
 Резюме. Для трёх групп птиц-«производителей» дупел — дятлов, поползней и гаичек — Wiebe et al. (2007) показана большая выгодность гнездования в прошлогодних дуплах сравнительно с долблением новых. Данные пары раньше гнездятся, у них выше величина кладки и т. д. Однако повторно использовать дупла возможно не везде: нужно многолетнее постоянство участков обитания, внутри которого в известных владельцу местах (там ежегодно подыскивается место для гнезда) копятся старые дупла, и нужно, чтобы последние не удалялись при уходе за лесом, не ликвидировались отдыхающими, не занимались скворцами и другими «пользователями», которых владельцы не в состоянии вытеснить и пр. Для специализированных видов, вроде трёхпалого дятла, первое наблюдается лишь в малонарушенных лесах (где и фиксируется необычно высокий — почти четверть! — процент гнездования в старых дуплах, и больший успех размножения подобных пар), для видов-генералистов, вроде желны и большого пёстрого дятла — в более широком спектре лесных ландшафтов, в том числе в «полуприродных» лесах.
Резюме. Для трёх групп птиц-«производителей» дупел — дятлов, поползней и гаичек — Wiebe et al. (2007) показана большая выгодность гнездования в прошлогодних дуплах сравнительно с долблением новых. Данные пары раньше гнездятся, у них выше величина кладки и т. д. Однако повторно использовать дупла возможно не везде: нужно многолетнее постоянство участков обитания, внутри которого в известных владельцу местах (там ежегодно подыскивается место для гнезда) копятся старые дупла, и нужно, чтобы последние не удалялись при уходе за лесом, не ликвидировались отдыхающими, не занимались скворцами и другими «пользователями», которых владельцы не в состоянии вытеснить и пр. Для специализированных видов, вроде трёхпалого дятла, первое наблюдается лишь в малонарушенных лесах (где и фиксируется необычно высокий — почти четверть! — процент гнездования в старых дуплах, и больший успех размножения подобных пар), для видов-генералистов, вроде желны и большого пёстрого дятла — в более широком спектре лесных ландшафтов, в том числе в «полуприродных» лесах.
[Век живи — век учись, этот момент биологии дятлов я осознал только недавно. См. далее перевод статьи Тимо Паккала с соавт. (2017) про трёхпалого дятла. Сам это не видел именно потому что долговременно прослеживал птиц лишь в подмосковных лесах, а они слишком нарушенные. Прим.публикатора]
***
Введение
Содержание
- 1 Введение
- 2 Результаты
- 2.1 Частота повторного использования дупел и обилие многодупельных деревьев
- 2.2 Роль многодупельных деревьев в повторном использовании дупел
- 2.3 Даты гнездования при повторном использовании дупел и в многодупельных деревьях.
- 2.4 Успех размножения при использовании старых дупел и в многодупельных деревьях
- 3 Повторное использование гнёзд/гнездовых деревьев и успех гнездования на уровне территории. Обсуждение
- 3.1 Долбить заново или использовать повторно?
- 3.2 Распространение повторного использования дупел/гнездовых деревьев у трёхпалых дятлов и их родственников
- 3.3 Повторное использование дупел и гнездовые деревья трёхпалых дятлов
- 3.4 Повторное использование дупел на уровне территории и популяции вида: адаптация к существованию в динамичном лесном ландшафте.
Timo Pakkala et al. (2017) в 1987-2015 гг. прослеживали использование дупел трёхпалыми дятлами, обитавшими на территории 170 км2 в южной Финляндии.
Прослежено использование 520 гнездовых деревьев, 645 активных гнездовых дупел (на одном дереве их может быть несколько) и 833 случая гнездования на 86 территориях пар, в том числе 211 случай повторного использования дупел (195 — старых дупел также трёхпалого дятла, 11 — большого пёстрого, 5 — повешенных дуплянок). 25% гнездований происходили в ранее использовавшихся дуплах, 28% дупел и 25% гнездовых деревьев использовались более одного раза. Повторное использование дупел повышает репродуктивный успех, способствует более раннему размножению пары в год, следующий за первым гнездованием в данном месте. Стволы с несколькими гнездовыми дуплами повторном использовались в 15% случаев; 62% случаев повторного используемых дупел приходилось на такие деревья. Эти данные контрастируют с расхожим мнением, что дятлы — и особенно виды — специалисты вроде трёхпалого — выдалбливают гнездовое дупло каждый год заново. До этой работы повторное использование дупел этим видом никем отмечено не было.
Использование старых дупел для гнездования и многодупельные деревья наиболее распространены в наилучших стабильных во времени местообитаниях вида, где возможно многолетнее использование одной и той же территории, со стабильным маршрутом обследования и пр. Он предпочитает спелые / перестойные хвойные и смешанные леса, с обилием мёртвой древесины (как сухостоя, так и упавших стволов) и отмирающих деревьев (Hess 1983, Pechacek 1995, Ruge et al. 1999, Bütler et al. 2004).
В бореальных лесах к повторному использованию дупел и гнездовых деревьев «подталкивает» короткий сезон размножения, «требующий» экономить время и силы. Район исследований общей площадью 170 км2 находился в муниципалитетах Хямеенлинна, Падасйоки и Асиккала на юге Финляндии (около 61°15 северной широты, 25°03 восточной долготы; рис. 1), в подзоне южной тайги. Там преобладают зрелые хвойные леса (в основном коммерческие, т. е. управляемые, а не развивающиеся в спонтанном режиме) чередуясь с «пятнами» других древостоев разного возраста и несколькими, обычно небольшими, олиготрофными озерами. Населенных пунктов в этом районе мало (рис.1), высока мелкомасштабная изменчивость форм рельефа, создающая мозаику сухих и влажных лесных биотопов.
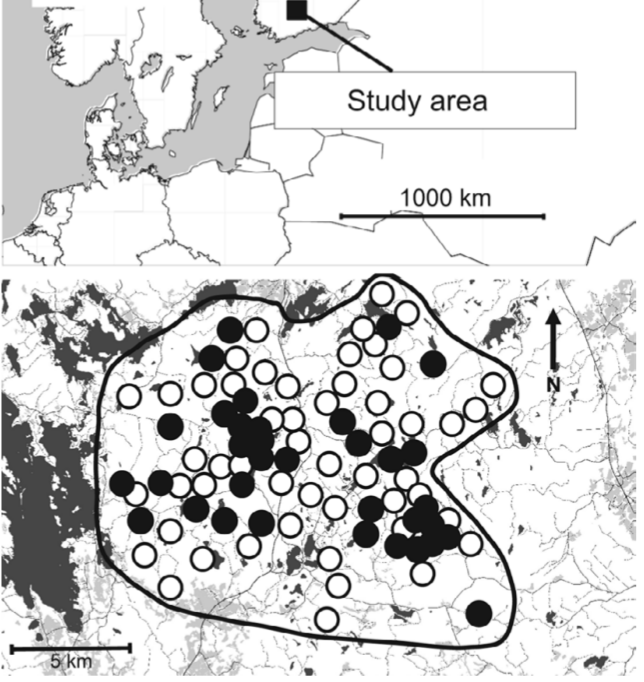
Рисунок 1. Озёра — тёмносерые, с/хугодья — светлосерые, дороги — тонкие линии, чёрные кружки — места регулярного гнездования трёхпалых дятлов (присутствует в >90% лет наблюдений, в среднем 97%, n=34), белые — нерегулярного (<60%, в среднем 33%, n=52)
В годы исследований в этой местности проводились небольшие предписанные выжигания (считается противопожарной мерой, на деле крайне опасна для тех самых массивов, которые вроде бы хотят сохранить, а не только сжигаемых). В её центре находится несколько ООПТ (conservation area) со старовозрастными лесами. В их буферных зонах в последних 20 лет проведена экореставрация — комплекс мероприятий по восстановлению эндогенной мозаичности влажных еловых лесов, развивающихся в спонтанном режиме, в том числе увеличению количества мертвой древесины. Здесь уже >50 лет живут интродуцированные канадские бобры Castor canadensis, чья деятельность ведёт к затоплению низменных форм рельефа и увеличивает обилие мертвых и гниющих деревьев [что создаёт местообитания для трёхпалого дятла как внутри ареала, так и вовне]. Леса в центральной части территории принадлежат государству (1/3 всей площади), почему экоопасная экспансия коммерческих лесов в последние 10 лет там была отчасти подавлена: скажем, поваленные деревья оставались в лесу, и проводились названные работы по экореставрации (Nordström 2015). С другой стороны, здесь — как и по всей Европе в связи с подъёмом т. н. биоэкономики — участились сплошные вырубки зрелых, ранее относительных непрерывных лесных массивов. В последние 15 лет подобные рубки были довольно интенсивны на частных землях вокруг государственного лесного участка.
Результаты
Частота повторного использования дупел и обилие многодупельных деревьев
В 179 обследования найдено 520 гнездовых деревьев, 507 были собственными трёхпалого дятла, 11 — старыми гнездовыми деревьями большого пёстрого, на 4 вместо дупла использовалась дуплянка. На 2 деревьях трёхпалые гнездились в старых дуплах большого пёстрого, но там же были и дупла трёхпалого, почему их отнесли к «настоящим» гнездовым деревьям трёхпалого.
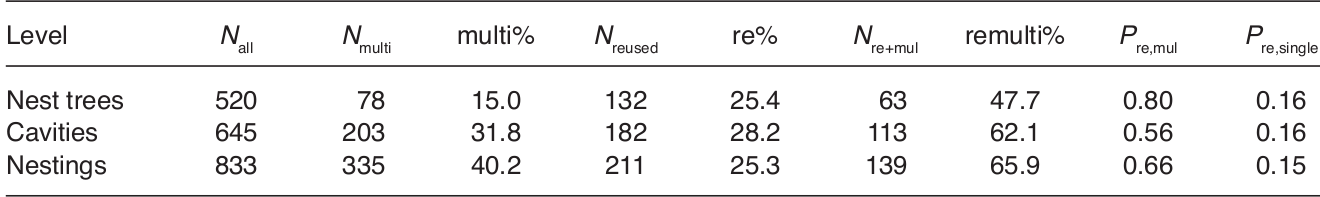
Таблица 1. Встречаемость многодупельных деревьев и повторного гнездования в прослеженной популяции трёхпалого дятла. Строки сверху вниз: гнездовые деревья, активные дупла, случаи гнездования, столбцы слева направо: всего первых, вторых и третьих, в том числе число многодупельных, оно же в %, общее число повторно используемых, оно же в %, общее число многодупельных стволов, используемых повторно, вероятность повторного использования многодупельного дерева, то же для обычного. Разница двух последних столбцов высокозначима: z = 12,2, p < 0,001 при анализе на уровне отдельных гнездовых деревьев, z = 10.5, p < 0.001 на уровне отдельных дупел, z = 8.80, p < 0.001 на уровне отдельных случаев гнездования.
Общее количество действующих дупел и гнёзд на одном гнездовом дереве составило 1-8 и 1-13 соответственно (рис. 2). За годы наблюдений отмечено 78 многодупельных деревьев (15,0% или 15,4 %, если включать только “настоящие” гнездовые деревья). Однако в них найдено 208 действующих гнёзд, там вылетело 335 птенцов, что составляет почти треть (31,8%) всех гнёзд и 40,2% всех птенцов соответственно. Повторно использовались дупла в первую очередь в год, следующей за их изготовлением, потом — сильно реже (году; 112 случаев, что 53,0 % от всех повторных использований). 10 раз дупла использовались и на третий год, но не в следующие (только 4 случая). Отмечено повторное гнездование через 19 лет после первого случая, в этом гнезде было до 4-х птенцов (рис. 2).
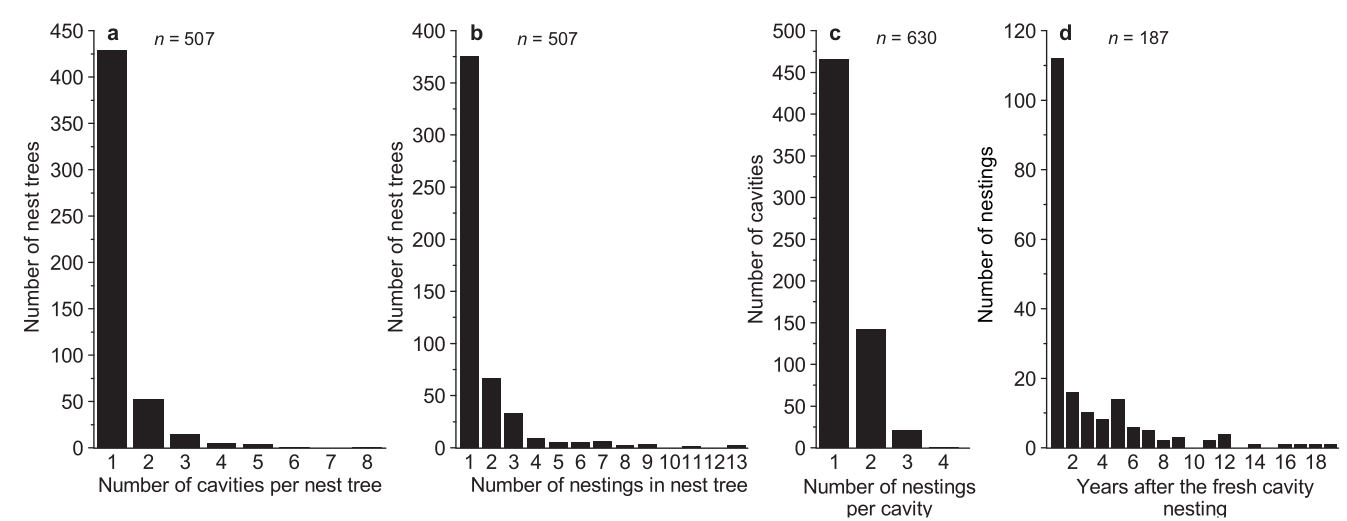
Рисунок 2. Структура повторного использования гнездовых деревьев для долбления дупел (а) и дупел для размножения (b) трехпалым дятлом, а также продуктивность размножения вообще (с) и продуктивность повторного использования дупел (d). Обозначения осей диаграмм: (а) ось Х — число дупел на гнездовом дереве, ось У — общее число деревьев с таким числом дупел; (b) — число гнездований на данном дереве, число деревьев с таким числом гнездований, (с) птенцы в гнездовых деревьях, (в) гнездования в полостях и (г) гнездования в полостях во времени после свежей полости.
Роль многодупельных деревьев в повторном использовании дупел
Кроме того, что они дают непропоционально большой % мест для размещения гнёзд и % произведённых слётков, они дали 47,7% дупел, используемых повторно. 62,1% последних находились в стволах многодупельных деревьев, и 62,9 % всех повторных гнездований в старых дуплах были именно там. гнезд были в повторно используемых полостях. Вероятность повторного использования дупел в стволах, где их несколько, значительно выше, чем для стволов лишь с одним дуплом (табл.1).

Даты гнездования при повторном использовании дупел и в многодупельных деревьях.
Данные теста Краскелл-Уоллеса показывают, что 17 лет наблюдений из 20 (1996-2015 гг.) птицы, гнездящиеся повторно и/или в многодупельных деревьях, начали размножение раньше. Однако использование поправки Бонферрони для множественных сравнений показывает, что только 11 из погодовых различий в сроках остались значимыми.
Сравнение сроков гнездования в свежеизготовленных дуплах и повторно используемых (или в них же с повторно используемыми не первый год как) показывает, что в 8 из 20 лет, где возможны такие сравнения, второе начинается значимо раньше (тест Данна: p <0,05). Однако использование поправки Бонферрони на множественные сравнения делает эти различия незначимыми (р <0,0025). Размножение в дуплах, используемых повторно после более чем 2-х лет перерыва в гнездовании дятлов, напротив, запаздывали сравнительно со свежими дуплами (тест Данна: Р < 0,05, 8 случаев; Р < 0.0025, 5 случаев). Повторное использование дупла на следующий год после его изготовления или повторного использования старого дупла также вело к более позднему гнездованию (тест Данна: Р < 0,05, 12 случаев; Р < 0.0025, 4 случая) (табл. 2).
Гнездование в многодупельных деревьях, напротив, начиналось раньше, чем в стволах с одним дуплом, в каждый из 20 лет исследования (U-тест Манна-Уитни значим в 12 (р <0,05), с поправкой Бонфероони — 5 годах (р <0,0025) наблюдений, таблица 2. Следовательно, тенденция к более раннему размножению при гнездовании в старых дуплах определяется чёткой связью последнего с многодупельными деревьями на участках.
Успех размножения при использовании старых дупел и в многодупельных деревьях
Вообще репродуктивный успех был высоким: успешно закончились 90,9 % всех случаев размножения (n = 833) и 90,0 % случаев размножения в дуплах, выдолбленных данным видом (n = 817). При повторном использовании дупел он был выше, чем при выдалбливании свежих: 93,8 % (n = 211) (все случаи повторного использования дупел) и 94,4% (n = 195) (повторно используются дупла именно трёхпалого дятла) vs 89,9 % (n = 622). Различия на грани статистической значимости (тест двух пропорций: все свежие дупла vs все повторно используемые: z = 1,73, p = 0,08; все свежие vs все повторно используемые дупла трёхпалых: z = 1,91, p = 0,06), что подтверждает авторскую гипотезу.
Успешней всего размножение при повторном использовании дупла в год, следующий за выдалбливанием — 97,3 % (n = 112). Это значительно выше, чем в свежих дуплах (z = 2,54, p = 0,01) или всех прочих повторно используемых (90,4 %, n = 83, z = 2,08, p = 0,04). Видно, что успех размножения в данном году стимулирует птиц использовать то же дупло в следующем: каждому из 126 случаях гнездования предшествовало успешное размножение в предыдущем году. И в прочих 69 случаях повторного использования дупел было лишь 2, когда птицы гнездились там, где предшествующее размножение было неудачным. Это показывает связь прошлой «истории использования» каждого дупла и/или размножений в той части участка, где оно выстроено, и вероятности их занятия и в следующем году.
Гнездование трехпалых дятлов в многодупельных деревьях было успешно в 94,3% случаев (n = 335), но в однодупельных (n = 498) лишь в 88,6 % случаев; разница статистически значима (z = 2,84, p = 0,005). Успех размножения в свежих дуплах в первом случае также значимо выше, чем во втором (n = 196, 93,9 % vs n = 426, 88,0 %; z = 2,25, p= 0,025;), при повторном использовании дупел разница в успехе размножения не значима (n = 139, 95,0 % vs n = 72, 91,7 %: z = 0,94, p= 0,35).
Таким образом, лучший общий успех гнездования в деревьях с несколькими полостями обусловлен как лучшим успехом гнездования в новых полостях, так и гораздо большей частотой повторных гнездований в полостях с высоким успехом гнездования по сравнению с деревьями с одной полостью (табл.1). и подтверждает нашу гипотезу. На основных территориях 29 % всех активных гнезд были в повторно используемых полостях и почти половина из них (48 %) в деревьях с несколькими полостями, но на других территориях соответствующие доли составляли всего 8 % и 9 % соответственно.
Следовательно, больший успех размножения дятлов в многодупельных деревьях связан как с большим успехом гнездования там в свежих дуплах, так и повышенной частотой повторного использования этих — лучших в смысле успеха размножения — дупел на данных деревьях по сравнению с однодупельными [т. е. при долговременном обитании на определённой территории дятлы определяют «лучшие» деревья в плане возможностей выкармливания выводка, его сохранения от разных опасностей и в последовательные годы гнездятся именно в них, почему они и оказываются многодупельными. Однодупельными остаются деревья, удобные для строительства гнезда по механическим качествам древесины и пр., но не очень — для собственно размножения]. Действительно, на постоянно используемых территориях 29% гнёзд были в старых дуплах, и 49% из них — в многодупельных деревьях, но на территориях, где вид гнездился не постоянно — всего 8% и 9%. Качество местообитаний на первых значимо выше, чем на вторых, однако между ними не найдено значимых различий в успехе размножения (табл.3).
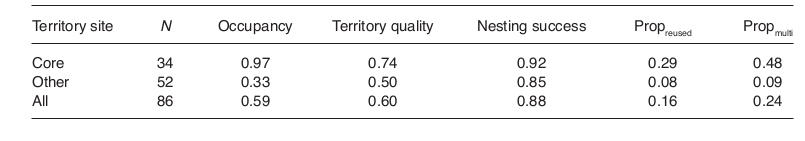
Характеристика территорий трёхпалых дятлов в местной популяции. Все участки обитания (n = 86) разделены на постоянно занятые («ядро») и занятые не каждый год. Показаны среднее качество территорий и успех размножения в обоих частях популяции, также как доля гнездований в старых дуплах («reused») и в многодупельных деревьях («multi»). Групповые средние получены усреднением показателей для отдельных территорий. Видны значимые различия в их качестве, доле использования старых дупел и многодупельных деревьев, без значимых различий в успехе размножения (Mann-Whitney U-test для качества территорий: U = 212.0, использования старых дупел: U = 168.5, многодупелных деревьев: U = 203.5, p < 0.001 для всех случаев; успех размножения: U = 861.5, p = 0.84).
Повторное использование гнёзд/гнездовых деревьев и успех гнездования на уровне территории. Обсуждение
Отмечена позитивная корреляция между постоянством обитания птиц на своих территориях за годы исследований и долей повторного использования дупел (ранговый коэффициент Спирмена, rS = 0.76, p < 0.001, n = 86), также как с долей гнездования в многодупельных деревьях (rS = 0.71, p < 0.001, n = 86). Однако не найдено зависимости между постоянством использования территории и успехом размножения (rS = 0.03, p = 0.80, n = 86). С другой стороны, фиксируется позитивная связь того же показателя с общим количеством успешных размножений в данном месте (rS = 0.93, p < 0.001, n = 86), а также между долговременностью занятия территории и её качеством (rS = 0.62, p < 0.001, n = 86), показывая большую важность территорий постоянного обитания для популяции трёхпалого дятла сравнительно с периодически заселяемыми. Повторное использование дупел на первых встречалось чаще, чем во вторых, также как и многодупельные деревья [притом что потенциально подходящих деревьев там было больше, а не меньше, чем на вторых].
Вопреки предшествующим наблюдениям, повторное использование дупел обычно для трёхпалого дятла. Полученные авторами результаты соответствуют трём объяснениям (1) оно позволяет раньше начать размножение; (2) оно делает последнее успешней, и (3) оно учащается с ростом долговременного использования территории — а может, и лучших местообитаний.
Долбить заново или использовать повторно?
Зачем дятлам повторно использовать прежние дупла вместо долбления новых. Простейшее объяснение — экономия времени и энергии, необходимых для выдалбливания, чтобы направить их на отдых или питание. Согласно Sollien et al. (1978) трёхпалому дятлу нужно 12 дней для изготовления дупла. По данным авторов, в 5 пяти случаях, когда выдалбливание прослеживали с самого начала, на это требовалось от 8 до 18 дней (медиана 11 дней; Т.Паккала неопубликован. данные).
Разные виды дятлов отличаются друг от друга по выбору гнездовых деревьев, и определяющих это условий а, значит, и по необходимости долбить дупло каждый год заново (Short 1979, Winkler & Christie 2002, Wiebe et al. 2007). Как известно, многие виды дятлов, в том числе и трехпалый, каждую весну долбят несколько дупел в разных частях участка (так называемые пробные или инициальные дупла): из них завершается только одно, а остальные находятся на разных стадиях изготовления (Blume 1968, Ruge 1974, Cramp 1985, Pechacek & d’Oleire-Oltmanns 2004, Pakkala et al. 2006). Старые, незавершённые дупла дятлы могут доделывать до гнездовых даже несколько лет спустя (Blume 1968, Ivanchev 1997, Rolstad et al. 2000, Matsuoka 2010). В 3-х случаях, когда необходимое на это время удалось точно оценить, у трёхпалых дятлов это заняло 5, 7 и 8 дней [т. е. значимо короче, чем долбление заново, и время сильно зависит от «степени готовности» инициального дупла].
Трёхпалые дятлы также начинают долбить сразу несколько дупел, зачастую — на одном и том же дереве (Schifferli & Ziegeler 1956, Blume 1968, Ruge 1974, Pakkala et al. 2006, Matsuoka 2010). Видимо, таким образом они проверяют пригодность дерева, особенно твёрдость древесины [в начале выдалбливания, и пригодность устанавливающегося с ходом весны микроклимата — в середине или конце процесса] (Matsuoka 2008, 2010, Lorentz et al. 2015). В исследовании авторов инициальные дупла обычны на многодупельных деревьях и часто используются как «заготовки» новых дупел. Использование незаконченных и инициальных дупел сэкономит время на долбление, а также время и энергию поиска подходящих мест для гнезда. Поскольку на гнездостроение уходит много сил, использование старых дупел позволяет извлечь выгоду из более раннего гнездования или нахождения в лучшем состоянии [на момент яйцекладки и насиживания], что увеличит успех размножения таких птиц.
Зачем дятлы тратят так много сил на выдалбливание дупел каждый год заново? Самый очевидный ответ — качество старых дупел падает со временем (Wiebe 2001, Edworthy & Martin 2014). Срок жизни гнездовых деревьев также ограничен: дупла в живых деревьях остаются пригодными много дольше, чем в сухих или усыхающих стволах (Wesołowski 2011, Blanc & Martin 2012, Edworthy et al. 2012). Если подходящие старые дупла наличествуют [не удалены вместе с сухими деревьями, не испорчены прежними квартирантами и птица знает, где они размещаются, в силу долговременного обитания на участке], выбор, использовать ли их или долбить новые определяется соотношением платы и выигрыша от того или иного решения [и запасом времени на строительство, которого у данной птицы может не быть, если она запоздала с образованием пары]. Результат виден на популяционном уровне в виде % повторного использования дупел у разных дятлов.
Распространение повторного использования дупел/гнездовых деревьев у трёхпалых дятлов и их родственников
Отмеченная частота повторного использования дупел (25,3%) и гнездовых деревьев (15%) исключительно высока, с учётом того, что в прошлые годы такое рассматривалось как чисто случайное (e.g. Svärdson & Durango 1950, Haftorn 1971, Winkler & Christie 2002). Разрозненные наблюдения его гнездования в дуплянках (Hortling 1929, von Haartman et al. 1963–1972, Ryssy 2004, Bolund 2010) показывают, что у вида есть потенциал гнездования в старых дуплах. Однако Wiebe et al. (2006) вообще не выявил этого у американской формы P.t.dorsalis. В южной Финляндии, в районах, прилегающих к месту нашего исследования, повторное использование гнездовых дупел трёхпалым дятлом отмечено с частотой 5-10% (H. Lokki личн.сообщ., T. Nurmi личн.сообщ., P. Pouttu личн.сообщ.).
Североамериканский P.arcticus повторно использует дупла также существенно реже, чем обнаружено авторами (2.8%–7.1%; Dixon & Saab 2000, Saab et al. 2004, Wiebe et al. 2006). Из североевропейских дятлов повторное использование дупел отмечено у желны Dryocopus martius и больших пёстрых дятлов (Dementiev & Gladkov 1966, Glutz & Bauer 1980, Cramp 1985). Доля повторного использования дупел у этих видов сильно варьирует: для высоких широт, т. е. короткого по сравнению с основным ареалом годового цикла, это 26% у желны в южной Швеции и 30% в центральной Норвегии (n = 264; Nilsson et al. 1991 и n =106; Rolstad et al. 2000), у большого пёстрого дятла это 25.8% в Рязанской обл. (n = 198; Ivanchev 1997), 21.4% в южной Англии (n = 135; Smith 1997).
[Отечественные данные этому автору — в отличие от поляков — почти неизвестны, хотя вышла сводка «Птицы России», где в разделе про дятлов они все суммированы. В плотных поселениях большие пёстрые дятлы обычно новых дупел не строят. Из 47 гнёзд в Гессене 33 были устроены в прошлогодних дуплах (лишь несколько расширенных), 10 — в доработанных инициальных дуплах и 4 выстроены заново (Blume, 1961, 1968). В Московской обл. из 24 прослеженных гнездовых пар в 1985-1989 гг. 6 долбили новое дупло, 11 расширяли инициальные дупла и 7 устроили гнёзда в прошлогодних (данные В.С.Фридмана). В Ленинградской обл. 10,8% дупел используются повторно (Сироткин, 1978), по другим данным лишь 2% (Мальчевский, Пукинский, 1983). В Окском зап. в старых дуплах ежегодно гнездились 19,2-33,3%, в среднем 25,8% пар (Иванчев, 1997). Прим.публикатора]
Большой массив данных по желне (n = 300; P. Pouttu pers. Comm.) из южной Финляндии показывает, что он чаще гнездится повторно (около 60%), чем долбит новые дупла. [возможно, рост тщательности и долговременности наблюдений за территориальными парами, увеличение выборок дупел с установленной судьбой повысят % повторного использования и для других видов.] Ещё меньше имеется исследований использования многодупельных деревьев нашими северными видами дятлов. Из хорошо изученных видов многодупельные деревья бывают у Colaptes auratus (e.g. Lawrence 1967, Moore 1995, Fisher & Wiebe 2005), большого пёстрого дятла (там их доля среди всех гнездовых деревьев колеблется между 8%–49%; Hågvar et al. 1990, Hansson 1992, Glue & Boswell 1994, Stenberg 1996, Mazgajski 1998) и чёрный дятел (25%–47%; Hågvar et al. 1990, Rolstadt et al. 2000, Kosiński et al. 2011).
В центральной Европе трёхпалые дятлы не используют гнездовые деревья повторно, или делают это случайно (Pechacek 2001, Pechacek & d’Oleire-Oltmanns 2004, P. Pechacek pers. comm., K. Ruge pers. comm.) также как в восточной Финляндии (Fayt pers. comm., J. Lakka & T. Nurmi pers. comm.). Близкий % повторных использований гнездовых деревьев — 15-20% — отмечен в южной Финляндии, рядом с местом исследования (J. Ahola & M. Nääppä pers. comm., P. Pouttu pers. comm.), и Stenberg (1996) отметил 20% для западной Норвегии. Общая картина повторного использования неясна из-за редкости популяционных исследований трёхпалого дятла, но данных достаточно, чтобы утверждать его зависимость от структуры и динамики лесной среды [скорее долговременной устойчивости использования территорий, которое не простое отображение качества местообитаний из-за контагиозного распределения многолетних территорий птиц, почему часть вполне подходящих биотопов используется не каждый год или пустует].
Повторное использование дупел и гнездовые деревья трёхпалых дятлов
Более раннее гнездование рассматривается как фактор увеличения репродуктивного успеха (больше выращивается слётков) у широкого круга групп птиц (Perrins 1970, Nilsson & Smith 1988), этот паттерн наблюдали также и у дятлов (e.g. Hogstad & Stenberg 1997, Wiktander et al. 2001, Fayt 2003, Mazgajski 2002, Pechacek 2006, Smith 2006). Wiebe et al. (2007) показал, что в 6 из 8 популяционных исследований дятлов кладка при повторном использовании дупел начиналась раньше, но только двое различий были статистически значимы, из-за небольших выборок. Повторное использование также может быть «быстрым приспособлением» к меняющимся условиям гнездования. Большое исследование золотых дятлов 1998-2006 гг. (всего проанализировано 1006 гнездований с 63,1% в старых дуплах, Wiebe et al. 2006, 2007) не было различий дат первой кладки в свежеизготовленных и старых дуплах, но большая часть вторых кладок пришлась на последние.
В данной работе повторное использование дупел распадалось на два класса:
(1) гнездование там, же где птицы гнездились в прошлом году (это может быть новое дупло или также повторное),
(2) гнездование после перерыва в 2 года и более.
Гнездования (1) начинались раньше (чем в свежих дуплах) и были весьма успешны, (2) — наоборот, позже, успех размножения был сравним с таковым в свежих дуплах. Они связаны с двумя разными ситуациями: в (1) дупло используется повторно после успешного гнездования прошлого года, (2) наоборот, есть проблемы, из-за которых начало гнездования запаздывает [скажем, птицы долго не могут образовать пару, обычно во фрагментированных местообитаниях], или первое размножение было сорвано, птица гнездится повторно. На решение, долбить новое дупло, или гнездиться в старом [если оно есть на участке, для чего птица должна здесь обитать достаточно долговременно, достаточно изучить имеющийся «фонд» дупел], также как на дату начала гнездования, влияют одновременно несколько факторов.
Отсюда разницу успешности гнездования в свежих и старых дуплах следует сравнивать надлежащим образом, учитывая неоднородность второй выборки. Для (2) ожидается позднее гнездование с успешностью ниже среднего, но именно выбор старого дупла экономит трехпалому дятлу время [иначе затрачиваемое на поиск подходящего ствола и выдабливание]. Большинство дупел использовалось повторно после успешного гнездования в первый год; при неудаче размножения в свежем дупле, оно не использовалось на следующий год и очень редко занималось позже.
Наблюдения за птицами, меченными цветными кольцами, показывают, что одни и те же особи используют одно и то же дупло в последовательные годы (тогда как другие — каждый раз долбят новое), тем самым демонстрируя память об истории использования разных дупел на участке и её влияние на последующий выбор птицы. Этот паттерн совместим с результатами исследований других дуплогнёздников [в том числе отличие от дятлов, вторичных], например, гоголя Bucephala clangula (Dow & Fredga 1983), мохноногого сыча Aegolius funereus (Sonerud 1985), чёрного дятла (Nilsson et al. 1991), мухоловки-пеструшки Ficedula hypoleuca (von Haartman 1949), и большой синицы Parus major (Harvey et al. 1979).
Как ранее обсуждалось, изготовленное дупло продолжает гнить [выдалбливание и насиживание этот процесс стимулируют], почему его качество падает со временем. Хотя у нас нет данных о качестве дупел трёхпалого дятла, можно заключить, что на второй год оно скорее близко к таковому свежеизготовленного. Паттерн повторного использования дупел, найденный авторами, и сходные паттерны у других видов дятлов показывают, что повторное использование отражает общий паттерн условий, определяющий качества дупел после выдалбливания [т. е. если птице доступны старые дупла, не сильно потерявшие в годности, с благоприятной историей прошлого размножения, она выберет их вместо долбления заново].
Ivanchev (1997) у большого пёстрого дятла отметил 80.0% (24 of 30) гнездований в старых дуплах 2хлетнего «возраста», Smith (1997) у него же — 41.4% (12 of 29) на второй и 34.5% (10 of 29) на третий год после изготовления дупла. Однако оба не сравнивали даты и успех гнездования в разных типах повторно используемых дупел. Pekka Pouttu (личн.сообщ.) установил, что по меньшей мере 50% повторно используемых гнёзд желны — это гнёзда 2го года жизни. Тем не менее, выгнивание дупел и гнездовых деревьев сильно варьирует во времени, и случаи гнездования через несколько лет после выдалбливания показывают, что часть дупел достаточно долго остаётся пригодной.
В этой работе не изучалось изменение свойств гнездовых деревьев [важных для выдалбливания], но несколько непрерывно используемых деревьев, и большинство многодупельных деревьев были живыми елями среднего размера, Picea abies, росшими в лощинах, занятых заросшими елью болотами (неопубл. данные авторов). Здесь гнездовые деревья стоят долго и сохраняют пригодность для долгосрочного использования.
В свежих дуплах на многодупельных деревьях птицы гнездятся раньше, чем в однодупельных. Авторы полагают, что это следствие использования уже имеющихся там незавершённых или инициальных дупел как закладок для долбления новых. Экономится время на поиск подходящих гнездовых деревьев и долбление, почему птицы здесь раньше гнездятся, чем в однодупельных. Здесь выше и успех размножения. Как и повторное использование старых дупел, повторное использование того же самого гнездового дерева (с долблением свежего дупла, делающего дерево многодупельным) происходит после успешного размножения там же в прошлом году. Это частично объяснит больший успех размножения в многодупельных деревьях, независимо от гнездования в свежем или старом дупле.
Деревья со старыми дуплами неизменно притягивают дятлов; трёхпалые во время размножения также регулярно обследуют старые гнездовые деревья на своей территории. Правда, утверждалось, что дуплогнёздники избегают гнездиться в многодупельных деревьях, мол, те воспринимаются как «низкого качества», скажем, из-за подверженности угрозе падения или повреждения (Aitken & Martin 2004). Риск хищничества (см. ниже) или конкуренция за дупла (Hågvar et al. 1990 и ниже) также могут снизить использование старых гнездовых деревьев. Однако, если они безопасны, а ствол годится для выдалбливания ещё одного дупла, последнее в долгосрочной перспективе может быть лучшим вариантом, чем повторное гнездование в старых дуплах, возможно, ставших негодными.
Для размножения в старых дуплах предполагается больший риск хищничества, чем в свежих, они могут успеть выучить «здесь есть пища» (Sonerud 1985, 1989). Так, лесная куница Martes martes чаще губит желну в повторноиспользуемых дуплах, чем в свежих (Nilsson et al. 1991). Однако обзор исследований разных видов дуплогнёздников Wiebe et al. (2007) не обнаружил различий успеха размножения в свежих и повторно используемых дуплах. У золотого дятла он несколько выше в первых (Wiebe et al. 2007), у больших пёстрых дятлов не отличался (Ivanchev 1997, Mazgajski 2002) или слегка повышается во вторых (Smith 1997).
Rolstad et al. (2000) не выявил разницы в частоте хищничества между свежими и прошлогодними дуплами желны, он также не коррелировал с числом старых дупел, прилегающих к данному. Сам процесс выдалбливания дупла может быть сигналом для возможных хищников, но это совсем не изучено (e.g. Wiebe et al. 2007).
Эктопаразитизм рассматривается как фактор, отталкивающий от повторного использования гнёзд (e.g. Loye & Zuk 1991, Møller 1997), в том числе и у дуплогнёздников (Mazgajski 2007a, Wiebe 2009). Смена места для гнезда его сокращает (Collias & Collias 1984, Hansell 2000), и дятлы могут избегать паразитов, долбя гнездовые дупла каждый год заново (Short 1979).
Однако значение эктопаразитизма для размножения в естественных дуплах почти не исследовано (см. Wesołowski & Stańska 2001, TomȧS et.al. 2007); несколько имеющихся работ не выявили четких различий в этой угрозе между свежеизготовленными и старыми дуплами (Wiebe 2009). Поскольку дятлы не приносят в дупло материал для гнезда (то, что можно так называть — это щепочки, счищаемые со стенок изготавливаемой полости) (например, Short 1982, Winkler & Christie 2002), риск эктопаразитизма для птенцов здесь не должен различаться между старыми и свежими дуплами.
У авторов не было данных о причинах гибели гнезд или срывов размножения. Исходя из полевых данных, наиболее очевидны дождливая и холодная погода, [снижающая успешность кормодобывания], и хищничество большого пёстрого дятла (Pakkala et al., 2006, см. Ниже). Иногда гнездование прерывалось падением гнездового дерева от сильного ветра или гибелью одного из родителей. Потенциально возможные нападения на гнёзда таких хищников, как мелкие куньи, белки Sciurus vulgaris, воробьиный сыч Glaucidium passerinum или большой пёстрый дятел (Glutz & Bauer 1980, Pakkala et al., 2006), были редки; в районе исследований хищничество — несущественный фактор снижения репродуктивного успеха. Он в целом очень высок, и риск хищничества скорее всего незначим при выборе «повторно использоваться дупла vs долбить заново»? Однако он может географически варьировать и быть более значим для населения вида в Центральной Европе, где повторное использование дупел и гнездовых деревьев крайне редко (Pechacek 2001, 2006, Ruge 1974, P. Pechacek личн.сообщ., K. Ruge личн.сообщ. T. Wesołowski личн.сообщ.).
Теоретически на обсуждаемый выбор дупла в следующих гнездованиях может влиять конкуренция с прочими дуплогнёздниками (а также на срок и успех размножения, Pakkala et al. 2006). Ряд исследований показал, что скворцы Sturnus vulgaris агрессивно захватывают дятловые дупла [заставляя долбить новые и/или гнездиться позже] и, вообще, вторичные дуплогнёздники агрессивно конкурируют за имеющиеся дупла [а точнее — за меньшинство «лучших» из них. В достаточно старых лесах «предложение» дупел всегда избыточно, но в ней есть меньшинство «наиболее популярных» у потребителей. Их неизменно недостаёт, конкуренция за подобные дупла может быть весьма интенсивна. Другое дело, что она по преимуществу не снижает репродуктивный успех даже у проигравших — в конце концов те удовлетворяются «худшими» дуплами.]. И если скворцы в районе исследований — редкость, занятия ими дупел трёхпалого не отмечено, то воробьиный сыч и большой дятел постоянно присутствуют в местах гнездования трёхпалых(Pakkala et al. 2003, 2006). Оба они в целом гнездятся раньше, почему редко захватывают свежие дупла трёхпалого, но часто занимают использованные. Они также предпочитают для этого многодупельные деревья, один сыч занимал там дупло несколько лет (T. Pakkala неопубл.данные), что делает их негодными для трёхпалого. Конкуренция с этими видами может уменьшить репродуктивный успех трёхпалых дятлов, заставив гнездиться позже, подтолкнуть их к использованию старых дупел, но со снижением репродуктивного успеха.
Повторное использование дупел на уровне территории и популяции вида: адаптация к существованию в динамичном лесном ландшафте.
Авторы обнаружили резкую разницу частот повторного использования дупел между районами регулярного и неежегодного гнездования трёхпалых дятлов («ядро» и периферия изученной территории). В первых она обычна, во второй — редкость: раньше такие мелкомасштабные различия не изучались. Первое интересно, так как трёхпалый дятел считается видом, адаптированным к типической динамике нарушений бореальных лесов (e.g. Winkler & Christie 2002, Fayt et al. 2005). [Обычные там] ветровалы, лесные пожары и затопления леса, создают территории с обилием мёртвых деревьев, гниение древесины которых привлекает сапроксильных жуков, что исключительно привлекательно для трехпалого дятла. Эти последствия возмущений часто бывают недолгими, и дятлы переселяются в новые районы. Такого рода пространственные изменения могут способствовать выдалбливанию дупел каждый раз заново у вновь возникших «скоплений» корма и/или подходящих микростаций [только вот заселение эти «пятен», возникающих и исчезающих каждый раз заново, осуществляют нетерриториальные особи из «подвижного резерва» популяции. Резидентные же, как показывает данное исследование, ориентированы на долговременное постоянство участков, и повторные использования гнездовых деревьев, несмотря на подобные нарушения. Если их интенсивность так высока, что не позволяет подобного, её следует счесть следствием хозяйственного использования массива, в доагрикультурных лесах такого не было].
Действительно, ранее авторы показывали позитивную корреляцию между постоянством гнездования вида и характеристиками лесного сообщества: его возрастом, структурным разнобразием древостоя, мелкомасштабной гетерогенностью, фиксируемой в разных пространственных шкалах (Pakkala et al. 2002, 2006, 2014). Увы, эти участки более привлекательны и для конкурентов: однако (Pakkala et al. 2003, 2006) предполагает, что большая структурная сложность лесного полога непропорционально увеличивает обилие дупел (и многодупельных деревьев) настолько что снимает остроту конкурентной борьбы за них]. Резко растёт и возможность использования старых дупел их «производителями», трёхпалыми дятлами (сравнительно с территориями нерегулярного гнездования). Поэтому оно может быть индикатором «лучших» местообитаний для данного вида и ключевых участков внутри популяционного ареала [вид угрожаем в Европе, где из-за экспансии коммерческих лесов почти не осталось не только малонарушенных, но и относительно нарушенных, но сохраняющих более или менее природный облик].
Для устойчивого существования и высокой плотности трёхпалого дятлов нужно, чтобы вышеописанные нарушения, обеспечивающие временную гибель деревьев, и благоприятные микростации для вида [вроде описанных в истории использования местных лесов], происходили на территориях, много меньших гнездовой территории пары, были точечными, случайно рассредоточенными по массиву, а не крупными, сконцентрированными в одном месте. Радиуопрослеживание птиц из территориальных пар трёхпалых дятлов также показывает, что обычно используемые места кормления могут быть далеко от мест гнездования (Pechacek 2004, Pakkala et al. 2005). Поэтому в среде обитания, соответствующей «ядру» популяции, необязательно менять места гнездования из года в год, что стимулирует привязанность к дуплам и многодупельным деревьям, «известным» успешным размножением в прошлом. И наоборот, при значительных масштабах нарушений (в десятки и сотни раз больше описанных в данной работе) фиксируется быстрое увеличение (а потом спад) численности трёхпалых дятлов, в котором не могут не участвовать «подвижные» особи со стороны (см. Fayt et al. 2005). Здесь стоит ожидать доминирования гнездования в свежих дуплах, в отличие от ситуации долговременного обитания, когда птицам известны все изменения растительности на «своей» территории, как и история существующих дупел. Долговременные нарушения, скорее всего, и здесь подтолкнут птиц к повторному использованию дупел.
Рекомендуем прочесть
 О гнездовых дуплах и эволюции гнездостроения у дятлов Picidae:
О гнездовых дуплах и эволюции гнездостроения у дятлов Picidae: Как в блокаду спасли Ленинградский зоопарк
Как в блокаду спасли Ленинградский зоопарк Болит ли голова у дятла?
Болит ли голова у дятла? Так кто всё-таки сэкономил на мозгах?
Так кто всё-таки сэкономил на мозгах? Зачем большим пёстрым дятлам всю зиму охранять территории?
Зачем большим пёстрым дятлам всю зиму охранять территории? «Гены урбанизации» и приспособление к средовому стрессу
«Гены урбанизации» и приспособление к средовому стрессу Кто далеко летает, живёт рискованней
Кто далеко летает, живёт рискованней Пресечение урбанизации птиц как зеркало московского экоцида
Пресечение урбанизации птиц как зеркало московского экоцида
