

В.А. Красилов
Палеонтологический институт РАН, Москва E-mail: vkrassilov@hotmail.com
Для парадигм современного естествознания характерны упрощенные решения сложных проблем. При этом значительные массивы научной информации оказываются излишними. В утверждении таких парадигм немаловажную роль играют вненаучные — политические, коммерческие — факторы. Примерами могут служить молекулярная филогения, тектоника плит, парниковая модель климатических изменений и импактная модель массовых вымираний. В основе этих моделей лежат допущения, умозрительные механизмы и неправомерные обобщения (молекулярные часы, конвекционный механизм движения литосферных плит, потепление последних десятилетий как доказательство парникового разогрева атмосферы, иридиевая аномалия как свидетельство вымирания динозавров от столкновения Земли с астероидом). Такие теории не только дискредитируют теоретические исследования, но и наносят ущерб связанным с ними областям практической деятельности. В качестве более перспективных рассматриваются альтернативные теории эпигенетических процессов как основы филогенеза, ротационной тектоники, перераспределения тепла по поверхности Земли, изменения структуры биологического разнообразия как причины вымирания. В обосновании этих теорий ведущую роль играют палеонтологические исследования.
Юбилей палеонтолога и организатора палеонтологических исследований — подходящий повод для разговора о ситуации в палеонтологии, которую едва ли удастся прояснить безотносительно к ситуации в естественных науках в целом. Известный философ Томас Кун (1975) утверждал, что наука развивается от одной парадигмы к другой. Смену парадигм он описывал как сложный процесс, включающий накопление данных, не укладывающихся в рамки общепринятых представлений, исчерпанность исследовательских программ, разочарование в авторитетах, смену поколений и многое другое.
Люди моего поколения, с увлечением читавшие Куна тридцать лет назад, теперь с удивлением наблюдают за тем, как парадигмы раздуваются и лопаются на глазах подобно мыльным пузырям. Раньше препятствием на пути научного прогресса был консерватизм ученых, с большим недоверием относившихся к новым идеям. Теперь опасения вызывает скорее та легкость, с которой огромные массивы накопленных знаний предаются забвению, не будучи востребованы современными парадигмами. Парадигма из инструмента мобилизации накопленных знаний превращается в инструмент элиминации. Это новая ситуация, и мы пока не знаем, во что она разовьется.
Симптоматично, что в самых престижных международных журналах «Nature» и «Science» первичный отбор статей производят не ученые, а журналисты, лучше знающие, что интересно массовому читателю. Именно эти журналы сыграли решающую роль в стремительном утверждении таких парадигм, как тектоника плит, парниковая теория изменения климата, катастрофические столкновения Земли с астероидами, определяющие ход геологической истории, и тому подобное. Вообще деление научных журналов на престижные и непрестижные выбило почву из-под ног любителей отстаивать, как Грегор Мендель, свое особое мнение в каких-нибудь «Записках естествоиспытателей Брно». Если ваше мнение чего-то стоит, почему вас не публикует «Science»? Записки естествоиспытателей не принесут вам грантов, вы не приобретете современного оборудования, а монастырей, в которых можно было бы бесплатно ставить опыты по гибридизации растений, уже практически не осталось.
Рыночные механизмы гораздо более эффективно обеспечивают единомыслие, чем репрессирование ученых. Но может быть это и хорошо, что бесконечные споры сменились завидным единодушием, а научные идеи стали понятны массам. К сожалению, эта ситуация требует простых и однозначных решений там, где они в принципе невозможны в силу сложности и неоднозначности решаемых проблем. Упростить проблему легче всего, рассматривая ее с высоты птичьего полета, переводя ее на виртуальный уровень, где никакие факты вообще не нужны, а ведь палеонтолог склонен рыться в земле и выкапывать новые проблемы.
Далее будут кратко рассмотрены современные парадигматические модели, более подробный анализ которых и обсуждение альтернатив можно найти в других работах автора. Общее впечатление таково, что в современном естествознании можно обойтись без классических дисциплин, роль которых в формировании новых парадигм неуклонно уменьшается. Так, эволюция органического мира сейчас в основном реконструируется на основе молекулярной филогении, развитие земной коры уже объяснили геофизики, а климатические изменения, как известно, связаны с выбросами СО2. Нужна ли в таком случае палеонтология?
Молекулярная филогения
Содержание
Раньше считали, что поскольку палеонтолог располагает сведениями о хронологической последовательности событий, о том, как все происходило на самом деле, то ему и заниматься филогенетическими построениями, привлекая данные других дисциплин. Но в наши дни эта область исследований практически монополизирована молекулярной филогенией. Палеонтологу, чтобы не отстать от времени, приходится признать, что молекулярные часы точнее, чем палеонтологическая летопись, что молекулярная филогения во всех отношениях превосходит традиционную и что должным образом отпрепарированные палеонтологические данные все это подтверждают. Правда, спрос на такого рода подтверждения неуклонно сокращается.
Методы молекулярно-филогенетических построений непрерывно совершенствуются, и их программное обеспечение становится все более сложным. Тем не менее сохраняется ряд исходных установок, вносящих в эти построения неустранимый элемент условности. Так, постоянная скорость мутирования (молекулярные часы) — это не более чем допущение, частично, может быть, оправданное в случае нефункциональных участков, таких как вкрапленные в ядерный геном фрагменты митохондриальной ДНК, но в общем случае не имеющее ни теоретических, ни экспериментальных оснований. Монофилия не вытекает из модели, а заложена в ней и не имеет никаких предпосылок, кроме мифа об Адаме и Еве (филогения человеческого рода сейчас как раз и сводится к реконструкции молекулярной Евы). Негласно принимается необратимость развития, хотя все элементарные мутационные акты обратимы.
В любом случае молекулярная филогения отражает эволюционную историю одного или немногих элементов генома. Как она соотносится с филогенией организмов — это, пожалуй, самый сложный вопрос эволюционной теории, решение которого лежит за пределами молекулярно-филогенетических построений и связано с противоречивыми представлениями о роли элементарных генетических и системных эпигенетических событий. Вопреки хрестоматийным представлениям об отборе единичных мутаций как основном механизме эволюции, эволюционно значимые изменения происходят на уровне генома как системы. Данные сравнительной морфологии и палеонтологии недвусмысленно свидетельствуют о системном (макромутационном) изменении функционирования генома как первом шаге на пути эволюционных преобразований, за которым следует микромутационная доводка отдельных генов в новых условиях функционирования. Современные данные молекулярной генетики подтверждают значение эпигенетических механизмов передачи наследственной информации. Но если Чарлзу Дарвину хватило мужества на склоне лет признать недооценку таких механизмов своей величайшей ошибкой, то мужества современных теоретиков эволюционизма обычно хватает лишь на то, чтобы слегка подновить в прошлом отвергнутую ими же теорию и выдать за свою.
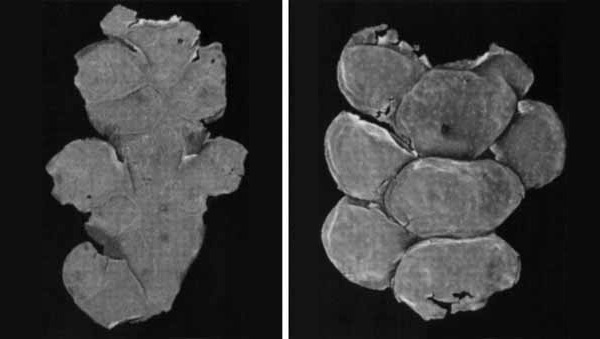
Рис. 1. Полиморфизм переходных форм от чешуелистных побегов к филлокладию у голосеменного растения из пермских отложений Европейской России. Изображение «Экология и жизнь»
В основании расходящихся филогенетических линий обычно находятся макрополиморфные популяции, пределы морфологической изменчивости которых нередко превышают морфологические дистанции не только видового, но и родового или даже семейственного уровней. Пример приведен на рис. 1: преобразование трехмерных чешуелистных побегов в дорсовентральные филлокладии у макрополиморфного вида пермских голосеменных дает спектр переходных форм между хвойными и пельтаспермовыми. На рис. 2 представлен пример макромутационного возникновения мезоперигенного устьичного аппарата с большим числом побочных клеток в стволовой линии пермских хвойных. Мезозойские формы как с перигенными, так и с мезогенными устьичными аппаратами могли возникнуть путем обособления соответствующих вариантов из макрополиморфной предковой популяции. Эти примеры показывают, что дивергенция не единственный способ формирования филогенетического древа.
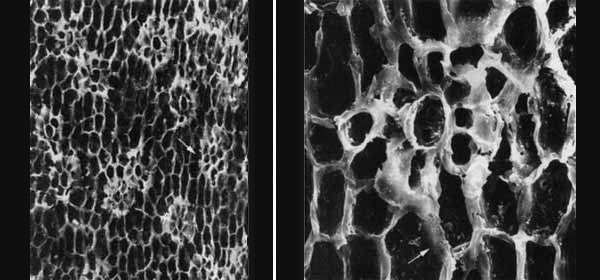
Рис. 2. Полиморфизм устьичных аппаратов у пермского хвойного Ullmannia cf. bronnii Goeppert. Абортированный устьичный аппарат (стрелка) в виде скопления мелких мезогенных клеток, образующих в зрелом мезоперигенном аппарате (справа) кольцо побочных клеток, прерванное вклинивающейся перигенной клеткой. Изображение «Экология и жизнь»
Противоречия между палеонтологической и молекулярной филогенией отчасти обусловлены тем, что в генетике системное представление о геноме еще не сформировалось. Однако именно генетике сейчас принадлежит монополия на теоретизирование, вследствие чего она не получает стимула для пересмотра своих базовых установок от других биологических дисциплин, добровольно сложивших с себя функцию генерирования продуктивных гипотез.
Тектоника плит
Аналогичная ситуация сложилась и в науках о Земле, теоретические разработки в которых монополизированы плит-тектонической парадигмой. Уже стало настолько привычным видеть в палеонтологических работах виртуальные карты смещенных континентов, построенные на основе плит-тектонической модели, что они воспринимаются как некая реальность. На ранней стадии становления этой парадигмы иногда вспоминали о необходимости фактологической проверки умозрительных построений, но теперь вопрос так не стоит: если палеобиогеография противоречит плит-тектоническим реконструкциям, то тем хуже для нее. Неудивительно, что палеонтологи в массе своей перешли на виртуальную географию, тем самым внося свою лепту в утверждение плит-тектонической парадигмы.
Тем не менее базовые установки биогеографии настолько фундаментальны, что не считаться с ними не может ни одна теория, претендующая на научность. Поступающая солнечная радиация убывает от экватора к полюсам, в связи с чем возникает широтная климатическая зональность, определяющая характер растительности. В ходе геологической истории глобальные фитоклиматические зоны сужались и расширялись, но при этом оставались параллельными широтам. Смещение на сфере означает вращение, поэтому на смещенном континенте линии координат и параллельные им биогеографические элементы должны быть расположены под углом к современным. Если же они параллельны современным, то смещения не было. Не все биогеографические элементы в равной мере изучены, но границы основных фитоклиматических зон подтверждены обширными и вполне убедительными данными. С девона по мел граница тропической и умеренной зон параллельна современным широтам (отклонения, скорее всего, связаны с крупносдвиговыми дислокациями, рис. 3), и ее положение в разные эпохи нередко совпадает.
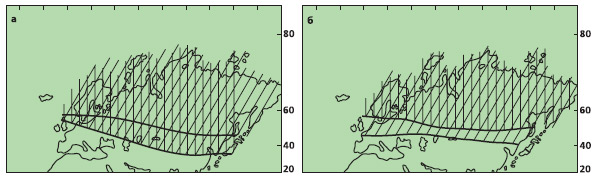
Рис. 3. Положение границы субтропической и умеренной (заштрихована) фитоклиматических областей в средней юре — раннем мелу (а) и позднем мелу — палеогене (б). Изображение «Экология и жизнь»
Мы, таким образом, имеем действенный инструмент для проверки плит-тектонических реконструкций, и они ее не выдерживают. Сейчас уже редко вспоминают о том, что в основе тектоники плит лежит рутинное геофизическое исследование *, подтвердившее известное с 20-х годов прошлого века деление литосферы на устойчивые кратоны («плиты»), окруженные подвижными поясами (сейсмичными зонами), дополненное облегченной теорией дрейфа континентов, из которой удален ротационный механизм. Собственно плит-тектонический механизм растаскивания плит конвекционными потоками и их субдукции — поддвига под другие плиты и погружения в мантию — с самого начала был и по сей день остается сугубо умозрительным.
Попперовский критерий опровергаемости тут не работает: субдукцию невозможно ни подтвердить, ни опровергнуть. Единственный пригодный для таких случаев инструмент — это старая, но все еще грозная бритва Оккама. От лишних умозрений (а именно такова субдукция) надо решительно избавляться.
Хотя плитовая тектоника ничего не объясняет, не предсказывает и уже длительное время не развивается, что указывает на ее исчерпанность, она успешно подавляет развитие альтернативных моделей, в частности, ротационной, в практическом плане необходимой для предсказания землетрясений. Сильные повторяющиеся землетрясения связаны с активизацией элементов планетарной сети разломов, образовавшихся в результате ротационной деформации Земли. Теоретические исследования в этом плане с появлением плитовой тектоники были практически прекращены. Плитовая тектоника, претендуя на совершенно новую модель развития Земли, никак не могла принять ротационный механизм: ведь на него опиралась старая теория перемещения континентов, разработанная А. Вегенером, и тогда что же нового предложено плитовой моделью? Неуязвимость плитовой тектоники для конструктивной критики объясняется ее включением в систему национальных приоритетов, а это уже не наука, а политика.
Парниковая модель климатических изменений
Еще в большей мере обязана своим успехом политике парниковая парадигма изменения климата, используемая для поддержки ядерной энергетики (не дающей выбросов парниковых газов в атмосферу), от которой прямой путь к ядерному оружию массового уничтожения. Физическая основа теории кажется безупречной — парниковые газы разогревают атмосферу, это общеизвестно. Но когда речь идет о сложных процессах, ничто так не вводит в заблуждение, как общеизвестное. В одномерной парниковой модели не учтены обратные связи в системе «парниковые газы — температура» (рис. 4).

Рис. 4. Схема регуляции содержания атмосферной СО2. Изображение «Экология и жизнь»
Поскольку растворимость углекислого газа в воде и его участие в реакциях с силикатными соединениями зависят от температуры, то именно колебания температуры являются ведущим компонентом системы. Иными словами, повышение температуры по причинам, не имеющим ничего общего с парниковым эффектом, вызывает кратковременное (поскольку существуют мощные буферные механизмы) увеличение содержания парниковых газов в атмосфере. Поэтому колебания СО2 в атмосфере всегда сопровождают как второстепенный эффект температурные циклы, связанные с солнечной активностью, динамикой вращения Земли и другими периодическими процессами.
Одним из показателей концентрации СО2 в атмосфере является устьичный индекс — отношение числа устьиц к числу эпидермальных клеток. В связи с парниковой моделью был выполнен ряд исследований по изменению устьичного индекса на листьях ископаемых растений из различных систематических групп. Ни в одном случае не обнаружено таких колебаний устьичного индекса, которые соответствовали бы масштабу климатических изменений, засвидетельствованных геологической летописью, и, таким образом, ведущая роль колебаний СО2 в эволюции климата не подтверждается.
Предполагаемая угроза потепления (таяние полярных льдов и поднятие уровня моря) также не учитывает механизм обратных связей. Таяние полярных льдов происходит в начале температурного цикла. С дальнейшим повышением температуры увеличиваются конвективность атмосферы и количество осадков в виде снега, способствующих росту полярных ледяных шапок.
Если же обратиться к палеонтологической летописи, то она недвусмысленно свидетельствует о том, что существенные климатические изменения происходили вследствие перераспределения тепла по земной поверхности, а не разогрева атмосферы в целом. Так, по палеоботаническим данным, в позднемеловую эпоху высокоширотный климат был теплее, а тропический холоднее, чем сейчас, причем в районах, которые, как на географической карте, так и на виртуальных плит-тектонических реконструкциях, попадают в тропики или субтропики, и растительность ровных водораздельных пространств имела более умеренный облик, чем в наши дни.
Международная борьба с техногенными выбросами СО2, инспирированная одномерной парниковой моделью климатических изменений, едва ли может существенно повлиять на направленность климатических процессов, но отвлекает значительные средства от предотвращения реальных угроз, таких как наводнения, землетрясения, цунами, частота которых возрастает в связи с глобальной тектонической активизацией. К тому же широко распропагандированная теория техногенного потепления приводит к тому, что люди оказываются морально и психологически неподготовленными к холодным зимам, неотвратимо приходящим на смену относительному потеплению. Конечно, механизмы изменения глобального климата еще во многом неясны, их изучение должно быть основано на объективном анализе всей доступной информации, в том числе палеонтологической. Здесь уместны любые модели, кроме одномерных.
Импактная модель массовых вымираний
Особенно впечатляет стремительное превращение в парадигму гипотезы массового вымирания вследствие столкновения Земли с крупным небесным телом, ведь авторитетные теоретики эволюционизма долгое время категорически отрицали теорию катастроф, восходящую к Ж. Кювье. Правда, в прошлом не было такого единомыслия, как сейчас, и катастрофические гипотезы время от времени возникали на обочине эволюционизма, неизменно подвергаясь резкой критике. Не избежал упреков в приверженности катастрофизму и автор этих строк, писавший о периодичности эволюционных процессов, вызванных внешними воздействиями на экосистемы.** По этим представлениям, длительные дестабилизирующие воздействия изменяют адаптивную стратегию видов, в устойчивых условиях направленную на разделение экологических ниш, а под влиянием стрессовых факторов приобретающую противоположную направленность. Экологическая экспансия приводит к перекрытию ниш и в конечном счете к сокращению биологического разнообразия. Наиболее конкурентоспособны в стрессовых условиях быстро размножающиеся виды, способные противостоять высокому уровню смертности. Популяциям таких видов свойственны резкие колебания численности, которые дестабилизируют трофическую структуру экосистемы и служат дополнительным фактором сокращения разнообразия.
Эта модель, разумеется, не имеет ничего общего с однофакторными катастрофическими гипотезами вымирания вследствие прямого воздействия низкой температуры, радиации, выбросов угарного газа или загрязнения тяжелыми металлами. В подобных гипотезах обычно смешиваются два понятия — массовая смертность и массовое вымирание. Массовая смертность может быть частью адаптивной стратегии вида (как у леммингов, например) и, как правило, не угрожает его существованию. В отличие от этого массовое вымирание, т. е. исчезновение доминирующих видов, связано с перестройкой экосистем и радикальным изменением адаптивной стратегии видов.
Однофакторные гипотезы вымирания до последнего времени оставались в значительной мере умозрительными и не привлекали большого интереса. В конце 1970-х годов группа исследователей, изучавших содержание иридия в разрезах меловых отложений, наткнулась на крупную аномалию в пограничных слоях мела и палеогена. В поисках причины этого явления могли быть плодотворно обсуждены такие гипотезы, как замедление осадконакопления (что подтверждается большой частотой хиатусов в пограничном интервале), появление дополнительных — земных или космических — источников иридия (в частности, таким источником могли быть выведенные на поверхность в меловых подвижных поясах офиолитовые комплексы, перидотитовый компонент которых нередко содержит промышленные месторождения иридия), а также широкое распространение аноксии, способствующее концентрации этого элемента и других металлов в донных осадках; может быть, совместное действие этих факторов. Однако исследователи, обнаружившие иридиевую аномалию, предпочли более сенсационный вариант: космический удар, подобный грандиозному ядерному взрыву, мгновенно уничтоживший динозавров.
Возможно, благодаря апокалипсическим обертонам, напоминающим об угрозе ядерной войны, эта гипотеза завоевала исключительную популярность у широкой публики. Несколько труднее объяснить ее успех среди ученых (впрочем, как мы уже говорили, ученые в наше время не могут пренебрегать общественным мнением). Во многих музеях мира созданы эффектные экспозиции, показывающие агонию динозавров на фоне ядерной вспышки. Рассчитаны вес и траектория «болида смерти», найден даже его гигантский кратер — Чиксулуб в Мексике. Правда, скрупулезные исследования Герты Келлер и ее сотрудников показали, что Чиксулуб никак не может быть «тем самым» кратером, поскольку образовался за 300 тыс. лет до предполагаемого импактного события. Следов иридия в нем тоже не обнаружено. Но это, кажется, никого не обескуражило.
Палеонтологи ищут и, как ни удивительно, находят дополнительные доказательства катастрофического импакта на границе мела и палеогена и других рубежах, отмеченных массовым вымиранием. Так, в спорово-пыльцевых комплексах из пограничных слоев мела и палеогена нередко наблюдается повышенное содержание спор папоротников. Этот «папоротниковый пик» объясняется, в духе импактной теории, гибелью наземной растительности и прорастанием спор на образовавшихся обширных пустошах. На рубеже перми и триаса в прибрежно-морских отложениях часто встречаются клеточные нити Tympanicysta и близких морфотипов, которые были описаны как остатки грибов и послужили основанием для реконструкции «грибного эпизода» — массового размножения сапрофитных грибов вследствие уничтожения растительности очередным столкновением Земли с небесным телом. На худой конец, если никаких следов этого события не обнаружено, то можно написать статью об удивительно быстрой регенерации растительности после импакта.
На самом деле папоротниковые пики приурочены не только к границе мела и палеогена, но и к другим стратиграфическим уровням, отмеченным вулканической активностью, например, к альбским отложениям Дальнего Востока, соответствующим по времени началу образования Тихоокеанского вулканического пояса. Зарастание папоротниками свежевыпавших пеплов и сейчас наблюдается в различных вулканических областях, подсказывая гораздо более правдоподобное объяснение палинологических папоротниковых пиков, чем импактная модель.
Что же касается шумно разрекламированного «грибного эпизода», то он обернулся совсем печальной историей. Исследования, проведенные в лаборатории палеоботаники ПИН РАН с участием альголога С. С. Бариновой, показали, что «грибные нити» — это филаменты зеленых водорослей (рис. 5). Для объяснения их массового развития в глобальных масштабах была предложена гипотеза эвтрофирования пресноводных водоемов и подпруженных эстуариев рек, откуда эвтрофные воды выплескивались в шельфовые моря, вызывая аноксию и сокращение разнообразия как планктонных, так и бентосных сообществ. Определенную роль в развитии этих процессов мог играть обширный наземный вулканизм основного состава как источник железа, ускоряющего рост биомассы водорослевых сообществ. Одновременное распространение эвтрофных вод и придонной аноксии способствовало концентрации металлов в донных отложениях.
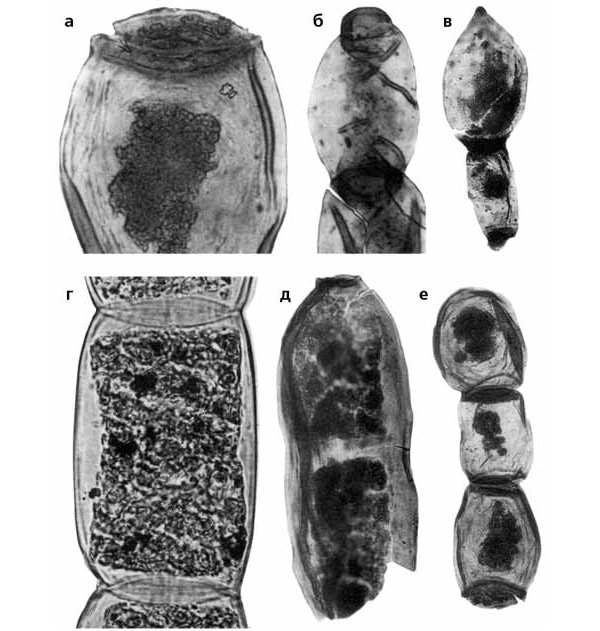
Рис. 5. Tympanicysta из пограничных отложений перми и триаса Русской платформы. Колонии вегетативных клеток и акинеты (слева внизу для сравнения фрагмент нитчатой колонии современной водоросли Spirogyra sp.). Длина масштабной линейки: а, б, г, д — 10 мкм; в — 20 мкм; е — 30 мкм. Изображение «Экология и жизнь»
Может показаться, что речь идет об альтернативных объяснительных моделях: одни связывают важнейшие геоисторические события с космическими ударами, другие — с действием множественных экологических факторов. Первые проще и эффектнее, поэтому и находятся в центре внимания, вторые сложны, неопределенны, во многом недоработаны и пока не могут претендовать на ведущую роль.
В такого рода рассуждениях имеется один существенный изъян. Многофакторные гипотезы требуют определенных знаний и мобилизуют большие массивы сведений из области экологии, палеонтологии и других естественных наук. Импактная гипотеза в этих науках вообще не нуждается. Хотя она претендует на объяснение вымирания динозавров, связь этого события с импактом остается неясной. Если речь идет об уничтожении всего живого ударной волной и/или ядерной вспышкой, то приходится предположить, что жизнь на Земле исчезла и зародилась заново, но так далеко, кажется, никто не заходит. Если причиной был холод «ядерной зимы», то среди динозавров были виды, обитавшие за Полярным кругом и лучше многих не вымерших животных преадаптированные к подобным условиям. Если причиной было нарушение лесной растительности, то для крупных растительноядных животных предпочтительна открытая растительность, и можно даже предположить, что распространение лесов в конце мелового периода сократило кормовую базу динозавров, способствуя их вымиранию.
Первооткрыватель кундурских динозавров Приамурья Ю. В. Болоцкий говорил мне, что у большинства их обнаруживаются те или иные патологии, свидетельствующие о неблагополучном состоянии популяции. Здесь вымирание началось уже в середине Маастрихта. Нельзя забывать и о том, что в истории динозавров было несколько крупных вымираний, которые не стали окончательными, вероятно, потому, что в динозавровых сообществах сохранялись достаточно быстро размножающиеся пионерные формы, позднее замещенные млекопитающими.
Дарвин писал, что для объяснения вымирания динозавров нужно знать, как они жили. Импактная модель в этом не нуждается. Мы изучаем прошлое как ключ к будущему. Но если история Земли — это серия случайных импактов, то ключом к чему она может служить?
К счастью, никаких практических выводов (кроме нежелательности ядерной войны) из импактной модели не следует, разве что возникающее чувство безнадежности (не от бомбы погибнем, так астероид прикончит) все же косвенно способствует развязыванию ядерной войны. А в эпистемологическом плане импактная модель чрезвычайно поучительна и уже в силу одного этого ее авторы безусловно заслуживают всех тех престижных премий и почестей, которых они удостоены.
Заключение
Импактная модель убедительно показывает, как простое объяснение сложных событий, по причине своей очевидной неадекватности, дискредитирует саму возможность объяснения, выпуская из бутылки джинна по имени Индетерминизм. Этот джинн обладает заклинанием, делающим всю содержательную науку ненужной. Звучит оно как Конвенционализм. Раз нам все равно не дано ничего понять — один считает так, другой иначе, — так давайте облегчим себе жизнь и условно примем элементарную импактную модель (или плит-тектоническую модель, парниковую модель, молекулярную филогению) за основу. Это позволит нам избежать бесплодных споров и заняться наконец серьезным делом, например, вероятностной оценкой суммарного количества пыли, попадающей от этих виртуальных импактов в стратосферу, с помощью сложных вычислений, без которых немыслима современная наука. А кто нашим договоренностям не следует, с тем мы сотрудничать не будем.
Это серьезная угроза. Но если мы хотим сохранить науку как инструмент познания, а не просто выжить в ней, то ни договариваться, ни принимать на веру, ни приспосабливаться не нужно, потому что стремление к истине для науки важнее единомыслия.
Примечания:
* Isaaks В., Oliver J., Sykes L.R. Seismology and the new global tectonics // J. Geophys. Res. 1968. Vol. 73. № 18. P. 5855–5900.
** См. Красилов В.А. Эволюция и биостратиграфия. — М.: Наука, 1977.
Рекомендуем прочесть
 Динамика психологического состояния российского общества: экспертная оценка
Динамика психологического состояния российского общества: экспертная оценка Спортсмен
Спортсмен О происходящем на Украине
О происходящем на Украине Футбол как глобальный феномен: Исторические перспективы
Футбол как глобальный феномен: Исторические перспективы Одну треть рыбных запасов оставить птицам — новый закон в экологии
Одну треть рыбных запасов оставить птицам — новый закон в экологии Оценка вклада внешних причин смерти в изменение ожидаемой продолжительности жизни в России в 1990-2010 гг.
Оценка вклада внешних причин смерти в изменение ожидаемой продолжительности жизни в России в 1990-2010 гг.  Закон о «запрете» заемного труда дискриминирует одиноких и многодетных родителей
Закон о «запрете» заемного труда дискриминирует одиноких и многодетных родителей Преследования гражданских немцев в Британии в Первую Мировую
Преследования гражданских немцев в Британии в Первую Мировую
