
 Резюме. Вольфганг Шлейдт — это ученик Лоренца, решивший ту действительную проблему этологии, за которую её атаковали социобиологи в 1970-е — произвольность выделения единиц поведения — демонстраций из потока активности, когда разделение на «фигуры» и «фон» оставлялось на мнение квалифицированного наблюдателя, созданное его «накопившимся впечатлением» от просмотров разных взаимодействий. Подробней об этой «атаке», вызвавшей концептуальный кризис сравнительной этологии, потом и прекращение исследований в рамках её программы, я писал ранее и рассказывал на «22 веке» («Взлёт и падение сравнительной этологии»).
Резюме. Вольфганг Шлейдт — это ученик Лоренца, решивший ту действительную проблему этологии, за которую её атаковали социобиологи в 1970-е — произвольность выделения единиц поведения — демонстраций из потока активности, когда разделение на «фигуры» и «фон» оставлялось на мнение квалифицированного наблюдателя, созданное его «накопившимся впечатлением» от просмотров разных взаимодействий. Подробней об этой «атаке», вызвавшей концептуальный кризис сравнительной этологии, потом и прекращение исследований в рамках её программы, я писал ранее и рассказывал на «22 веке» («Взлёт и падение сравнительной этологии»).
Другое решение независимо от него предложили в Израиле Ноа Эшкол и Абрахам Ваксман, создавшие систему формализованной записи движений танца (Eshkol-Wachmann Movement Notation). Я рассказываю об обоих, вместе с грустной историей, как эти решения, способные защитить этологов от социобиологической критики, которая в данном вопросе била в «больное место» им оказались совсем не нужны. Это одно из подтверждений современного «общего места» в логике и методологии науки, что научное знание в истории каждой дисциплины не только накапливается поступательно, но и забывается в силу разных, иногда вненаучных причин. Поэтому их развитие — не поступательное движение «вперёд и вверх», ложно считал К.Поппер, но местами «шаг вперёд, два шага назад».
Это хорошо показано в диссертации Ивана Кузина, посвящённой истории борьбы Гулда и Левонтина с социобиологией; так получилось, например, с симметричной версией полового отбора Дж.Хаксли, много лучше соответствующей фактам, чем господствующая сейчас асимметричная, Фишера-Бейтмана).
Я переписывался со Шлейдтом, поскольку 40 лет спустя решаю ту же проблему (несколько по другому, о чём сейчас пишу книгу). Надеюсь что за это время негодность социобиологических концепций стала ясна, плюс исследования в рамках данной парадигмы добыли массу полезного знания, которое почти всё больше подходит для развития сравнительно-этологических теорий (исследования в рамках которых не ведутся с конца 1980-х, кроме некоторых одиночек) чем теорий «родной ей дисциплины», закрывают многие концептуальные изъяны в первых (о них см. первую часть статьи «Как социобиология себя отрицает») и, напротив, высвечивают их во второй.
Проблема объективного выделения единиц видоспецифического поведения («демонстраций»)
Содержание
Вольфганг Шлейдт — это ученик Лоренца, решивший ту действительную проблему этологии, за которую её атаковали социобиологии в 1970-е — произвольность выделения единиц поведения — демонстраций из потока активности, когда разделение на «фигуры» и «фон» оставлялось на мнение квалифицированного наблюдателя, созданное его «накопившимся впечатлением» от просмотров разных взаимодействий. Подробней об этой «атаке», вызвавшей концептуальный кризис сравнительной этологии, потом и прекращение исследований в рамках её программы, я писал ранее и рассказывал на «22 веке» («Взлёт и падение сравнительной этологии»).
А уже тогда было известно, что это впечатление наблюдателя гарантированно обманывает: категориальность человеческого восприятия не обойти. Известные опыты Фрица Хайдера & Марианны Зиммель (1944) или Альберта Мишотта (1947) показывают, что все мы воспринимаем сближение и соприкосновение как взаимодействие и влияние, а непрерывный поток изменений склонны делить на устойчивые фрагменты — «слайды», чтобы их формами оперировать в следующем восприятии таких же потоков, даже когда они объективно невыделимы.

Фриц Хайдер (в центре), на Небрасском симпозиуме по психологии. 1960 г.
Квалификация тут не помогает, из-за чего разные и равно квалифицированные наблюдатели дают очень разные наборы единиц у одного и того же вида в одних и тех же типах взаимодействий (скажем, брачных или охраны территории). В результате где-то к середине 1970-х выяснилось, что этология оказалась в странном положении науки, лишённой адекватного ей метода описания реальности. Об этом много писал Е.Н.Панов, особенно в «Судьбах сравнительной этологии». Правда, его решение вместе с водой выплёскивает и ребёнка. См. анализ в «Как социобиология себя отрицает», ч.1 и 2: вместо устранения изъянов метода он пробовал «отменить» сравнительную этологию вовсе, однако попытка была с негодными средствами (как и у социобиологов, атаковавших те же самые аспекты этологических теорий, хоть и с противоположных позиций, хотя Евгений Николаевич — крупнейший критик социобиологии в нашей стране). Пока этологов было мало, сходило и более приблизительное/субъективное выделение единиц, поскольку установление действенности их как релизеров и исследование на этой основе механизмов коммуникации было важней точного выделения единиц, отделения их друг от друга и «фона» при составлении этограмм — «словарей» демонстраций и их «значений» (эффектов). Критика социобиологами этологического метода била в больное, в действительную проблему (в отличие от критики ими этологической теории, вовсе неадекватной).
Вольфганг Шлейдт нашёл решение: в первой половине 1980-х гг. появляются его работы по брачным демонстрациям селезня кряквы, Finley et al., 1983; расписного перепела Synoicus chinensis, Schleidt et al., 1984, и общетеоретические работы Schleidt, 1980, 1982 о методе, позволяющем наблюдателю в отслеживаемых потоках активности направленно «нащупывать» действительные отдельности — демонстрации (характеризующиеся инвариантом формы и дифференцированные друг от друга и от «фона») и одновременно отвергать кажущиеся, проверяя их на отдельность, инвариантность формы и т.д. важные характеристики. Устойчивое впечатление наблюдателя, даже квалифицированного, здесь обманывает в силу неустранимой склонности нашего сознания категоризировать наблюдаемое, даже если это континуум, делить его на отдельные объекты, приписывая движению и сближению объекта причинно-следственные объяснения вроде сигнала и ответа (как это показано в опытах Хайдер-Зиммель и Мишотта).

«Накопившись» за время отслеживания, оно делается ригидным, как «схватывается» гипс, после чего некритично воспринимает действительные отдельности и кажимости на равных, как реально существующие. Необходима аналитическая процедура, отделяющая те и другие среди отмеченных наблюдателем единиц. Она следует гипотетико-дедуктивному методы — выдвигает и проверяет гипотезы о реальности существования каждой из единиц, почему представляет собой второй шаг описания демонстративного поведения вида в территориальных, брачных, или иных взаимодействиях (первый — индуктивная фиксация единиц, основанная на «схватывании» их чётко очерченных и характерных образов вниманием наблюдателя по контрасту между ними и «фоном» образующим идеальный континуум, или контрасту между последовательными единицами, созданному резкой границей между ними во времени даже если составлены одними и теми же или близкими телодвижениями и действиями).
Вольфганг Шлейдт, анализируя фильмы ухаживания селезня кряквы Anas platyrhynchos, подтвердил реальность существования первых (шести) и отбросил вторые (три) из 9 демонстраций, выделенных классиком в рамках чисто индуктивной процедуры. Демонстрации здесь выделялись аналитически из потока движений, фиксированных киносъёмкой, т. е. непрерывно, без предрешения где проходит граница между формами отдельных единиц.
Выделение демонстраций «геометрическим методом» В.Шлейдта
[Шлейдт произвёл] «последовательный анализ брачных демонстраций селезня кряквы в присутствии самки (рис.1). Они были детально изучены ещё отцами-основателями этологии и переисследовались каждым следующим поколением учёных. Соответственно, сохранились кино- и фотоматериалы старых работ, на которых возможно тестирование новых методов описания демонстраций со сравнением точности и устойчивости выделения единиц в новых и традиционных методиках, см. Schleidt, Oeser, 2011.

Рис. 1А. Ухаживательные демонстрации кряквы, выделенные как «характерные кадры» процесса (см. также фильм). Это подход классической этологии (по: Lorenz, 1941, 1952), оказавшийся уязвимым для критики, ибо выделение индуктивное, по «устойчивому впечатлению» наблюдателя, а оно неизменно обманывает, требует проверки и корректировки. Как это и показывают работы Шлейдта.
Возможны три разных подхода к описанию брачных демонстраций селезня кряквы во время ухаживания за уткой на воде (рис.1А-Б). Первый применён классическими этологами, которые на основании устойчивых впечатлений квалифицированного наблюдателя выделили 10 характерных «кадров» — демонстраций (индуктивная процедура, Lorenz, 1939b).
Второй способ классификации исходит из предположения, что ухаживание самца за самкой есть непрерывный поток действий, континуум последовательных телодвижений, который развёртывается так, что как бы проецируется на экран, заданный координатами «высота глаза над водой – высота хвоста над водой», в плоскости которого находится самка (рис.1Б). Описание подчёркивает непрерывность изменения видимых очертаний тела токующего самца; градуальные изменения координат разных «опорных точек» на корпусе демонстратора вносят больший вклад в эффект сигнала, подаваемого силуэтом селезня, чем дискретное «перещёлкивание» отдельных демонстраций (Schleidt, Crowley, 1980).
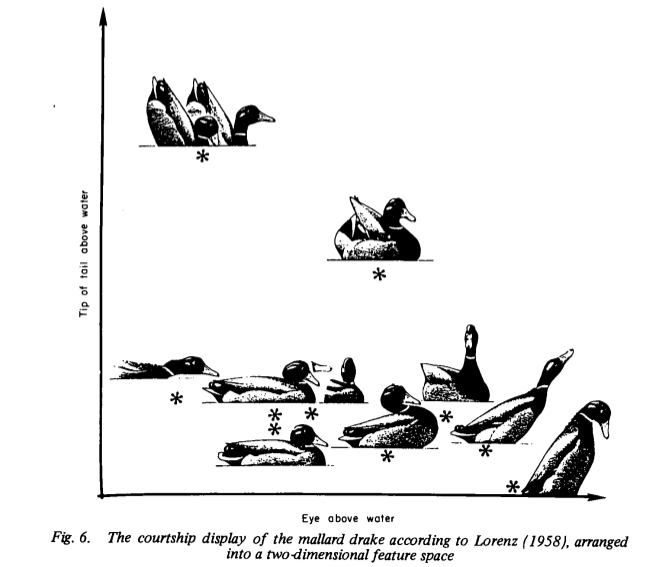
Рис.1Б. Та же последовательность телодвижений ухаживающего селезня в двухкоординатном признаковом пространстве (ось Х — высота положения глаза над водой, У — верхушки хвоста над водой). Из: Schleidt, Crowly, 1980.
Этот способ на первом шаге анализа представляет коммуникацию фильмом, где все девять «кадров» — демонстраций выделены предельно условно. Иными словами, метод не предрешает подразделения потока на единицы, как бы к этому наблюдателя не «подталкивали» его впечатления, неустранимо склонные к категориальности. На втором этим же методом в данном потоке изменений выделяются дискретные единицы — «слайды», долго удерживающие специфический инвариант формы, а затем «перещёлкивающиеся» скачком, если они в нём реально присутствуют. Если нет, рабочая гипотеза о подразделённости отслеживаемого потока активности на дискретные единицы, «схваченные» вниманием наблюдателя, опровергается на той же самой выборке данных.
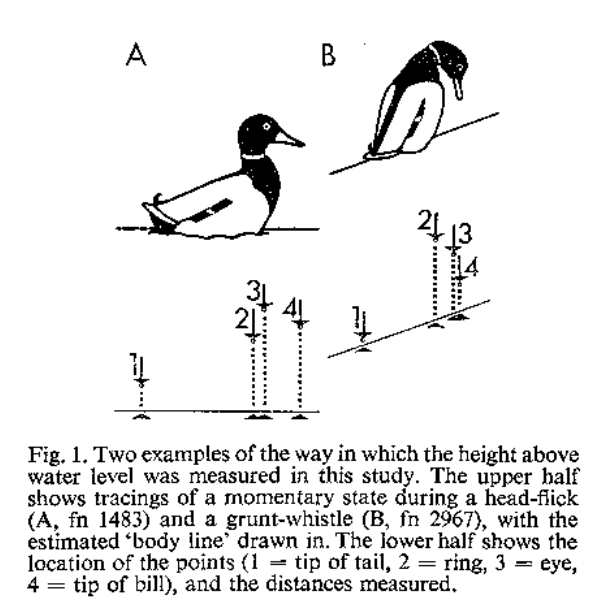
Рис.1Б. Два примера пути выделения демонстраций как «фигур» в двухкоординатном пространстве изменений визуального образа самца, репрезентируемого самке, из: Schleidt, Crowly, 1980. Обозначения. 1-4 – «опорные точки», сдвиги в положении которых описывают изменения силуэта птицы в процессе перехода между демонстрациями. 1 – высота кончика хвоста над водой, 2 – то же для шейного кольца, 3 – для глаза, 4 – высота кончика клюва над водой. Рисунки справа и слева показывают способы измерения параметров 1-4 при разном положении линии тела в демонстрациях «кивки головой» (head-flick) и «ворчливый свист» (grunt-whistle).
Сам процесс демонстрирования представляется кинолентой, откуда наблюдатель «выхватывает» самые впечатляющие «кадры» — демонстрации, исключительно чтобы маркировать ими некоторые ключевые моменты непрерывного развёртывания ухаживаний селезня, а не потому что соответствующие демонстрации объективно существуют для крякв, то есть воспринимаются как отдельные единицы, которые существенны, поскольку что-либо «значат» (стрелки на рис.1А). Собственно, смысл второго подхода в том, чтобы объективно зафиксировать плавное превращение демонстраций друг в друга в процессе ухаживания через регистрацию изменений в положении опорных точек 1-4 (рис.1Б), точно выделить те временные отрезки взаимодействия, к которым могут быть привязаны «кадры» 1-10, отделить их от времени перехода между демонстрациями.
Сравнив результат первого метода описания со вторым, видим хорошее соответствие в наборов выделенных структур – демонстраций, но первый игнорирует резкую неравномерность времени, «отводимого» на разные этапы взаимодействия. А она крайне значима для осуществления коммуникативной функции демонстраций и для решения вопроса о реальности их как дискретных единиц поведения. Ведь устойчивые и долго удерживающиеся элементы, выделенные наблюдателем, априори представляются «реальнее» кратких и мимолётных. А если демонстрации – это стимулы, то и коммуникативное воздействие вторых будет слабее.
Пример. Исследование предсовокупительного поведения обыкновенного тритона Tritirus vulgaris показало, что демонстрации, выделенные наблюдателем, вполне достаточны для объяснения связей между стимулом и реакцией на тех стадиях взаимодействия, где обмен сигналами происходит согласованно. Однако, исходя из установленных эффектов воздействия разных демонстраций, нельзя спрогнозировать момент перехода обоих участников – и демонстратора, и объекта воздействия/реципиента сигнала — к своим следующим демонстрациям (Halliday, 1975). Эта низкая прогностичность в отношении моментов смены демонстраций сохраняется на всём протяжении процесса ухаживания, даже в периоды высокой согласованности и взаимной комплементарности поведения обоих (которые чередуются с периодами несогласованной активности).
Этограмма, созданная вторым методом, только иллюстрирует ухаживание подбором ярких «кадров» — демонстраций, но отнюдь не исчерпывает его. Цепочка «кадров» лишь подчёркивает непрерывность процесса ухаживания и условность его подразделения на отдельные стадии, маркированные каждая «своей» демонстрацией. Таково представление ухаживаний селезня в традиции классических этологов — как последовательной смены фиксированных комплексов действий, каждый из которых обозначается как «демонстрация» (рис.1А, 1-10).
Иными словами, в основе второго способа описания лежит представление о потоке поведения как континууме. Несмотря на это, он подтвердил объективное существование демонстраций как дискретных структур, обладающих специфической формой, чётко дифференцированных друг от друга и от «фона» в виде несигнальной активности. Только этих «объективно существующих» демонстраций в брачном поведении кряквы выделено шесть, а не десять, как то представляется наблюдателю извне (рис.1В).
Более тщательный покадровый анализ «киноленты» ухаживания селезня показал, что процесс последовательной трансформации образа демонстратора на двумерном «экране» — не непрерывный «фильм», а скорее «слайд-шоу», составленное из дискретных кадров (Finley et al., 1983). Брачные демонстрации селезня адекватней описываются дискретной последовательностью из 6 чётко обособленных элементов (рис.1В, 1), чем моделью непрерывного континуума действий, где выделяются 9-10 демонстраций-«кадров», обычно фигурирующих в этологических описаниях (Lorenz, 1939b; Schleidt, Crowly, 1980). Иными словами, разбиение на 9 или 10 демонстраций актуально для нас, но незначимо для самих особей.

Рис.1В. Аналитическое выделение шести демонстраций как дискретных структур или дифференцированных образов (здесь и далее по: Finley et al., 2013). В.I. 1. Обозначения. Ось Х – время исполнения, ось У изменения параметров 1-4 в процессе исполнения демонстрации и перехода к следующей. положение точек 1-4. Звёздочки – высота хвоста над водой, треугольники – высота глаза над водой, кружки – высота шейного кольца и квадратики – высота кончика клюва.
Как уже было сказано, брачные демонстрации самца как бы проецируются на экран, в плоскости которого находится самка. В такой системе любая «позиция» демонстрирующего селезня описывается путём регистрации координат 4-х опорных точек на корпусе (кончик клюва, хвоста, темя и дорзальная поверхность шеи). Они и претерпевают самые резкие и порывистые изменения в ходе последовательных трансформаций силуэта самца.
Оперируя этими переменными, выделили 6 основных моторных координаций в поведении демонстрирующего селезня, показав устойчивость воспроизведения инварианта каждой демонстрации в реакциях разных особей, в разные моменты процесса, в разных контекстуальных обстоятельствах, а также дифференцированность всех шести демонстраций друг от друга (рис.1В, 2). Выделение форм демонстраций проводили дискриминацией «фигур», «очерчиваемых» в двухкоординатном пространстве силуэтом особи при последовательном исполнении телодвижений ухаживания (рис.1В, 3).
Здесь хорошо видим сопряжённые движения разных частей тела при исполнении демонстраций: этими телодвижениями «вычерчивается» инвариант формы, устойчивость и точность «черчения» (также как типичность и узнаваемость «обрисованного» ими образа) определяется стереотипностью исполнения каждой демонстрации, которая может быть объективно описана и количественно оценена в рамках подхода Шлейдта. Обратите внимание на устойчивые различия в исполнении демонстраций близкой формы (head-shake и grunt-whistle), сменяющих друг друга в процессе ухаживания, при неизменности исполнения каждой из них, инвариантность формы обоих не отрицается, а подчёркивается межиндивидальными различиями в исполнении, которые имеют место. Следовательно, здесь отдельные демонстрации – не «кадры», а «слайды» — реально существующие единицы процесса, значимые для самих особей, а не моменты, выделенные условно (Finley et al., 1983).
Анализ одних только временных характеристик телодвижений ухаживания недостаточен для разделения потока на дискретные единицы – демонстрации. Причина в исключительно высокой вариабельности времени предъявления каждой из них: средняя длительность исполнения меняется от 0,69 до 2,25 секунд с коэффициентом вариации 0,14-0,52 с. Это не позволяет выделить демонстрации напрямую, по «устойчивым образам», накапливающимся во впечатлении наблюдателя: при просмотре серии последовательных предъявлений в попытках объективной характеристики образа каждой из них в действительности он будет расплывчатым и нечётким из-за сильных вариаций длительности удержания, так что даже при высокой стереотипности предъявления не удастся их отделить друг от друга, впечатление же о чёткости разделения потока телодвижения — из ряда ошибок восприятия, описанных выше.
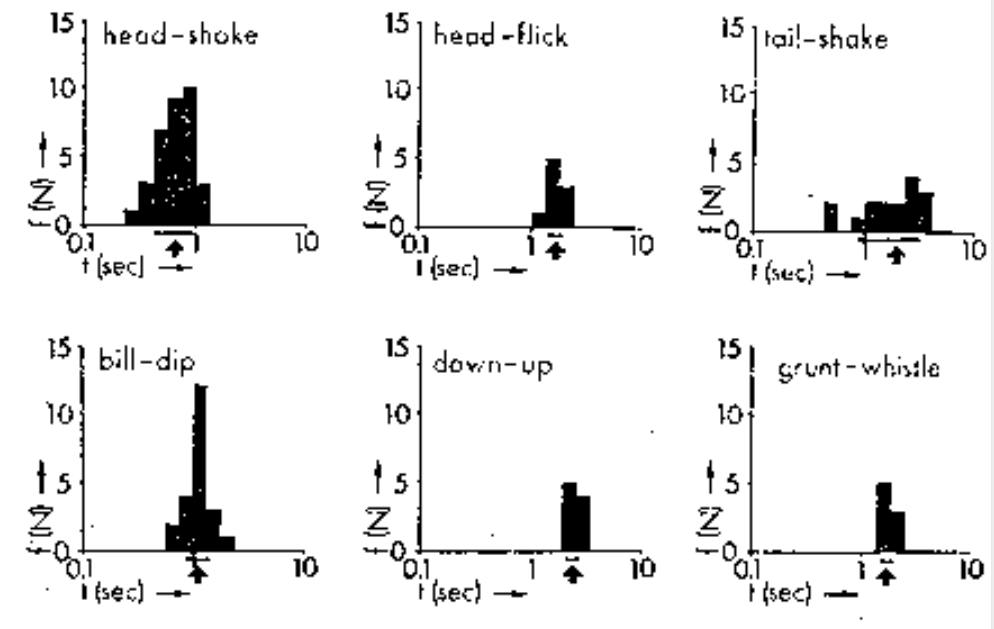
Частотное распределение продолжительности удержания 6 разных демонстраций. Стрелка под осью абсцисс — средняя, горизонтальная линия — плюс-минус стандартное отклонение. Из: Finley et al., 1983, рис.2
К тому же «устойчивость впечатления» и «чёткость» образов каждой из них обратно пропорциональны колеблемости времени его удержания в разных актах предъявления. Ведь сознание должно зафиксировать отличие ярких и экстравагантных демонстраций от несигнального поведения и (самое сложное) друг от друга, провести между ними границу во времени. Если время удержания демонстрации сильно варьирует, наблюдатель затрудняется составить о неё устойчивое впечатление, оно останется ярким, но неопределённым, и в дело включается воображение, чтобы впечатление закрепить. Это и есть источник неустранимых ошибок в любом описании ритуализованных демонстраций из состава видового репертуара.
Поэтому нужен анализ форм, «описываемых» движениями селезня, с доказательством, что они не переходят друг в друга плавно, а меняются скачком. Он был проведён В.Шлейдтом с соавторами (рис.1В) и подтвердил безусловную дискретность всех шести выделенных элементов поведения. Постулируемого континуума выразительных движений просто нет; во всяком случае, нет континуума в тех движениях, комплексы которых реализуют самоё демонстрации. С точки зрения формы все шесть демонстраций селезня — дискретные структуры, это замкнутые контуры, образованные движущейся точкой (любой из четырёх опорных). Контур каждой демонстрации начинается и заканчивается именно в том самом участке системы координат, где находятся значения, присущие обыденной (недемонстративной) позе (рис.1В).
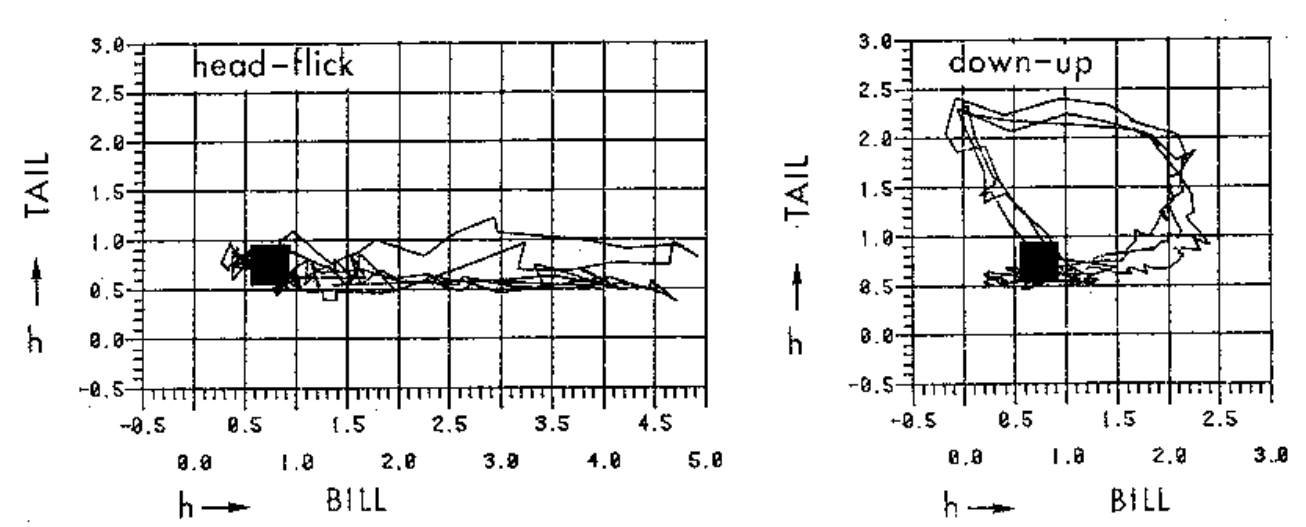
Рис.1В.I.2. Точность воспроизведения форм демонстраций в телодвижениях индивидов. Показано три независимых исполнения демонстраций «вверх-вниз» (down-up) и «кивки головой» (head-flick) разными особями в разных взаимодействиях. Обратите внимание на точность «вырисовывания» формы демонстрации во всех этих случаях. Обозначения. Ось Х – высота клюва над водой, ось У – высота хвоста над водой, чёрный квадрат — обыденная поза спокойно сидящей птицы. Отклонение силуэта птицы от обыденной позы в сторону области, очерченной замкнутыми линиями на рис.1.В.I.2 означает исполнение соответствующей демонстрации; важно, что у всех шести демонстраций эти области не пересекаются (рис.1В.II).
При смене демонстраций самца все 6 моторных координаций, последовательно реализуемых в процессе ухаживания, переходят друг в друга не непрерывно, а через «точку 0», соответствующую обыденной позе. Этим подтверждается дискретность демонстрирования: форма каждой из 6 демонстраций описывается уникальной траекторией, которая уже в силу замкнутости не предполагает плавного перехода к траектории другой демонстрации (Finley et al., 1983).
Вообще, во всех случаях, где это специально исследовали (что вовсе непросто), смена одной демонстрации на другую никогда не происходит так, как предполагал Е.Н.Панов (1978) — через постепенную замену выразительных движений первой комбинации на выразительные движения второй. В этом случае поток демонстративного поведения животного представлял бы собой идеальный континуум, что важно для теоретических построений данного автора. Однако нет – смена демонстраций скачкообразна: две последовательные демонстрации разделяются принятием на мгновение обыденной позы, но не трансформируются плавно одна в другую.
Каждая траектория очерчивает специфическую форму демонстрации, как движением огонька в темноте вычерчивают буквы или фигуры. Форма устойчиво воспроизводится в разных актах предъявления данной демонстрации разными особями, в разных обстоятельствах контекста, в ответ на разные элементы поведения партнёра и т.д.. Устойчивость воспроизведения инварианта формы сохраняется на всём соответствующем разнообразии ситуаций (=в максимально гетерогенной выборке предъявлений сигнала), несмотря на мощные факторы «помех», препятствующие сохранению инвариантности формы и дифференцированности демонстраций друг от друга. Так в шести демонстрациях селезня раз за разом воспроизводятся одни и те же инварианты формы сигналов, чётко дифференцированные друг от друга и от континуума несигнальных движений, образующих «фон».
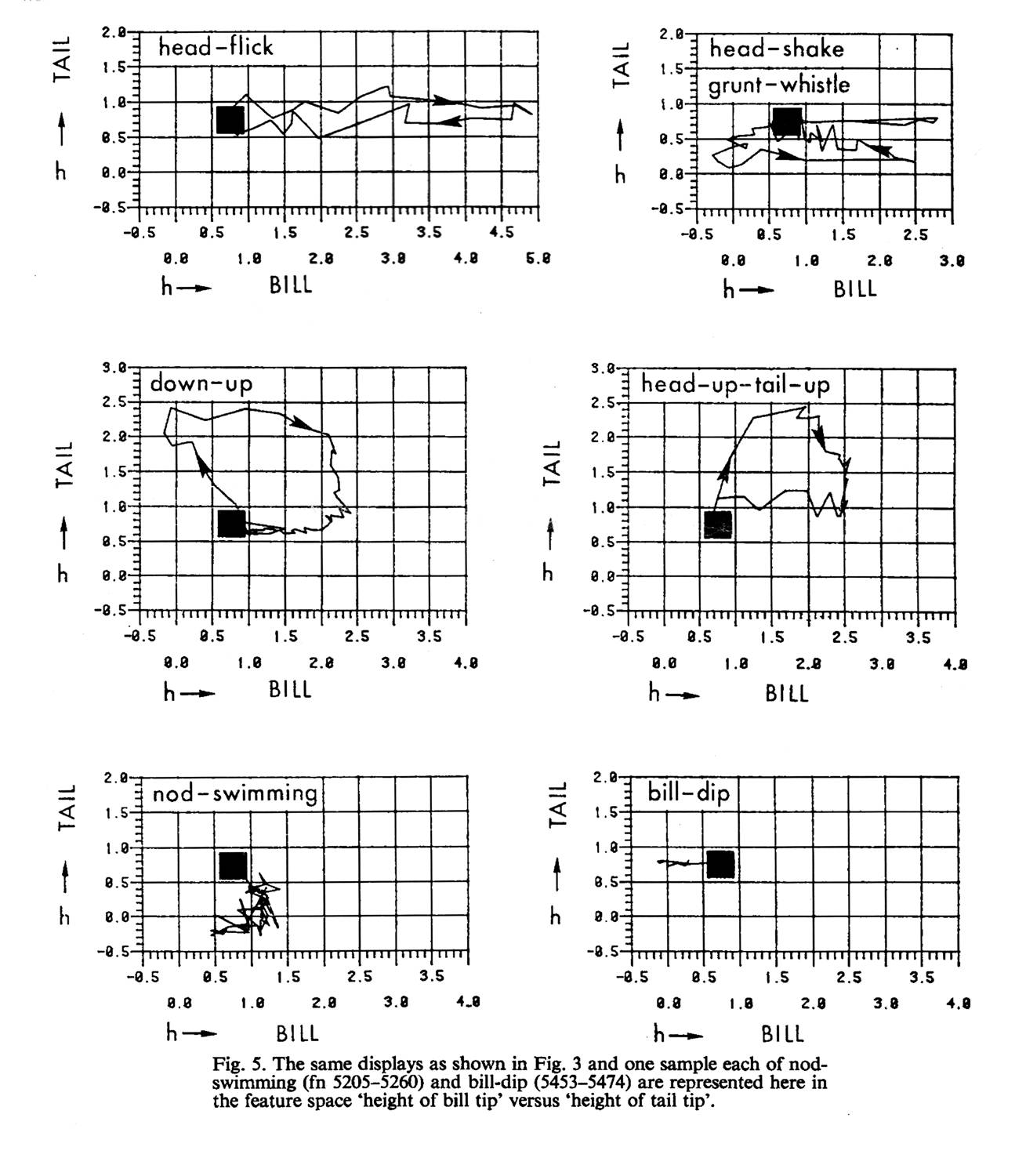
Рис.1В.II.1. Дискретность выделенных брачных демонстраций кряквы. При непосредственном следовании друг за другом в потоке поведения токующего самца переходы между этими шестью демонстрациями не плавны и постепенны, они происходят скачком через «точку 0» — обыденную позу, даже если последовательные демонстрации включают близкие телодвижения. Обозначения см.рис.1В.I.2.
Следовательно, дискретность демонстраций видового репертуара, возможность функционирования их как знаков в коммуникативном процессе определяется степенью скоррелированности исполнения разнородных движений из состава КФД, образующих демонстрации, а не стереотипностью и шаблонностью исполнения каждого элементарного действия из состава демонстрации, как считали классические этологи (Dane et al., 1959; Moynihan, 1970).
Мы видим, что демонстрации самца исполняются стереотипно именно в смысле воспроизведения раз за разом одних и тех же инвариантов формы на уровне точности, достаточном для однозначного распознавания компетентным участником коммуникации, но недостаточно шаблонно и схематично, чтобы их «зацепило» внимание внешнего наблюдателя. Вообще, интуиция исследователя сильно преувеличивает эту шаблонность и схематичность (Dane et al., 1959). Поэтому при расшифровке сигнального репертуара формы демонстраций следует выделять аналитически, дифференцируя их друг от друга и от «фона» процесса. Например, так, как предложено в работе J.Finley et al. (1983) или как сделано в наших работах (Фридман, 1992а, 1993а, 1996а, 1998, 1999, 2006, 2007). Следовательно, необходимы формальный анализ элементов поведения и формальные методы оценки дискретности разных демонстраций репертуара.
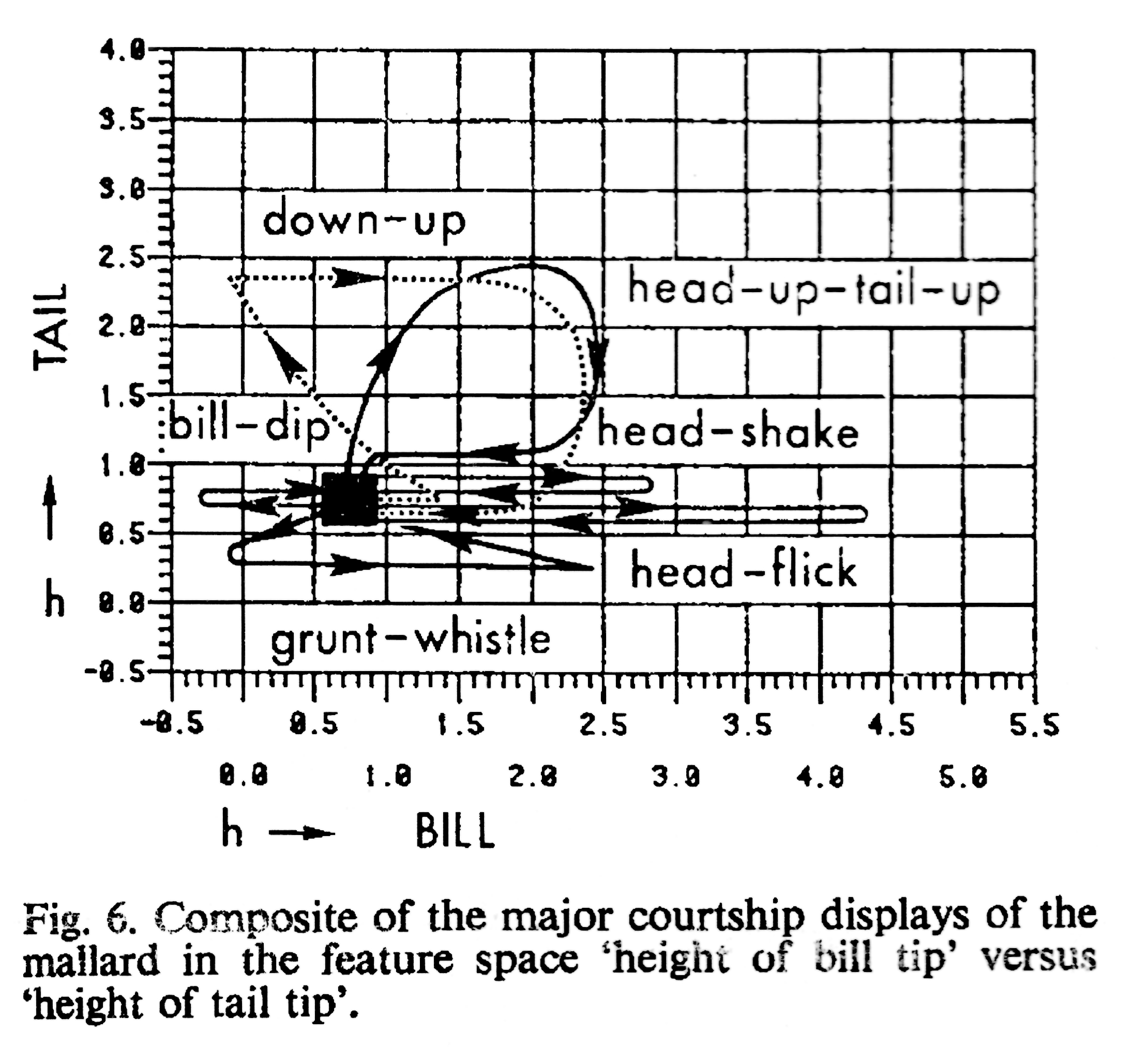
Рис.1В.II.2. В выборке брачных взаимодействий разные демонстрации занимают разные «области» в двухкоординатном пространстве, с устойчивым разграничением «областей». Обозначения см.рис.1В.I.2.
Значит, если демонстрации описывать как специфические комбинации выразительных движений, исполняемых скоррелированно в составе комбинации, то устойчивые комбинации обладают специфическим образом, присущим именно и только им как целому, и в потоке поведения образуют дискретную последовательность – слайд-шоу, а не фильм. Элементы последовательности («слайды», сменяющие друг друга в соответствие с динамикой проблемных ситуаций процесса) есть демонстрации классических этологов в точном смысле данного понятия, пусть они и выделены другим методом.
Во-первых, эти специфические элементы существуют объективно и значат для регуляции поведения особей-участников взаимодействия больше, чем они значимы как элементы классификации видового поведения внешним наблюдателем. Для верификации этого достаточно показать, что формы соответствующих демонстраций используются как идеальные «образцы», под которые «подгоняются» телодвижения индивида в конкретных актах коммуникации – что и обнаруживается в опытах, где распознаваемый образ воспроизводился искусственными нейронными сетями (Ghirlanda, Enquist, 1998; Enquist, Ghirlanda, 2005; Peters, Davis, 2006). Другие доказательства существования «образцов» структуры сигнала получены в исследованиях онтогенетеза песни птиц и роли Sollwert’ов – идеальных «образцов» видоспецифической структуры песни в становлении и стабилизации последней в развитии реальных вокализациях конкретного индивида (Slater, Catchpole, 1995). Применительно к визуальной или запаховой коммуникации в природе данный вопрос лишь начинает исследоваться (Hurd, Enquist, 2001, 2005; Peters, Evans, 2003a-b, 2007).
Во-вторых, коммуникативный эффект демонстраций достигается лишь в связи с дискретизацией их форм в потоке поведения, производимой обоими участниками процесса — демонстратором при предъявлении и реципиентом при распознавании сигнала, при том, что комплементарность поведения первого и второго усиливает дискретизацию значащих элементов на следующей стадии процесса и т.д. При такой дифференцированности «значащих элементов» лишь дискретные демонстрации существенны для партнёров как отдельные единицы поведения и единицы коммуникации.
***
И никого из этологов эти работы, спасающие дисциплину, не заинтересовали (!), в т.ч. и других учеников Лоренца, живших и работающих в данное время. Чему Шлейдт посвятил грустную короткую статью 1985 г. — «В защиту стандартной этограммы» («In Defense of Standard Ethograms«), где констатирует, что не сумел «защитить» ни её, ни в целом исследовательскую программу сравнительной этологии. В том числе в кругу «научных детей» или «внуков» Лоренца с Тинбергеном, казалось бы, максимально в этом заинтересованных, уже отдавших этологии 25-50 лет жизни и наблюдающих её кризис и пресечение в конце 1980-х)!
В защиту стандартной этограммы
Вольфганг Шлейдт
Предложенная нами стандартная этограмма [расписного перепела Coturnix cinensis, Schleidt et al., 1984] вызвала сравнительно большое число обращений за перепечаткой за короткое время. Учитывая, что использованный модельный вид, синегрудый перепел, не принадлежит к обычно используемым в поведенческих исследованиях, можно заключить, что нужда в перепечатках вызвана интересом к собственно теоретической задаче: какой дизайн стандартной этограммы или, точнее, какова организация процесса описания видоспецифических форм поведения чтобы сделать информацию о них [в отслеживаемом поведении] более доступной для извлечения, более удобной для представления и использования и одновременно способствующей извлечению релевантной информации?

Расписной перепел
Я очень признателен за полученные комментарии, что были представлены на предшествующих страницах. Однако мне не скрыть своего разочарования, с одной стороны, от отсутствия каких–либо прямых атак [на эту методику. Здесь и далее в квадратных скобках прим.публикатора] («как абсурдно…»), при отсутствии достойных внимания альтернатив («много лучше было бы…»), и, главное, от отсутствия явных подтверждение от пионеров в сравнительном исследовании поведения («мы пробовали так и так, и это работает…»). Вместо этого я получил несколько писем от старых друзей (тех самых «пионеров»), которые я воспринял как мнение: совершенно замечательная работа, но нереалистически романтичная.
«Мне кажется невозможным перекинуть мост над пропастью между прагматикой и идеалом» (P.G.Baerends, личн.сообщ.).
Многие из обычных непониманий нашего предложения состоят в том, что оно берётся слишком буквально, слишком жёстко. Мы явно не смогли объяснить достаточно ясно, что [в рамках нашей методики при построении этограммы] каждый новый случай потребует креативности в поиске дополнительных или альтернативных признаков во времени и пространстве, особенно для новых видов, отдалённо родственных данному, для которого этограмма уже существует, чтобы отрегулировать систему признаков к его телодвижениям.
Например, был поднят вопрос, что мы будем делать с мерцанием светлячка или со вспыхиванием и сменой цветов у головоногих. Я сожалею, что не включил в нашу перепелиную этограмму сонаграммы вокализаций, где применим тот же подход (поведение 38, 51, 52, тревожный крик и различные варианты контактного крика). По сравнению с мерцанием или вспыхиванием они с большей очевидностью будут представлены в визуальном домене в графике, сходной с нашими наброскам устойчивых поз (climax postures) и/или в терминах числовых значений (темп повторения, диапазон частоты и т.д.). Радует, однако, что наш формат был оценен как «легко адаптируемый» для переднежаберных (Leonard, Lukowiak, 1985).
Другое непонимание провоцируется нашим списком 17 состояний признаковых переменных для каждого поведения. Мы сделали так поскольку в ранних версиях, я подозреваю, были слишком оптимистичны в наших оценках применимости нашей системы кодирования. Было заявлено, что ещё многие признаковые переменные нуждаются в отделении от наших 60 единиц поведения. Относительно расточительное представление кодов в верхней части нашей таблицы было выбрано чтобы сделать систему более ясной. Эти коды не были предназначены для возбуждения толков [о выделений признаков, т.к. значимы не переменные а понимаемые через них образы], но взамен служит, в сочетании с рисунками, заменителем длинного описания и потенциально как переменные для контроля за анимированным перепелом на компьютерном экране (см. Schleidt et al., 1984, 198-202).
Если было принято стандартное кодирование, этограмма расписного перепела (и что важно, многих других видов животных) может быть сокращена до 2 стр. в этом журнале и потенциально может печататься штрих-кодом, какой можно считывать на компьютерах световым карандашом, и до 1 стр.рисуночной информации. В этой форме, публикация этограмм станет осуществимей, тем более что проза жизни состоит в следующем: их принимают лишь в исключительных случаях (как показали Леонард и Луковяк, по статистике отказов редакций поведенческих журналов).
Я принимаю валидность критицизма общего характера что [ориентируясь только на форму единиц, противопоставленную «фону»] мы упустили средовые и контекстуальные референты [сопоставляемых каждой из единиц в такой этограмме]. Однако я предлагаю заниматься проблемой по одному шагу за раз, оптимистически веря, что паттерны таксон-специфического поведения остаются элементарными единицами поведения в биологии (units, единственное оформленное подразделение в составе отслеживаемой активности особи), как химические элементы или биологические виды заняли своё место в химии и таксономии, несмотря на продолжающиеся дискуссии интересующиеся деталями которые эти концепции представляют.
Нельзя не заметить, однако, что информация об ориентации [демонстратора] относительно специфического объекта или положения, вызывание или сдерживание внешними «пространственно-временными констелляциями» (объекты, события, положения и ситуации, Шлейдт, 1985) может быть добавлена к любому поведенческому элементу без необходимости изменения алгоритма. Например, с ориентацией в терминологии 26–стороннего тела [сторона – направление изменений облика по одной из «степеней свободы», связанной с движением того или иного органа] ориентация в терминологии специфического объекта [еда, хищник, брачный партнёр, кладка и т.д.] может быть добавлена, если там применимо, к туловища или голове (переменные TrO или НО). А пространственно–временные констелляции, вызывающие данные единицы (например, специфические «релизеры» тем более это обычно другие юниты из того же списка) можно добавлять к каждому поведению как дополнительную переменную.
Кто- то может захочет вспомнить фон Юкскюля и Гибсона относительно средовых признаков и соответственно пытаться составить частичный репертуар релевантных вызывающих факторов для каждого вида. «Экограмма» самое подходящее название для такого списка. Я думаю, осмысленно ждать, что у каждого вида и каждой особи этограмма и экограмма взаимосвязаны специфическим образом что само по себе может рассказать нам что-либо интересное про связь поведения и среды.
Я склоняюсь к прагматическому подходу. Я не думаю, что «теории контекстно–зависимого поведения» могут рассматриваться (а тем более быть) альтернативой нашему подходу «больше этограммы» [(в смысле, обоснованней выделение единиц, точней их разграничение и пр.] (Gordon, 1985). Теории непригодны как замена поведенческим данным. Мы ищем общую теорию поведения, как пучок согласующихся между собой и всеми принятых априорных утверждений [обычно основанных на некой «более общей» теории, общепринятой здесь и сейчас, почему полагаемой верной] что ведёт нас к «объяснению» поведения, к прочерчиванию «стволовых» линий причинности в комплексной сети причин и следствий [управляющих поведением].
Однако, такие объяснения обречены оставаться чисто теоретическими, если мы не можем разрешить совпадающим [с природой] образом проблему строгого описания базовых поведенческих наблюдений. Если «контекст», в котором встречается частная форма поведения, суть значимый признак, его надо включить, если нет, смотреть на что-либо ещё интересней. Однако, что ни обозначь ярлыком «контекст», никак не уйти от принятия того или иного стандарта для эффективного описания поведенческих наблюдений [нельзя оставлять это на произвол «схватывания» авторским впечатлением].
В исторической перспективе мы видим общее принятие поведенческих паттернов как «кусочков» поведения, нечто среднего между движением отдельных мускулов и целыми инстинктами, как элементарными составляющими реализуемого поведения. См. Schleidt, 1974. Оглядываясь назад по меньшей мере на полстолетия [в историю этологии, включая историю] практического использования таких паттернов (в) для описания видовых сигнальных репертуаров, видим полный склад сильно избыточной информации (избыточной из–за некачественного использования логики, слов и чернил), и ещё больше потеряли информации в уже никак не восстановимых регистрациях. Мы должны остановить этот мусор.
Ещё есть время чтобы перестроиться в сторону большей эффективности сбора и хранения данных. Если мы согласны, что р<0.05 отвергает нуль–гипотезу о неструктурированности потока в пользу рабочей [о разделении потока активности, отслеживаемого этологом, на данные единицы и «фон»], мы договоримся также о неком «уровне согласия» вклада признаковых переменных в наблюдаемые паттерны поведения [по превышении которых он признаётся), см. предложенный нами 95%-ный интервал вокруг средних признаковых переменных, см. Schleidt, 1972, с.236–237. Давайте создадим базовый стандарт для хранения поведенческой информации и найдём пути достижения этой цели.
Источник Zeitschrift für Tierpsychologie (ныне Ethology). 1985. V.68. №4.
***
Однако этологи не обратили внимание на решение, устраняющее проблемы их собственного метода: сменившая её социобиология, в период критики указавшая на этот изъян, также не восприняла это решение, а, победив, использовала прежний сравнительно-этологический подход к выделению единиц по «устойчивому впечатлению» наблюдателя со всеми его минусами. Такая вот грустная история, как в силу причин вненаучного характера сравнительные этологи сами отказались от развития исследовательской программы, более перспективной и лучше соответствующей эмпирии, чем социобиологическая конкурентка, даже в условиях, когда проблемы, вскрытые её критикой, были (Вольфгангом Шлейдтом и независимо от него Ноа Эшкол-Абрахамом Ваксманом в Израиле) разрешены.
Тогда социобиология была на подъёме, предельно модной, особенно среди научной молодёжи, этологию стали считать устарелой и — что немаловажно — присущие ей большие натуралистические статьи делались всё неудобней для публикации, а публиковаться требовали всё настойчивей — и его достижения остались втуне.
Я с ним переписываюсь, поскольку 40 лет спустя решаю ту же проблему (несколько по другому, о чём сейчас пишу книгу). Надеюсь что за это время негодность социобиологических концепций стала ясна, плюс исследования в рамках данной парадигмы добыли массу полезного знания, которое почти всё больше подходит для развития сравнительно-этологических теорий (исследования в рамках которых не ведутся с конца 1980-х, кроме некоторых одиночек) чем теорий «родной ей дисциплины», закрывают многие концептуальные изъяны в первых (о них см. первую часть статьи «Как социобиология себя отрицает») и, напротив, высвечивают их во второй.

Первое издание системы описания движений Эшкол-Ваксмана
«Хореографическое» решение той же проблемы в Израиле
Одновременно со Шлейдтом ключ к решению этой проблемы подобрали Ноа Эшкол и Абрахам Ваксман в Израиле. Сперва они задались способами объективного описания составляющих танцевальных движений у человека, иерархии их соподчинения в процессе танца, и создали так называемую систему описания (тело)движений Эшкол-Вахмана (EWNM). Cперва её использовали для задач хореографии (объективное описание телодвижений танцоров и пространственно-временной структуры танца), а затем применили к «хореографии» обмена демонстрациями во взаимодействиях животных, вроде «ритуализированной борьбы» у волков, в понимании двигательно-координационных проблем, следующих из дефектов социальной коммуникации у аутистов, разработки способов их терапии и пр. Здесь наиболее известны работы Илана Голани 1970-80-х гг.

Ноа Эшкол и Абрахам Ваксман, 1955 г.

Работа И.Голани, посвящённая паттернам телодвижений, структурирующим демонстративное поведение обыкновенного шакала Canis aureus
Система сначала была разработана для записи танца, позволяющей полностью воспроизвести его в соответствие с той же музыкой и музыкальным ритмом. В отличие от других систем записи она предназначена для фиксации любых скоординированных телодвижений в группе, не только танцевальных, в данном методе нет ограничений определёнными танцевальными стилями и даже движениями человеческого тела вообще – те же принципы успешно переносятся на животных (пример+другие работы Илана Голани).
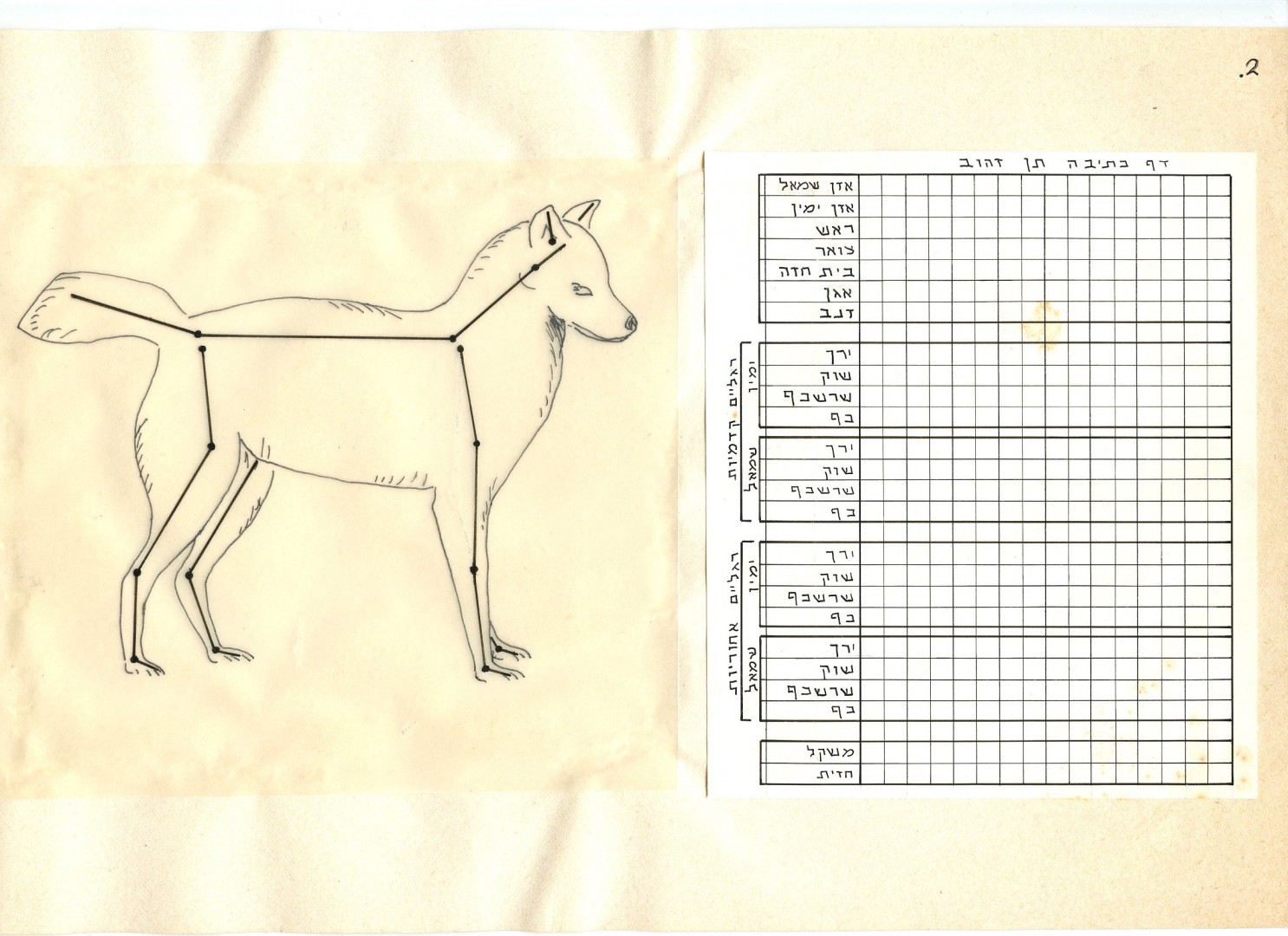
EWMN рассматривает тело как фигуру из палочек. Оно делится на единицы (которыми производятся элементарные движения) по сочленениям скелета; каждая пара соединений (или соединение и окончание) даёт линейный сегмент («член»). Например, foot – это член, ограниченный лодыжкой и концом носка (рис.2). В нашей классификации это соответствует изменению положения части тела птицы – крыла, шеи, хвоста и пр. на единичную позицию в данном направлении изменений, заданного разными «степенями свободы» изменения внешнего облика птицы (отведение крыльев, вытягивание шеи, качание корпусом и пр.). См. «От стимула к символу«, ч.2, разд.1.1.
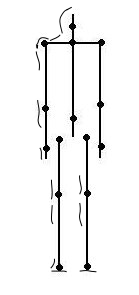
Рис.2
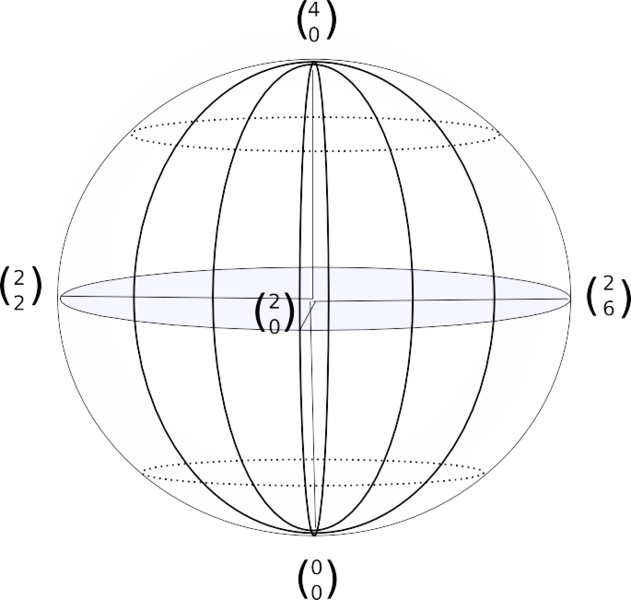
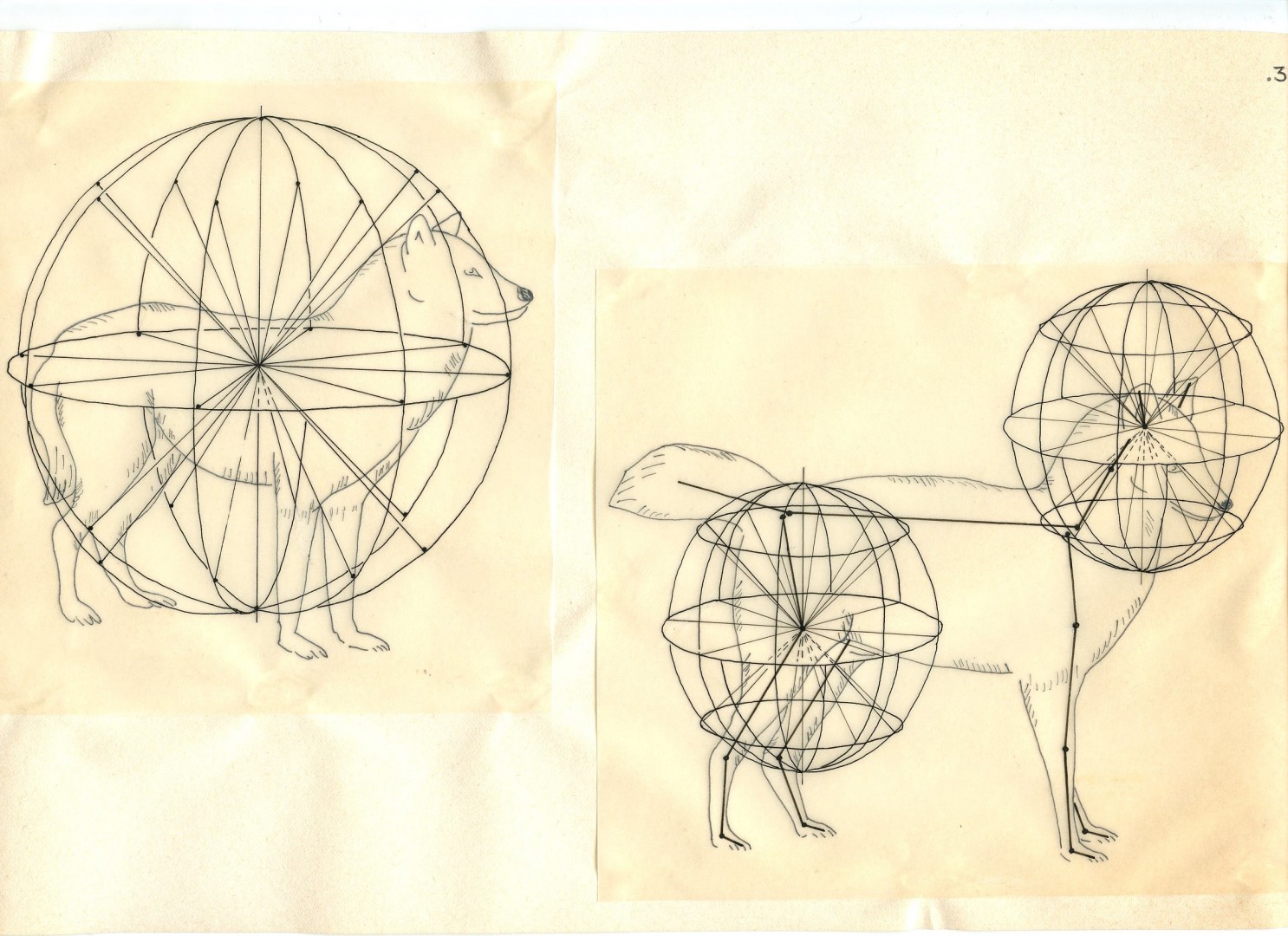
Для характеристики (и записи) пространственных отношений членов используется сферическая система координат. Если один из концов сегмента удерживается в фиксированной позиции, он служит центром сферы, радиус которой равен длине данного члена. Позиция свободного конца сегмента определяется двумя сферическими координатами, наподобие широты и долготы на глобусе.
Позиция самой конечности описывается комбинацией вертикального и горизонтального номера, где вертикальный записан над горизонтальным. Нижний горизонтальный компонент считывается первым. Эти два номера заключаются в квадратные или обычные скобки, чтобы показать, определяется ли позиция по отношению к примыкающему члену или по отношению ко внешнему ориентиру, такому как помост.
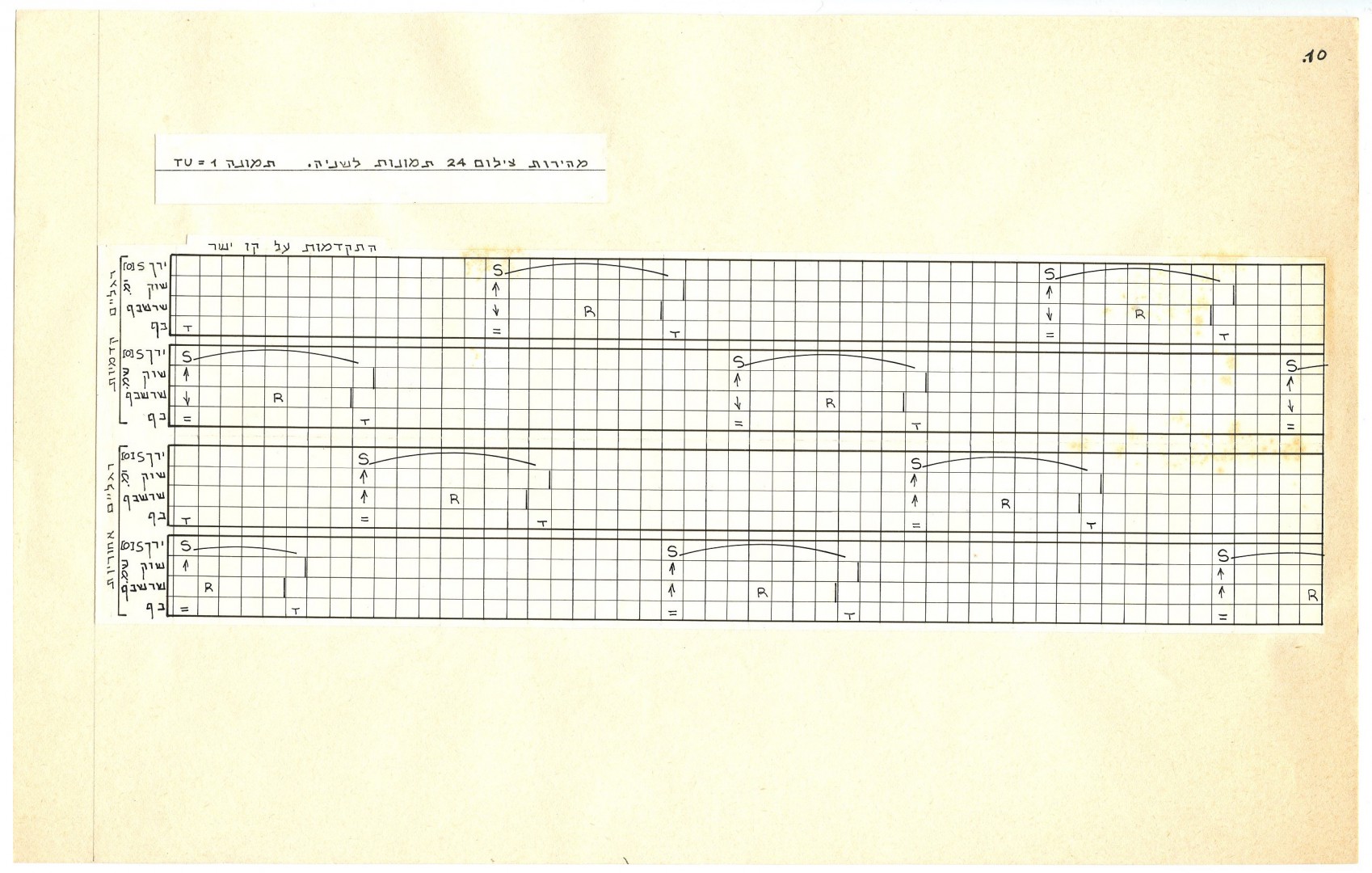
Знаки нотации записываются в сетке, где горизонтальная строка фиксирует положение и движение одной конечности, а каждая вертикальная колонка представляет единицу времени. Движение записывается как переход между координатами начала и конца (рис.3).
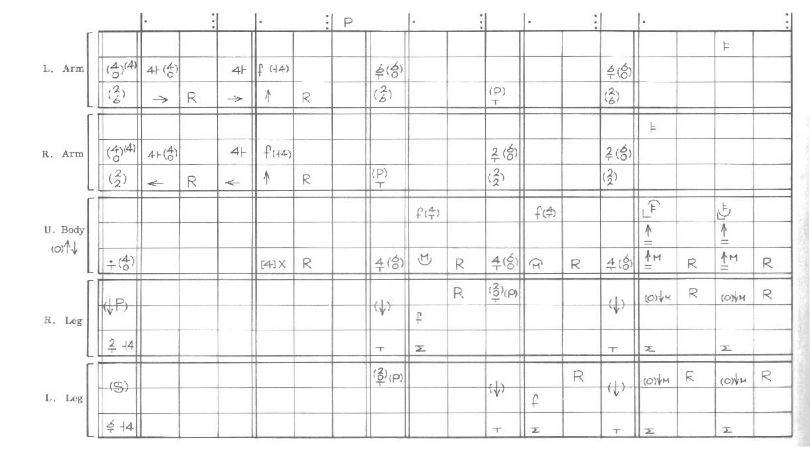
Рис.3. Образец записи телодвижений.
История EWMN
Книга с её описанием впервые вышла аж в 1958 г. Ноа Эшкол написала пять танцев, предназначенных для исполнения без музыки, для сосредоточения на вызываемых ими внутренних эмоциях. Ещё раз подчеркну, что с самого начала это была система записи телодвижений, а не танца – во втором случае функциональный момент неизбежно привносится в описание. Она считала, что на основании анализа движений мы можем понять, как те или иные движения вызывают те или иные чувства и что использование записи движений поможет открыть законы композиции для определённых танцевальных стилей, наподобие того, как это уже было сделано для музыки и других видов искусства.
Тело. Как изображение для всех тел предлагается абстрактное тело «человека без свойств». Каждый член тела изображается осевой линией постоянной длины (рис.2).
Закон «легких» и «тяжёлых» соединений. Когда человек ходит ногами, остальная часть тела (туловище, руки, голова) следует за движением ног. В EWNM это соответствует закону «лёгких» и «тяжёлых» соединений. Структура тела рассматривается как ветвящаяся связь. База – это «тяжёлый» сегмент тела. Когда «тяжёлый» член движется, он несёт все присоединённые к нему «лёгкие» сегменты. При вертикальном положении ступни рассматриваются как самый «тяжёлый» член, далее ноги, далее – туловище (рис.4), т.е. движущие части тела «тяжелее» движимых, а те, соответственно, «легче» (рис.5).
Рис.5
Рис.4
Вид страницы записи. EWMN записывается, а не рисуется. Как уже говорилось, движения записываются на горизонтальных строчках. Вертикальные линии делят таблицу на колонки, обозначающие единицы времени. Символы движения записываются в определённом порядке, слева направо. См. порядок членов, которого придерживаются по умолчанию (рис.2). Настройка является гибкой, что позволяет разделить тело на разные части в зависимости от того, запись каких движений необходима. Движения, записанные в нотации Эшколь-Вахмана, могут быть положены на музыку. Но это не является необходимым.
Система отсылок (SOR). Свободный конец каждого члена описывает криволинейную траекторию на поверхности сферы, размеченной координатами долготы и широты (рис.6). Одна из позиций на сфере выбирается в качестве стартовой и обозначается 0, по мере изменения позиции на 45о добавляется по 1. Положение и движение конечностей определяется координатами SoR (см. примеры записи 1-2-3). Для документирования перехода между статическими позициями система учитывает движение, количество движения, пространственную ориентацию и движение по или против часовой стрелки.
Типы движения. Выделены 3 типа движений. Первый – вращение члена вокруг своей оси без изменения положения в пространстве, «вращение дверной ручки». Второй – «движение самолёта» — кратчайшее перемещение между двумя вариантами положения члена в пространстве. Примером третьего типа движения является вращение хула-хупа, это «коническое движение».
Применения. EWMN была использована для обозначения движения рук и пальцев на языке жестов, композиции танцев, народных танцев, в медицинской области, записи ухаживательного поведения шакалов и других этологических исследований. Она использована также в системе физиотерапии Моше Фельденкрайса, применяющейся при болях или ограниченной подвижности.
Метод применялся в визуальном и кинетическом искусстве, была создана компьютерная система, способная построить любое движение, записанное в EWMN. Система может быть использована в робототехнике, анимации или при съёмках фильмов. Она применялась для обнаружения раннего появления движений, являющихся предвестниками аутизма. Исследования Prof. Philip Teitelbaum и Osnat Teitelbaum в Университете Флориды было целиком основано на применении системы для изучения детского движения и показало, что специфические закономерности движения, проявляющиеся в первые несколько месяцев жизни, являются надёжными предвестниками появления в дальнейшем аутизма и синдрома Аспергера.
Илан Голани
Затем Илан Голани и другие авторы распространили этот метод на выделение единиц (демонстраций) в потоке активности взаимодействующих животных, и чисто структурное описание их формы, с критериями отделения друг от друга и от «фона»). См. «характерные кадры» телодвижений шакалов и пример записи их поведения в системе EWMN.

 Здесь и далее источник
Здесь и далее источник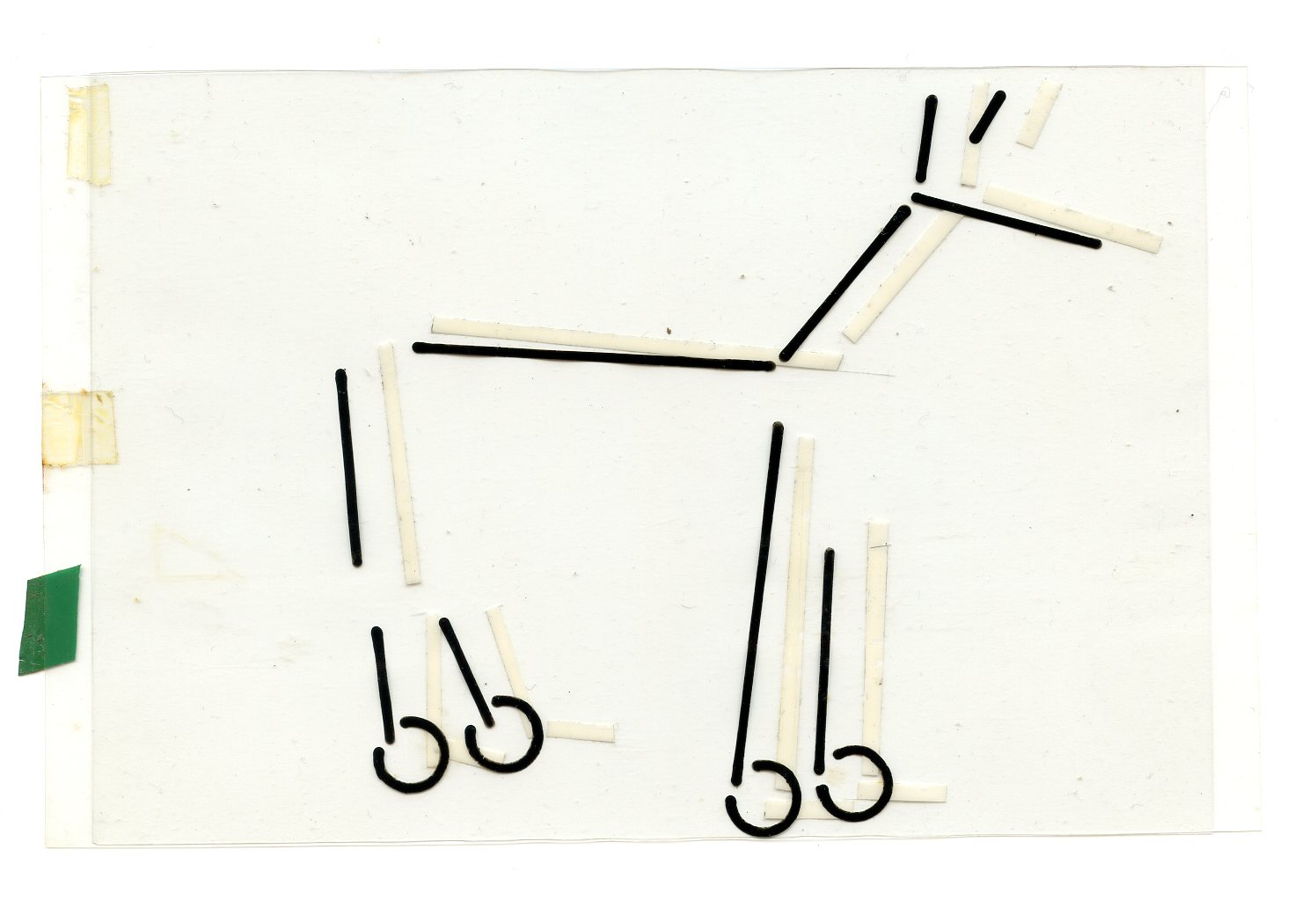


 Другой пример — анализ движений волков в ритуализированной борьбе, впервые произведённое Шенкелем в 1947 г., а Голани с коллегами переисследованное. Процитирую из «От стимула к символу»:
Другой пример — анализ движений волков в ритуализированной борьбе, впервые произведённое Шенкелем в 1947 г., а Голани с коллегами переисследованное. Процитирую из «От стимула к символу»:
«С одной стороны, отсутствие адекватного языка описания, превалирование функционального подхода над структурным, привело к смешению формальных, целевых, функциональных и мотивационных критериев в описании демонстраций, так что выделенные сигналы даны в первом приближении и требуют переописания. С другой — Шенкель верно уловил устойчивую связь сигнала и ответа в том процессе коммуникации, которым является «ритуальная борьба». Им показано, что «связанные конфигурации» демонстраций сигнала и ответа партнёра оказываются более устойчивым и однозначным (по форме) комплексом действий, чем каждая из этих демонстраций, взятая не в сопряжении с другой, а в одиночестве, когда волк демонстрирует не в ответ на сигналы партнёра (Golani, 1976).
Далее, в «ритуализованной борьбе» волков структура коммуникативных связей между сигналами и ответами индивидов (то есть управляющие воздействия процесса, опосредованные сигналами) надёжней прослеживается и легче характеризуется типологически, чем структура последовательностей демонстраций каждой особи (выражающая эндогенные изменения мотивации животного и/или интенсивности стимуляции партнёра). Вторая вне первой практически не выделяется, поэтому при описании сигнального репертуара «ритуализованной борьбы» следует выделять «связанные конфигурации» одновременных демонстраций партнёров (Golani, 1976; Moran et al., 1981)
Работа G.Moran, J.Fentress, J.Golan (1981) демонстрирует возможности структурного подхода к выделению таких «связанных комбинаций» движений волков в ритуализованной борьбе и тем самым его способность объективно анализировать процессы обмена демонстрациями даже в «трудном случае», когда животные используют недифференцированную систему сигналов-стимулов. Из семи переменных, использованных для описания, первые три описывают связи между движением двух волков: дистанция между животными, взаимная ориентация тел и точки противостояния между партнёрами (участки их тел, обращённые друг к другу). Другие четыре переменные описывают развёртку поведения особи в пространстве как последовательность развития демонстраций и последовательные изменения ориентации особей.
Комбинация семи переменных формирует трёхмерное пространство взаимодействия, где отдельные демонстрации (конфигурации тел взаимодействующих волков) соответствуют отдельным областям. Показано существование четырёх наиболее устойчивых конфигураций, в которых демонстрации оппонентов строго сопряжены, и восьми областей переходов между ними, в которых демонстрации неустойчивы, несопряжены с демонстрациями партнёра и животные их стараются как можно быстрее сменить, перейдя в одну из «устойчивых областей». Переход через «области неустойчивости» совершается участниками независимо друг от друга, но демонстрации, соответствующие «областям устойчивости», строго сопряжены (Moran et al., 1981; Golani, Moran, 1983).
Постоянство всех девяти областей, занимаемых ритуализованными паттернами в пространстве взаимодействия показывает, что движения участников в значительной степени обусловлены одновременными демонстрациями партнёра. Социальная роль животного определяет индивидуальный вклад каждого партнёра в координацию действий и управление процессом перехода от одной «связанной конфигурации» к другой. Как в других аналогичных исследованиях, доминант оказывается управляющим процессом развития взаимодействия в целом, субъектом происходящей смены сигналов, подчинённая особь, проигрывающая взаимодействие – управляемым объектом тех же смен (Moran et al, 1981)».
Что следует из всего этого?
«Пресечение этологических исследований не позволило распространиться и этим подходам среди тех, кто исследует видоспецифическое поведение животных в природе. Подобную ситуацию пора исправлять, что я и собираюсь сделать в ближайшие годы.
Если же обособленность элементов в поведении особей невелика, и дифференцированных элементов нет, то сигнально значимым будет поток поведения в целом или отдельные характеристики непрерывного процесса (например, изменения скорости демонстрирования во времени или разнообразия элементов поведения в неких «выделенных точках» процесса, соответствующих демонстрациям чисто условно). Ни об объективности существования, ни о «форме» сигнала здесь уже говорить не приходится, и сам «сигнал» выделяется исключительно для удобства классификации.
Вне сферы социальной коммуникации (просто при специфическом возбуждении животного), отсутствует та совместность и одновременность исполнения элементарных движений, которая образует демонстрацию. Специфические структуры процесса здесь «складываются» только случайно, соответствующая им комбинация элементарных движений «выпадает» редко и держится очень недолго из-за статистической невероятности устойчивого сохранения пика возбуждения, поддерживающего данную комбинацию, в том случае, когда пик возник эндогенно, а не установлен и удерживается направленным воздействием сигналов извне. Во втором случае эта «невероятная» комбинация складывается закономерно, маркирует специфику проблемной ситуации, созданной последней демонстрацией партнёра, и стереотипно удерживается до тех пор, пока один из участников не воспримет информацию, связанную с данным сигналом, и не скорректирует «в нужную сторону» собственную модель поведения выбором следующей демонстрации. Так доминошник корректирует игру выбором и выкладыванием следующей кости.
Это заставляет нас видеть демонстрации специфическими «значащими структурами» процесса взаимодействия, а не «пеной» на гребне волны общего возбуждения «втянутых» туда индивидов, как это делает Е.Н.Панов (1978). Поэтому демонстрации должны обладать специфичностью формы, выделенностью из потока событий, и быть специализированными к исполнению сигнальной функции, что и подтверждается в соответствующих исследованиях. Другие наблюдения за демонстрированием у птиц, млекопитающих, ящериц и костистых рыб также обнаружили столь высокую стереотипность исполнения данных элементов, что переход от одной демонстрации к другой возможен только скачком и лишь через неритуализованное состояние обыденной позы. См. Dane et al., 1959; Simpson, 1968; Schleidt, 1982; Schleidt et al., 1984; Ord, Evans, 2003; Peters, Evans, 2007. Это и делает демонстрацию дискретной структурой.
Всё вышесказанное позволяет дать положительный ответ на четыре вопроса, вокруг которых в 1970-х-1980-х гг. развернулась интенсивная дискуссия о природе и коммуникативной функции демонстраций: насколько формы последних существуют объективно в потоке действий животного, устойчиво реализуясь в специфических ситуациях взаимодействия? Насколько объективна дифференцированность демонстраций друг от друга и их всех вместе взятых – от несигнальных действий животного? И если да, насколько дискриминация демонстраций (элементов с потенциальной сигнальностью) друг от друга и от «фона» существенна для самих особей, насколько успешно телодвижения демонстратора способы эту дискриминацию провести, насколько она существенна для участников коммуникации и для эффективности всего процесса?
По-видимому, правы классические этологи, отстаивавшие дискретность демонстраций и наличие у них специфических инвариантов формы, точность воспроизведения которых достигается за счёт стереотипности демонстрирования. И наоборот, неправы их критики, отрицавшие всё вышесказанное (главы 2.1-2.3 книги).
Другое дело, что концепции классической этологии обладали одним ключевым недостатком (который не преминул отметить такой внимательный критик, как Е.Н.Панов, 1978, 2005а). В выделении демонстраций они целиком и полностью полагались на «устойчивое впечатление» квалифицированного наблюдателя и так и не создали формального метода описания форм демонстраций, дифференциации их друг от друга и от «фона» по данным непрерывной регистрации поведения, позволяющего это делать объективно и аналитически, — так, как было показано выше.
К сожалению, все работы, в которых объективируется процесс выделения демонстраций, и на основании которых можно сформировать методы описания сравнительной этологии, адекватные объекту данной дисциплины (Moran et al., 1981; Golani, Moran, 1983; Finley et al., 1983; Schleidt, Crowly, 1980; Schleidt et al., 1984; Фридман, 1992а, 1993а, 1994), существенно запоздали. С конца 1980-х гг. интерес к сравнительному анализу поведения резко упал и к концу 1990-х практически упал до нуля (Панов, 2005а) [см. также рис.7 из статьи Е.Н.Панова, 2005а].
Сейчас, по сути, его надо восстанавливать заново, опираясь на те работы по объективному описанию инвариантов формы сигнала, которые с других позиций решают те же задачи выделения сигнальных инвариантов в изменчивых телодвижениях животного, образующих «демонстрацию», количественной оценки инвариантности формы сигнала, степени дискретности его материальных носителей – демонстраций и пр.

Литература
Dane D., Walcott C., Drury W.H., 1959. The form and duration of the display action of the golden-eye (Bucephala clangula)// Behaviour. Vol.14. P.265-281.
Enquist M., Ghirlanda S., 2005. Neural networks and animal behavior // Monographs in behavior and ecology. Princeton University Press. 253 pp.
Enquist M., Ghirlanda S., Hurd P.L., 1998. Discrete conventional signalling of continuously varying resource value // Anim. Behav. Vol.56. P.749–753.
Finley J., Ireton D., Schleidt W., Thompson T., 1983. A new look at the features of mallard courtship display// Anim. Behav. Vol.31. №2. Р.348-354.
Ghirlanda S., Enquist M., 1998. Artificial neural networks as models of stimulus control // Animal Behaviour. Vol.56. P.1383-1389.
Golani I., 1976. Homeostatic motor process in mammalian interactionbs: a choreography of display// Perspectives in ethology. Vol.2. P.69-104.
Golani I., Moran G., 1983. A motility-immobility gradient in the behavior of the «inferior» wolf during «ritualized fighting» // Advances in the study of Mammalian Behavior (Eds. J.F.Eisenberg & .G.Kleiman). American Society of Mammalogists Special Publication. American Society of Mammalogists, Shippensburg, PA. Vol.7. P.65-94.
Halliday T., 1975. An observational and experimental study of sexual behaviour of the smooth newt Tritirus vulgaris (Amphibia, Salamandridae)// Anim. Behav. Vol.23. №2. Р.291-322.
Lorenz K., 1939b. Vergleichendes über die Balz der Schwimmenten // Journal fur Ornithologie. Bd. 87. S.172—173.
Moran G., Fentress J.C., Golani I., 1981. A description of relational patterns of movement during ritualized fighting in wolves (Canis lupus) // Animal Behavior. Vol.29. P. 1146-1165.
Peters R.A., Evans Ch.S., 2003а. Design of the Jacky dragon visual display: signal and noise characteristics in a complex moving environment// J. Comp Physiol. A. Vol.189. P.447–459.
Peters R.A., Evans Ch.S., 2003b. Introductory tail-flick of the Jacky dragon visual display: signal efficacy depends upon duration// Journal of Experimental Biology. Vol.206. P.4293-4307.
Peters R.A., Davis C.J., 2006. Discriminating signal from noise: recognition of a movement-based animal display by artificial neural networks //Behav. Process. Vol.72. №1. Р. 52-64.
Peters R.A., Evans Ch.S., 2007. Active space of a movement-based signal: response to Jacky dragon (Amphibolurus muricatus) display is sensitive to distance, but independent of orientation // Journal of Experimental Biology. 210. P.395-402.
Schleidt W., 1982. Stereotyped feature variables are essential constituents of behavior patterns// Behaviour. Vol.79. P.230-238.
Schleidt W., Crawley J.N., 1980. Patterns in the behaviour of organisms// J.Soc. and Biol. Structures. Vol.3. №1. Р.1-15.
Schleidt W., Yakalis G., Donnelly M., McCarry M., 1984. A proposal for a standard ethogram, exemplified by an ethogram of a blue-breasted quail (Coturnix chinensis)// Zeitschr. für Tierpsychol. Bd.64. H.3-4. S.193-220.
Рекомендуем прочесть
 Как птицы обучаются инстинктивному поведению
Как птицы обучаются инстинктивному поведению Синдром социальности: сравнительное исследование поведения песчанок
Синдром социальности: сравнительное исследование поведения песчанок Что не так с социобиологией?
Что не так с социобиологией? Ритуализация «по Благосклонову» и её сопряжение с «тинбергеновской»
Ритуализация «по Благосклонову» и её сопряжение с «тинбергеновской» «История идей» сравнительной этологии
«История идей» сравнительной этологии К этологии леммингов
К этологии леммингов «Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования
«Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования Зачем большим пёстрым дятлам всю зиму охранять территории?
Зачем большим пёстрым дятлам всю зиму охранять территории?
