

Сэмюэль Боулз, наиболее известный из аналитических марксистов — исследователей, решающих традиционные для марксизма проблемы методами экспериментальных наук, в первую очередь о поведении. Работы и биография
Резюме публикатора. «Коэволюция парохиального альтруизма и войны» — известная статья Сэмюэля Боулса и Йона-Кью Чо (перевод Максима Дмитриева), представляющая модель прогрессивного развития альтруизма у нашего вида, где движитель — межгрупповые конфликты, и шире, деление на «своих» и «чужих», первых поддерживают и защищают, ко вторым враждебны и дискриминируют. Модель обосновывает, что альтруизм у Homo sapiens развивался именно как парохиальный, распространяющийся лишь на «своих», именно благодаря войнам он вытеснил свою альтернативу (толерантный альтруизм, который для всех, притом что деление на группы остаётся и значимо). Авторы специально подчёркивают, что их модель чисто гипотетическая — «так может быть при ряде предположений», реалистичность которых не обсуждается. Одновременно они показывают, где именно сомнительные места, в чём большой плюс. Правда, в следующей работе, 2009 года (по-русски пересказана Александром Марковым на Элементах.ру) они всё же пытаются представить её эмпирической зависимостью, выведенной из полевых данных и/или объяснением, предложенным для них. Увы, это попытка с негодными средствами: те наблюдения, что авторы могут подогнать к их модели, они подгоняют, что не укладывается в её прокрустово ложе — игнорируют, а так делать не стоит. Впрочем, моделирование, исходящее из социобиологических концепций - отбора генов, прямой связи между генными вариантами и поведенческими альтернативами, обсуждаемый авторами отбор по поведению движущий, в пользу особей с определёнными качествами (распространяемыми в популяции), а не стабилизирующий, в пользу определённых моделей поведения — скорей всего и не будет соответствовать данным, из-за несостоятельности всего направления. Поэтому по ходу изложения мы показываем, где именно построения авторов оказываются нереалистичными, или выводы — сомнительными.
Сегодня уже точно известно, что идея «биологической» связи между групповой солидарностью, кооперацией, альтруизмом и войной не работает не только у людей, но и обыкновенных шимпанзе, выступающих здесь своего рода «образцом». С одной стороны, человеческий коллективизм утвердился задолго до войн, предполагаемых в модели Боулса и Чо, и независимо от них, у предка всей человеческой ветви, в социальном плане близкого к бонобо Pan paniscus. С другой, первобытные войны стали нормой, и достигли жестокости, необходимой для действенности модели, очень поздно в нашей истории — после того, как «биологически» обусловленная агрессивность упала, приведя к т.н.краниофациальной феминизации.
Т.е. первобытные войны, не говоря о последующих — следствие не «биологии», изменяемой эволюцией в сторону солидарности со «своими» и агрессии против «чужих», но идеологии (причём связанной с требованиями первобытной религии для «своих», а не того, что может быть интерпретировано как агрессия — вражда, страх, ненависть к «чужим»). Идеология же определяет границу «своих» и «чужих», оказывающейся ситуативно подвижной, связанной не с репродуктивными сообществами (как было бы при установлении её отбором), а с родоплеменным делением, означенным в культуре, и никогда не являющимся полным вражды (в ряде ситуаций — например, брачных обменов — чужие закономерно делаются «своими», и наоборот).
***
Йон-Кью Чо, Сэмюэль Боулз
Альтруизм, готовность помочь другим членами группы за собственный счет, и парохиальность, враждебность ко всем, кто не принадлежит к “своей” группе (этнической, расовой, или иной) — два типа поведения, характерных для людей. Пересечение этих типов поведения, парохиальный альтруизм, вызывает некоторое недоумение, если смотреть на него с “точки зрения” эволюции. И альтруистическое, и парохиальное поведение уменьшает выигрыш индивида в сравнении с выигрышем, который он получил бы избегая обоих типов поведения, и уж тем более их пересечения.
Но парохиальный альтруизм мог быть фактором эволюции, если парохиальность провоцировала межгрупповые конфликты. В таком случае сочетание альтруизма и парохиальности способствовало бы успеху в конфликтах.
Мы выполнили теоретико-игровой анализ и агентное моделирование данного предположения. Полученные результаты показывают, что в условиях, в которых, вероятно, жили люди позднего плейстоцена и раннего голоцена, ни парохиализм, ни альтруизм не были “жизнеспособны” по отдельности, как типы поведения. Но под воздействием групповых конфликтов — и вызывая эти конфликты — они могли развиваться совместно.
В конце XIX века ученые, от Чарльза Дарвина (1) до Карла Пирсона (2), называли войну мощной эволюционной силой, которая может способствовать социальной солидарности и альтруизму к членам своей группы. Гамильтон (3) выдвигал предположения о работе этой эволюционной силы. Но до сих пор не выполнено исследований, в которых системно изучено, как военные конфликты могли стать настолько распространенными, чтобы способствовать эволюции альтруизма
[если — и только если — биологически детерминированы как войны (организованное и вооружённое насилие одних групп людей в отношении других), так и альтруизм, включая взаимопомощь при решении сложный интеллектуальных задач, какими являются успешные войны. Современные данные показывают, что вряд ли верно как первое, так и второе: оба детерминированы социально, их характерные формы обусловлены культурой и непосредственно на войне опосредованы мотивациями, не имеющими к агрессии отношения.
В т.ч. межгрупповое насилие у людей стало заметным явлением очень поздно, и лишь после существенного снижения «биологической» агрессивности в ходе т.н. краниофациальной феминизации. Больше того, антропогенез сейчас описывается как «самодоместикация» человека, т.к. к происхождению Homo sapiens применяется концепция доместикации Д.К.Беляева, неотделимая от отбора на миролюбивый характер, неагрессивное восприятие других людей. Прим.публикатора].
И, наоборот, еще нет исследований о том, как альтруизм, обусловленный членством в группе, мог стать причиной и способствовать необычайно высокому уровню убийств в межгрупповых конфликтах.
Альтруизм и враждебность по отношению к членам других групп хорошо изучены на эмпирических данных. Результаты экспериментов и наблюдений показывают, что люди зачастую охотно помогают незнакомцам, вознаграждают за добрые дела и наказывают людей, нарушающих социальные нормы — даже если цена наказания высокая (4). В то же время люди отдают предпочтение членам “своей” группы при выборе друзей, выбирают для обмена и других ассоциированных взаимодействий партнеров из “своих”, отдают предпочтение “своим” при распределении ценных ресурсов (5).
[И вообще, предпочитают «своих» при любом разделении на своих и чужих; правда, лишь после его возникновения, производимого механизмами не природы, а культуры, и ими же быстро меняющегося. Межгрупповая вражда усиливается конкуренцией и, в свою очередь, делает её напряжённей. Больше того, помощь незнакомцам наиболее вероятна у тех, чей жизненный опыт не ограничен «своими», а включает людей разного происхождения, социального статуса, убеждений, пола и возраста: так легче понять, что адресат альтруизма — «человек вообще».
«Люди, так же как и животные, реагируют на тех, кто был рядом с ними, когда они взрослели, словно это их родственники (Aldhous, 1989; Wells, 1987). Хотя этот признак общего родства может время от времени сбивать нас с толку, он чаще оказывается правилен, потому что дома, как gравило, находятся члены настоящей семьи, то есть группы, где почти каждый включает себя в общее «мы».
Интересный вывод состоит в том, что те люди, родители которых регулярно приглашали в дом разных людей, с разным образованием, обычаями и внешностью, когда вырастают, чаще всего помогают незнакомцам. Так происходит потому, что их концепция «мы» расширяется и в дальнейшем включит не только членов семьи, но и других людей. Для них вдохновляющее на помощь чувство «мы» простирается на весь род человеческий (Hornstein, 1982; Piliavin, Dovidio, Gaertner & Clark, 1981)».
(Из статьи Роберта Б. Чалдини и др., 1997 про связь эмпатии и альтруизма).
Иными словами, люди вырастающие в более разнообразном окружении (по соцпроисхождению, занятию, нации, полу, цвету кожи и пр.) — т.е. в более эгалитарном обществе, с меньшим уровнем расизма и ксенофобии, более склонны помогать не только «ближним», но и всем людям вообще. По той же причине а) в одном и том же обществе люди более коллективисткой культуры — т.е. бедняки в сравнении с богатеями — более склонны помогать незнакомцам (см.первую ссылку), в) то же фиксируется у эгалитарных шимпанзе бонобо, но не у деспотико-иерархических Pan troglodytes.Ситуация, заметим, трудно совместимая с готовностью воевать с «чужаками»; и действительно, взаимное доверие и готовность помогать незнакомцам в современном мире растут как функция не житейского благополучия, но мира и социального равенства. Прим.публикатора].
Например, в недавнем эксперименте “наказание третьей стороной” в Папуа-Новой Гвинее участники демонстрировали сильный фаворитизм по отношению к собственной лингвистической группе, при распределении “ресурсов”. В том же эксперименте участники использовали большее наказание — в сравнении со “своими” — для “чужаков” из другой лингвистической группы, если “чужаки” действовали недоброжелательно по отношению к членам “своей” для участника эксперимента группы (6).
Межгрупповая враждебность и агрессия в чем-то схожи с альтруизмом — индивид, ведущий себя парохиально, как и ведущий себя альтруистично, может подвергаться смертельному риску. Придерживающийся парохиальности также может упустить благоприятные возможности для коалиций, взаимного страхования и для обмена. Он несет потери в приспособленности по сравнению с индивидами, которые избегают враждебности к другим группам. Если мы наблюдаем такое поведение, и если члены “своей” группы получают выгоду от враждебных действий индивида к другим группам — мы наблюдаем “парохиальный альтруизм” (условимся, что будем называть подобное явление именно так). Пример парохиального альтруизма — результаты экспериментов в Папуа-Новой Гвинее (см. ниже).
Ни парохиализм, ни альтруизм, по отдельности, видимо, не сохранятся в процессе отбора, который благоприятствует поведенческим чертам, увеличивающим “выигрыш”, выгоду для индивида. Но парохиальный альтруизм, их сочетание, мог возникнуть и распространиться среди предков, потому что они жили в среде с [жестокой] конкуренцией за ресурсы. К такой среде лучше приспособятся группы со значительным количеством парохиальных альтруистов — последние готовы и желают участвовать в конфликтах с чужаками, от имени представителей своей группы.
Подобный выигрыш для группы мог бы компенсировать внутригрупповой отбор как “против” альтруизма, так и “против” парохиальности. В отличие от многоуровневых моделей отбора, в которых существование групповых конфликтов взято, как простое предположение (7-9), мы предоставляем объяснение для причин войны и ее уникальной “смертоносной природы” в поведении людей. Правдоподобно ли это объяснение — вопрос эмпирической проверки.
Данные этнографических и археологических исследований “свидетельствуют”, что война, сражения — причина высокой смертности у некоторых групп охотников-собирателей и в ранних племенных обществах (10, 11). В межгрупповых конфликтах наши предки, возможно, теряли на порядок больше людей, чем европейцы в воинственном 20 веке.
Самые жестокие межгрупповые конфликты — непрерывные или с “перемириями”, с сопутствующими жертвами — вероятно, больше походили на пограничные конфликты у шимпанзе (12), чем на современные войны. Однако и среди охотников-собирателей случались “генеральные сражения”, вроде описанного Лорандосом конфликта двух коалиций австралийских аборигенов, в котором участвовало около 700 воинов (13).
Исследователи вопроса установили, что наиболее важные корреляты войн между группами охотников-собирателей — это стихийные бедствия и периодическая нехватка ресурсов (14). [Более тщательное антропологическое исследование, однако, показывает, что даже в суровых условиях агрессия у охотников-собирателей в целом снижена, с увеличением (или ростом разнообразия) ресурсной базы она скорей увеличивается, а различия в агрессивности между разными этносами охотников-собирателей обусловлены разной культурой, не ресурсами, и не физиологической агрессивностью. Прим.публикатора].
Поэтому представляется вероятным, что неустойчивый климат позднего Плейстоцена (примерно 125 000 — 10 000 лет до н.э.) способствовал высокому уровню насилия и частым межгрупповым конфликтам. При этом группы, которые избегали конфликтов, выиграли от лучшего доступа к ресурсам в тех областях, которые в противном случае становились бы защитными буферными зонами, из которых не удастся извлечь ресурсы (15, 16). Также, группы, избегающие конфликтов, выигрывали от межгруппового распределения рисков (17) и взаимовыгодного обмена (18), часто на значительных расстояниях [см. (19)].
Мог ли возникнуть и распространиться парохиальный альтруизм в такой среде? Мы моделируем эволюцию генетически передаваемых типов поведения в популяции охотников-собирателей, которые участвуют как во внутригрупповых, так и в межгрупповых взаимодействиях.
Индивидуумы в группе могут быть альтруистичны (или нет) и парохиальны (или нет). В модели мы представляем эти типы поведения через две гипотетические аллели, каждая из двух локусов. Таким образом, моделируется четыре типа поведения:
-
парохиальных альтруистов (parochial altruists, PA, носители обладают P и A аллелями)
-
парохиальных не-альтруистов (PN)
-
толерантных, не-парохиальных альтруистов (TA)
-
толерантных не-альтруистов (TN).
Парохиальные индивиды (любого типа) враждебны по отношению к другим группам. Но только парохиальные альтруисты участвуют в боях, потому что парохиальные не-альтруисты не хотят рисковать жизнью для блага своих товарищей по группе. При отсутствии межгрупповых конфликтов толерантные члены группы извлекают выгоду из помянутых выше обмена, распределения риска с другими группами и из ресурсов доступных в не-буферных зонах.
В представленной модели действуют два вида отбора. Внутригрупповой отбор благоприятствует толерантным не-альтруистам и способствует уничтожению парохиальных альтруистов, а также толерантных альтруистов и парохиальным не-альтруистам. Второй вид отбора, выборочное истребление (selective extinction) в результате межгруппового конфликта, напротив, может благоприятствовать парохиальным альтруистам, несмотря на то, что они рискуют жизнью даже в победоносных боях.
Чтобы лучше объяснить роль войны, парохиальности и выборочного истребления, мы не моделируем другой механизм распространения альтруизма — избирательную миграцию (20). Таким образом, в отсутствие территориальной экспансии через завоевание, мы предполагаем, что размер групп фиксирован, поэтому высоко альтруистичные группы не воспроизводятся значимо в следующих поколениях. Наша модель не способствует эволюции альтруизма, поскольку эквивалентна моделям, в которых “локальный” отбор зависит от плотности населения и смещает-компенсирует преимущества альтруизма для групп (21, 22).
Парохиальные альтруисты, выжившие в бою, получают личную репродуктивную выгоду — ведь они разделяют со своей группой повышенную вероятность победы, благодаря своему статусу и умениям “бойца” (относительно ожидаемого исхода конфликта, если бы индивид предпочитал другое поведение). Победа в бою дает два вида репродуктивных преимуществ членам группы: больше шансов на выживание и — для выживших — возможность произвести больше потомства, на замену погибшим, в побежденной группе.
Преимущества заманчивы, но, по результатам нашего моделирования (19), повышенный риск смертности в бою для парохиальных альтруистов с лихвой их перекрывает. То есть, каждого парохиального альтруиста ожидает значимо больший репродуктивный успех, стоит только переключиться на толерантное и/или неальтруистичное поведение. И шанс на успех выше даже с учетом того, что “переключение” на другой тип поведения увеличивает вероятность поражения группы в конфликте с чужаками. Поэтому тех, кто сражается за свою группу, можно назвать альтруистами и в общепринятом смысле слова (23).
В нашей модели, в каждом поколении, все члены группы случайным образом спариваются с другими членами (своей группы), чтобы получить потомство [здесь нужно добавить, что большая склонность убивать «чужих» на войне в среднем позитивно связана с убийствами «своих» криминальным образом, что дополнительно снижает приспособленность «воителей». Авторы это не учитывают. Прим.публикатора]. Ожидаемое число потомков пропорционально доле родительской пары в их заслугах перед группой (group’s payoffs). Заслуги описаны чуть ниже. Чтобы не “подыгрывать” нашей гипотезе о коэволюции парохиальности и альтруизма, зависящей от двух видов поведения, связанных статистически, мы делаем предположение, что процесс передачи (поведения) из поколения в поколение происходит без “встроенной” тенденции к корреляции парохиальных и альтруистичных аллелей. Таким образом, мы предполагаем отсутствие внутригрупповой сортировки в спаривании и допускаем возможность полной рекомбинации. Так, например, родительская пара, состоящая из парохиального альтруиста (PA) и толерантного не-альтруиста (TN) с равной вероятностью может родить потомство всех четырех поведенческих типов.
Кроме того, процесс передачи поведения между поколениями модифицируется мутацией. С некоторой вероятностью μ потомки любого члена группы случайным образом наследуют поведенческий тип, независимо от родительских типов. С вероятностью не выше 1 − μ, происходит не-мутантная репликация. И, наконец, с еще одной вероятностью m каждое поколение переходит в другую, произвольно выбранную группу “чужаков”.
Взаимодействие между группами выглядит следующим образом. Каждое поколение в группе взаимодействует с другой группой, выбирая либо сотрудничество, либо вражду (График 1).
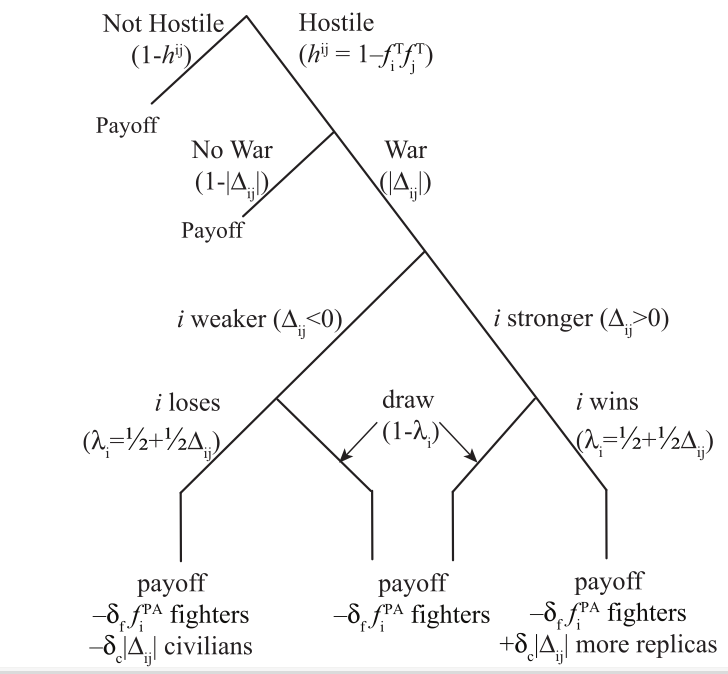 Вражда в межгрупповом взаимодействии возникает, если доля парохиальных альтруистов хотя бы в одной группе достаточно велика. Выбор в пользу кровопролития, использования силы в отношениях между двумя группами, происходит, когда одна из групп с высокой вероятностью победит. Такой “избыточный” выбор отражает факт, что — как и в случае с другими приматами — равные по возможностям группы людей стремятся избежать дорогостоящих конфликтов (24).
Вражда в межгрупповом взаимодействии возникает, если доля парохиальных альтруистов хотя бы в одной группе достаточно велика. Выбор в пользу кровопролития, использования силы в отношениях между двумя группами, происходит, когда одна из групп с высокой вероятностью победит. Такой “избыточный” выбор отражает факт, что — как и в случае с другими приматами — равные по возможностям группы людей стремятся избежать дорогостоящих конфликтов (24).
Вероятность того, что группа победит в конфликте, зависит от разницы в количестве бойцов, парохиальных альтруистов, в каждой из групп. В боях погибает какое-то количество бойцов с обеих сторон, но — в проигравшей группе победители убивают и выживших бойцов, и часть “гражданских”, которые статусом бойцов не обладают. Доля смертей среди “гражданских” равна постоянной разнице (в количестве?) между группами в доле парохиальных альтруистов, поэтому чем больше дисбаланс сил, тем более тяжкая участь ждет проигравших. Однако если группа с большим количеством бойцов не побеждает, и итогом боев становится “ничья” — обе группы теряют часть своих бойцов, но не “гражданских” (не-бойцов) [ещё одно не вполне реалистичное предположение, у многих охотников-собирателей границ между ними по сути нет. Прим.публикатора].
Те, кто выбыл из обеих групп, по результатам конфликта, заменяются потомством от случайно выбранных родителей — по механизму, описанному выше — из победившей группы. То есть победители присоединяют (или присоединяются) проигравшую группу, доводя численность обеих групп до их первоначальной “емкости”. Мы смоделировали и альтернативу этому “миграционному” сценарию (19): бойцы победившей группы убивают меньше проигравших, вместо этого “спариваясь” с большим количеством поверженных, “перенаселяя” группы подобным образом.
Такой сценарий “спаривания” благоприятствует эволюции парохиального альтруизма даже больше, чем смоделированные результаты ниже. “Спаривание” вместо “мигрирования” дает привилегии парохиальным альтруистам — потому что именно они, как бойцы, будут в первую очередь оплодотворять членов поверженной группы, в процессе повторного заселения. В сценарии “миграции”, те, кто воспитывает колонистов для повторного заселения группы и территории поверженных, выбираются случайным образом из населения победившей группы.
Если группы выбирают взаимодействие, а не вражду, каждый индивид с толерантным поведением (TA) получает чистую выгоду от каждого члена с поведением TA из противоположной группы. Парохиальные альтруисты выгоды не получают. Таким образом, при отсутствии вражды, ожидаемая выгода толерантных альтруистов в данной группе превышает выгоду парохиальных альтруистов, независимо от доли толерантных альтруистов в группе. Без вражды толерантность становится доминирующей стратегией [т.е. толерантным альтруистам в каждой группе выгодно унять и/или смирить выделяющихся боевитостью, что у видим у бушменов и других охотников-собирателей. Разделение групп остаётся, поскольку функционально в плане непревышения предельной нагрузки на пищевые ресурсы, и общего рассредоточения, но их отношения вполне мирные. Прим.публикатора].
Это сценарии взаимодействия между группами. Как распределяются индивидуальные выигрыши при внутригрупповых взаимодействиях? В каждом поколении альтруисты, “уплачивая” цену c, вносят свой вклад в общественное благо, ценность которого b поровну распределяется между n членами группы, принадлежащими к одному поколению. Общественным благом трактуется здесь широко — это может быть обмен информацией, разделение рисков с членами группы и другое подобное поведение. Не-альтруисты вклада не вносят. Поскольку b > c > b/n, вклад повышает средний выигрыш членов группе. Но выигрыш вкладчика-альтруиста будет “увеличен” (уменьшен) на сумму c — b/n, в сравнении с теми, кто не внес вклад, независимо от количества других альтруистов в группе.
В результате, в “мирное время” как толерантные альтруисты, так и парохиальные альтруисты сталкиваются с неблагоприятным внутригрупповым отбором. Механизм работы отбора и “выигрышей” продемонстрирован в Таблице 1.

Таблица 1. Ожидаемые выгоды (польза, выигрыш) для четырех типов поведения: общественные блага и мирное межгрупповое взаимодействие. Доля альтруистов в группе i обозначена, как . Все члены группы получают выгоду от общественного блага, которая обозначена, как . Толерантные члены группы, обоих типов, TA и TN, получают выгоду от мирного межгруппового взаимодействия, , где g — это благо от мирного группового взаимодействия, n — количество членов группы в одном поколении, а -это доля толерантных членов другой группы, которые вступают в обмен. Если межгрупповое взаимодействие враждебное, то толерантные члены группы не получают выгоды, равной . Имитационные контрольные значения-бенчмарки параметров в таблице следующие: c = 0,01, b = 0,02, g = 0,001, m = 0,25, μ = 0,005, n = 26.
|
Парохиальность |
Толерантность |
|
|
Альтруисты |
bfiA — с |
bfiA— с + gnfjT |
|
Не альтруисты |
bfiA | bfiA + gnfjT |
Последовательность событий для каждого поколения следующая: на первом этапе происходит групповые взаимодействия, за которым [если выбрана вражда] следует перезаселение той группы, которая потерпела поражение в войне. Члены восстановленных групп затем взаимодействуют в распределении общественных благ, по теоретико-игровым моделям, после чего размножаются пропорционально своей доле от общего выигрыша группы. В среднем, родительские пары обзаводятся как минимум двумя доживающими до возраста зреллости потомками, чтобы сохранить размер группы. Наконец, родительское поколение умирает и происходит переход к последовательности событий для нового поколения.
Мы посчитали в коде агентную модель, в которой задали — для целей нашего исследования — значения параметров (19), откалиброванных так, чтобы они походили на среду обитания людей позднего Плейстоцена и раннего Голоцена, т. е.приблизительно соответствовали условиям жизни 7000 лет назад. Размер контрольной группы в модели — 78 индивидов, размер одного поколения — n = 26 индивидов. Можно представить себе контрольную группу, как три поколения, или коалицию из трех групп времен позднего Плейстоцена (25). Принятый базовый уровень миграции, 25% на поколение, основан на имеющихся наблюдениях за жизнью современных групп охотников-собирателей. Метапопуляция в модели состоит из 20 групп, общепринятый размер для изучаемых поздне-плейстоценовых этно-лингвистических групп.
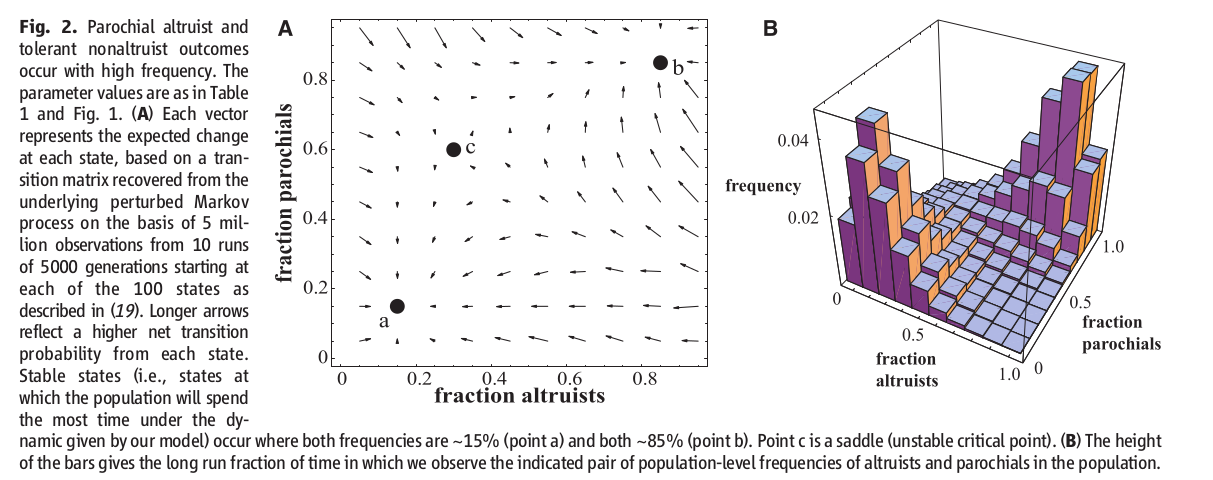
График 2. Парохиальные альтруисты и толерантные не-альтруисты “возникают” с высокой частотой и преобладают в распределении. Значения параметров приведены в Таблице 1 и на Графике 2А. На изображении каждый вектор — это ожидаемое изменение в каждом состоянии [распределения], на основе матрицы переходов, взятой из лежащего в основе расчетов Марковского процесса со случайными возмущениями. Данные для процесса — это 5 миллионов наблюдений за 10 прогонов для 5000 поколений, каждое из которых начинает с одного из 100 состояний [распределения типов поведения в группе], как описано в (19). Длинные вектора-стрелки отражают более высокую вероятность перехода из каждого состояния. Стабильные состояния — те состояния, в которых население проводит больше всего времени, в соответствии с динамикой, положенной в основу модели — наблюдаются (происходят) там, где частотность распределения лежит в диапазоне от ~15% (точка a) до ~85% (точка b). Точка c — это седло, то есть неустойчивая критическая точка. На Графике 2Б высота столбцов отражает долю времени, долгосрочную перспективу, в которой мы наблюдаем распределение в популяции числа (частотности) альтруистов и приверженцев парохиальности.
На Графике 2А приведена аппроксимация явной динамики Марковского процесса. Стрелками помечен [эволюционный] отбор против парохиальных не-альтруистов, в отсутствии альтруистов, и наоборот, как и ожидалось. На Графике 2B показано, что происходит в динами Марковского процесса. В течение очень длительного периода времени смоделированное население живет, преимущественно, в двух распределениях. Первое — состояние, при котором много парохиальных альтруистов и мало индивидов трех других поведенческих типов. Второе — когда преобладают толерантные не-альтруисты.
В первом распределении, высокий уровень парохиальности ведет к частым конфликтам, причем победителями становятся те группы, в которых преобладают альтруисты. Во втором распределении, с многочисленными толерантными не-альтруистами, вражда встречается редко, выигрыш межгрупповых взаимодействий существенен, а преобладающий внутригрупповой отбор направлен против парохиальности и альтруистов.
Из статистического анализа [поведения] многих и многих поколений, представители которых сосредоточены вблизи точки b на Графике 2А (распределение “много парохиальных альтруистов”), следует любопытный вывод. Количество межгрупповых конфликтов и количество смертей в бою, которые “требуются”, чтобы поддерживать и альтруизм, и парохиализм на высоком уровне — значительно ниже того количества боев и смертей, которые сопутствовали жизни наших предков в позднем Плейстоцене и раннем Голоцене, по оценкам археологических и этнографических данных (19). При воинственном распределении вблизи точки b на Графике 2А в каждом поколении 3,6% от популяции умирают в бою. И это приблизительно в три раза меньше, чем тот уровень смертности в бою, который сопутствовал предкам, если судить по этнографическим и археологическим данным (11). Для результатов нашего моделирования не требуется какого-то невероятно высокого уровня смертности, вызванного войной.
На Панели 3, на первых четырех графиках (сверху вниз), проиллюстрирован процесс перехода между распределениями, от распределения вокруг точки a к распределению вокруг точки b. Переходы в модели случаются не часто и резко, причем переходы идут в обоих направлениях. Из-за случайного складывания родительских пар и групп, случайных результатов групповых взаимодействий, миграции и мутации — популяция может переместиться из окрестностей точки a (или точки b) в ту область, где силы отбора, представленные стрелками на Графике 2A, способствуют “передвижению” населения в противоположный угол распределения поведений.
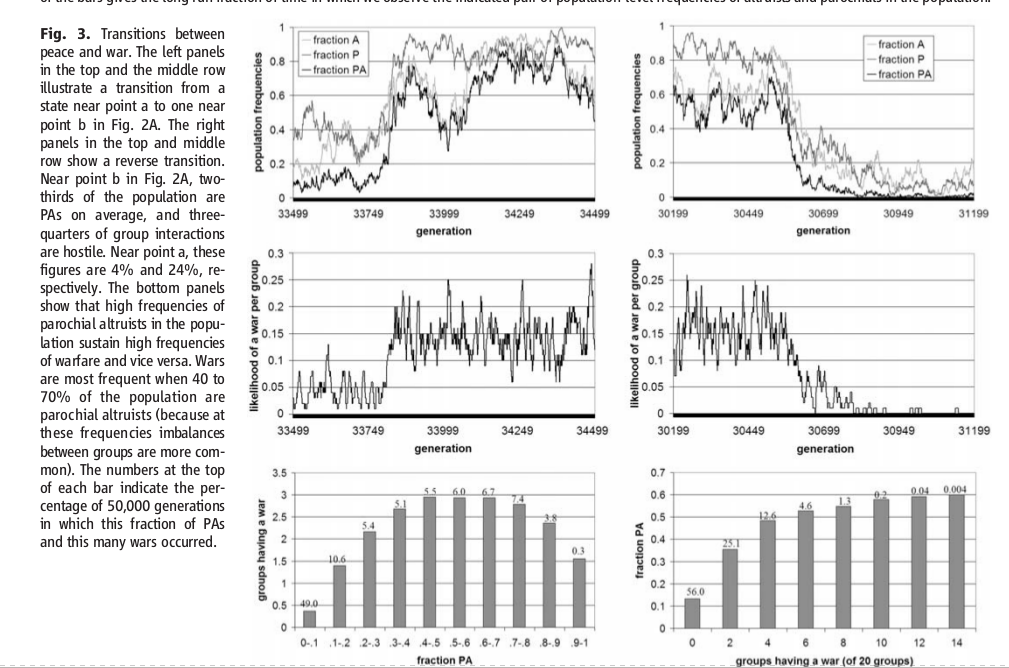
Панель 3: Переход от мира к войне. В верхнем и среднем ряду, левые графики иллюстрируют переход из состояния, близкого к точке а — много толерантных не-альтруистов — в состояние, близкое к точке b, с Графика 2A. Правые графики в верхнем и среднем ряду иллюстрируют обратный переход. Вблизи точки b на Графике 2A, в среднем, две трети популяции — это парохиальные альтруисты, а три четверти групповых взаимодействий это война. Вблизи точки a эти показатели равны, соответственно, 4% и 24%. Нижние графики показывают, что высокая частотность распределения парохиальных альтруистов в популяции поддерживают высокую частотность войны, и наоборот. Войны становится больше, когда от 40 до 70% популяции приходится на парохиальных альтруистов — потому что на такой частотности распределения поведенческих типов дисбаланс между группами более распространен. Цифры над столбцами указывают процент частотности войн — сколько войн пришлось на каждое из 50 000 поколений в модели, с данной частотой распределения парохиальных альтруистов.
Нижняя панель Графика 3 — совокупные данные по распределениям, на большом количестве запусков модели. Левая (нижняя) панель интересна тем, что показывает, что когда среди населения начинают преобладать парохиальные альтруисты, войн происходит… меньше. Потому что группы, как правило, равны. Нижняя правая панель показывает, что, с ростом числа войн начинает расти и число парохиальных альтруистов, среди населения.
Для дополнительной проверки мы поэкспериментировали со значениями параметров в модели (19). “Эксперименты” показывают, что частота распределения парохиальных альтруистов в популяции и уровень смертности в войнах могут обратно зависеть от размера группы и скорости миграции. Такой тип зависимости следует из того, что указанные “параметры” населения уменьшают различия между группами, в распределении поведенческих типов. Тем самым уменьшается частота войн и ослабляются последствия выборочного истребления во время войны. Возможна и прямая зависимость: смерти на войне и частота распределения парохиальных альтруистов зависят от масштабов потерь “гражданских” в поверженных группах.
При этом изменение уровня смертности среди бойцов сперва увеличивает “общее” количество смертей на войне, а затем понижает — при очень высоком уровне смертности среди бойцов немногие парохиальные альтруисты выживают. В популяции становится меньше бойцов, и как следствие, становится меньше войн. Полученные результаты не очень чувствительны к вероятным изменениям в выгодах и стоимости альтруизма.
Однако таких результатов мы не получаем: как описано выше, у парохиальных альтруистов есть прямая выгода от увеличения вероятности победы в войне. И действительно, при большинстве распределений поведенческих типов в модели, от поколения к поколению, индивид, который гипотетически перешел бы в парохиальные альтруисты, понес бы потери большие, чем стоимость с вклада в общественные блага внутри группы.
Два стабильных состояния распределения, точки a и b на Графике 2А, возникают потому, что поведение парохиальных альтруистов и толерантных не-альтруистов соответствуют поведенческим типам из модели “избирательного отбора” (selective assortment) Эшеля и Кавалли-Сфорца (26). Привилегированный отбор (preferential assortment) с генетическим родством (genetic kin) в модель не включен — группы достаточно велики и как миграция, так и внутригрупповое деторождение являются случайными.
Важнейшие процессы отбора, которые объясняют распределение поведенческих типов в группах, возникают, скорее, эндогенно, из условий межгрупповых взаимодействий.
Когда группы кооперируются, толерантных не-альтруистов становится больше, потому что они извлекают выгоду из “позитивного” отбора — выгоду из, собственно, кооперации и обмена групп. Да и сотрудничать могут только те группы, у которых большинство членов толерантны.
Когда группы воюют, начинает работать “негативный” отбор (negative assortment), благоприятный для парохиальных альтруистов. При равномерном распределении типов поведения среди членов разных групп, такие группы избегают войн. Чем пользуются группы с большим количеством парохиальных альтруистов: войны, в которых они участвуют (и побеждают), они ведут, как правило, против групп с равномерным распределением типов поведения. Высокий репродуктивный успех групп парохиальных альтруистов, следующий успехами их групп в войне — дар, который парохиальные альтруисты даруют своим товарищам по группе. И этот дар — приносит непропорционально высокую ценность другим парохиальным альтруистам и членам группы, чем и объясняется их успех.
В модели мы показали, что переход от преобладания толерантных не-альтруистичных индивидов — и, следовательно, от состояния мира — к парохиальным альтруистам и войне может быть очень быстрым. Доминирующие распределения меняются менее, чем за 200 поколений, или за приблизительно 5000 лет (График 3, с панелями).
Более высокий репродуктивный успех групп с преобладанием парохиальных альтруистов, при взаимодействии с группами без значимого число парохиальных альтруистов, может, таким образом, объяснить реальную историю. Объяснить “паттерн” быстрой экспансии, который считается общим для некоторых человеческих групп, племен, живших в позднем Плейстоцене. Используя модель, можно частично объяснить все еще до конца непонятную вторую “великую диаспору гоминидов”, которая за каких-то 10 тысяч лет рассеялась по планете, из Африки добравшись даже до Австралии. [Что произошло после снижения биологической агрессивности; отбор здесь если имел место, то лишь стабилизирующий, на приспособление наших тел, их физиологии и пр. к меняющейся культуре. Прим.публикатора]
Описанная в статье динамика коэволюции парохиального альтруизма и войны предоставляет правдоподобное объяснение для результатов поведенческих экспериментов, вроде проведенного в Папуа-Новой Гвинее. Если взять нашу модель, как объясняющую, то можно ожидать, что толерантные альтруисты согласятся нести издержки, чтобы распределять выигрыш как для “своих”, так и для “чужаков”, и наказывать нарушителей норм. Учитывая важность взаимовыгодного межгруппового взаимодействия, толерантные альтруисты должны одинаково жестоко наказывать нарушителей норм и среди чужаков, и среди своих [и будут выигрывать в обычной для первобытных народов ситуации, когда соседние группы связаны брачными обменами, т.е. многие люди в каждой из них отчасти свои, отчасти чужие, и в каждом конфликте должны самоопределяться, что очень непросто; проще с «обоих сторон» действовать в пользу мира, что способствовало прогрессу процедур примирения, тем более изощрённых и действенных, чем выше была военная активность до того. Прим.публикатора]. А вот парохиальные альтруисты благосклонны к своим и более жестоко наказывают чужаков, которые причиняют вред своим.
В нашей модели показано, как враждебное поведение к чужакам — и другие типы поведения в эксперименте — стало бы развиваться, на пользу другим членам группы, при последующих межгрупповых конфликтах. Эксперимент очень хорошо соответствует модели: в нём наказание для “чужаков” увеличивало относительный выигрыш для “своих”, потому что стоимость удачного завершения эксперимента в три раза превышала стоимость наказания (“чужаков” наказывать дешевле).
Распределение в пользу других в эксперименте — даже в пользу членов своей группы — не удастся объяснить одним только родственным альтруизмом (kin altruism). Стоимость распределения благ (выигрыша) равна выгоде для получателя, поэтому члены группы не выбрали бы такое поведение, даже если бы являлись идентичными близнецами.
Наконец, модель, с разными параметрами, помогают дать “эволюционное” объяснение причин, по которым границы групп — культурные, территориальные, иные — так сильно влияют на поведение человека (27-29).
Мы объяснили, как Homo sapiens мог стать и воинственным, и альтруистичным видом. Но нет никаких доказательств, что гипотетические аллели из нашей модели существуют в реальности. А даже если бы они существовали — вряд ли их можно было бы выразить, “вычленить” из такого сложного поведения, как помощь другим, или участие в войне. Таким образом, мы не доказали, что у людей существует некая генетическая предрасположенность к войне. Мы доказали только то, что если бы такая предрасположенность существовала, она могла бы сосуществовать с альтруизмом — в том виде, в котором мы описали выше.
Модель вертикальной передачи генов (от родителей к детям) можно улучшить и расширить, чтобы включить в неё культурное обучение и учесть влияние сверстников, а также взрослых и родителей. Расширение модели было бы важно, если бы выводы о современном поведении делались только исходя из тех же предположений, что лежат в основании модели. Однако существует множество доказательств того, что человеческую парохиальность можно перенаправить и даже преодолеть целенаправленным обучением, случайными воздействиями и другими социализирующими процессами.
Источники и сноски
-
C. Darwin, The Descent of Man (Prometheus Books, Amherst, NY, 1998).
-
K. Pearson, Fortnightly Review 56, 1 (1894).
-
W. D. Hamilton, in Biosocial Anthropology, R. Fox, Ed. (Wiley, New York, 1975), pp. 133–155.
-
E. Fehr, U. Fischbacher, Nature 425, 785 (2003).
-
T. Yamagishi, N. Jin, T. Kiyonari, Adv. Group Process. 16, 161 (1999).
-
H. Bernhard, E. Fehr, U. Fischbacher, Nature 442, 912 (2006).
-
K. Aoki, Evolution 36, 832 (1982).
-
S. Bowles, J.-K. Choi, A. Hopfensitz, J. Theor. Biol. 223, 135 (2003).
-
R. A. Guzman, C. Rodriguez-Sickert, R. Rowthorn, Evol. Hum. Behav. 28, 112 (2007).
-
L. Keeley, War Before Civilization (Oxford Univ. Press, New York, 1996).
-
S. Bowles, Science 314, 1569 (2006).
-
J. H. Manson, R. W. Wrangham, Curr. Anthropol. 32, 369 (1991).
-
H. Lourandos, Continent of Hunter-gatherers (Cambridge Univ. Press, Cambridge, 1997).
-
C. R. Ember, M. Ember, J. Conflict Resolut. 36, 242 (1992).
-
R. C. Kelly, Warless Societies and the Origin of War (Univ. of Michigan Press, Ann Arbor, MI, 2000).
-
S. A. LeBlanc, Constant Battles (St. Martins, New York, 2003).
-
P. Wiessner, Evol. Hum. Behav. 23, 407 (2002).
-
S. McBrearty, A. Brooks, J. Hum. Evol. 39, 453 (2000).
-
See Online Materials.
-
R. Rogers, Am. Nat. 135, 398 (1990).
-
P. D. Taylor, Evol. Ecol. 6, 352 (1992).
-
D. S. Wilson, G. B. Pollock, L. A. Dugatkin, Evol. Ecol. 6, 331 (1992).
-
B. Kerr, P. Godfrey-Smith, M. Feldman, Trends Ecol. Evol. 19, 135 (2004).
-
M. Wilson, M. D. Hauser, R. W. Wrangham, Anim. Behav. 61, 1203 (2001).
-
F. Marlowe, Evol. Anthropol. 14, 54 (2005).
-
I. Eshel, L. L. Cavalli-Sforza, Proc. Natl. Acad. Sci. U.S.A. 79, 1331 (1982).
-
R. McElreath, R. Boyd, P. Richerson, Curr. Anthropol. 44, 122 (2003).
-
R. L. Axtell, J. M. Epstein, H. P. Young, in Social Dynamics S. Durlauf, H. P. Young, Eds. (MIT Press, Cambridge, 2001) pp. 191-211.
-
R. A. Hammond, R. Axelrod, Theor. Popul. Biol. 69, 333 (2006).
Science, Vol. 318, 26 october 2007
Перевод Максима Дмитриева
Об авторах: Йон Кью Чо — профессор школы экономики и торговли Кёнбукского Национального Университета (School of Economics and Trade, Kyungpook National University, 1370 Sankyuk-dong, Buk-gu, Daegu 702-701, Korea).
Сэмюэль Боулз — профессор института Санта-Фе, Нью-Мексико, и департамента экономической политики Сиенского Университета (Santa Fe Institute, 1399 Hyde Park Road, Santa Fe, NM 08571, USA; and Dipartimento di Economica Politica, University of Siena, Piazza San Francesco, 7, 53100 Siena, Italia).
Мы благодарны за возможность воспользоваться программой поведенческих наук в Институте Санта-Фе, Фонду Рассела Сейджа и Сиенскому университету. Мы благодарим Роберта Бойда, Эрнста Фера, Стивена Ле Блана, Питера Ричерсона, Роберта Роуторна, Тошио Ямагиси, Полли Весснер и Элизабет Вуд за помощь и вклад в наши исследования.
Рекомендуем прочесть
 «Кровавые когти и клыки профессора Пинкера». Ч.1.
«Кровавые когти и клыки профессора Пинкера». Ч.1. Что не так с социобиологией?
Что не так с социобиологией? 130 лет после «антропологических» работ Энгельса: что подтверждается и что — нет?
130 лет после «антропологических» работ Энгельса: что подтверждается и что — нет? Война и мир в первобытном обществе
Война и мир в первобытном обществе «Животное» и «человеческое», расчёт и равенство
«Животное» и «человеческое», расчёт и равенство Затопит ли нас глобальное поглупение?
Затопит ли нас глобальное поглупение? Человеческая коллективность: социальные сети
Человеческая коллективность: социальные сети Проблемы эволюционной психологии
Проблемы эволюционной психологии
