![Print PDF Как пятна на крыльях помогают кормиться, но среднего размераСодержание1 Как пятна на крыльях помогают кормиться, но среднего размера2 P.S. публикатора: в чём смысл стабилизирующего отбора? 3 Эволюция — […]](http://www.socialcompas.com/wp-content/uploads/2025/05/Hooded_Warbler.jpg "Как пятна на крыльях помогают кормиться, и про стабилизирующий отбор")

Самец капюшонной вильсонии
Как пятна на крыльях помогают кормиться, но среднего размера
Содержание
Век живи, век учись. Не знал, что у певчих птиц из сем. Древесницевых, Parulidae, белые пятна на хвосте и/или крыльях помогают в ловле добычи, и узнал из работы Ronald R. Mumme (2023). Скажем, каёмки на внешних опахалах рулевых 4-6 у капюшонной вильсонии Setophaga citrina (рис.1): взмахивая ими, птичка движется в кроне, эти взмахи пугают насекомых, заставляют взлететь и их ловят в угон. Кормление древесницевых эксплуатирует эволюционно консервативную реакцию бегства насекомых; специальные опыты показывают, что закрашивание пятен резко снижает уловистость такого кормления, по сравнению с контролем, где закрашивание лишь изображалось (рис.1 внизу, полевые опыты с S.citrina, а с древесницами р. Myoborus проводили и лабораторные).
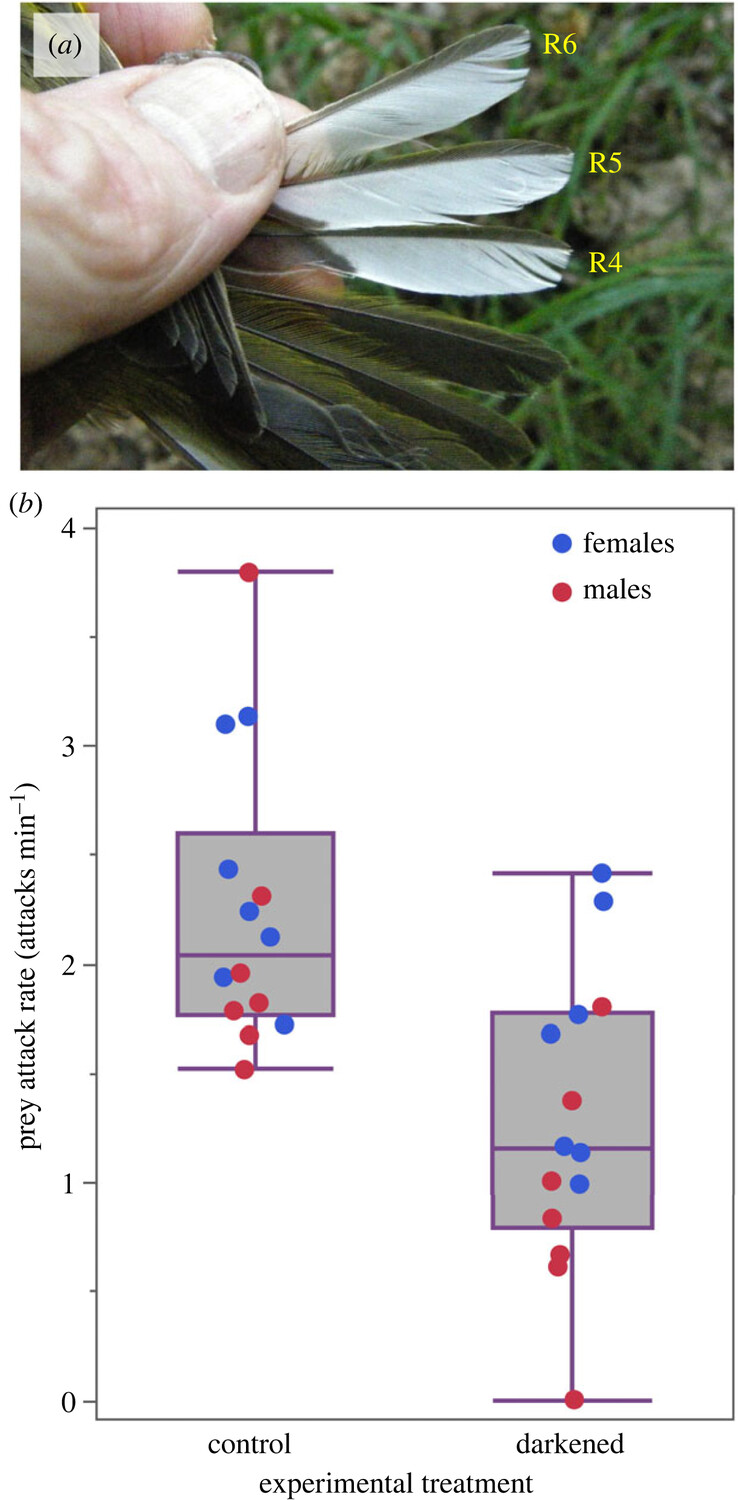
Рисунок 1. Белые каймы на рулевых капюшонных вильсоний (сверху) и снижение уловистости кормления (частота успешных схватываний жертв, в мин, ордината) при их закрашивании. Слева контроль, где закрашивание лишь изображали, справа опыт.
Это верно и для самок и для самцов, хотя у данного вида они на зимовках (т.е. большую часть года, это дальний мигрант, см.карты ареалов) используют разные местообитания: самцы сомкнутые тропические и субтропические леса, самки открытые пространства с кустарниками (скрэб), т.е. пола ландшафтно разделены. Однако взгляд на помахивание хвостом/крыльями как способ повысить уловистость кормодобывания работает не везде работает не везде, скажем, не соответствует фактам попытка объяснить взмахивание хвоста трясогузки использованием этого кормового метода1.
Степень белого на 4-6 парах рулевых у этих птиц варьирует индивидуально, у разных особей их очертания устойчиво сохраняются из года в год (повторяемость 75%) и не меняются при позднелетней линьке, что заставляет говорить о наследуемости, хотя прямо её не мерили. У данного вида это вообще трудно сделать из-за низкой степени филопатрии и очень высокой доле птенцов, происходящих от внебрачных копуляций.
И вот у капюшонной вильсонии (или древесницы, раньше её относили туда) Setophaga citrina (рис.3) на данных 14 лет наблюдений за индивидуально меченной популяцией в с.-з. Пенсильвании (Hemlock Hill) в 2010-2023 гг. показан стабилизирующий отбор по величине «ловчих» пятен. В мае-августе здесь картировали все гнездящиеся пары (в разные годы их было 45-70), искали гнезда и фиксировали репродуктивный успех, чтобы связать с ним и с физическим состоянием птиц величину белых кайм на рулях.
У птиц, пойманных для кольцевания, фиксировали домну крыла, вес, максимальную линейную длину белого на каждом из трёх опахал, а дальше считали индекс развития белого цвета, относя сумму этих трёх промеров к утроенной длине хвоста. Индекс коррелирует с развитием белого на каждом из перьев и учитывает колебания длины хвоста. У 28% самцов и 14% самок есть также белое пятнышко на третьей паре рулевых, но его размер не учитывали, ибо он не коррелирует с величиной других кайм. Для всех птиц, гнездившихся в 2010-2023 гг., и меченных цветными кольцами, посчитана долговременная выживаемость по числу лет присутствия на гнездовании (она была 0, если птица гнездилась лишь 1 сезон).
В среднем это 1,05 (0-8) для самцов (n=317) и 0,78 (0-7, n=308) для самок. Развитость белого на рулях нормально распределена у обоих полов, у самцов она выше, ненамного, но значимо (рис.2, верхняя и средняя илл.). У среднего самца каймы примерно на 2 мм длинней, чем у средней самки. Дольше живут особи с пятнами среднего размера, как самцы, так и самки (0,50-0,53, рис.2, нижняя илл.).
Миграционный путь данного вида: жёлтым — гнездовой, синим — зимовочный ареал
У самцов эффект стабилизирующего отбора выражен сильней, чем у самок, на 5й паре рулевых сильней, чем на 4й и 6й. А вот на репродуктивный успех в данный сезон или на массу птиц при размножении развитость белого не влияет. Единственный предиктор продукции слетков — число лет пребывания в гнездовой группировке (объясняет 60% изменчивости успеха размножения), массы тела — длина крыла, т. е. общий размер и пол.
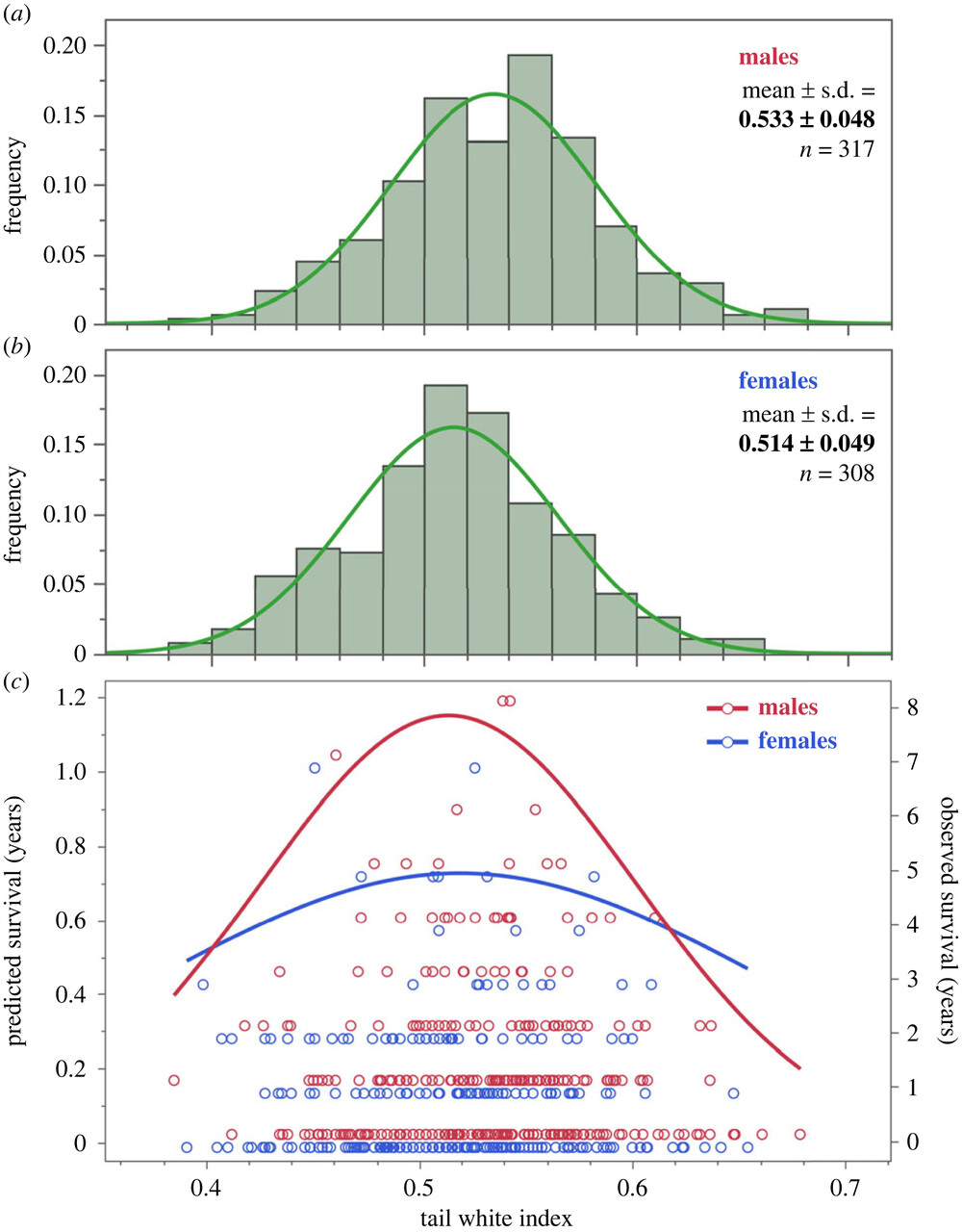
Рисунок 2. Стабилизирующий отбор по развитию белого на опахалах рулевых. Абсцисса — индекс развития белого, ордината — частота встреч особей с таким индексом у самцов (верхняя), самок (средняя), и прогнозируемая выживаемость (нижняя картинка, красные точки и аппроксимирующие кривые — самцы, синие — самки).
Считали, что наблюдаемая выживаемость — случайно распределённая переменная (по Пуассону), её связь с полом и развитием белого на рулях исследовали регрессионным анализом в обобщённой линейной модели со смешанными эффектами (GLMM). Авторы отказались от обычных способов её оценки, вроде модели Кормака — Джолли-Себера, основанной на мечении и повторном отлове, или модели Кокса, основанной на оценке пропорциональных опасностей, поскольку их интересовали не среднепопуляционные значения этих параметров, а связь наблюдаемой продолжительности жизни [в изученной группировке] и степени развития белого для разных индивидуумов.

Молодая самка на зимовке
Эту связь аппроксимировали сперва линейной, а затем квадратичной зависимостью (табл.1-2), это позволяет «поймать» и исследовать все возможные виды отбора, если они «работают» с ней: движущий (положительный или отрицательный), стабилизирующий, дестабилизирующий или дизруптивный. Пол и развитость белого были фиксированными эффектами в регрессионной модели, год начала наблюдений — случайный эффект. Наблюдаемая долговременная выживаемость недооценивает реальную продолжительность жизни птиц, по двум причинам:
1. Вильсонии, окольцованные в более позднем возрасте, имели впереди меньше лет вероятной жизни, чем окольцованные более молодыми: это корректируется включением года начала исследований как случайного эффекта в модель регрессии.
2. Окончательное исчезновение птиц с территории контролируемой группировки означает не только смерть, но и эмиграцию, которая авторами никак не отслеживалась (она особенно актуальна для самок, их филопатрия ниже, чем у самцов). Однако большая или меньшая развитость белого на рулях вряд ли влияет на вероятность эмиграции и вряд ли «толкает» птиц эмигрировать в какой-то конкретный год жизни: поэтому долговременное пребывание в контролируемой популяции представляется пусть неточной, но и не смещённой оценкой продолжительности жизни в контексте её связи и/или зависимости с развитием белого на рулях, а через него — с эффективностью кормодобывания.
Результаты работы строго доказывают что развитость белого на рулях контролируется стабилизирующим отбором: средние значения развития белого связаны с более долгой жизнью, чем низкие и высокие (у самцов оптимальная степень развития белого значимо выше, чем у самок). Поскольку те, другие и третьи не влияют на успех размножения в каждом из лет, проводимых птицей в данной группировке, её приспособленность прямо пропорциональна продолжительности жизни там. Скорее всего эта связь объясняется вспугивающим эффектом белых пятен определённого размера для тех насекомых, что служат кормом вильсониям, так что зависимость между размером пятен и эффективностью кормодобывания описывается оптимальной кривой.
Сниженная приспособленность птиц с большим развитием белого может быть связана также с ещё двумя факторами: белые участки пера легче изнашиваются, чем пигментированные, возможно это ухудшит лётные качества при изнашивании оперения птиц с более белыми рулями или более крупные пятна уже не столько облегчают вспугивание насекомых, сколько делают птицу заметней для хищников. Хотя ни одна из этих двух причин не объясняет пониженной выживаемости птиц с меньшим развитием белого, видимо, здесь действует только меньшая эффективность кормодобывания.
Выявленный авторами стабилизирующий отбор на самцов действует сильней, чем на самок и, главное, поддерживает значимо большую степень развития белого на их рулях, чем у самок. Возможных причин этому две. Первая главная: отбор действует главным образом на зимовке, когда самцы кормятся в тёмных и сомкнутых тропических лесах, самки — в более открытых и светлых зарослях кустарников. В первом случае размер «вспыхивающих» белых пятен для эффективного вспугивания насекомых, обеспечивающего их отлов, должен быть выше, чем во втором, чему есть и экспериментальные доказательства. К тому же растёт количество данных, что доступность пищи на зимовке или, наоборот, смертность из-за её недостатка — главный детерминант продолжительности жизни у дальних мигрантов.
Тем более что у капюшонной вильсонии размер пятен не влияет ни на репродуктивный успех, ни на массу птицы в текущем году (а эти два отклика очевидно связаны с эффективностью кормодобывания в гнездовое время, не на зимовке). Действительно, успех размножения у вильсоний определяется (отрицательно) в первую очередь гибелью гнёзд от хищников и гнездовым паразитизмом коровьих трупиалов Molothrus ater, не эффективностью кормодобывания.

Взрослая самка на гнезде
Вторая причина дополнительная — при ухаживании за самками из чужих пар самцы регулярно расправляют хвосты, так что белые пятна увеличиваются: нужно проверить, имеет ли это сигнальное значение, действительно ли самцы с более крупными пятнами успешней во внебрачных копуляциях. Если да, большая белизна рулевых у самцов связана с ним, тем более что рулевые самцов действительно несколько белей, чем обеспечивало бы наибольшую выживаемость (вершина параболы на рис.2).
Самое интересное и/или важное: выявленный авторами отбор формирует паттерн окраски вне обычно исследуемого ухаживания или, шире, социальной сигнализации.
Источник Proc. of the Royal Society B. Biol.Sci. 2023. V.290
P.S. публикатора: в чём смысл стабилизирующего отбора?
Понятие естественного отбора (англ. natural selection) отражает повсеместную ситуацию, когда в одной и той же среде обитания носители одних признаков разножаются лучше и/или дольше живут, носители же других, противопоставляемых им — хуже: отбор в популяции «поддерживает» первых и элиминирует вторых, что отражается в долговременном изменении их частот, а дальше — и в сдвиге степени выраженности и/или изменении характера детерминации обоих признаковых альтернатив.
Действие отбора (селективное давление) количественно оценивается по данным демографической статистики данным демографической статистики о дифференциальной размножаемости (If) и дифференциальной смертности (Im) индивидов индексом тотального отбора Дж.Кроу, предложенным в 1958 г.,
I = Im + 1/Ps*If,
Im = Pd/Ps; If = Vk/k2, Pd – доля лиц, умерших до наступления репродуктивного возраста, Ps – доля лиц, достигших репродуктивного возраста, k – среднее число детей, приходящееся на одного индивидуума, достигшего репродуктивного возраста, Vk – варианса числа детей. Вклад компонент изменяется на протяжении истории вида и/или при разных взаимодействиях его со средой. У Homo sapiens компонента Im максимальна в аграрных обществах доиндустриальной эпохи, и минимальна в промышленно развитых странах, If — наоборот (Алтухов, 2003, табл.7.23). На качественном уровне отбор действует, когда в контексте изменений среды обитания (в первую очередь процессов ландшафтной и биотопической трансформации, но не только) одни варианты организации (формы строения) биосистемы сильней размножаются и/или лучше выживают, другие нет (Костерин, Колесникова, 2009).

Работа по горным гаичкам на обложке Science. «Память работает: гаички с лучшей пространственной памятью живут дольше»
Скажем, у горных гаичек Parus gambeli, обитающих на больших высотах, выживаемость взрослых повышена вдвое по сравнению с живущими ниже, вопреки более суровым условиям, зимой особенно (рис.3). Их зимнее выживание зависит от качества пространственной памяти и пространственного обучения. Владельцам участков следует знать, где находятся созданные осенью запасы корма, и помнить, какие уже початы или разграблены, какие нетронуты. Действительно, жители больших высот создают больше запасов и лучше помнят их размещение. Молодые с лучшей пространственной памятью/обучением лучше переживают в первую, самую трудную зиму, почему дольше живут во взрослом состоянии (Benedict et al., 2020). Гаички с лучшей пространственной памятью — реже ошибающиеся в определении дверцы, за которой их раньше кормили — в естественных местообитаниях и живут дольше (Welklin et al., 2024).

Рисунок 3. А. Тестовая задача: надо помнить «свою» кормушку как минимум 4 дня. Синица с геолокатором (их носили все особи в исследовании, почему и удалось определить выживаемость) наблюдает за открыванием дверцы кормушки
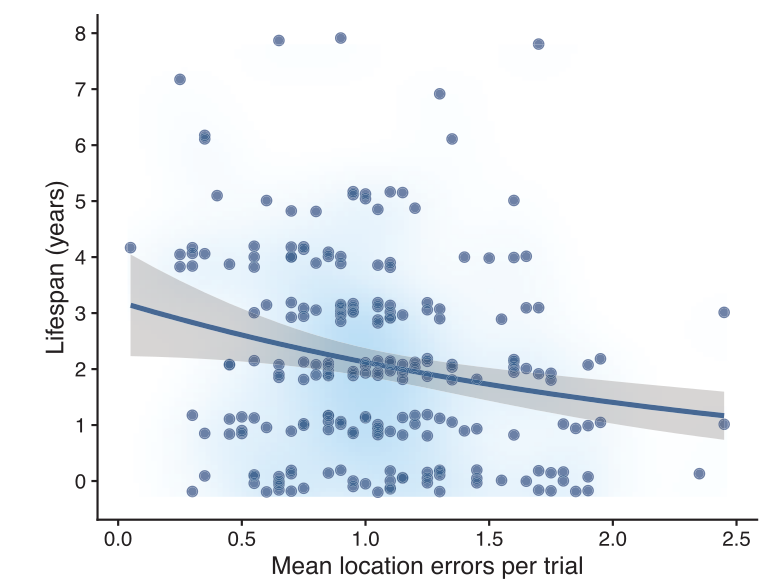
Б. Продолжительность жизни горных гаичек (годы, ордината) положительно связана со способностью к пространственному обучению и «мощью» пространственной памяти (абсцисса, средняя ошибка в определении «своей» кормушки).
Другой пример: из 126 фазанов Phasianus colchicus, выращенных в сходных условиях и выпущенных на волю в сельской Англии, 45 попало в зубы лисиц. Больше шансов погибнуть было у особей, хуже справлявшихся с тестами на пространственную память, когда были птенцами. Они же имели меньший участок, и наоборот: площадь участка пропорциональная когнитивным «достижениям» индивида: случаи гибели всех птиц (и хорошо справлявшихся) были в основном приурочены к периферии участков, хуже известной владельцам (Heathcote et al., 2023, см.пересказ А.В.Марковым).
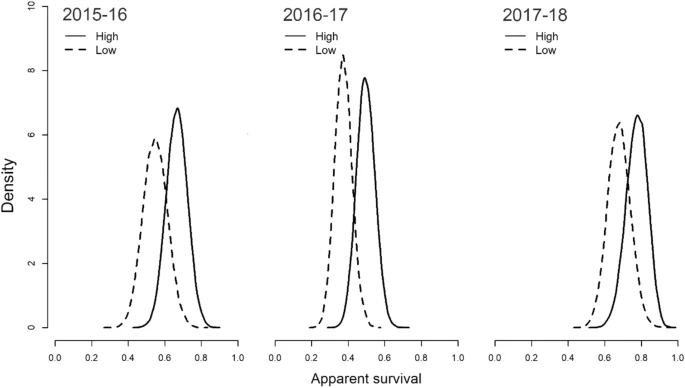
Рисунок 3. Распределения апостериорной вероятности выживания гаичек на высоко- и низкогорных участках (сплошная линия и пунктир) на протяжении трёх последовательных лет с разными зимами. Источник. Benedict et al., 2020, fig.2.
Эволюция — не только приспособление, но и вымирание
Зачем нужен стабилизирующий отбор? см.заголовок раздела: поддерживая жизнеспособность эволюционирующей популяции здесь и сейчас, в каждый данный момент изменений, запущенных отбором движущим, он минимизирует риск вымирания, сопровождающий всякое — и в первую очередь успешное — изменение под действием естественного отбора, ведь жизнеспособность популяции падает при всяком увеличении дисперсии по приспособленности, неважно, вследствие случайной гибели или успешной селекции. Увы, этот момент обычно остаётся “в забвении”.
Оценка отбором всякой биологической организации (как и социальной организации — конкуренцией) внутренне противоречива. С одной стороны, живому надо меняться, приспосабливаясь к изменениям среды, делающим обжитую, исторически типичную среду обитания экстремальной, обычно в режиме медленной катастрофы. Таковы все быстрые антропогенные трансформации ландшафтов (англ. HIREC — Human-Induced Rapid Environmental Change). Лучший пример здесь урбанизация, «пережёвывающая» и «дробящая» исходные местообитания вида в регионе (не только природные, но и антропогенные, скажем в традиционном деревенском ландшафте): она создаёт самую быструю средовую динамику, с наиболее радикальными ландшафтными трансформациями (Фридман, Суслов, 2022) — и именно здесь в разных группах биоты, от сосудистых растений до птиц и млекопитающих фиксируется самое быстрое эволюционирование! (Alberti et al., 2017, fig.3).
С другой стороны, живому приходится оставаться самим собой и воспроизводить именно себя любимого (разумеется, вместе с приспособительными изменениями), а не «не мышонка, не лягушку, а неведому зверушку», иначе отбор поддержит и распространит не тебя самого, а именно что эту зверушку. Поэтому приспособительные изменения, вырабатываемые отбором, могут быть не любыми, они должны удачно дополнять, усиливать и улучшать организацию (следуя «преимуществам» уже имеющейся конструкции или исключая и компенсируя её «слабые места»), но никак не рушить её, не быть песком в паровозных буксах или пятым колесом в телеге. Поэтому движущий отбор, а успешный особенно, имеет оборотной стороной снижение жизнеспособности, тем большее, чем более эффективное приспособление он формирует и чем быстрее идут изменения/чем жёстче отбор.
Что прямо подтверждено в опытах с сочетанием движущего и стабилизирующего отбора у Drosophila melanogaster. Оба вида отбора вели по признакам, друг с другом не связанным:
i) на положительную фотоактивность (сочетающую в себе фототаксис и локомоторную активность) и комплексу 9 метрических признаков крыла соответственно (Имашева и др., 1989а);
ii) на увеличение экспрессии мутации radius incompletus (ri) и комплексу метрических признаков крыла.
У гомозигот по ri она проявляется в разрыве второй радиальной жилки крыла, с образованием 2–х фрагментов, проксимального и дистального. Мух отбирали на уменьшение длины проксимального фрагмента (Имашева и др., 1989б; Лазебный и др., 1990; Лазебный и др., 1991). Движущему отбору везде подвергали оба пола, с интенсивностью 80%, стабилизирующему — только самок с интенсивностью 50%, в течение 16 поколений. При скрещивании мух исследуемой линии с тестерной, несущей компаунд–аутосомы. Эта интегральная оценка учитывает плодовитость и выживаемость вместе.
Опытные популяции делили на 3 повторности: в одной вели направленный отбор в сочетании со стабилизирующим (НС), в другой – только направленный (Н), в третьей отбора не было (К). В первых двух ответ на отбор равно значителен, в Н — с ростом вариабельности отбираемых признаков при ухудшении адаптивных свойств популяции (её численность и конкурентоспособность падали, флюктуирующая асимметрия росла, будучи почти неизменной в контроле). В НС везде происходит:
1) снижение эффекта направленного отбора (с 5 поколения);
2) падение уровня изменчивости по признаку, по которому ведётся первый, при постоянстве изменчивости признаков, по которым ведётся второй;
3) восстановление адаптивности, пониженной направленным отбором (с 12 поколения);
4) коррелированный ответ на отбор — начиная со второго поколения, дистальный фрагмент уменьшается также как проксимальный, хотя они контролируются разными генетическими системами (Имашева и др., 1989аб; Лазебный и др., 1990; Лазебный и др., 1991).
Т.е. стабилизирующий отбор восстанавливает адаптивные свойства популяции, разрушенные направленным в меру своей эффективности (список литературы).
Эволюция биологической организации всегда сопровождается риском вымирания, и чем быстрей и успешнее изменения под действием отбора, тем этот риск выше. Этот риск — оборотная сторона всякого эволюционирования, успешного или нет. Поскольку эволюция — это отбор не генов, а фенотипов, и сколько-нибудь сложная организация реагирует на него по прутковскому «щёлкни кобылу в нос, она махнёт хвостом», никогда нет гарантии, что начавшиеся изменения поднимут приспособленность организации в целом, даже когда это успешное приспособление к конкретному фактору неблагоприятности.
Действительно, не всегда изменения под действием естественного отбора адаптивны. Достаточно часто они “загоняют” вид или популяцию в “ловушку”, ведущую к вымиранию, как глобальное потепление — южных соловьёв (Remacha et al., 2020). В чём «ловушка»? На зимовку вид совершает транссахарские перелёты и там в Африке, сильно страдает в годы прихода засух (учащающиеся в связи с потеплением, особенно на юге и Африканском роге) — должны вылетать раньше, их больше гибнет в эти годы, прилетает и гнездится у нас меньше и пр. Важный фактор снижения этих рисков — более длинные крылья, и вроде приспособление должно действовать в эту сторону?
Однако работа по двум популяциям из Центральной Испании показывает, что в последние 20 лет, когда потепление развёртывается вовсю и сдвигает фенодаты не только прибытия, но и отлёта с зимовок, в т.ч. у дальних мигрантов, соловьи меняются в противоположную — и невыгодную — для них сторону, их крыло укорачивается! что делает их ещё более уязвимыми в условиях ухудшения обстановки на зимовках и трассе пролёта.
Почему так получается? В связи с потеплением в Центральной Испании лето стало суше, засухи чаще, суровей и дольше, почему временное «окно» для успешного размножения сократилось, оптимальным стал меньший размер кладки. А, как назло, до всякого потепления, у перелётных птиц, и особенно дальних мигрантов, сформировался так называемый «пакет генов, связанных с миграцией» («migratory gene package«), фактически суперген, регулирующий не только перелёты, но и подгоняющий жизненный цикл к их требованиям. И здесь большая длина крыла ассоциирована с большим уровнем базального метаболизма, большим размером кладки и меньшей продолжительностью жизни — либо наоборот.
Поэтому местный отбор в пользу меньших размеров кладки и более длинной жизни имеет последствия, неблагоприятные для перелётов, в том числе уменьшение длины крыла. Это противоречит общей тенденции приспособления: у североамериканских перелётных певчих птиц за время с 1978 г. вес и размер тела в целом стали меньше, а крылья длиннее. Изменения масштабней в более южных широтах, где потеплее, т.е. они отражают действие правила Бергманна (Weeks et al., 2020, Youngflesh et al., 2022).
И вот благодаря таким корреляциям, установленным в филогенетическом прошлом вида, отбор на приспособление к местным, испанским условиям, имеет губительную «отдачу» в виде увеличения уязвимости при перелётах (при тех же условиях потепления, что его вызвали). Это показывает относительность — и высокую инерционность — адаптации, объясняет случаи так называемой инадаптивной эволюции, когда отбор эффективен в каждый данный момент, но организация не выдерживает изменений и дохнет. Последнее вещь довольно обычная в с/х практике, почему для предотвращения этого приходится отбор по хозяйственно важным признакам сочетать с отбором по приспособленности (Васильева, Никоро, 1976; Frankham et al., 1988; Лазебный и др., 1991).
“Выжить” для вида и популяции безусловно важнее, чем “приспособиться”, этим взаимно противоречивым требованиям приходится удовлетворять в каждый из моментов процесса. [Повторюсь, именно в этих двух формах происходит существование живых организмов, они не независимы друг от друга (“россыпь”), а система. Типические паттерны связей между ними, образующие “конструкцию” этой системы (управляющая часть её, интересующая этологов — пространственно-этологическая структура популяции), для повседневного существования, приспособления и эволюции много важней собственных характеристик особей, первые управляют вторыми, а то и определяют их, но не наоборот].
Поэтому успешная эволюция (ведущая к биологическому успеху изменившейся популяции — росту численности, расширению ареала, устойчивости к прежним факторам неблагоприятности и даже использованию их во благо) требует одновременного действия двух давлений отбора — одного движущего, изменяющего один или несколько признаков, прямо взаимодействующих с факторами неблагоприятности, и одного стабилизирующего, повышающего жизнеспособность за счёт множественных координированных изменений разных аспектов организации биосистемы. Все вместе вторые компенсируют уменьшение жизнеспособности вследствие эффективности движущего отбора: это своего рода “тень” или “отдача” успешного эволюционирования, впервые установленная Эрнстом Майром в работе 1954 г. “Change of genetic environment and evolution”, показавшей, что изменения под действием отбора и стабилизация новой нормы происходят практически одновременно. Cегодня, увы, она подзабыта.
Другой пример: ставшие классическими доместикационные эксперименты Д.К.Беляева, Л.Н.Трут, О.В.Трапезова показывают, что у отбираемых по поведению лис, норок, крыс под селективным давлением “движется” не только целевой признак, но и множество других, связанных с первыми общими регуляторами (генными сетями, Суслов, Колчанов, 2009), или изменения которых участвуют в процессе регуляции отбираемого признака. Что хорошо видно по изменениям ряда морфологических признаков дружелюбных особей, в пользу которых отбор, сравнительно с агрессивными: дисперсия в первом случае не больше, чем во втором (Трут, 2007, рис.14-16, 18).
Благодаря этой “тени” или “отдачи” отбора вместо распада организации в переходные моменты эволюционирования (дестабилизации) мы видим нечто противоположное — её прецизионную подстройку к новой норме, причём быструю, без груза элиминации, превышающего “фон” в популяции, находящейся в стазисе.
Происходящее напоминает мораль в хасидской истории:
«как слабый человек может избежать греха, ведь в мире столько соблазнов? Представь себе человека, идущего по проволоке над бездной. Как сохранить устойчивость? Если твоё тело потянет в одну сторону, ты сам тянись в другую».
Для верующего соблазн угрожает духовной смертью, для биологических видов эволюционирование — физической, оно происходит лишь вынуждено, когда популяция захвачена экстремальным обострением борьбы за существование, в меру собственной устойчивости и сложности организации особей она “пробует” данные изменения “обезопасить”, т.е. направить и укоротить во времени.
А.С. Северцовым также показано, что сравнимая прецизионная подстройка к новой норме обеспечивается контрбалансом противонаправленных селективных давлений. Например, предпочтение больших значений признака Х, отражающих развитость брачного украшения или мощность ухаживательных демонстраций самцов неограниченно сдвинуло бы популяционную среднюю, действуй оно в одиночку (как и вышеописанные преимущества лучшей долговременной памяти горных гаичек или фазанов). Однако все подобные изменения в сторону большей развитости структуры, поддерживаемой отбором, большей мощности поведения и т.д. влекут за собой, кроме выигрыша, также плату в виде больших затрат или большего риска употребления.
Возникает противонаправленное давление отбора, определяющее наилучшее с точки зрения приспособленности значение Х здесь и сейчас. В итоге половой отбор в реальных сообществах животных половой отбор носит не движущий, а стабилизирующий характер: предпочтения самок (или самцов — процесс вполне симметричен), как всякие иные условия среды, отбирают не максимальное, а оптимальное выражение в соответствующих аспектах брачного поведения и/или брачных «украшений» вида, т.е. отбираемые признаки «брачных украшений» и других структур, связанных с привлекательностью, имеют предел не только снизу, но и сверху.
Скажем, самки ткачика-вдовушки Euplectes jackstone однозначно предпочитают самцов с хвостом подлиннее, чем длиннее, тем лучше, даже если удлинение явно приделано искусственно. То же видим у других ткачей рода Euplectes: у разных видов длина хвоста сильно варьирует, от довольно короткого, до очень длинного, и везде самки предпочитают «подлиннее«.
Таким образом, это общий тренд для рода, естественным образом вызывающий направленную эволюцию в сторону всё более длинного хвоста. Но! в ряде случаев половой отбор вступает в конфликт с задачей распознавания видов, когда Euplectes orix с хвостом порядка 4 см контактирует с длиннохвостыми видами с хвостом порядка 7-50 см., и мог бы с ним скрещиваться. В этом случае самки первого вида в опытах на предпочтение (самку помещают в клетку с тремя рукавами, открывающимися в клетки с соответствующими самцами; самка может предпочесть, проводя время около рукава с конкретным самцом, показать равнодушие или отвергнуть, избегая соответствующего «рукава»), предпочитают самцов с естественным хвостом (5 см.), равнодушны к контрольным (4 см.) и отвергают с коротким (3 см.), также как и со сверхнормальным 8 см. Т.е. при наличии ограничений, связанных с селективными давлениями другого рода (а контрбаланс противоположных давлений отбора — это норма, а не исключение; именно он обеспечивает стазис морфологии вида) половой отбор из движущего становится стабилизирующим.
Также как в исследовании предпочтительных уровней агрессивности самцов трёх видов грызунов с разными системами спаривания: 2-х промискуитетных (водяная полёвка Arvicola amphibius и домовая мышь Mus musculus) и 1ого моногамного (степная пеструшка Lagurus lagurus). М.А.Потаповым с соавт. (2010) показано, что у всех трёх видов разнокачественность самцов по обоим признакам очень велика, вплоть до выделения альтенативных стратегий, так что в популяции репрезентативно представлены все без исключения классы агрессивности и/или привлекательности. У моногамных пеструшек отчётливо предпочтение неагрессивных самцов, у 2-х полигиных видов предпочитаются более агрессивные, но не слишком агрессивные (рис.3).
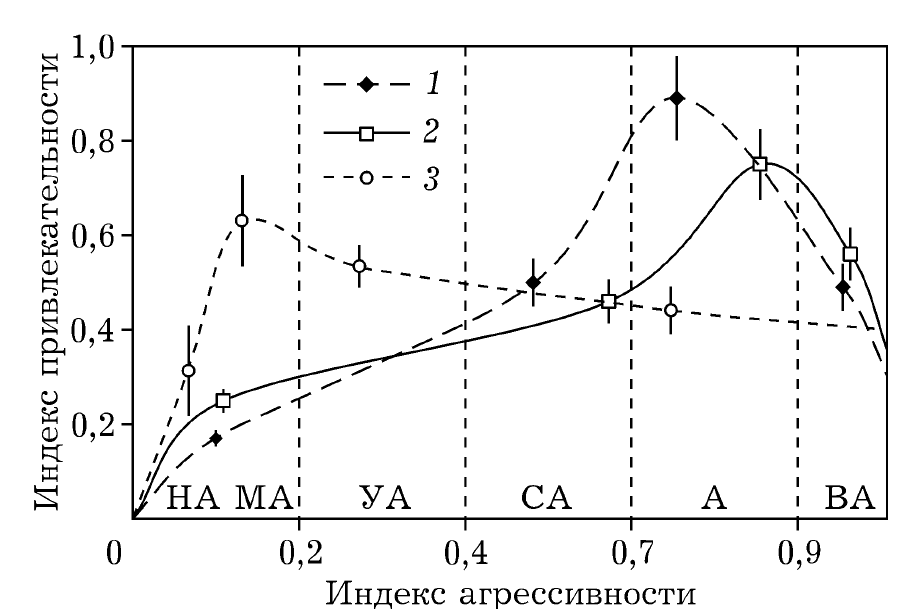
Нелинейная зависимость ольфакторной привлекательности самцов от их агрессивности. 1- домовая мышь, 2- водяная полёвка, 3 — степная пеструшка
Спаривания с более агрессивным самцом кроме «выигрыша» (большей вероятности эструса) влекут за собой «плату» — выше перинатальная смертность детёнышей, риски их гибели от каннибализма самца; высокагрессивные самцы обращаются с самкой так, что она оказывается худшей матерью (данные по домовым мышам, Потапов, 2010). Поэтому спаривания с более у многих видов животных, от жуков-мертвоедов до грызунов, самки при прочих равных ценят менее драчливых самцов — не только за «лучшее обхождение», но и потому что они адаптивней, меньше «повреждаются» в стычках с другими самцами.
Важно подчеркнуть, что у всех трёх видов половой отбор по агрессивности (производимый самками по ольфакторным критериям) равно оказывается не движущим, а стабилизирующим. Они снижает шансы на размножение как слишком агрессивных самцов, так и недостаточно агрессивных, устанавливает некий оптимум. Для каждого вида он свой, в зависимости от социальной системы и конкретно, системы скрещивания. Думаю, то же верно для любых изменений уровня специфической мотивации, как и всего что требует компромисса между несовместимыми затратами на размножение и витальные потребности. Тогда как отбор на развитость брачных украшений может быть чисто движущим, если они «более сигнальны, чем затратны».
Примечание
Рекомендуем прочесть
 Понятие приспособленности в структуре эволюционной теории
Понятие приспособленности в структуре эволюционной теории «Сортировка» вместо отбора в контексте урбанизации вида
«Сортировка» вместо отбора в контексте урбанизации вида Размножаемость и образование англичанок
Размножаемость и образование англичанок Быстрая эволюция в связи с урбанизацией: примеры
Быстрая эволюция в связи с урбанизацией: примеры О половом отборе в социальной истории нашего вида
О половом отборе в социальной истории нашего вида Против «отбора на ум» у евреев
Против «отбора на ум» у евреев Взаимоотношения полов у позвоночных и модели полового отбора
Взаимоотношения полов у позвоночных и модели полового отбора Про это
Про это
