

Кассийский клёст самец — самая крупноклювая форма из североамериканских. Смог обособиться таксономически (хорошо дифференцированный подвид а, может, и отдельный вид, в отличие от остальных 9 форм) благодаря не столько изолированности от других форм, сколько благодаря отсутствию белок.
Резюме. Один из создателей СТЭ, Николай Владимирович Тимофеев-Ресовский, крупнейшим обобщением в биологии называл «некисельность жизни». Разнообразие биологических форм не перетекает друг в друга, но разделено на дискретные единицы — виды (понятно, какие-то из них мы застали в процессе дифференциации и тогда границу провести трудно, но когда — или если — данный процесс дойдет до конца, она обнаруживается и объективна); это применимо к отдельностям живого покрова планеты — биогеоценозам и т.д. Клёст-еловик Loxia curvirostra — лучший пример некисельности, в первую очередь потому, что здесь её не объяснить жёсткостью территориальных связей особей, вызывающих изоляцию расстоянием и локальные адаптации.
У еловиков всего этого нет: вид в ареале представлен не территориальными популяциями, а кочевыми стаями, постоянно перемещающимися в места богатого урожая; там стая оседает на гнездование, практически без распада (нет территориальности). Стаи структурированы системой иерархии, на основе которой в сезон размножения выделяются пары. Они отнюдь не изолированы демографически изолированные: как между территориальными популяциями, между ними есть переток особей, до 10–15% приходит со стороны.
Несмотря на всё это, изменчивость еловика не континуальна, выделены два ряда дифференциации. Первый морфологический, создан большей или меньшей специализацией к разным видам хвойных, на его основании Craig W. Benkman для территории США выделил 10 экотипов, различающихся в основном общей величиной и формой клюва. Он также исследовал дивергентный (дизруптивный) отбор, разделяющий население вида на экотипы: от формы клюва (особенно высоты с шириной бороздок рогового нёба), её «подогнанности» к размерам и жёсткости чешуй шишек того вида (или видов) хвойных, к которым сппециализирован, причинно зависит успешность кормления — чем точнее «подгонка», тем быстрее кормящиеся клесты потребляют семена, что важно в условиях короткого зимнего дня.
Ниже приведён перевод статьи Бенкмана 2003 г., в основном посвящённой самому специализированному из экотипов — клесту Южных Холмов в штате Айдахо, питающегося семнами сосны скрученной широкохвойной, но также и другим формам, приспособленным к добыванию семян из шишек псевдотсуги Мензиса, сосны жёлтой и пр. хвойных. В работе приводится адаптивный ландшафт по Сьюэллу Райту, поскольку красиво показаны жёсткие и однозначные причинно-следственные связи не только (1) специализации клюва и эффективности кормления клестов на шишках разных видов хвойных, но и (2) эффективности кормления с приспособленностью особей, через дифференциальную выживаемость индивидуально меченых птиц, что и означает давление дивергентного отбора, соответствующее «пикам» и «долинам» адаптивного ландшафта (рис.3 статьи). Автор показывает, что (1) и (2) сильней выражены у более специализированных экотипов (из 10 выявленных в США и 7 — в Западной Европе таких только часть).
Ещё интересней, что отбор «подгоняет» клюв к точному соответствию шишкам определённого вида хвойных только в отсутствие красной (бурундуковой) белки и других белок, выступающих конкурентами. Их высокая численность «мешает» клесту эволюционировать: белка ест семена, отгрызая чешуйки, т.е. ей не надо по-разному приспосабливаться к каждому виду хвойных. Клесты Южных Холмов продвинулись в сторону узкой специализации дальше других экотипов благодаря тому, что за примерно 5000 лет обособленного существования данной популяции (данные ДНК-генеалогии, строившейся сперва по полиморфизму длин амплифицированных фрагментов генома (AFLP), затем по сходству/различию SNP, полученным в ходе полномасштабного секвенирования геномов разных форм, и показавшем значительно большую дивергенцию ДНК данной формы от прочих), белки отсутствовали. Поэтому когда ей решили присвоить видовой статус, назвали sinesciurus, лат. «без белки».
Второй ряд дифференциации форм создан поведенческими механизмами, обеспечивающим различение «своих» и «чужих» особей и, благодаря ему, поддерживающих гетерофобию — объединение со «своими» и отделения от «чужих», например в ситуации контакта и перемешивпания стай, скажем при встречах с кормлением на соседних (да даже и на одном дереве). Они используют различия в акустической форме контактных криков, полётных позывов, щебечущих слогов в песне самцов и некоторых других вокализациях; исходя из них выделяются «племена», соответствующие экотипам. Работы Craig Benkman, Kendra Sewall и других авторов (их я переведу далее) показана безусловная эффективность таких механизмов аффилиации-дифференциации (МАД), поддерживающих изолированность «племён» вопреки достаточно сильному притоку со стороны.
Эти позывы клестов, как и прочих вьюрковых, не врождённые, но выучиваются от взрослых. Было показано, что птицы в кочующей стае клестов, особенно выркастающие там молодые, сближают вокализации в сторону некого общего «образца» (общего для каждого из типов сигналов); сходный феномен отмечен в зимних стаях черноголовых гаичек. Пришельцы в общем случае отличаются по звучанию тех же типов сигналов, однако они «подгоняют» вокализации к «образцу» своего нового социума только при принадлежности к тому же самому «племени». А если они из другого, то наоборот — изменяют вокализации в сторону ещё больших отличий от этой стаи, чем увеличивают вероятность из неё выпасть, присоединившись к «своим» при первом же контакте с ними, оседании в одном месте, кормлении на одном дереве и т.д.
И наоборот, присоединившись к «своим», клёст благодаря конвергенции вокализаций, описанной K.Sewall, последовательно повышает свой шанс и дальше остаться с ними, потом размножаться в этой же стае и т.д. Такое группирование со «своими» и отделение от «чужих» не только всё больше и больше дифференцирует племена друг от друга, что видно и на коротких временных интервалах — 20 лет наблюдений показали рост разницы акустических форм позывов клестов Южных Холмов и ближайшего к ним экотипа №2, специализированного к сосне жёлтой, при отсутствии каких-либо изменений степени отличий от позывов экотипа №5, также специализированного к сосне скрученной широкохвойной (Porter, Benkman, 2019). МАД в пределе способны вести к обособлению подвидов и даже видов; однако важнее, что в каждый момент времени, здесь и сейчас, они увеличивают приспособленность стай, составленных птицами одного «племени» по сравнению со смешанными.
Для определения качества кормового дерева (стоит его покинуть или можно кормиться дальше) клесты используют как свою собственную скорость потребления семян [из тамошних шишек], так и аналогичную скорость поступления корма у сочленов по стае, т. е. социальную информацию. Однако ориентация на неё повышает эффективность кормления лишь если все клесты в стаи обрабатывают шишки и достают семена примерно одинаково (т. е., они одного экотипа, имеют в среднем одну форму клюва и т.д.). И наоборот: эффективность кормления снижена, когда стая смешанная: птицы кормятся разными способами и с разной скоростью, и социальная информация от них дезориентирует. В такой ситуации МАД, дифференцирующий «племена» на основе социального обучения с различением «своих» и «чужих», ведущий к ассортативности образования стай, выгоден всём экотипам. Действительно, проведённые в прошлом полевые опыты по проигрыванию стаям клестов позывов симпатрических экотипов показали, что они оседали тем вероятней, чем меньше были различия между формой их собственных клювов и клювов тех экотипов, чей крик звучал (Porter, Benkman, 2019).
Следовательно (добавляю уже от себя), обособление «племён» усиливает дивергентный отбор, создающий разделение экотипов. Больше того, эти исследования клестов (и другие, цитируемые в обсуждении ниже) ясно показывают неспособность дивергентного отбора дойти до конца, «разорвав» колокол изменчивости соответствующих признаков надвое, натрое и т.д., без сопровождения МАД, обеспечивающими обособление данных отдельностей (экотипов, рас, подвидов и пр. форм) как коммуникативных сообществ, и основанных на использовании каких-то сигналов, различающих — а потом раздеделяющих — «своих» и «чужих». У животных это, понятное дело, этологические механизмы, у растений — химические сигналы, образующие фитогенное поле, поведенческим изменениям соответствуют изменения роста и т.д.
Так или иначе, благодаря им эволюция (превращении внутригрупповой изменчивости в межгрупповую с появлением нового, включающего а) приспособление, выработку адаптаций отбором, и б) формообразование, обособление групп, маркированных данными адаптациями) в той или иной степени контролируется и управляется самими особями, оценивающими друг друга в рамках каких-то важных взаимодействий, скажем образования пар, где действует половой отбор, или как у клестов, в случае социальной аффилиации, где работает отбор социальный. Больше того, только это оценивание сопрягает оба аспекта микроэволюции (адаптацию и обособление внутривидовых форм, вплоть до появления новых видов) в общий процесс изменений, и без этого сопряжения ни один не дойдёт до конца.
***
Клест-еловик (Loxia curvirostra complex) — идеальная модель для количественной оценки связей между признаками фенотипа [особенно клюва, которым он добывает семена хвойных], «работой» признаков в соответствующем поведении и приспособленностью, с использованием полученных цифр для построения адаптивного ландшафта по Стьюэллу Райту [см.один из примеров. Прим.публикатора]. Во-первых, клесты демонстрируют четкую и прямую связь морфоэкологии с пищевыми ресурсами (Newton 1972; Benkman 1987a,b, 1993, 1999; Benkman and Miller 1996; Benkman et al. 2001). Это решающий момент для понимания, почему [и, главное, как] фенотипическая изменчивость ведет к различиям в приспособленности (Arnold 1983; Schluter 2000).
Клесты сильно специализированы к добыче семян из шишек разных видов хвойных, эффективность кормления определяется морфологией клюва и обрабатываемой им шишки [и, как дальше увидим, точностью соответствия между ними для каждой из дифференцировавшихся форм (экотипов, или «племëн») еловика и предпочитаемых ими видов хвойных] (Newton 1972; Benkman 1987a,b, 1993, 1999; Benkman and Miller 1996; Benkman et al. 2001). Клест использует скрещенные над- и подклювье для разрезания черепицеобразно лежащих чешуй шишки; затем они раздвигаются отводящим движением подклювье, открывая семена, лежащие у их основания (Tordoff 1954; Newton 1972; Benkman 1987a; Benkman and Lindholm 1991). Высота клюва, признак с высокой наследуемостью (h2 = 0.7; R. Summers, перс. сообщ.), влияет на время, затрачиваемое на извлечение семян (Tordoff 1954; Newton 1972; Benkman 1987a; Benkman and Lindholm 1991).
Ширина боковых бороздок на небе связана с временем обработки семян в клюве (Benkman, 1993). Клесты языком закрепляют семечко в боковой бороздке на роговом небе (на стороне, противоположной той, которую пересекает нижняя челюсть), затем [сжав клюв] отшелушивают оболочку семени и глотают ядро (Benkman 1993, 1999; Benkman and Miller 1996; Benkman et al. 2001; Parchman and Benkman 2002). [За короткий зимний день нужно набрать максимум семечек и в желудок и в подъязычный мешок — что птенцам, что на долгую зимнюю ночь. Поэтому] кормовое поведение влияет на выбор клестами местообитаний, использование [разных видов] хвойных, перемещения внутри и размножение [особенно в части мест и сроков] (Benkman 1987b, 1989, 1990, 1992). Во-вторых, адаптивная радиация у клестов вещь недавняя (Questiau et al., 1999; Arnaiz-Villena et al., 2001; Piertney et al., 2001), дифференциация форм продолжается и сегодня
[отчасти поэтому не у всех клестовых «племён» видна чёткая специализация к конкретному виду хвойных, см.краткую русскую выжимку из англоязычной сводки по 10 «племенам» в Северной Америке (в Европе их 7): некоторые используют широкий набор видов той местности, где обитают. Две другие причины обсуждаются ниже.
Первая — поведенческие механизмы обособления форм, превращающие экотип в «племя», обосабливают стаю/группу стай некоторой местности как коммуникативное сообщество и в отсутствие специализации, когда их нельзя назвать экотипом, см. их экспериментальные исследования (Sewall, 2009, 2011; Sewall, Hahn, 2009). Определяемая данным механизмом гетерофобия — распознавание «своих» и «чужих», объединение с первыми, отвержение вторых — работает независимо от экологии. Вторая — красная белка Tamiasciurus hudsonicus, «мешающая» клесту эволюционировать. Прим.публикатора].
Самка кассийского клеста с шишкой сосны скрученной
Она же добывает семя


До недавнего времени фенотип не мог быть связан с приспособленностью, потому что автор не мог найти постоянную популяцию клестов, где можно было бы измерить и пометить особей.
Еловики Северной Америки делятся на десять форм, распознаваемых по чётко различным вокализациям (Groth 1993; Benkman 1999). В Западной Европе выделено 7 форм, в нашей стране их выделение только начато (Редькин и др., 2023).
Минимум 7 из 10 североамериканских специализированы к обработке шишек разных видов хвойных — тех, что всю зиму хранят семена частично закрытыми (Benkman 1993, 1999; Benkman and Miller 1996; Benkman et al. 2001; Parchman and Benkman 2002). Между формами (или популяциями), частично разошедшимися в последние 10000 лет, развивается репродуктивная изоляция. Авторские исследования в Южных Холмах (South Hills, штат Айдахо, горный хребет на юге штата, вдоль границ с Невадой и Ютой) показывают ассортативность скрещиваний разных форм в природе.
[В Шотландии ассортативно скрещиваются три симпатричные формы — еловик, сосновик Loxia pуtyopsittacus и шотландский клёст L.scotica (двух последних не всегда выделяют как отдельные виды). У британских клестов также выявлены различия позывов у птиц с разным размером клюва, при ассортативности скрещиваний между ними (Паевский, 2018). Прим.публикатора]
Из 163 пар, размножавшихся там в 2001 году, 161 была образована «со своими»: 157 пар местной формы [тип криков 9; сейчас им присвоили видовое название Loxia sinesciurus, русск. кассийский клёст, по имени графства], 4 пары формы, специализированной на сосне жёлтой или орегонской Pinus рonderosa1 (тип 2) и две гибридные [Точный тест Фишера, P<0,0001; J.W. Smith, неопубликованный. данные]). Предварительные сравнения форм по разнообразию сайтов рестрикции (AFLP, использованы 3 пары праймеров) также показывают их расхождение. 42 маркера, использованные в AFLP, у клестов Южных Холмов присутствуют с частотой от 20% до более чем 50% (n=11), но отсутствуют у [генеалогически] самых близких к ним клестов экотипа жёлтой сосны (n=13 птиц; и наоборот) (Benkman et al., 2001).
До недавнего времени фенотипы клестов не получалось связать с приспособленностью: не находилось постоянной популяции, где можно было индивидуально пометить птиц, померить их клювы и затем выживаемость, в норме клестовые стаи широко кочуют по ареалу, а надо их отслеживать длительное время. Другая проблема, обычная для эволюционных исследований: как именно «работа» органа [скажем, кормодобывание разными клювами] обеспечивает большую или меньшую приспособленность разных вариантов биологической формы [и у клестов, где последние чётко дифференцированы, это тоже почти не изучено]?
На счастье, в 1997 г. автор нашёл оседлую изолированную популяцию еловиков, коэволюционирующую с сосной скрученной широкохвойной2 (Pinus contorta ssp. latifolia). Эти клесты живут в сосняках площадью 100 км2 на Южных холмах и прилегающих к ним горах Альбион, графство Cassia (Benkman 1999; Benkman et al. 2001) [в каталоге «племён» они №9 — самая крупноклювая и специализированная форма. Прим.публикатора]. Там можно использовать метод повторного отлова меченых птиц для определения выживаемости и по ней приспособленности.

Сосна скрученная широкохвойная и леса из неё, в которых живёт эта форма
 Клесты Южных холмов обладают уникальными вокализациями (Benkman, 1999); были зарегистрированы только в сосняках из сосны скрученной на этих двух изолированных горных хребтах. В 1998 году автор начал ловить, измерять и описывать цвет оперения взрослых клестов Южных Холмов.
Клесты Южных холмов обладают уникальными вокализациями (Benkman, 1999); были зарегистрированы только в сосняках из сосны скрученной на этих двух изолированных горных хребтах. В 1998 году автор начал ловить, измерять и описывать цвет оперения взрослых клестов Южных Холмов.
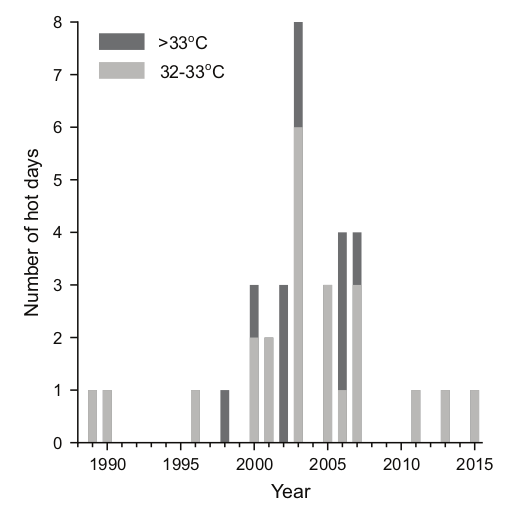
[Сейчас ситуация изменилась: после масштабных пожаров 2020 г. впервые отмечено их гнездование в штате Колорадо, видимо, катастрофа среды обитания, как часто бывает, стимулировала дальний залёт. Вообще учащение лесных пожаров, обусловленное глобальным потеплением, грозит уничтожить совсем кассийских клестов, тем более что они малочисленны — исходно не более 6000 птиц.
Рост числа аномально жарких (32-33 и >33 градусов Цельсия) дней в ареале клеста Южных Холмов
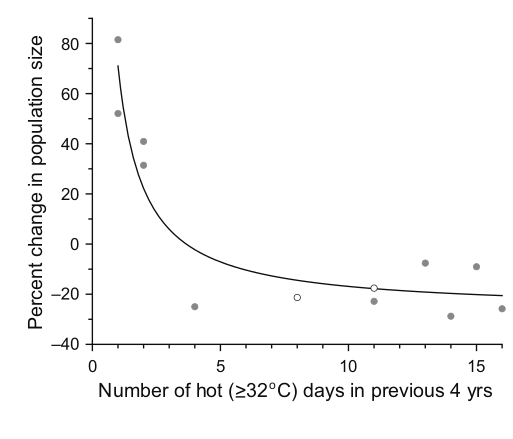
Снижение численности популяций клестов Южных Холмов (в %) с ростом числа аномально жарких (32 и более градусов Цельсия) в предшествующие 4 годы. Эта и предыдущая илл. из: Benkman, 2016.
Прим.публикатора]
Дальше по данным долговременного отлова и мечения автор установил две зависимости: (1) между высотой клюва и выживаемостью, (2) между высотой клюва и эффективностью кормления, используя логистическую регрессию (Janzen and Stern, 1998). Их форма оказывается очень похожей, из чего следует
(а) эффективность кормления детерминирует выживаемость,
(б) отбор «работает» с высотой клюва, приспосабливая его [у разных форм еловика] к обработке конструктивно различных шишек хвойных. Всё это хорошо согласуется с имеющимся анализом эффективности кормодобывания (Benkman, 1993). Опираясь на эти данные, автор реконструировал
(в) адаптивный ландшафт в смысле С. Райта, связывающий разные «конструкции» клюва (ось Х — высота, У — ширина клюва, задаёт ширину бороздок рогового неба) с приспособленностью (ось Z) для 5 экотипов еловиков запада Северной Америки (Fear and Price, 1998).
[Грустно, что, активно рассказывая про «племена» клестов, известный орнитолог и палеонтолог К.М.Михайлов опускает в них всё, что касается специализации, микроэволюции и естественного отбора (притом что это важнейшая часть метода и результатов Крэйга Бенкмана!), чтобы «уложить» реальность в прокрустово ложе собственных взглядов, оказывающихся ему важней эмпирических данных, чем необратимо искажает последние. Это не есть хорошо. Прим.публикатора]
Использовать выживаемость как суррогат приспособленности возможно, поскольку у птиц и многих других таксонов первая лучше всего репрезентирует вторую, нежели прочие демографические показатели (Crone 2001). Перекрывание ареалов разных экотипов, их частичная симпатия делает частотно-зависимый отбор наиболее вероятным, если не неизбежным; поэтому пики адаптивного ландшафта не обязательно соответствуют эволюционному равновесию (Fear and Price, 1998).
Однако при построении адаптивного ландшафта, который связывает приспособленность разных особей в популяции с их фенотипом, есть одна трудность (Fear and Price 1998; Schluter 2000). Его пики отражают устойчивую / равновесную ситуацию только в случае, если частотно-зависимый отбор не приводит к значительному отклонению от пика.
Методы исследования
Содержание
Повторный отлов маркированных клестов
Авторы ловили клестов паутинными сетями и на манную птицу в 2х местах Южных Холмов. В 1998 и 2000 гг. пятьдесят шесть [пойманных так птиц] индивидуально помечены. Цифровым штангенциркулем им измерили с точностью до 0,01 мм 1) высоту клюва у переднего края ноздри; 2) длину надклювья от кончика клюва до переднего конца правой ноздри; 3) длину подклювья от кончика нижней челюсти до пересечения её ветвей. Также линейкой измеряли длину сложенного крыла с точностью до 0,5 мм, массу тела — весами Песола с точностью до 0,5 г (в 1998 году) и цифровыми весами с точностью до 0,01 г (в 1999-2001 годах). Клюв измеряли два разных человека (CWB во все годы и W. C. Holimon в 1998 году), получая почти идентичные и хорошо воспроизводимые результаты. Каждый из них мерил высоту клюва 3-5 раз у каждой птицы и дальше использовали среднее. Например, из 17 клестов Южных Холмов, которых независимо измерили с интервалом примерно в месяц, средняя разница высоты клюва составила 0,001 мм (повторяемость = 0,992, F=245,1; P<0,001) (Lessells and Boag, 1987).
После выпуска полётные крики птиц были записаны с помощью микрофонов Sennheiser ME66 (Old Lyme, Коннектикут) и магнитофонов Marantz PMD222 (Аurorа, Иллинойс). Записи анализировали программой Canary (Корнелльская орнитологическая лаборатория, Итака, Нью-Йорк) на компьютере Macintosh, чтобы подтвердить принадлежность криков каждого меченого клеста к определённому типу. В 2000 и 2001 г. авторы искали и повторно отлавливали помеченых клестов в 2000 и 2001 годах и обнаружили 20 (35,7%) из них через один-три года после первой поимки.
Интенсивность отбора определяли, используя логистическую регрессию (Janzen and Stern 1998) в предположении, что повторно отмеченные птицы выжили, ненайденные нет. [В дальнейшем анализе] использовали кубический корень из массы тела и логарифмы средних значений других показателей. Наибольшая корреляция между признаками составила r=0,52 (между длинами над- и подклювья). В работе представлен коэффициенты логистической регрессии (α). Дальше данные интерполировали кубическим полиномом (Schluter, 1988) для определения формы отбора. Здесь использовали данные по всем 56 клестам, но в логистической регрессии — лишь 5 (не все признаки смогли померить для всех птиц).
Эффективность кормления
Авторы рассчитали время кормления 28 содержащихся в неволе клестов Южных холмов, кормящихся местной сосной, чтобы выяснить, действительно ли однотипны связи между высотой клюва и эффективностью кормления, с одной стороны, и высотой клюва и выживаемостью, с другой.
Методы измерения эффективности кормления опубликованы (Benkman 1993; Benkman and Miller 1996; Benkman et al. 2001) и кратко изложены ниже. Пойманных клестов держали в закрытых вольерах (1,6-32,7-32,2 м или больше). По крайней мере несколько дней до и во время экспериментов птиц держали впроголодь, с низкой массой тела, давая кормиться в основном семенами из шишек сосны скрученной широкохвойной.
Чтобы обеспечить постоянную и высокую мотивацию к кормодобыванию, всякую пищу убирали за ~15 часов до опыта. Для каждой птицы фиксировали общее время, нужное для извлечения и шелушения (удаление оболочки и проглатывание ядра) 10 семян, начиная с момента съедения первого семени; и так при питании на 10-11 шишках этой сосны. Также определили время лущения семян, вычли его средние значения, чтобы получить среднее время извлечения семян для каждой птицы. В анализе использовали время извлечения семян, поскольку именно она, но не время лущения тесно связано с высотой клюва (Benkman, 1993).
Эффективность кормления оценивали как 1/(время извлечения семян из шишек разных хвойных * предполагаемую суточную потребность в энергии) (Benkman 1993; Benkman and Miller 1996; Benkman et al. 2001). Последняя оценена как функция массы тела М, Е=13,05*М0,6052(Walsberg 1983).
Масса тела зависит от многих факторов, включая количество накопленного жира и, особенно в неволе, потерю мышечной массы в полете. Её оценивали по аллометрическому соотношению между массой тела (Y, в граммах) и высотой клюва (X, в мм): Y= -3,78+3,82X (r2=0.68, df = 77; P <0, 0001), где массу измеряли на пойманных наименее жирных клестах. Высоту клюва, наиболее эффективную в удовлетворении суточных потребностей в энергии (т.е. оптимальную высоту), определили так такую, которая минимизирует время кормления, необходимое для удовлетворения суточных энергопотребностей (1993; Benkman and Miller 1996; Benkman et al. 2001).
Установление «рельефа» адаптивного ландшафта
Эффективность кормления клестов семенами разных хвойных определяли, как описано выше, скармливая шишки этих видов птицам с разной комбинацией высоты клюва и ширины небных бороздок. Единственно, время кормодобывания здесь брали как сумму времен извлечения семян из шишек и их лущения. Для анализа выбраны 5 видов хвойных, ежегодно дающих урожай шишек по всему ареалу или в какой-то его части, и долговременно сохраняющих семена в закрытых шишках: оба фактора благоприятствуют специализации [к питанию семенами этих видов] (Benkman 1993; Thompson 1994). 4 из 5 широко распространены на северо-западе США или в северной части Скалистых гор, к ним специализированы четыре обычных и широко распространенных формы еловиков, населяющих запад Северной Америки (Benkman, 1993).
Пятый вид хвойное дерево — сосна скрученная широкохвойная, растущая на Южных Холмах (Benkman 1999; Benkman et al. 2001). Автор использовал уже опубликованные уравнения (Benkman, 1993) в определении времени извлечения из шишки и шелушения семян западной тсуги (Tsuga heterophylla) и псевдотсуги (дугласовой пихты, Pseudotsuga menziesii ssp. menziesii), а также 2х видов сосен, сосны жёлтой Скалистых гор (Pinus ponderosa ssp. scopulorum) и сосны скрученной широкохвойной. Поскольку при шелушении семян желтой сосны клесты иногда теряют ядрышко, оценки затрат времени корректировались: среднее время обработки одного семени делили на число ядер, потреблëнных на одно очищенное семечко, получилось 475,28–455,20*(ширина нёбных бороздок)-110,27*(ширина нёбных бороздок)2, r2=0,81, df=10, p<0,0002). Использованы уже опубликованные уравнения (Benkman and Miller 1996; Benkman et al. 2001) для оценки времени извлечения семян сосны Скалистых гор, а также для оценки времени извлечения семян из шишек жёлтой сосны и сосны скрученной широколистной.
У автора не было данных о времени лущения семян в зависимости от ширины небных борозд для сосны скрученной Южных Холмов; однако его зависимость от ширины небной бороздки одинакова для разных типов семян (Benkman, 1993). Поэтому [в его определении] пользовались уравнением для лущения семян сосны Скалистых гор (Benkman, 1993), скорректированное так, чтобы оптимальная ширина бороздки соответствовала прогнозируемой (оптимальная ширина бороздки 1,28+0,21*(масса семян)-3, r2=0,998, df=2, P< 0,002) исходя из средней массы семян этого вида, на 7,01 мг (Benkman et al., 2001). Уравнение для времени извлечения семян из шишек псевдотсуги скорректировали так, чтобы оптимум соответствовал средней высоте клюва потребляющих их еловиков (8,75 мм), поскольку их средняя ширина нёбных борозд идентична оптимальной для лущения именно этих семян, что подразумевает адаптацию к псевдотсуге (Benkman, 1993). Наконец, эффективность кормления преобразовали в приспособленность на основе степенной зависимости, связывающей оба параметра (см. рис.2). Абсолютную высоту пиков адаптивного ландшафта не определить, поэтому автор использовал относительную, доли единицы.
Таблица 1. Логистическая регрессия, показывающая связь разных морфологических признаков клестов Южных Холмов в выживаемость (т.е. их адаптивную значимость, n=50 птиц). Общая модель значима, χ2 =31,1, P<0.0006). Представлен коэффициент регрессии α± SE, X2 и его значимость. Признаки сверху вниз: высота клюва мм, её квадрат, длина надклювья мм, её квадрат, домна подклювье мм, её квадрат, длина крыла, масса тела, г., её квадрат.
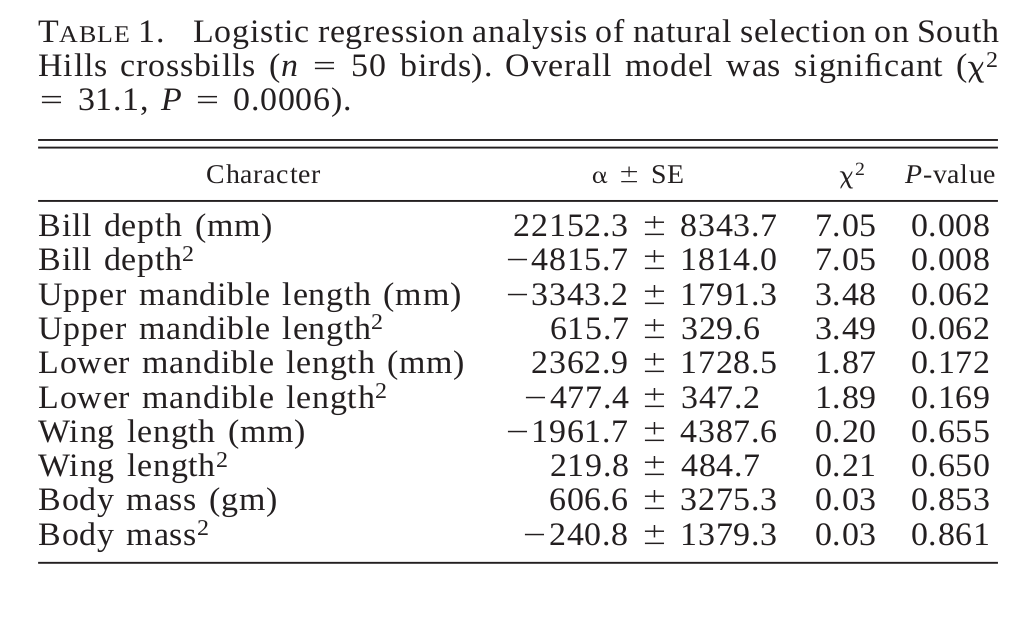
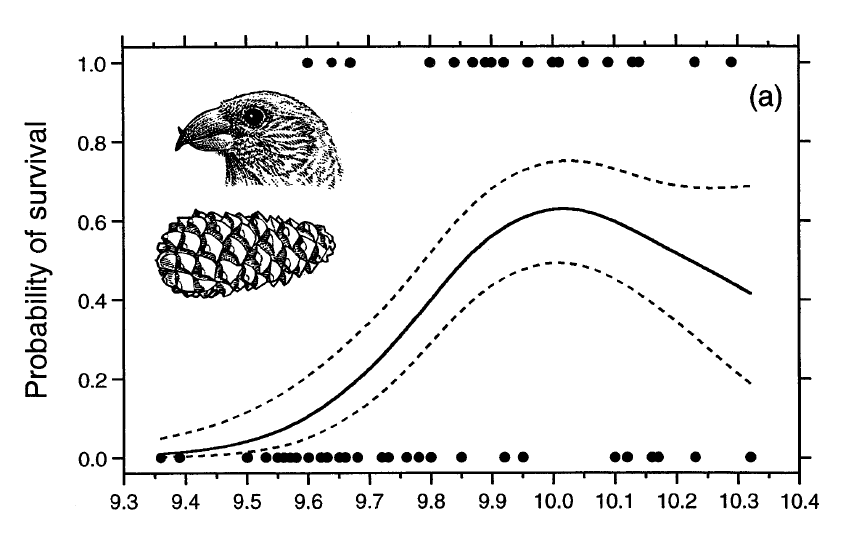
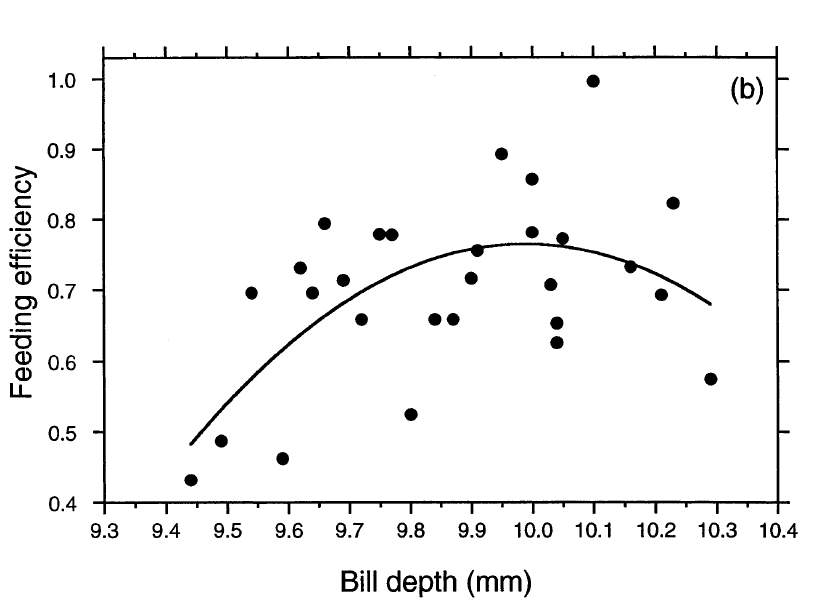 Рисунок 1. Относительная выживаемость взрослых клестов Южных Холмов на протяжении хотя бы года (а) и эффективность питания семенами местной сосны (b) в зависимости от высоты клюва. Обозначения. (а) Сплошная кривая — аппроксимация кубическим многочленом, пунктир — стандартная ошибка, ±SE. Сплошные кружки сверху графика — число индивидуально меченых птиц, пойманных или наблюдавшихся в природе через год, два или три после отлова взрослыми (n=20), они же внизу — число не отмеченых после поимки (n=36). Также изображена типичная шишка сосны Южных Холмов. (b) Кривая — аппроксимация квадратным трехчленом, эффективность кормления = -92,94+18,76*(высота клюва)-0,94*(высота клюва)2, r2=0.35, df=25, P<0.005, P=0.020 для квадратичного значения). Кубический многочлен дал почти идентичную кривую. Каждая чёрная точка — средняя эффективность кормления для одной особи, извлекавшей по 10 семян из каждой из 10-11 шишек.
Рисунок 1. Относительная выживаемость взрослых клестов Южных Холмов на протяжении хотя бы года (а) и эффективность питания семенами местной сосны (b) в зависимости от высоты клюва. Обозначения. (а) Сплошная кривая — аппроксимация кубическим многочленом, пунктир — стандартная ошибка, ±SE. Сплошные кружки сверху графика — число индивидуально меченых птиц, пойманных или наблюдавшихся в природе через год, два или три после отлова взрослыми (n=20), они же внизу — число не отмеченых после поимки (n=36). Также изображена типичная шишка сосны Южных Холмов. (b) Кривая — аппроксимация квадратным трехчленом, эффективность кормления = -92,94+18,76*(высота клюва)-0,94*(высота клюва)2, r2=0.35, df=25, P<0.005, P=0.020 для квадратичного значения). Кубический многочлен дал почти идентичную кривую. Каждая чёрная точка — средняя эффективность кормления для одной особи, извлекавшей по 10 семян из каждой из 10-11 шишек.
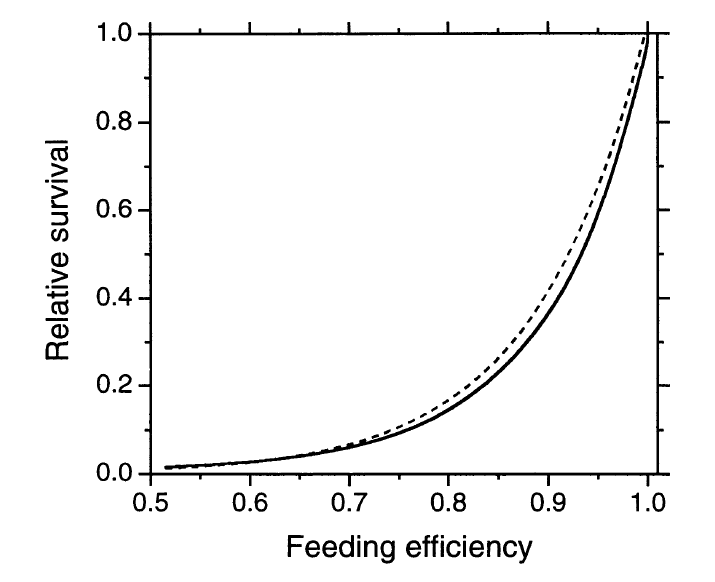 Рисунок 2. Расчетная относительная выживаемость в зависимости от расчетной относительной эффективности кормления (сплошная линия) и регрессия, используемая для аппроксимации, пунктир; Выживаемость = е[-9.032+9,057*эффективность кормления] (рис.3). Примечание. Мы ограничили анализ высотой клюва <10,00 мм: клещи Knemidokoptes jamaicensis, обитающие в коже ног, грызя её, поднимают чешуйки, чем препятствуют кольцеванию. Они чаще бывают на крупноклювых клестах, поэтому оценки выживаемости точнее для более мелких птиц (см. доверительные пределы на рис.1а).
Рисунок 2. Расчетная относительная выживаемость в зависимости от расчетной относительной эффективности кормления (сплошная линия) и регрессия, используемая для аппроксимации, пунктир; Выживаемость = е[-9.032+9,057*эффективность кормления] (рис.3). Примечание. Мы ограничили анализ высотой клюва <10,00 мм: клещи Knemidokoptes jamaicensis, обитающие в коже ног, грызя её, поднимают чешуйки, чем препятствуют кольцеванию. Они чаще бывают на крупноклювых клестах, поэтому оценки выживаемости точнее для более мелких птиц (см. доверительные пределы на рис.1а).
Результаты
Логистическая регрессия включила все признаки; наибольший из значимых коэффициентов был при высоте клюва, почему её можно считать главной мишенью отбора (таблица 1). Ею может быть также длина надклювья (таблица 1): в этом случае отбор может быть дизруптивным или как минимум больше благоприятствующим более длинноклювым особям. Однако влияние этого признака на выживаемость не было значимо, когда он был единственным в уравнении регрессии (P=0,40 как для линейного, так и для квадратичного коэффициентов). Напротив, стабилизирующий отбор по высоте клюва значим (P<0,05) независимо от включения в анализ других признаков. [Поэтому скорее всего, влияние длины надклювья на выживаемость — и, соответственно, селективное давление на неё — осуществлялись по корреляциям].
На Южных Холмах сосняки из сосны скрученной в массе распространились около 5000-7000 лет назад. Вслед за ними здесь обосновались клесты и стали увеличивать высоту клюва для лучшего открывания шишек. В ответ их чешуйки утолстились, что вызвало следующее увеличение клюва и т. д. Коэволюция клестов и местных сосен продолжалась, пока не был достигнут пик адаптивного ландшафта: этому способствовало отсутствие на Южных холмах красных белок Tamiasciurus hudsonicus. Поэтому название формы sinesciurus означает по-латыни «без белки».
Специализация клювов разных «племён» к добыванию семян конкретных видов хвойных идёт лишь пока низка численность белки, то есть при не очень больших урожаях. Когда урожай хорош, белки быстро увеличивают численность, и успешно конкурируют с клестом. Они настолько успешно выедают «наиболее типичные» шишки хвойных каждого вида, что клесты вынуждены изменить направление своей адаптации. Вместо потребления «наиболее типичных» шишек каждого из хвойных (ведущего к внутривидовой дифференциации форм по клюву и, в той мере, в какой стаи константны по составу, по поведению), они вынужденно переключаются на использование шишек промежуточного размера и содержания семян. То есть, при возрастании численности белки вектор специализации у клестов сдвигается в противоположную сторону «объединения» морф, выделившихся на прошлом этапе, при падении – вновь идёт дифференциация по видам шишек и пр.

Поэтому сообитание с белкой лишает клестов значительной части семян и эффективно блокирует специализацию на отдельных видов хвойных. Возможно, обособление местной формы произошло в том числе генетически, дошло как минимум до подвидового (а может, и видового) уровня, именно благодаря отсутствию конкурента. В отличие от клестов, белки полностью сгрызают чешуйки, оставляя лишь стерженёк, поэтому им не надо не надо по-разному приспосабливаться к каждому виду. Плюс в отличие от сильно специализированного клеста они потребляют массу других сопутствующих кормов, так что в каждой из местностей их численность куда меньше зависит от урожая семян, чем клестовая.
Визуализация выборки по высоте клюва дополнительно подтверждает колоколообразную зависимость выживаемости с максимумом при высоте клюва 10,01 мм (рис.1а). Это близко к оптимальной высоте клюва 9,99 мм, рассчитанной по зависимости между ней и эффективностью кормодобывания (рис.1б). Хотя оба значения почти совпали (рис. 1), при отклонении высоты клюва от оптимума выживаемость падает быстрей эффективности кормления, поскольку здесь зависимость экспоненциальная (рис.2).
Автор ее использовал, чтобы преобразовать эффективность кормления пяти видами хвойных северо-запада США в приспособленность. Получается адаптивный ландшафт с пиками, соответствующими специализации какого-то «племени» клестов на одном из хвойных, и долинами (рис.3).
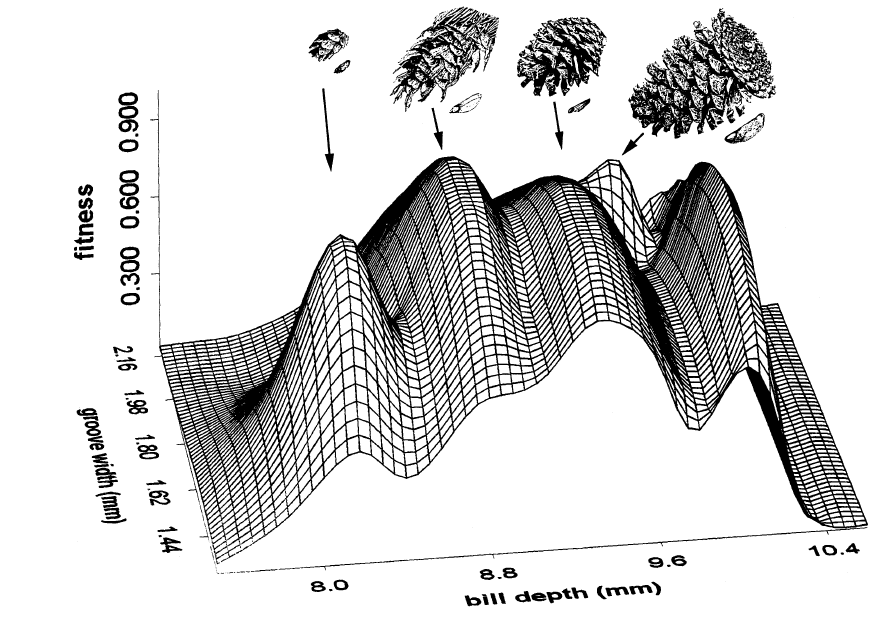 Рисунок 3. Адаптивный ландшафт для клювов клестов-еловиков, специализированных к 5 видам хвойных: слева направо — тсуга западная, псевдотсуга Мензиса, сосна скрученная широкохвойная, сосна жëлтая, сосна скрученная скалистых гор. Показаны шишки и крылатые семена первых 4х. Ось Х — высота клюва, У — ширина нёбных бороздок, Z — приспособленность, рассчитанная по выживаемости на основании её зависимости от эффективности кормления (рис. 2).
Рисунок 3. Адаптивный ландшафт для клювов клестов-еловиков, специализированных к 5 видам хвойных: слева направо — тсуга западная, псевдотсуга Мензиса, сосна скрученная широкохвойная, сосна жëлтая, сосна скрученная скалистых гор. Показаны шишки и крылатые семена первых 4х. Ось Х — высота клюва, У — ширина нёбных бороздок, Z — приспособленность, рассчитанная по выживаемости на основании её зависимости от эффективности кормления (рис. 2).
Обсуждение
Прошлые результаты автора показывают, что высота клюва, дополненная шириной небной бороздки — главные признаки, влияющие на скорость кормления (Benkman, 1993). Поэтому они должны быть целью отбора, если скорость кормления влияет на физическое состояние птицы. Что подтвердилось в анализе селективной значимости разных признаков (табл. 1).
Однако длина надклювья связана с выживаемостью как минимум слабо. Неслучайно в лаборатории не выявлена её со скоростью потребления семян сосны скрученной широкохвойной: в многофакторной регрессии [Lande and Arnold 1983], включающей высоту клюва с длинами над- и подклювья, коэффициенты для длины надклювья и его квадрата не значимы (P=0,24 и 0,26), тогда как высоты клюва и её квадрата высоко значимы (P=0,002 и 0,003). Почти значимый коэффициент регрессии для квадрата длины надклювья у клестов Южных Холмов (таблица 1) показывает рост выживаемости у более длинноклювых птиц. Причины такой предпочтительности неясны, особенно на фоне предпочтения более короткоклювых птиц при образовании пар в популяции Южных холмов (Benkman et al., 2001).
Возможное объяснение в том что это побочное следствие меньшей изношенности рогового чехла челюстей у птиц, лучше извлекаюших семена. Он же постоянно растёт: хуже извлекающие (дольше возящиеся) изнашивают сильнее чехол сильнее, так что у них клюв короче. Т.е. это модификационные изменения, созданные внешней средой, а не отбор наследуемых признаков.
Сходство зависимостей эффективности кормления с одной стороны, выживаемости, с другой, от высоты клюва (рис.1) означает, что фактором отбора по высоте клюва выступает морфология шишек, на чьих семенах птицы специализируются. Более того, нелинейный характер зависимости между выживаемостью и эффективностью кормления (рис. 2) заставляет считать, что даже небольшое снижение последней (и скорости поступления корма за день) сильно и негативно влияет на физическое состояние существенные последствия для физической формы. Это подтверждает фундаментальное предположение теории оптимального поиска пищи (Stephens and Krebs, 1986), помогая понять, почему многие аспекты поведения и экологии клестов кажутся связаны с кормодобыванием и максимизируют поступление корма (Benkman, 1989, 1990, 1992).
Нелинейная связь выживаемости и «работы» клестиных клювов определённой формы делает поверхность приспособленности (рис.3) намного сильнее изрезанной, чем ландшафт, построенный только на эффективности «работы» клюва. Всё это усиливает дивергентный отбор экотипов, специализированных к добыче семян из шишек разных хвойных, т. е. ускоряет «движение» каждого из них к вершинам адаптивного ландшафта (Fear and Price, 1998; Schluter, 2000).
Дивергентный отбор при специализации к использования альтернативных ресурсов — ключевое объяснение процессов адаптивной радиации (Schluter, 2000). Оно подтверждается этим исследованием и еще одним (Schluter and Grant 1984), в котором была дана количественная оценка крутизны адаптивного ландшафта, отражающей направление и темпы радиации форм (Grant, 1999). Действительно, средние высоты клюва каждого из 5 «племён» еловка — зарождающихся видов — приурочены к пикам ландшафта или находятся очень близко к ним (Benkman, 1993; Benkman and Miller 1996; Benkman et al. 2001).
Это значит, что частотно-зависимый отбор не сильно влияет на эволюционное равновесие и подразумевает дивергентный отбор между формами, производимый специфическими особенностями шишек и семян разных видов. Полученный результат намного яснее, чем любое другое исследование, через «работу» органа определённой формы (клюва) в определённой обстановке (шишки различной толщины и формы чешуй) связывает фенотип с приспособленностью в дикой природе и, в конечном счете, с дифференциацией форм.
Последующие работы автора посвящены стайному поведению, развитию и использованию вокализации, обеспечивающим единство внутри каждого «племени» (в том числе предпочтительное образование пар «со своими»), одновременно обособляя их от иных «племён», а также их генетической дифференциации и изоляции друг от друга. Отдельные «племена» возникают как экотипы, специализированные к определённому виду хвойных, и обретают обособленность (делаясь закрытым коммуникативным сообществом) в силу особенностей коммуникативной системы этого вида в стаях, кочующих вслед за урожаями соответствующих хвойных. Объединяя особей одного «племени», она обладает гетерофобией, т. е. эффективно разделяет » своих» и «чужих», распределяя их в разные стаи (в которых образуются пары, почему они в основном со » своими») даже при кормлении на одной территории или на соседних деревьях.
Уже довольно давно показано, что дизруптивного (в англоязычной литературе дивергентного) отбора недостаточно, чтобы «разорвать» поддерживаемые им альтернативные варианты, нужно одновременное действие полового отбора или любой иной системы оценки самими особями фенотипов друг друга, с определением «своих» и чужих», и присоединением к первым (неважно, в общие стаи или конспецифичные пары) и отвержением «вторых» несмотря на симпатрию форм, или их синтопию и прочее тесное соприкосновение.
В Science была интересная статья, в которой показано путём компьютерного моделирования, что обычный дизруптивный отбор по признакам, влияющим на просто на приспособленность особей к среде, не способен «разорвать» единую популяцию, в которой есть широкая изменчивость этих признаков, и привести к распадению её на две и дальше к симпатическому видообразованию. Дизруптивный отбор – это отбор в пользу высоких и низких значений таких признаков, и против носителей средних значений, так что ранее единый колокол распределения делится на два, почти не перекрывающихся друг с другом. А вот если дополнить естественный отбор половым по ещё одному признаку, значения которого сами по себе на приспособленность не влияют, но влияют на выбор полового партнёра, дизруптивный отбор доходит до конца, единая популяция делится на две, и возможно симпатическое вилообразование. Там дана и математическая модель явления, см. русский пересказ А.В.Маркова.
Данный результат известен с 1955 г., но, увы, был в забвении у исследователей.
«В 1955 г. Тудей получил разделение единой популяции дрозофилы при одновременном отборе в (+) и (-) – направлениях по числу стерноплевральных щетинок. Однако ряд попыток других исследователей повторить эти опыты оказались неудачными: дизруптивный отбор вёл лишь к увеличению изменчивости признака, а при продолжительном проведении – к формированию 2-х или большего числа дискретных «морф» и их стабилизации, как у парусника Papilio dardanus стабилизированы миметические морфы. Это же касается и дизруптивного отбора по другим признакам, например, на увеличение или уменьшение массы тела. Чтобы дизруптивный отбор был эффективен, его требовалось дополнить половым (который проводился исследователями «насильственно», т.е. был искусственным: при дизруптивном отборе по массе тела мелких мух принудительно спаривали с мелкими, а крупных с крупными (Huges, 1986)».
А затем ситуация разделение исходной популяции на две почти не смешивающиеся друг с другом формы за счёт дизруптивного отбора, сопряжённого с половым, была найдена и в природе.
«Дизруптивный отбор эффективен, если идёт по признакам, ограничивающим свободное скрещивание особей, отличающихся экологическими предпочтениями, т.е. является одновременно и отбором на их изоляцию. Примером … может служить дивергенция по величине цветков у растения Polemonium viscosum. У этого вида часть растений имеет мелкие цветки с неприятным гнилостным запахом, которые опыляются мухами, а часть – крупные цветки с приятным сладким ароматом, опыляемые шмелями (вообще, так что от сладости до гнилости один шаг…). Из семян, полученных в перекрёстном опылении морф, вырастают вполне нормальные растения с промежуточным размером цветков. Однако из-за различия опылителей в природе подобного скрещивания практически не происходит, и дизруптивный отбор по величине цветков продолжается (Galen et al., 1978). Из учебника теории эволюции А.С.Северцова, стр.180.].
«Племена» у еловиков дают новый и интересный вариант механизма, обеспечивающего гетерофобию, и обеспечивающего дифференциацию форм — иногда в сопряжении с экологической специализацией, на основе экотипов, иногда без, основанного на социальном отборе и конвергенции сигналов, объединяющих особей в стаях и парах, к «типу», специфическому для каждого «племени», за счёт социального обучения. Здесь формы обособляет (в тех обстановках, где это возможно, т.е. в отсутствие белки, «не секс», а «культура»). Все названные механизмы коммуникативны по своей природе — как и механизмы внутрипопуляционной регуляции, обеспечивающие «дистанционное наведение» нерезидентов в места, где необходимо восстанавливать численность после локальных катастроф.
Если репродуктивная изоляция — результат дивергентного отбора (т.е. экологического видообразования), то диверсификация клестов обеспечивает дополнительную поддержку экологической теории адаптивной радиации (Schluter, 2000). Сигналы и механизмы коммуникации, «склеивающие» воедино членов сообщества и побуждающие их рассматривать тех как «своих», дающие всем коммуникативные средства объединения со » своими» и отделения от «чужих», выступают как «закрепители» и «усилители» дифференциации, начавшейся как экологическая, превращать разные экотипы в более или менее закрытые коммуникативные сообщества.
Источник Evolution. 2003. V.57. №5. Р.1176-1181.
Примечания
1одно из самых жароустойчивых хвойных, лесах из неё профильны.
2имеет серотиновые шишки, раскрывающиеся лишь после (и вследствие) сильных пожаров.
Рекомендуем прочесть
 Дивергентно-конвергентные механизмы в эволюции биоразнообразия
Дивергентно-конвергентные механизмы в эволюции биоразнообразия Быстрая перестройка фенологии в новых местах гнездования ласточек
Быстрая перестройка фенологии в новых местах гнездования ласточек Популяция как система: механизмы устойчивости и саморегуляции у мелких млекопитающих
Популяция как система: механизмы устойчивости и саморегуляции у мелких млекопитающих Красивые теории, отвергнутые из-за сопротивления некрасивых фактов
Красивые теории, отвергнутые из-за сопротивления некрасивых фактов Как изменение климата способствует быстрой эволюции
Как изменение климата способствует быстрой эволюции Жизнеспособность важнее специальных адаптаций: приморские формы саванной овсянки
Жизнеспособность важнее специальных адаптаций: приморские формы саванной овсянки Понятие приспособленности в структуре эволюционной теории
Понятие приспособленности в структуре эволюционной теории Гиперболы Виллиса: попытка интерпретации
Гиперболы Виллиса: попытка интерпретации
