

В продолжение социальной релятивности действия гормонов приведены данные об этой релятивности для окситоцина, часто (и ложно, ибо «биологическая» агрессивность мужчин и женщин не отличается, это не мыши, и не обыкновенные шимпанзе) называемого «гормоном любви и счастья», ответственным за «доброту, миролюбие, верность» и прочие позитивные эмоции, связанные с просоциальностью и кооперацией, включая взаимодействия людей-хозяев с домашними животными.
Он только закрепляет и усиливает уже существующие социальные объединения и разделения, будь то группы или отдельные (знакомые) личности, однако не создаёт их, последнее — целиком сфера действия социальных факторов. Поэтому он усиливает позитивное отношение к «своим», ухудшает к «чужим», одновременно способствуя различению тех и других, т. е. социальной категоризации. И наоборот, изменение тех или других отношений, требует прежде всего изменения социальных норм, если индивид им следует, или дополнительного социального влияния на человека, чтобы он перестал противостоять этих нормам, иначе окситоцин бессилен.
Однако формы таких отношений и способы их реализации во взаимодействиях между людьми — или мышами, крысами, другими исследуемыми особями — определяются социально, наличным репертуаром телодвижений и действий в соответствующих контактах.
На деле зависимость между социальной связанностью и окситоцином обратная — каждый акт подтверждения и укрепления этой связанности, происходящий в социальных контактах (а они в свою очередь следствие существующих представлений и культурных норм) стимулирует выделение гормона и усиливает специфичность воздействия.
Отсюда необязательность окситоцина для прочности парных связей у знаменитых моногамных полёвок Microtus ochrogaster и забавное следствие для людей: их национальные общности, как и объединения на основе религии основаны на иллюзии, это воображаемые сообщества, в отношении наций верен конструктивизм, а примордиализм нет. Однако «конструкция», раз созданная и воспроизводимая средствами социальной трансляции (воспитание в семье, обучение в школах, индоктринация в социуме через книги, карты, музеи и прочий печатный капитализм) «склеивают» людей в системное целое несравненно сильней, чем любой из мыслимых биологических факторов общности, «мы-концепция» управляет биологией, а не наоборот.
Социальное влияние — самое сильное из того, что создала биологическая эволюция ещё у животных, и ещё больше усилила социальная история уже у нашего вида. Поэтому расовые предубеждения ослабляются или исчезают, когда детей убеждают думать о соответствующих различиях как межиндивидуальных, а не межгрупповых (и это имеет долговременный эффект). А всё вышесказанное про гормоны и социальные влияния также верно для разных видов позвоночных животных, различающихся пространственно-этологической структурой популяций.
Новый взгляд на механизмы доместикации: окситоцин как маркер поведения
Содержание
Гербек Ю.Э.
Резюме. Доместикация животных – важный этап в истории человечества, который совпал, а, возможно, и способствовал переходу от охоты и собирательства к сельскому хозяйству. Главным фактором эволюционных изменений на первых этапах доместикации был, по-видимому, отбор по поведению по отношению к человеку: сначала естественный, а затем бессознательный, искусственный. Согласно концепции акад. Д.К.Беляева этот отбор был направлен на системы, которые регулировали поведение, снижая ответ на стресс реакцию и агрессию, а затем формируя эмоционально-положительную реакцию на человека. В статье обсуждается ряд работ последнего десятилетия, указывающие на возможную ключевую роль центральной окситоциновой системы не только в снижении агрессии и стресс-ответа, но и в формировании социальных взаимоотношений между человеком и доместицированными животными.
Одомашнивание растений и животных – одно из самых значительных культурных и эволюционных событий в истории нашего вида за 300 000 лет (т.е. с начала его существования). В процессе одомашнивания человек создал для себя новую среду, значительно повлиявшую на все стороны его жизни. Доместицированные животные стали важным источником пищи, материалом для изготовления одежды, средством передвижения и физической работы. Важнейшим изменением, произошедшим при доместикации животных, являлось появление дружелюбия по отношению к человеку («ручного» поведения), которое не наблюдается в дикой природе.
Собака, по-видимому, первое доместицированное животное. По археологическим и генетическим данным наиболее ранняя граница доместикации животных – предковсовременных собак – 16 – 17 тыс. лет назад. Важным отличием собаки от всех остальных доместицированных животных является то, что её доместикация, по-видимому, происходила задолго до возникновения сельского хозяйства, в среде охотников (Freedman et al., 2014; Guagin et al., 2017).
Выделяют два основных периода доместикации животных. Первый совпадает, а возможно тесно связан с началом производством продуктов питания – 12 000-9 000 лет назад в конце последнего ледникового периода и во время перехода к настоящему межледниковому периоду. В этот период доместикация происходила в основном на территории Ближнего Востока, а также современного Китая (это свинья, корова, коза, овца и кошка). Второй период – от 7 000 до 4000 лет назад – связан с развитым сельским хозяйством и происходил в основном в таких регионах, как Центральная и Восточная Азия (Larson et al., 2014). В этот период были доместицированы такие животные, как лошадь, курица, верблюд и зебу.
Если доместикация животных в первый период, по-видимому, происходила путём бессознательного отбора на отсутствие агрессии и дружелюбность к человеку, а, возможно, и путём естественного отбора на адаптацию к новой среде обитания рядом с человеком (Беляев, 1981; Коппингер, Коппингер, 2005; Larson et al., 2014), то второй этап доместикации, очевидно, происходил под действием сознательного отбора, причём не только на дружелюбное поведение, но и на хозяйственно-полезные признаки.
Ранние этапы доместикации, как указывал акад. Д.К. Беляев, были связаны со снижением, а затем исчезновением агрессии и стресса-ответа при появлении человека, которые свойственны диким животным (Беляев, 1981). Наиболее показательной в этом ключе является собака. Как описывал в своих наблюдениях известный исследователь собак Ричард Коппингер (Coppinger, Coppinger, 2002), полудикие собаки, по сравнению с волками и другими дикими животными имеют значительное преимущество, когда питаются на свалках, не убегая, едва завидев человека. Им не надо охотиться, чтобы добыть еду, а, значит, не надо тратить на выслеживание и преследование добычи много дополнительной энергии. Таким образом, отбор на толерантное отношение к человеку – это первый и важнейший шаг в адаптивном поведении.
При этом, как полагал Д.К.Беляев снижение стресса-ответа на человека, а возможно стресс-реакции вообще должно было способствовать этой адаптации (Беляев, 1981). К сожалению, за исключением единичных прямых сравнений стресс-ответа у доместицированных животных и их диких сородичей, таких как свинья, морская свинка, курица и певчие птицы (Weiler et al., 1998; Künzl,Sachser, 1999; Price, 2002; Suzuki et al., 2014; Ericsson, Jensen, 2016), существуют только косвенные подтверждения этих различий.
Одним из основных доказательств снижения стресс-ответа при доместикации являются результаты, полученные под руководством Д.К. Беляева и его ученицы и ближайшей соратницы проф. Л.Н. Трут на всемирно известной модели доместикации – серебристо-чёрной лисице. Эта модель получена путём многолетнего интенсивного отбора животных по единственному признаку – поведению по отношению к человеку, в начале на элиминацию агрессии, а затем на эмоционально-положительную реакцию (Беляев, 1981; Трут и др., 2004).
Однако в ту же сторону постепенно изменялись и недоместицируемые лисицы, на уменьшение агрессивности которой действовал не только бессознательный или сознательный отбор во всех зверосовхозах, но и естественный отбор на адаптацию к обитанию рядом с человеком. В неволе серебристо-чёрные лисицы разводятся с 1890-х годов и ведут своё происхождение с острова Принца Эдуарда, Канада (Westwood, 1989), хотя первые попытки разведения лисиц в России уходят в XVII в. Кроме того, отлов лисицы в разное время вёлся в разных регионах России – на Урале, в Якутии и Камчатке (Беспятых, 2009; Плотников, 2017). Чтобы поддержать снижающийся уровень агрессивности лисиц по отношению к человеку, необходимый в качестве контроля к интенсивному отбору на его снижение, был начать отбор по поведению по отношению к человеку в противоположном направлении – на усиление агрессии (Трут и др., 2004).
Важные результаты, полученные на этой модели указывали, что при отборе на ручное поведение происходит снижение активности гипоталамо-гипофизарно-надпочечниковой системы (ГГНС), одной из основных систем вызывающих ответ на стресс (Беляев, 1981; Трут и др., 2004). А также было показано, что нейромедиаторные системы мозга, связанные с агрессией также изменяют свою активность (Науменко и др., 1987). Однако эти данные дают представление только о самых ранних этапах даже не доместикации, а «прото-доместикации» – переходе к синантропному состоянию, естественном отборе на адаптацию к появившейся новой среде вблизи человека. Каковы же возможные механизмы возникновения не только эмоционально-положительных реакций, но настоящей социальной связи между разными видами – человеком и доместицированными животными? Одним из возможных ключевых механизмов может являться изменение центральной окситоциновой системы, которая способна одновременно снижать активность ГГНС и усиливать социальное поведение, а также обладает сильным эффектом на снижение тревожности (Neumann, 2008; Neumann, Landgraf, 2012).
Снижение агрессии под действием окситоцина, в частности, показано в исследованиях на отловленных из природы крысах Groningen 35-го поколения разведения в неволе, имеющих по-прежнему довольно большой разброс межсамцовой агрессии (Calcagnoli et al., 2014). Причём наибольший эффект снижения агрессии был обнаружен у самых агрессивных крыс, что может быть связано с большим количеством рецепторов окситоцина в мозге, чем у менее агрессивных (Calcagnoli et al., 2014). В целом, результаты, полученные на лабораторных животных, указывают на снижение агрессии под действием окситоцина, хотя есть и исключения (Jurek, Neumann, 2018).
Что же касается доместицированных животных, то результаты имеющегося на данный момент небольшого количества исследований демонстрирует противоположный эффект – усиление агрессии или ослабление дружелюбности под действием окситоцина. При его введения в нос собака может вести себя менее дружелюбно в ответ на угрожающее поведение хозяина (Hernádi et al., 2015). Показано, что введение окситоцина в нос является эффективным для попадания белка в мозг, минуя гемато-энцефалический барьер (Leng, Ludwig, 2016). Введение ОТ поросятам приводило к повышению агрессивных взаимодействий между конспецификами в стрессирующих условиях в возрасте 17 дней и 8 недель (Rault et al., 2013). У телок с эндогенным высоким уровнем ОТ было отмечено повышенное число как аффилиативных, так и, наоборот, агрессивных взаимодействий между собой (Yayou et al., 2015). Следует также отметить, что повышение агрессии при центральном введении ОТ наблюдалась у доминантных самцов squirrel monkeys (Winslow, Insel, 1991), а у шимпанзе ОТ в моче повышался при межгрупповых конфликтах, что, однако, сопровождается внутригрупповым сплочением (Samuni et al., 2017).
Исследование отдалённых эффектов неонатального введения окситоцина на агрессивность взрослых лисиц, селекционируемых на усиление агрессии по отношению к человеку, дало совершенно неожиданный результат. Также как у крыс Groningen, эффект снижения агрессии был наиболее сильным у самых агрессивных лисиц и отсутствовал у менее агрессивных. Но действие неонатального введения окситоцина на самых слабоагрессивных лисиц, практически толерантных к человеку оказалось обратным – агрессия усилилась (Гербек и др., неопубл. данные).
Таким образом, можно предполагать, что окситоцин мог играть роль в снижении агрессии к человеку на этапе прото-доместикации. Однако, можно ли предполагать, что он мог участвовать в формированииэмоционально-положительной реакции на человека на ранних этапах одомашнивания, если у слабоагрессивных лисиц, таксономически близкиз к собаке, он вызывает усиление агрессии? Однако и у исторически доместицированных животных введение окситоцина также может приводить к подобным эффектам. Вероятно, это связано с условиями проведения эксперимента. При стрессорных условиях, к которым относится и тест определения агрессивности лисицы, количество рецептора аргинин-вазопрессина 1A (AVPR1A) может повышатьcя (Grundwald et al., 2016), что может привести к связыванию окситоцина с этим рецептором (Koshimizu et al., 2012; Anacker et al., 2016). Известно, что AVPR1A способен повышать агрессию и усиливать стресс-ответ (Jurek, Neumann, 2018).
Поэтому в нестрессорных условиях обнаруженный эффект окситоцина на слабоагрессивных лисиц, возможно, не проявляется. Большое внимание в современном животноводстве уделяют благополучию животных (EFSA, 2012). Как одни из неинвазивных маркеров положительных эмоций животных предлагают определять количество окситоцина в моче (Mitsui et al., 2011). Однако, т.к. увеличение окситоцина в некоторых случаях может вызывать снижение дружелюбности и агрессию (Hernádi et al., 2015; Rault et al., 2017), то данный показатель выглядит более чем сомнительным. Разнонаправленные эффекты окситоцина на лисиц в зависимости от степени их агрессии (Гербек и др., неопубл. данные) также указывают на неприменимость этого показателя в пушном звероводстве.
За последние 20 лет на разных млекопитающих проведено множество исследований роли окситоцина во внутривидовых эмоционально-положительных контактах (Ross, Young, 2009; Bartz et al., 2011; Romero et al., 2014; Samuni et al., 2017). Ряд экспериментов проведённых на человеке также указывает, что введение ОТ (обычно в виде спрея в нос) стимулирует обмен взглядами, улучшает память на лица и потенцирует социальный эмоциональный ответ (Bartz et al., 2011).
Показано также, что эмоционально-положительные контакты собак между собой коррелируют с повышенным уровнем окситоцина в моче, и, обратно, введение окситоцина в нос коррелирует с увеличением эмоционально-положительных контактов. Кроме того, введение окситоцина повышало частоту и продолжительность игрового поведения у собак (Romero et al., 2014). Тем не менее, у поросят эффект введения окситоцина в нос зависел от эмоционального состояния – введение окситоцина после стрессорного воздействия повышал число эмоционально-положительных контактов с конспецификами, а введение после позитивного воздействия – снижал (Camerlink et al., 2016). Как уже упоминалось,повышенный уровень окситоцина в крови тёлок связан как с эмоционально-положительными, так и с агрессивными взаимодействиями (Yayou et al., 2015).
Доместицированные животные проявляют эмоционально-положительное поведение не только по отношению к конспецификам, но и переносят его на человекa. Верно и обратное в отношении человека и доместицированных животных. В течение последнего десятилетия активно проводятся исследования, которые направлены установить связь активности центральной ОТ системы современной собаки и других доместицированных видов и её взаимоотношениями с человеком. При этом исследуется и ответ ОТ системы человека.
В ряде работ показано, что контакт собаки с человеком в виде взаимных прикосновений, голосовых сигналов или даже только обменов взглядом повышает периферический окситоцин как у собаки, так и у человека (Odendaal, Meintjes, 2003; Rehn et al., 2014; Nagasawa et al., 2015). У ягнят концентрация ОТ в крови повышется как при сосании молока матери, так и при контакте с человеком. Более того, присутствие человека вызывает у ягнят повышение активности OT нейронов и ряда структур мозга, ими иннервируемых и участвующих в возникновении эмоций и социальном распознавании (Nowak, Boivin, 2015).
У свиней в спиномозговой жидкости также повышается концентрация окситоцина после эмоционально-положительного контакта с человеком (Rault et al., 2017). Это позволяет говорить о важной роли окситоцина не только во внутривидовых связях лабораторных животных, собак, человекообразных обезьян и человека, но и при контактах доместицированных животных с человеком.
Недавние эксперименты показали, что введение окситоцина в нос собаке повышает не только эмоционально-положительные контакты между собаками, но и частоту контактов с хозяином. При этом у хозяина повышалась концентрация ОТ в моче (Romero et al., 2014; Nagasawa et al., 2015). Обмен взглядами является важным социальным сигналом, которым пользуются собаки при эмоционально-положительных контактах с человеком. Собаки динго, подобно домашней собаке, способны инициировать обмен взглядами, хотя, вероятно, реже и на менее продолжительный период (Johnston et al., 2017). В то же время прирученные волки, по-видимому, не способны инициировать этот коммуникативный сигнал в отношении человека (Miklósi et al., 2003; Nagasawa et al., 2015).
Следует также отметить, что эффекты окситоцина на собак являются зависимыми от генотипа (пород) и, в частности, от полиморфизма рецептора окситоцина (Kovács et al., 2016; Kis et al., 2017).К сожалению, на данный момент почти нет достоверных данных о различии ОТ системы у волков и собак и, тем более, у других доместицированных животных и их сородичей, которые бы указывали на её роль в доместикации, ни на физиологическом, ни на молекулярном уровнях. Тем не менее, есть данные, полученных в экспериментах на доместицированных животных, которые, несмотря на их противоречивость, наводят на мысль о ключевой роли центральной окситоциновой системы в доместикации. С другой стороны, окситоциновая система хорошо известна своими видоспецифическими эффектами (Young, 1999). Это следует учитывать при сравнении результатов экспериментов на таких животных как, например, собака, лисица и свинья.
Наиболее перспективным представляется нейробиологическое сравнение ОТ системы в мозге доместицированных и диких животных, а также изучение эффекта введения окситоцина на агрессию и социальное поведение доместицированных животных и, в особенности, их диких сородичей. Генетические исследования практически не дают указаний на различия в генах окситоцина (von Holdt et al., 2010; Fam et al., 2018) и его рецептора между доместицированными и дикими животными, что однако, может быть связано с регуляцией их активности на других уровнях.
Обсуждая данные, полученные за последние годы при изучении ОТ, следует признать ряд ограничений, которые следует учитывать при их интерпретации (Rault et al., 2017). Во-первых, эффект ОТ как у животных, так и у человека индивидуален и зависит от многих факторов, таких как базальный уровень окситоцина (Bartz et al., 2011; Romero et al., 2014) и пол (Nagasawa et al., 2015; Oliva et al., 2015). Во-вторых, ОТ может иметь просоциальный эффект, направленный на знакомых индивидуумов и антисоциальный – на незнакомых (Shamay-Tsoory et al., 2009, de Dreu et al., 2011).
Исходя из приведённых данных можно предположить, что взаимоотношения человека и животных в период одомашнивания могли строится на отборе по усилению центральной окситоциновой системы, что могло снижать агрессию и стресс-ответ к человеку на начальных этапах доместикации. На следующем этапе такой отбор мог играть существенную роль в установлении особых отношений с человеком, называемых доместикационным поведением.
Использованная литература:
1. Беляев Д.К. Дестабилизирующий отбор как фактор доместикации // Генетика и благосостояние человечества. – М., 1981. – С. 53-66.
2. Беспятых О.Ю. Реакция лисиц разных генотипов на введение per os антиоксиданта янтарной кислоты // Информ. вестник ВОГиС. 2009. Т. 13. No 3. – С. 639-646.
3. Коппингер Л., Коппингер Р. Собаки. Новый взгляд на происхождение, поведение и эволюцию собак – М.: Софион, 2005. – 384 с.
4. Науменко Е.В., Попова Н.К., Иванова Л.Н. Нейроэндокринные и нейрохимические механизмы доместикации животных // Генетика. 1987. Т. 23. No 6. – С. 1011-1025.
5. Плотников И.А. Воспроизводительная способность лисиц (Vulpes Vulpes Linnaeus, 1758) разных типов поведения // Кролиководство и звероводство. 2017. No 3. – С. 69-70.
6. Трут Л.Н., Плюснина И.З., Оськина И.Н. Эксперимент по доместикации лисиц и дискуссионные вопросы эволюции собак // Генетика. 2004. Т. 40. No 6. – С. 794-807.
7. Anacker A.M.J., Christensen J.D., LaFlamme E.M., Grunberg D.M., Beery A.K. Septal oxytocin administration impairs peer affiliation via V1a receptors in female meadow voles // Psychoneuroendocrinology. 2016. V. 68. – P. 156-162.
8. Bartz J.A., Zaki J., Bolger N., Ochsner K.N. Social effects of oxytocin in humans: context and person matter // Trends Cogn. Sci. 2011. V. 15. – P. 301-309.
9. Calcagnoli F., Meyer N., de Boer S.F., Althaus M., Koolhaas J.M. Chronic enhancement of brain oxytocin levels causes enduring anti-aggressive and pro-social explorative behavioral effects in male rats // Horm. Behav. 2014. V. 65. – P. 427-433.
10. Camerlink I., Reimert I., Bolhuis J.E. Intranasal oxytocin administration in relationship to social behaviour in domestic pigs // Physiol. Behav. 2016. V. 163 – P. 51-55.
11. de Dreu C.K.W., Greer L.L., Kleef G.A.V., Shalvi S., Handgraaf M.J. Oxytocin promotes human ethnocentrism // PNAS. 2011. V. 108. – P. 1262-1266.
12. Ericsson M., Jensen P. Domestication and ontogeny effects on the stress response in young chickens (Gallus gallus) // Sci. Rep. 2016. V. 6. – P. 35818.
13. Fam B.S.O., Paré P., Felkl A.B. et al. Oxytocin and arginine vasopressin systems in the domestication process // Genet. Mol. Biol. 2018. V. 41. – P. 235-242.
14. Freedman A.H., Gronau I., Schweizer R.M. et al. Genome Sequencing Highlights the Dynamic Early History of Dogs // PLoS Genet. 2014 V. 10. – e1004016.
15. Grundwald N.J., Benítez D.P., Brunton P.J. Sex-Dependent Effects of Prenatal Stress on Social Memory in Rats: A Role for Differential Expression of Central Vasopressin-1a Receptors. J. of Neuroendocrinol. 2014. V. 28. No 4. – 14p.
16. Guagnin M., Perri A.R., Petraglia M.D. Pre-Neolithic evidence for dog-assisted hunting strategies in Arabia // J. Anthropol. Archaeol. 2018. V. 49. – P. 225-236.
17. Hernádi A., Kis A., Kanizsár O. et al. Intranasally administered oxytocin affects how dogs (Canis familiaris) react to the threatening approach of their owner and an unfamiliar experimenter // Behav. Processes. 2015. V. 119 – P. 1-5.
18. Johnston A.M., Turrin C., Watson L., Arre A.M., Santos L.R. Uncovering the origins of dog–human eye contact: dingoes establish eye contact more than wolves, but less than dogs // Anim. Behav. 2017. V. 133. – P. 123-129.
19. Jurek B., Neumann I.D. The Oxytocin Receptor: From Intracellular Signaling to Behavior // Physiol. Reviews. 2018. V. 98. No 3. – P. 1805-1908.
20. Kis A., Ciobica A., Topál J. The effect of oxytocin on human-directed social behaviour in dogs (Canis familiaris) // Horm. Behav. 2017. V. 94. – P. 40-52.
21. Koshimizu T., Nakamura K., Egashira N. et al. Vasopressin V1a and V1b receptors: From molecules to physiological systems // Physiol. Reviews. 2012. V. 92. No 4. – P. 1813-1864.
22. Kovács K., Kis A., Pogány Á., Koller D., Topál J. Differential effects of oxytocin on social sensitivity in two distinct breeds of dogs (Canis familiaris) // Psychoneuroendocrinol. 2016. V. 74. – P. 212–220.
23. Künzl C., Sachser N. The behavioral endocrinology of domestication: A comparison between the domestic guinea pig (Cavia aperea f. porcellus) and its wild ancestor, the cavy (Cavia aperea) // Horm. Behav. 1999. V. 35. – P. 28-37.
24. Larson G, Piperno DR, Allaby RG et al. Current perspectives and the future of domestication studies. PNAS. 111(17):6139–46.
25. Leng G., Ludwig M. Intranasal Oxytocin: Myths and Delusions // Biol. Psychiatry. 2016. P. 79. No 3. – P. 243-250.
26. Miklósi Á., Kubinyi E., Topál J. et al. A Simple Reason for a Big Difference: Wolves Do Not Look Back at Humans, but Dogs Do // Curr. Biol. 2003. V. 13. – P. 763-766.
27. Mitsui S., Yamamoto M., Nagasawa M. et al. Urinary oxytocin as a noninvasive biomarker of positive emotion in dogs // Horm. Behav. 2011. V. 60. – P. 239-243.
28. Nagasawa M., Mitsui S., En S. et al. Social evolution. Oxytocin-gaze positive loop and the coevolution of human-dog bonds // Science. 2015. V. 348. – P. 333-336.
29. Neumann I.D. Brain Oxytocin: A Key Regulator of Emotional and Social Behaviours in Both Females and Males // J. Neuroendocrinol. 2008. V. 20. – P. 858-865.
30. Neumann I.D., Landgraf R. Balance of brain oxytocin and vasopressin: implications for anxiety, depression, and social behaviors // Trends Neurosci. 2012. V. 35. – P. 649-659.
31. Nowak R., Boivin X. Filial attachment in sheep: Similarities and differences between ewelamb and human-lamb relationships // Appl. Anim. Behav. Sci. 2015. V. 164. – P. 12-28.
32. Odendaal J.S.J., Meintjes R.A. Neurophysiological correlates of affiliative behaviour between humans and dogs // Vet. J. 2003. V. 165. – P. 296-301.
33. Oliva J.L., Rault J.-L., Appleton B., Lill A. Oxytocin enhances the appropriate use of human social cues by the domestic dog (Canis familiaris) in an object choice task // Anim. Cogn. 2015. V. 18. – P. 767-775.
34. Price E.O. Animal Domestication and Behavior. – N.Y.: 2002. – 320 p.
35. Rault J.-L., Carter C.S., Garner J.P. et al. Repeated intranasal oxytocin administration in early life dysregulates the HPA axis and alters social behavior // Physiol. Behav. 2013. V. 112–113. – P. 40–48.
36. Rault J.-L., van den Munkhof M., Buisman-Pijlman F.T.A. Oxytocin as an Indicator of Psychological and Social Well-Being in Domesticated Animals: A Critical Review // Front Psychol. 2017. V. 8. – P. 1521.
37. Rehn T., Handlin L., Uvnäs-Moberg K., Keeling L.J. Dogs’ endocrine and behavioural responses at reunion are affected by how the human initiates contact // Physiol. Behav. 2014. V. 124. – P. 45-53.
38. Romero T., Nagasawa M., Mogi K., Hasegawa T., Kikusui T. Oxytocin promotes social bonding in dogs // PNAS. 2014. V. 111. – P. 9085-9090.
39. Ross H.E., Young L.J. Oxytocin and the neural mechanisms regulating social cognition and affiliative behavior // Front. Neuroendocrinol. 2009. V. 30. – P. 534-547.
40. Samuni L., Preis A., Mundry R. et al. Oxytocin reactivity during intergroup conflict in wild chimpanzees // PNAS. 2017. V. 114 – P. 268-273.
41. Shamay-Tsoory S.G., Fischer M., Dvash J. et al. Intranasal Administration of Oxytocin Increases Envy and Schadenfreude (Gloating) // Biol. Psychiatry. 2009. V. 66. – P. 864-870.
42. Suzuki K., Ikebuchi. M, Bischof H.-J., Okanoya K. Behavioral and neural trade-offs between song complexity and stress reaction in a wild and a domesticated finch strain //Neurosci. Biobehav. Rev. 2014. V. 46. – P. 547-556.
43. VonHoldt B.M., Pollinger J.P., Lohmueller K.E. et al. Genome-wide SNP and haplotype analyses reveal a rich history underlying dog domestication // Nature. 2010. V. 464. – P. 898-902.
44. Weiler U., Claus R., Schnoebelen-Combes S., Louveau I. Influence of age and genotype on endocrine parameters and growth performance: a comparative study in Wild boars, Meishan and Large White boars // Livest. Prod. Sci. 1998. V. 54. – P. 21-31.
45. Westwood R.E. Early fur-farming in Utah // Utah Historical Quarterly. 1989. V. 57. – P. 320-339.
46. Winslow J., Insel T.R. Vasopressin modulates male squirrel monkeys’ behavior during social separation // Eur. J. Pharmacol. 1991. V. 200. – P. 95-101.
47. Yayou K., Ito S., Yamamoto N. Relationships between postnatal plasma oxytocin concentrations and social behaviors in cattle // Anim. Sci. J. 2015. V. 86. – P. 806-813.
48. Young L.J. Frank A. Beach Award. Oxytocin and vasopressin receptors and species-typical social behaviors // Horm. Behav. 1999. V. 36. – P. 212-221.
Кролиководство и звероводство. 2019. №3.
Окситоцин: первичный механизм доместикации или её тормоз? Роль окситоцина при доместикации
Ю.Э. Гербек*, А.А. Малявко, Д.В. Шепелева, А.В. Владимирова
Резюме. В статье приведены результаты анализа поведения в парных тестах у взрослых самцов лисиц, селекционируемых на усиление агрессивно-оборонительной реакции по отношению к человеку («агрессивные» животные). Тестируемым животным в первые сутки после рождения внутрибрюшинно вводили окситоцин (ОТ) или физ.раствор. Оппонентами в парном тесте выступали лисицы, селекционируемые на усиление эмоционально-положительных реакций по отношению к человеку («ручные» животные), не подвергнутые предварительно никаким экспериментальным воздействиям. Показано, что введение ОТ агрессивным лисицам усиливает агрессивные реакции, направленные на ручных. На основании полученных результатов и анализа литературы, посвящённой экспериментам по введению окситоцина грызунам и доместицированным животным (в первую очередь, собакам), выдвинута гипотеза о том, что предки собак, дикие волки, должны были иметь пониженный уровень активности центральной ОТ–Системы в самый начальный этап доместикации, именуемые «протодоместикацией», вопреки высказываемым ранее предположениям. Следующие этапы доместикации могли, наоборот, сопровождаться повышением активности ОТ–Системы, что, вероятно, способствовало установлению эмоционально-положительного контакта с человеком.
На протяжении более сотни лет доместикация – крупнейшее культурно-эволюционное событие в истории человечества – находится в фокусе внимания исследователей. Так, Чарльз Дарвин описывал изменения, происходящие при доместикации еще в 1859 и 1868 гг. ХХ века [1, 2]. Важнейшим изменением, произошедшим при доместикации животных, являлось появление дружелюбия по отношению к человеку («ручного» поведения), которое не наблюдается в дикой природе. Несмотря на появление новых методов и возможностей исследования ответ на вопрос, как начиналась и происходила доместикация диких животных и что двигало её вперёд, по-прежнему далёк.
Первым одомашненным животным являлась собака, наиболее вероятным центром доместикации которой является Юго-Восточная Азия. Другие, хотя и менее вероятные центры происхождения могли располагаться на территории Ближнего Востока, Центральной Азии и Европы. Предполагаемое время расхождения волка и собаки также широко варьируется – от 11 до 30-40 тыс. лет до настоящего времени [3].
Доместикация собаки, вероятно, связана с тем, что некоторые из диких волков, предки собаки, могли подходить близко к стану человека, а не убегать, когда он появлялся. Это могло дать таким волкам преимущество в питании и размножении, т. к. не требовалось существенных затрат энергии на добычу пищи [4]. Этот этап перехода волка в синантропное состояние называют протодоместикации. Здесь начинается отбор на толерантное отношение к человеку. Он, по-видимому, был первым и важнейшим шагом в адаптивном поведении. При этом, как полагал Д.К. Беляев снижение стресса-ответа на человека, а возможно стресс-реакции вообще должно было способствовать этой адаптации [5].
Вероятно, сниженная активности гипоталамо-гипофизарно-надпочечниковой системы (ГГНС) является особенностью всех домашних животных по сравнению с их дикими сородичами [5]. На известной модели доместикации животных – серебристо-чёрной лисице, созданной многолетним отбором на эмоционально-положительное поведение по отношению к человеку, академик Д.К. Беляев и его сотрудники выявили снижение активности ГГНС во многих звеньях по сравнению со совхозными животными, а также лисицами, селекционируемыми в противоположную сторону, на усиление агрессивности по отношению к человеку [6]. Похожие закономерности показаны на параллельной экспериментальной модели доместикации, серой крысе [7] и при сравнении диких и исторически доместицированных морских свинок [8].
Экспериментальная доместикация лисиц уже не может рассматриваться как этап протодоместикации. Этот этап был пройден при переходе диких лисиц в содержание в неволе. Хотя лисицы по-прежнему оставались агрессивно-трусливыми в клетке и не подпускали человека ближе нескольких метров в вольере [3, 6, 9-11], однако они стали размножаться в неволе и к началу эксперимента по доместикации лисиц в 1958 году это не требовало стольких усилий и не приносило столько неудач как ещё в 1920-х гг. ХХ века [12-14].
Важнейшими вопросами стоящими перед исследователями, по-прежнему, являются следующие: как и с помощью каких механизмов появилась эмоционально-положительная реакция диких животных на человека на следующем шаге, на этапе собственно доместикации? Взаимоотношения человека и животных в период одомашнивания (особенно, собаки) могли строится на отборе по усилению активности центральной окситоциновой (ОТ) системы, что могло снижать агрессию и стресс-ответ к человеку на начальных этапах доместикации.
Известно, что она участвует в регуляции активности ГГНС [15] и связана не только с изменением уровня агрессии, но является важной в формировании дружелюбного, социального поведения у разных видов животных [16-17]. На грызунах в ряде экспериментов показано, что введение окситоцина в мозг взрослым животным или в первый день жизни внутрибрюшинно улучшает социальное распознавание, образование устойчивых пар или дружелюбное поведение [18]. Поэтому можно предположить, что предком собаки были волки с пониженной активностью ГГНС и повышенной активностью центральной ОТ–Системы. Это позволяло им питаться вблизи, а затем и в самом человеческом стане, не проявляя стрессорной реакции на человека и свободно размножаясь в этих условиях [19]. Для проверки этой гипотезы нами был проведён начальный эксперимент по введению ОТ внутрибрюшинно в первый день жизни лисицам, селекционируемым на усиление агрессивности по отношению к человеку.
Материалы и методы исследований
Внутрибрюшинные инъекции окситоцина новорожденным лисятам
В эксперименте использовали самцов серебристо-черных лисиц, селекционированных на усиление агрессивности по отношению к человеку («агрессивные»), более 50 лет содержащихся в условиях ЦКП «Генофонды пушных и сельскохозяйственных животных» ИЦиГ СО РАН. Периферическое введение окситоцина щенкам лисиц проводили в первые сутки после рождения. Были взяты щенки агрессивных лисиц, различимые по фенотипическим признакам: отличия в размере, форме и количестве белых пятен на груди, в паху, на пальцах и длине белого кончика хвоста – что позволило вводить ОТ или физ. раствор однопомётникам. Детеныши были ненадолго (~15 мин) отделены от матерей для введения 10 мкг/мл ОТ (Sigma-Aldrich), 1 мл раствора/100 г массы тела или физ.раствора фенотипирования, фото- и видеосъемки и взвешивания. Отрицательного эффекта от инъекций ОТ у щенков не наблюдалось, все они (n = 24) были приняты матерями.

Лисица с низким уровнем агрессивности. Из: Трут и др., 2017. Об отборе лисиц на агрессивность и его коррелированных последствиях. Вавиловский журнал генетики и селекции. Т.21. №4.

Высокоагрессивная лисица. Из: Трут и др., 2017.
Тестирование поведения
Экспериментальные агрессивные лисицы, которым был неонатально введён ОТ или физ. раствор были протестированы на поведение по отношению к другой лисице в возрасте 6–6,5 месяцев. В качестве оппонента использованы ручные лисицы с мутациями в окраске (в качестве цветового маркера при расшифровке видеозаписей). На 18 часов лисиц пересаживали в новую экспериментальную клетку для привыкания к новой территории. Клетка состоит из трёх последовательных отделений с непрозрачными перегородками, в 1-ом находилась экспериментальная лисица (агрессивная); 2 – нейтральная территория; 3 – лисица-оппонент (ручная). Перед началом теста, открывали входы в нейтральный отсек и проводили видеорегистрацию взаимодействия в течение 20 мин. С помощью программы «Реалтаймер» («Открытая наука», Россия) анализировали паттерн поведения на видеозаписях. Поведение относили к агрессивому, тревожному, дружелюбному (аффилиативному), двигательному и смешанному.
Статистический анализ
Статистический анализ проводили с помощью метода главных координат (PCo) [20] с использованием программного комплекса JACOBI 4 (Россия) для многомерного анализа биологических данных, который использует для вычисления главных компонент множества объектов матрицу евклидовых расстояний между ними, что снимает ограничения на количество анализируемых признаков у ограниченной выборки особей.
Результаты исследований и обсуждение
По вектору дисперсии вычисленных главных координат была определена целесообразность рассмотрения первых двух координат, чья дисперсия составила: PCo1 – 8,08, PCo2 – 6,94.
A. Поскольку распределение по PCo1 оказалось не связано с неонатально введённым окситоцином, а касалось внутригрупповой дисперсии, то для данного исследования целесообразно было рассматривать только распределение по PCo2, в которую достоверно даёт вклад фактор окситоцина (p<0,05). В PCo2 также дают достоверный вклад типы поведения обозначенные в таблице.
Наибольший вклад в PCo2 делают факторы с отрицательным значением, которые относятся к агрессивному или смешанному поведению. Данная компонента была интерпретирована как «неагрессивное поведение». Животные, в отрицательном множестве являются более агрессивными, чем в положительном (рис. А).
Агрессивные лисицы из контрольной группы, которым был введен физиологический раствор, образуют два чётко разделённых по второй компоненте множества – более агрессивные (вверху) и менее агрессивные (внизу) (рис. А). Лисицы, которым неонатально вводили окситоцин (10 мкг/мл) лежали в основном в нижней, отрицательной области, или приближаются к нулевым значениям (рис. Б). Таким образом, видно, что неонатальное введение окситоцина (10 мкг/мл) ассоциировано с агрессивным поведением по отношению к другим лисицам.
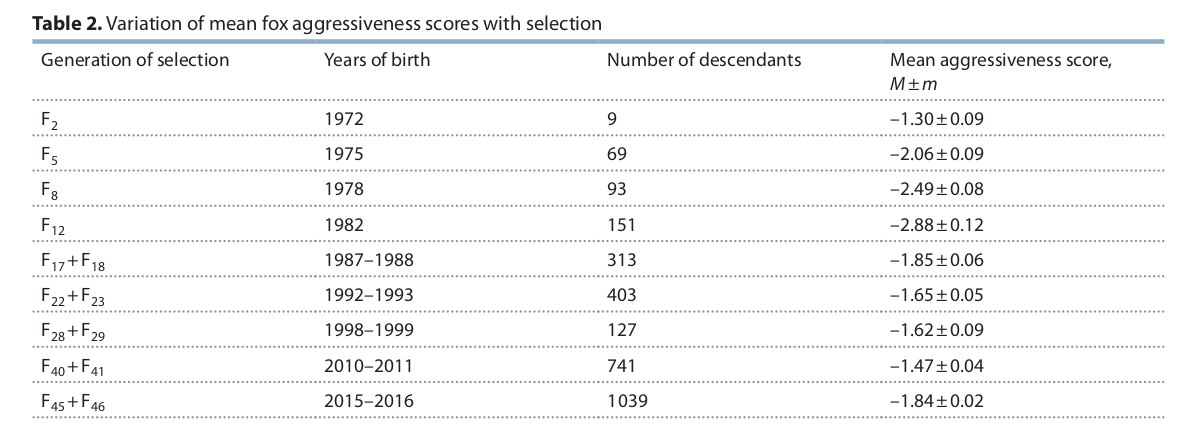
Изменения балла агрессивности в череде поколений у лисиц, селектируемых на рост агрессивности. Из: Трут и др., 2017
Полученные в данном исследовании результаты вписываются в парадигму, что введение окситоцина не всегда связано со снижением агрессивности, а в ряде случаев может вызывать её усиление.
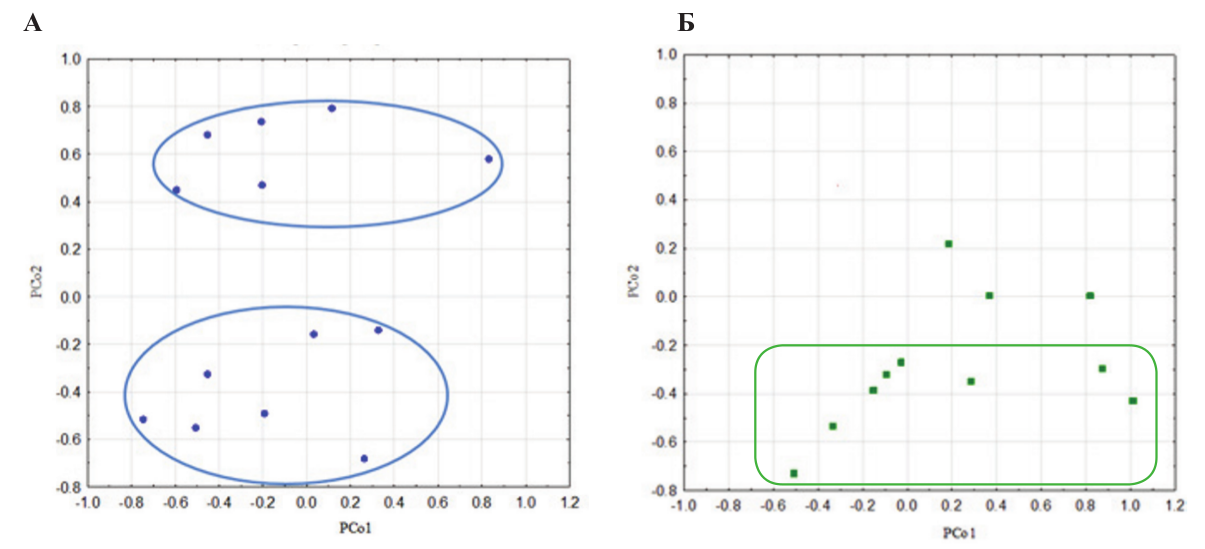
Рисунок А. Распределение лисиц по главным координатам PCo1 и PCo2. A – контрольная группа (физ.раствор), n = 13; Б – экспериментальная группа (окситоцин), n = 11.
Повышенная агрессия при центральном введении ОТ наблюдалась у доминантных самцов беличьих обезьян [21]. Кроме того, у доминантных крыс обнаружено повышенное количество рецептора ОТ в социально-значимых областях мозга [22]. При межгрупповых конфликтах у шимпанзе периферический уровень ОТ повышался [23]. Ручные серые крысы после пятидневного интраназального введения окситоцина становились более агрессивными по отношению к другим крысам [24].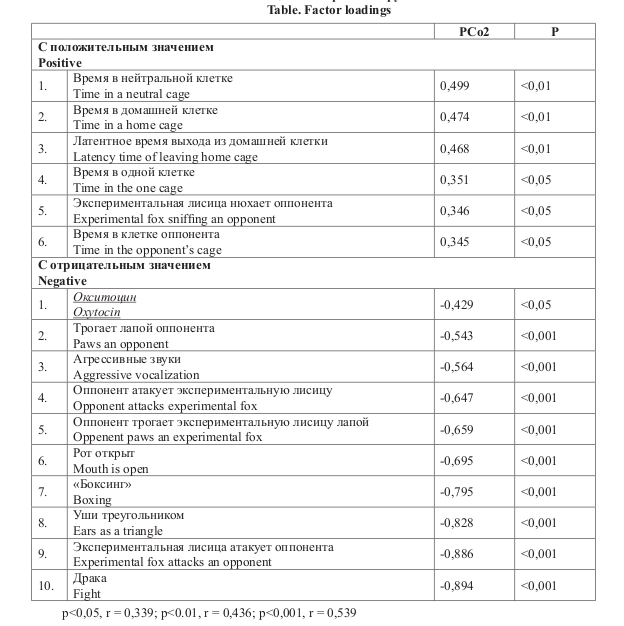 Важно, что похожие результаты были получены на доместицированных животных. После интраназального введения ОТ собака может вести себя менее дружелюбно по отношению к хозяину при его угрожающем поведении [25]. Введение ОТ поросятам интраназально в течение нескольких дней после рождения приводило к повышению в стрессирующих условиях агрессии, направленной на других поросят в возрасте 17 дней и 8 недель [26]. Эндогенный высокий уровень ОТ у тёлок был связан с повышенным количеством социальных взаимодействий в стаде: как дружелюбных, так и, наоборот, агрессивных [27].
Важно, что похожие результаты были получены на доместицированных животных. После интраназального введения ОТ собака может вести себя менее дружелюбно по отношению к хозяину при его угрожающем поведении [25]. Введение ОТ поросятам интраназально в течение нескольких дней после рождения приводило к повышению в стрессирующих условиях агрессии, направленной на других поросят в возрасте 17 дней и 8 недель [26]. Эндогенный высокий уровень ОТ у тёлок был связан с повышенным количеством социальных взаимодействий в стаде: как дружелюбных, так и, наоборот, агрессивных [27].
Приведённые исследования наводят на мысль, что в стрессогенных условиях высокий уровень ОТ вызывает повышение, а не понижение агрессии [28, 29]. Исходя из этих данных можно предположить, что в стрессогенных условиях присутствия человека центральная ОТ–Система предков собак при повышенной активности могла не понижать агрессивность, а, наоборот, её повышать. Исходя из моделей психофизиологического действия ОТ, основанных как на результатах, полученных на животных, так и на наблюдениях за человеком, ОТ, по-видимому, не является просто анксиолитическим и просоциальным фактором, но увеличивает значимость социальных сигналов [30-32].
Он способствует передаче информации о них в соответствующие области мозга, указывая, что происходящее событие может быть значимо для приспосабливаемости или с положительным, или с отрицательным знаком. Существует ряд исследований, в которых ОТ усиливает тревожность, а не ослабляет ее [30, 33]. Таким образом, в стрессогенных или неопределённых, но тревожных условиях, ОТ может усиливать оценку опасности и фокусировать на ней внимание. При отсутствии опасности, наоборот, ОТ ослабляет бдительность.
В данном эксперименте важно отметить, что оппонентом агрессивным всегда выступали ручные лисицы. Обнаружено (A.V. Kukekova, неопубликованные данные), что количество драк в смешенных парах ручных и агрессивных лисиц существенно больше, чем в парах агрессивных лисиц. Это, по-видимому, связано с повышенной социальностью ручных лисиц («навязчивое» игровое поведение). Возможно, уровень активности центральной ОТ системы, заданный ранним введением ОТ, способствует тому, что адаптивным ответом на дружелюбное (игровое) поведение в стрессогенной обстановке теста является агрессивный ответ. Кроме того, следует упомянуть о нелинейном дозозависимом эффекте введения окситоцина после рождения, обнаруженном в исследованиях на грызунах [18].
Эффект низкой дозы может быть ярче, чем большей, а может быть противоположным. Количества ОТ на грамм веса, вводимые в нашем эксперименте щенкам, были ниже, чем в предыдущих экспериментах на грызунах [18], что также могло повлиять на результат.
Заключение
Таким образом, исходя из представленных результатов и данных литературы, можно предположить, что на этапе ранней доместикации волк, предок собаки, который получил эволюционное преимущество, питаясь остатками добычи или на свалках рядом с человеком, скорее всего, имел пониженный уровень активности центральной ОТ–Системы, тревожности и агрессивности по сравнению с волками, которые боялись подойти к человеческому жилью из-за возможной опасности и добывали еду охотой. Однако на следующем этапе (собственно доместикации) проявление эмоционально-положительной реакции по отношению к человеку могло возникнуть при отборе на повышенную активность ОТ–Системы в благоприятных нестрессорных условиях.
Список литературы
1. Дарвин Ч. Происхождение видов – М.: Гос. изд-во сельхоз. лит., 1952. –С.192.
2. Дарвин Ч. Изменение животных и растений в домашнем состоянии. –М. –Л.: ОГИЗ –Сельхозгиз. 1941. –С.620.
3. Трут Л.Н., Харламова А.В., Пилипенко А.С., Гербек Ю.Э. Эксперимент по доместикации лисиц и эволюция собак с позиции современных молекулярно-генетических и археологических данных // Генетика. 2021. V. 57. No7. –C.67-785.
4. Коппингер Л., Коппингер Р. Собаки. Новый взгляд на происхождение, поведение и эволюцию собак. –М.: Софион, 2005 –С.400.
5. Беляев Д.К. Дестабилизирующий отбор как фактор доместикации // Генетика и благосостояние человечества. –М.: Наука, 1981. –С.53–66.
6. Трут Л.Н., Плюснина И.З., Оськина И.Н. Эксперимент по доместикации лисиц и дискуссионные вопросы эволюции собак // Генетика. 2004. Т. 40. No 6. –С.794-807.
7. Оськина И.Н., Гербек Ю.Э., Шихевич С.Г., Плюснина И.З., Гулевич Р.Г. Изменения гипоталамо-гипофизарно-надпочечниковой и иммунной систем при отборе животных на доместикационное поведение // Информ. Вестник ВОГиС. 2008. Т. 12. No 1-2. –С.39-49.
8. Künzl C., Sachser N. The behavioral endocrinology of domestication: a comparison between the domestic guinea pig (Cavia apereaf. porcellus) and its wild ancestor, the cavy (Cavia aperea) //Hormones and Behavior. 1999. V. 35. No 1. –P. 28-37.
9. Croft R.B. The culture of black and silver foxes // Rod and gun in Canada. 1912. No 8. (Цит. По: [11])
10. Keeler, C.E. Coat colour gene synthesis of tame behaviour in the rat, mink and fox // Mind Over Matter. 1964. No.9. –P. 16-30.
11. Генерозов В.Я. Промышленное разведение серебристо-черных лисиц и песцов в Северной Америке: Отчет по осмотру лисоводных питомников в Канаде. – Петроград: Типография М. Меркушева, 1916. – С.256.
12. Анфилов В.К. Разведение лисиц и лисьи фермы. 1916. Петроград: Изд-во Сойкина. – С.79.
13. Петряев П.А. Разведение серебристо-черных лисиц. Выпуск I. –1929. –М.: Изд-во «Госторга РСФСР». –С.112.
14. Афанасьев В.А., Перельдик Н.Ш. Клеточное пушное звероводство. –М.: Колос. –1966. –С.399.
15. Jurek B., Slattery D.A., Hiraoka Y., Liu Y., Nishimori K., Aguilera G., Neumann I.D., van den Burg E.H. Oxytocin regulates stress-induced Crf gene transcription through CREB-regulated transcription coactivator 3 // J. Neurosci. 2015. V. 35. – P. 12248–12260.
16. Гербек Ю.Э., Гулевич Р.Г., Шепелева Д.В., Гриневич В.В. Окситоцин: коэволюция человека и доместицированных животных // Вавиловский журнал генетики и селекции. 2016. V. 20. No 2. –С.220–227.
17. Jurek B., Neumann I.D. The oxytocin receptor: From intracellular signaling to behavior // Physiol. Rev. 2018. V. 98. No 3. P. 1805–1908.
18. Miller T.V., Caldwell H.K. Oxytocin during Development: Possible Organizational Effects on Behavior // Front. Endocrinol. 2015. V. 6. – 76.
19. Гербек Ю.Э. Новый взгляд на механизмы доместикации: окситоцин как маркер поведения // Кролиководство и звероводство. 2019. No 3. –С.45-49.
20. Gower J.C. Some distance properties of latent root and vector methods used in multivariate analysis // Biometrika. 1966. V. 53. No 3-4. –P. 325-338.
21. Winslow J.T., Insel T.R. Social status in pairs of male squirrel monkeys determines the behavioral response to central oxytocin administration // J. Neurosci. 1991. V. 11. No 7. –P. 2032-2038.
22. Lee W., Hiura L.C., Yang E., Broekman K.A., Ophir A.G., Curley J.P. Social status in mouse social hierarchies is associated with variation in oxytocin and vasopressin 1a receptor densities // Horm. Behav. 2019. V. 114. –P. 104551.
23. Samuni L., Preis A., Mundry R., Deschner T., Crockford C., Wittig R.M. Oxytocin reactivity during intergroup confl ict in wild chimpanzees // PNAS. 2017. V. 114. –P. 268-273.
24. Gulevich R., Kozhemyakina R., Shikhevich S., Konoshenko M., Herbeck Y. Aggressive behavior and stress response after oxytocin administration in male Norway rats selected for different attitudes to humans // Physiol. Behavior. 2019. V. 199. –P 210-218.
25. Hernádi A., Kis A., Kanizsár O., Tóth K., Miklósi B., Topál J. Intranasally administered oxytocin affects how dogs (Canis familiaris) react to the threatening approach of their owner and an unfamiliar experimenter // Behav. Processes. 2015. V. 119. –P. 1-5.
26. Rault J. -L., Carter C.S., Garner J.P., MarchantForde J.N., Richert B.T., Lay D.C. Repeated intranasal oxytocin administration in early life dysregulates the HPA axis and alters social behavior // Physiol. Behav. V. 112–113 –P. 40–48.
27. Yayou K., Ito S., Yamamoto N. Relationships between postnatal plasma oxytocin concentrations and social behaviors in cattle // Anim. Sci. J.V. 86. –P.806-813.
28. Herbeck Y.E., Gulevich R.G. Neuropeptides as facilitators of domestication // Cell Tissue Res. 2019a. V. 375. No 1. –P. 295-307.
29. Herbeck Y.E., Gulevich R.G. Correction to: Neuropeptides as facilitators of domestication // Cell Tissue Res. 2019b. V. 375. No 1. –P. 309-310.
30. Crespi B.J. Oxytocin, testosterone, and human social cognition // Biol. Rev. 2016. V. 91. No 2. –P. 390-408.
31. Ne’eman R., Perach-Barzilay N., Fischer-Shofty M., Atias A., Shamay-Tsoory S.G. Intranasal administration of oxytocin increases human aggressive behavior // Horm. Behav. 2016. V. 80. –P. 125-131
32. Egito J.H., Nevat M., Shamay-Tsoory S.G., Osório A.A.C. Oxytocin increases the social salience of the outgroup in potential threat contexts // Horm. Behav. 2020. V. 122. –P. 104733.
33. Eckstein M., Scheele D., Weber K., Stoffel
Wagner B., Maier W., Hurlemann R. Oxytocin facilitates the sensation of social stress // Human Brain Map. 2014. V. 35 –4741-4750.
Кролиководство и звероводство. 2021. №5.
Рекомендуем прочесть
 Эксперимент по доместикации лисиц и эволюция собак с позиции современных молекулярно-генетических и археологических данных
Эксперимент по доместикации лисиц и эволюция собак с позиции современных молекулярно-генетических и археологических данных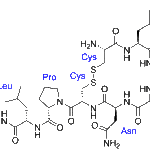 Окситоцин: коэволюция человека и доместицированных животных
Окситоцин: коэволюция человека и доместицированных животных «Самодоместикация»: модель, которая не везде работает
«Самодоместикация»: модель, которая не везде работает Как доместикация усложняет песни бенгальской амадины
Как доместикация усложняет песни бенгальской амадины Клеточные и молекулярные механизмы наркотической зависимости
Клеточные и молекулярные механизмы наркотической зависимости Выбор альтернатив в популяризации науки
Выбор альтернатив в популяризации науки Последнее слово о фетальном тестостероне
Последнее слово о фетальном тестостероне Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов
Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов
