной Европы в течение голоцена была средопреобразующая деятельность человека. Она проявлялась и проявляется в настоящее время:
1) в изменении границ ареалов ключевых видов животных в связи с их уничтожением на части ареалов, а также с антропогенно инициированными изменениями экотопов и климата, в первую очередь с аридизацией на юге лесного пояса и бореализацией на его севере;
2) в разрушении функциональных связей между ключевыми видами растений и животных,которые обеспечивали устойчивые потоки поколений в популяциях подчиненных видов разных трофических групп.
Широко распространенные в настоящее время варианты лесных сообществ, в том числе и в лесах ООПТ, отражают в своем составе и структуре способы природопользования, существовавшие до начала – середины ХХ в. (Восточноевропейские леса…, 2004; Vera et al., 2006; Кюстер, 2012).
Осознание этого чрезвычайно важного заключения принципиально меняет взгляды на охрану природы. Очевидно, что период пассивной охраны на большей части территории Земли должен смениться научно обоснованным содействием по восстановлению потенциальной биоты на основе существующих и вновь организуемых ООПТ. Только восстановление потенциальной биоты и формирование всеохватных сетей ООПТ даст возможность разработать комплексные модели природопользования, решающие задачи получения ресурсов и поддержания максимально возможного (в конкретных условиях) биоразнообразия.
Разработка программ по восстановлению потенциальной биоты ООПТ и организации экосистемного природопользования
Ко времени планомерных исследований растительности и животного мира (XIX–XX вв.) лесной пояс России в ходе традиционного природопользования был существенно преобразован. Неверная трактовка антропогенно преобразованного лесного пояса как природного привела к представлениям об абсолютной экотопической и климатической обусловленности состава, строения и биоразнообразия таких лесов, где следы предшествующей антропогенной деятельности не выявляются без тщательных исследований истории их формирования и развития.
Изменения представлений о природном облике лесного пояса произошли во второй половине ХХ в. в связи с большим объемом новых данных о палеоистории лесов; о временах и способах природопользования; о восстановлении природных ареалов ключевых видов животных и растений (Восточноевропейские леса…, 2004; Vera, 2004; Vera et al., 2006; Бобровский, 2010; Кюстер, 2012).
Модельная реконструкция лесного пояса Восточной Европы, полученная переосмыслением новых данных с позиций современных представлений синэкологии, позволяет предложить принципиально новый подход к решению проблемы оптимизации функции биоразнообразия лесного покрова.
На первом этапе на основе существующей сети ООПТ необходимо выяснить:
1) в какой степени каждая ООПТ и существующая система ООПТ в целом отражает биоразнообразие потенциального лесного покрова анализируемой территории;
2) какие меры необходимо принять для восстановления потенциального биоразнообразия в тех ООПТ, где его уровень ниже потенциально возможного;
3) как расширить систему ООПТ локального и регионального уровней и создать сеть рефугиумов потенциального биоразнообразия как источников биоразнообразия для эксплуатируемых лесов.
Следует отметить, что в настоящее время нет единой классификации ООПТ, которая учитывала бы наличие в них ключевых видов животных и растений, состояние их элементарных популяций, их размеры, степень антропогенной нарушенности территории в прошлом и настоящем, естественные потенции самовосстановления, а также состояние природных комплексов за их пределами.
Методология и методы оценки современного состояния ООПТ с позиций современной синэкологии изложены в монографиях (Мониторинг…, 2008; Методологические подходы…, 2010) и многочисленных публикациях (Краснитский, 1983; Мишнев, 1984; Смирнова, 2010). Однако обобщение этой огромной литературы с целью разработки локальных и региональных программ оценки и восстановления потенциального биоразнообразия – абсолютно необходимая и весьма трудоемкая задача.
В процессе разработки алгоритма оценки степени реализации экосистемной функции биоразнообразия в первую очередь необходимо провести комплексные исследования ООПТ, чтобы установить:
1) где восстанавливаются элементарные популяции крупных стадных копытных и/или бобров и устойчиво существуют элементарные популяции регионального пула видов теневыносливых и светолюбивых деревьев, кустарников, трав;
2) где сочетание ключевых видов деревьев, а также подчиненных видов растений и животных поддерживается за счет имитации режима традиционного природопользования (выпас домашних животных, сенокошение и пр.);
3) где сохранились фрагменты растительности, представители животного мира и других царств, характерные для других современных климатических зон (участки северо-таежных лесов в тундре, широколиственных лесов в степи и пр.).
Полученные данные, помимо большой значимости для типизации ОППТ, послужат основой для разработки комплексных программ природопользования, ориентированных на реализацию экосистемных функций в целом.
Несомненно, что для реализации поставленных задач требуется создание единой государственной программы, где были бы отражены все задачи, которые необходимо решить для квалифицированной оценки возможностей ООПТ восстановить потенциальное биоразнообразие, определить необходимые действия по его восстановлению и длительность этого процесса, а также оценить необходимые материальные затраты.
На втором этапе необходимо определить, какие принципиальные изменения лесопользования и лесоуправления необходимо произвести, чтобы поэтапно приблизиться к поставленной задаче – оптимизации экосистемных функций и в первую очередь функции биоразнообразия как наиболее доступной для решения при современном уровне знаний.
Общие подходы и конкретные методы экологически ориентированного лесопользования изложены как в классических, так и современных работах. Часть их успешно реализуется на практике. Однако решение проблемы оптимизации экосистемных функций лесного пояса России в целом требует перехода от лесопользования к природопользованию, т.е. сохранению всей потенциальной биоты и устойчивому использованию ее ресурсной части. Переход ко второму этапу потребует значительного времени, ресурсов и в первую очередь усилий для осознания невозможности восстановления и поддержания экосистемных функций лесного пояса в целом без использования принципиально новых теоретических представлений современной синэкологии о равноценной роли ключевых видов животных и растений в этом процессе.
Традиционное лесное хозяйство, основанное на проведении сплошных рубок и создании лесных монокультур на обширных территориях, приводит к снижению биоразнообразия, возрастанию риска повреждения культур в результате вспышек размножения патогенных микроорганизмов и энтомовредителей, уменьшению почвенного плодородия, снижению почвозащитных и водоохранных функций (Восточноевропейские леса…, 2004).
Очевидна необходимость разработки таких способов ведения лесного хозяйства, которые смогли бы обеспечить формирование разновозрастных полидоминантных лесов, отличающихся высоким уровнем биоразнообразия. Экспериментальные работы в этом направлении проводили в природно-историческом заповеднике “Горки” (Московская обл.) в 1988–1997 гг. в послепахотных березняках Богдановского лесопарка, в которых занос семян позднесукцесионных видов крайне ограничен большим расстоянием (Сукцессионные процессы…, 1999). В настоящее время под пологом березовых лесов формируется кустарниковый ярус из кустарников (лещины, крушины, рябины и других видов). В результате проведения рубок переформирования были созданы “окна” размерами 0.16–0.25 га, в которые высаживались зональные эдификаторы (дуб черешчатый, липа мелколистная, клен остролистный, ясень обыкновенный, ель европейская). Описание лесоводственных экспериментов и предварительные результаты опубликованы (Сукцессионные процессы…, 1999).
Опыт проведения экспериментов и анализ лесоводственной литературы дают возможность предложить наиболее важные направления работ по восстановлению лесов:
1) восстановление структурного разнообразия (системы разновозрастных мозаик окон возобновления) путем проведения группово-выборочных рубок. Имеющиеся расчеты и данные показывают, что оптимальные размеры окон составляют по диаметру 1.5–2 высоты окружающего древостоя (0.1–0.3 га);
2) восстановление видового разнообразия должно базироваться на естественном возобновлении в сочетании с созданием лесных культур недостающих древесных видов. Для целого ряда редких видов трав, отличающихся малым радиусом репродуктивной активности, восстановление их популяций целесообразно проводить путем реинтродукции; 3) восстановление генетического разнообразия популяций древесных видов. При закладке питомников необходимо использовать гетерогенный семенной материал, собранный из местных популяций древесных видов.
Восстановление полидоминантных разновозрастных лесов поможет обеспечить длительное неистощительное лесопользование, а также будет способствовать поддержанию высокого биоразнообразия лесных экосистем, сохранению и восстановлению почвенного плодородия, увеличению устойчивости лесных экосистем. Эти мероприятия мы рассматриваем, как необходимый первый шаг по пути оптимизации функции биоразнообразия в эксплуатационных лесах.
Заключение
Потребность в разработке единой стратегии охраны природы и природопользования, направленной на максимальную реализацию экосистемных функций живого покрова Земли как основы устойчивого развития цивилизации, из области теоретических построений становится насущной необходимостью. Огромная сложность этой проблемы требует разработки системы программ на государственном уровне путем объединения усилий специалистов, изучающих и сохраняющих лесной покров, и специалистов, использующих лесные ресурсы и управляющих ими. Реализовать разработанные программы можно поэтапно.
На первом этапе на основе имеющихся данных надо создать серии модельных реконструкций потенциального лесного покрова на разных уровнях – от локального до континентального. На втором этапе на основе разработанных алгоритмов оценки степени реализации экосистемных функций следует провести обследования существующих ООПТ и оценить их современные возможности поддержания экосистемных функций и динамические тренды. На третьем этапе надо разработать меры содействия наиболее полной реализации экосистемных функций сетями региональных и континентальных ООПТ (создание новых ООПТ, расширение территорий существующих ООПТ, реинтродукция природных ключевых и подчиненных видов). На четвертом этапе необходимо использовать разработанные алгоритмы оценки степени реализации экосистемных функций эксплуатируемыми лесами и начать поэтапный переход от лесопользования к природопользованию в целом.
Список литературы
Антипина Е.Е., Маслов С.П. Некоторые проблемы изучения истории взаимодействия хозяйственной деятельности человека с природными биоценозами // Эволюционная и историческая антропоэкология. М.: Наука, 1994. С. 111–120.
Бобровский М.В. Лесные почвы европейской России: биотические и антропогенные факторы формирования. М.: Тов-во КМК, 2010. 359 с.
Восточноевропейские леса: история в голоцене и современность / Под. ред. О.В. Смирновой. Кн. 1–2. М.: Наука, 2004. Кн. 1. 479 с.; Кн. 2. 575 с.
Завьялов Н.А. Бобры – ключевые виды и экосистемные инженеры // Экосистемы малых рек: биоразнообразие, экология, охрана. Лекции и материалы докладов Всероссийской школы-конференции. Институт биологии внутренних вод им. И.Д. Папанина РАН. 18–21 ноября 2008 г. Ярославль: Изд-во “Ярославский печатный двор”, 2008. С. 4–24.
Злотин Р.И., Ясный Е.В. Глобальная деградация биологического разнообразия // Изв. АН СССР. Сер. Геогр. 1992. No 2. С. 76–88.
Климанов В.А. Цикличность и квазипериодичность климатических колебаний в голоцене // Палеоклиматы позднеледниковья и голоцена. М.: Наука, 1989. С. 29–33.
Краснитский А.М. Проблемы заповедного дела. М.: Лесн. пром-сть, 1983. 191 с.
Крупенина Л.А. Признаки антропогенного влияния на растительный покров центральной части Среднерусской возвышенности в голоцене // Палинология голоцена и маринология. М.: Наука, 1973. С. 91–97.
Кузьмина И.Е. Лошади Северной Евразии от плиоцена до современности // Тр. Зоол. ин-та РАН. 1997. Т. 273. 223 с.
Кюстер Х. История леса. Взгляд из Германии. М.: Высш. школа экономики, 2012. 304 с.
Методические подходы к экологической оценке лесного покрова бассейна малой реки / Под ред. Л.Б. Заугольновой, Т.Ю. Браславской. М.: Тов-во научн. Изд. КМК, 2010. 383 с.
Мишнев В.Г. Заповедники и принципы жесткой резервации территорий // Ботан. журн. 1984. Т. 69. No 8. С. 1106–1112.
Мониторинг биологического разнообразия лесов / Под ред. А.С. Исаева. М.: Наука, 2008. 453 с.
Нейштадт М.И. История лесов и палеогеография СССР в голоцене. М.: Сов. наука, 1957. 404 с.
Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России / Под ред. Л.Б. Заугольновой. М.: Научный мир, 2000. 185 с.
Павлов Д.С., Букварева Е.Н. Климаторегулирующие функции наземных экосистем и экологоцентрическая концепция природопользования // Успехи современной биологии. 2011. Т. 131. No 4. С. 324–345.
Пучков П.В. Некомпенсированные вюрмские вымирания. Сообщение 2. Преобразование среды гигантскими фитофагами // Вестник зоологии. 1992. No 1. С. 58–66.
Растительный покров СССР. Пояснительный текст к геоботанической карте. М.; Л.: Изд-во АН СССР, 1956. Т. 1. Т. 2.
Реймерс Н.Ф. Надежды на выживание человечества. Концептуальная экология. М.: ИЦ “Россия молодая”, 1992. 367 с.
Серебрянная Т.А. К голоценовой истории лесов запада Среднерусской возвышенности // Бюлл. комиссии по изучению четвертичного периода. 1980. No 50. С. 178–185.
Смирнова О.В. Теоретические основы, система методов и программ стационарных экологических исследований в таежных лесах Урала // Труды Печоро-Илычского заповедника. Вып. 16. Сыктывкар, 2010. С. 157–162.
Смирнова О.В., Калякин В.Н., Турубанова С.А., Бакун Е.Ю. Генезис восточноевропейской тайги в голоцене // Закономерности вековой динамики биогеоценозов. XXI Чтения памяти академика В.Н. Сукачева. М.: Тов-во научн. изд. КМК, 2006. С. 18–65.
Смирнова О.В., Попадюк Р.В., Заугольнова Л.Б., Ханина Л.Г. Оценка потерь флористического разнообразия в лесной растительности (на примере заповедника “Калужские засеки”) // Лесоведение. 1997. No 2. С. 27–42.
Смирнова О.В. Популяционная организация биоценотического покрова лесных ландшафтов // Успехи совр. биологии. 1998. No 2. С. 25–39.
Соколов В.Е., Филонов К.П., Нухимовская Ю.Д., Шадрина Г.Д. Экология заповедных территорий России. М.: Янус-К, 1997. 575 с.
Сукачев В.Н. Растительные сообщества: Введение в фитосоциологию. 4-е изд. М.; Л.: Книга, 1928. 232 с.
Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия / Под ред. О.В. Смирновой, Е.С. Шапошникова. Спб.: РБО, 1999. 549 с.
Тюрюканов А.Н. О чем говорят и молчат почвы. М.: Агропромиздат, 1990. 224 с.
Удра И.Ф. Расселение растений и вопросы палео- и биогеографии. Киев: Наук. думка, 1988. 197 с.
Хотинский Н.А. Дискуссионные проблемы реконструкции и корреляции палеоклиматов голоцена // Палеоклиматы позднеледниковья и голоцена. М.: Наука, 1989. С. 12–17.
Цалкин В.И. Древнее животноводство племен Восточной Европы и Средней Азии. М.: Наука, 1966. 159 с.
Экономика экосистем и биоразнообразия: потенциал и перспективы стран Северной Евразии: Матер. совещ. “Проект TEEB – экономика экосистем и биоразнообразия: перспективы участия России и других стран ННГ”. М., 2010. 136 с.
Braun-Blauquet J., Pavillard J. Vocabulaire de sociologie vegetale. 2 nd ed. Montpellier, 1925. 22 p.
Brzeziecki B., Kienast F., Wildi O. A simulation map of the potential natural forest vegetation of Switzerland // J. Veget. Sci. 1993. V. 4. No 2. P. 149–156.
Ciccarese L., Mattsson A., Pettenella D. Ecosystem services from forest restoration: thinking ahead // New Forests. 2012. V. 43. No 5–6. P. 543–560.
Collins S.L., Glenn S.M., Roberts D.W. The hierarchical continuum concept // J. Veget. Sci. 1993. V. 4. No 2. P. 149–156.
Crain C.M., Bertness M.D. Hastings A., Byers J.E., Crooks J.A., Cuddington K., Jones C.G., Lambrinos J.G., Talley T.S., Wilson W.G. Ecosystem engineering in space and time. Review and synthesis // Ecology Letters. 2008. V. 10. P. 153–164.
Ellenberg H. Vegetation Ecology of Central Europe. Cambridge: Univ. Press, 1988. 731 p.
Feld C.K. Indicators of biodiversity and ecosystem services: a synthesis across ecosystems and spatial scales // Oikos. 2009. V. 118. No 12. P. 1862–1871.
Gamfeldt L., Hillebrand H., Jonsson P.R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning // Ecology. 2008. V. 89. No 5. P. 1223–1231.
Hector , A., Bagchi R. Biodiversity and ecosystem multifunctionality // Nature. 2007. V. 448. P. 188–190.
Hooper D.U., Chapin F.S., Ewel J.J., Hector A., Inchausti P., Lavorel S., lawton J. H., Lodge D.M., Loreau M., Naeem S., Schmid B., Setala H., Symstad A.J., Vandermeer J., Wardlw D.A. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge // Ecol. Monogr. 2005. V. 75. No 1. P. 3–35.
McCarthy J. Gap dynamics of forest trees: A review with particular attention to boreal forest // Environ. Rev. 2001. V. 9. P. 1–59.
MEA–Millennium Ecosystem Assessment Ecosystems and human well-being: biodiversity synthesis. Washington: World Resources Institute, 2005. 86 p.
Schwartz M.W., Brigham C.A., Hoeksema J.D., Lyons K.G., Mills M.H., Mantgem P.J. Linking biodiversity to ecosystem function: implications for conservation ecology // Oecologia. 2000. V. 122. P. 297–305.
The Ecological History of European Forests / Ed. by K.J. Kirby, C. Watkins. Wallingford, UK: CAB International, 1998. 384 p.
The Mosaic-Cycle Concept of Ecosystem / Ed. by H. Remmert. Berlin, Heidelberg, N.Y.: Springer-Verlag, 1991. 168 p.
Tuxen R. Die heutige potentielle naturliche vegetation als Gegenstand der Vegetationskartierung // Angew. Pfl anzensociologie (Stolzenaw). 1956. Bd. 13. S. 4–42.
Vera F.W.M. Grazing Ecology and Forest History. Oxon–New York: CABI Publishing, 2000. 506 p.
Vera F.W.M., Bakker E., Olff H. Large herbivores: missing partners of western European light-demanding tree and shrub species? // Large Herbivore Ecology, Ecosystem Dynamics and Conservation / Ed. by K. Danell, P. Duncan, R. Bergstrom, J. Pastor. Cambridge: Cambridge University Press, 2006. P. 203–238.
ЛЕСОВЕДЕНИЕ, 2015, No 5, с. 367–378
Рекомендуем прочесть
 Основные концепции и методы восстановления природных лесов Восточной Европы
Основные концепции и методы восстановления природных лесов Восточной Европы «Город как заказник»: (пока?) упускаемая возможность
«Город как заказник»: (пока?) упускаемая возможность Экологический риск заповедания и пути его преодоления: обзор современных представлений
Экологический риск заповедания и пути его преодоления: обзор современных представлений Разбор популярных мифов про охрану природы
Разбор популярных мифов про охрану природы Рельеф, реки, луга и болота Москвы
Рельеф, реки, луга и болота Москвы Теоретические основы единой стратегии охраны природы и природопользования
Теоретические основы единой стратегии охраны природы и природопользования Климатогенная роль городского озеленения
Климатогенная роль городского озеленения Первый класс и подонки общества
Первый класс и подонки общества
