
 Резюме. Приведены примеры быстрых эволюционных изменений в связи с урбанизацией у птиц и ящериц. Важно, что экологические (приспособление) и микроэволюционные изменения во всех этих случаях одновременны, а не следуют друг за другом во времени, как предполагается в СТЭ. Скорость изменений также существенно выше, чем можно ожидать на основании СТЭ. В ответ на изменения среды, представляющие собой «локальную катастрофу», эволюция «шьёт из готового», стабилизирующий отбор поддерживает все генные варианты/физиологические проявления имеющейся организации, «работающие» на рост её устойчивости и лучшее воспроизводство популяции вопреки катастрофе.
Резюме. Приведены примеры быстрых эволюционных изменений в связи с урбанизацией у птиц и ящериц. Важно, что экологические (приспособление) и микроэволюционные изменения во всех этих случаях одновременны, а не следуют друг за другом во времени, как предполагается в СТЭ. Скорость изменений также существенно выше, чем можно ожидать на основании СТЭ. В ответ на изменения среды, представляющие собой «локальную катастрофу», эволюция «шьёт из готового», стабилизирующий отбор поддерживает все генные варианты/физиологические проявления имеющейся организации, «работающие» на рост её устойчивости и лучшее воспроизводство популяции вопреки катастрофе.
В.В.Сусловым было показано, что для «поиска» генных вариантов, наилучших в плане локальной адаптации к неблагоприятным изменениям среды (сценарий, нормативный для СТЭ) потребует
а) уровня элиминации, несовместной с жизнеспособностью всякой реальной популяции, особенно у долгоживущих видов позвоночных (включая растущий риск вымирания от случайных причин при любом понижении средней численности) и
б) времени, много большего, чем потребное для средовых изменений.
Описанный механизм «обходит» это противоречие: реальная микроэволюция поэтому идёт много быстрей, чем предполагается СТЭ, и с несопоставимо меньшим риском элиминации (в том числе легко обходит известную дилемму Холдейна). Он позволяет снизить плату за отбор путём направленного «поиска» мутаций, сколько-нибудь перспективных в плане предполагаемых изменений, вместо их случайного «перебора» и «тестирования» отбором, несовместного с жизнью.
Параллельная эволюция термостойкости и конечностей у бурых анолисов в городах
Содержание
а) могут устойчиво действовать,б) пользоваться присадами,в) ещё переносят эту температуру тела,
Критический максимум температуры для «городских» анолисов в каждом из 4-х городов острова (Mayagüez, Aguadilla, Arecibo, San Juan) в среднем на 0,870С выше значимо выше, чем для «лесной» популяции от которой произошла данная городская (путём независимой колонизации). В г.Mayagüez ящерицы активны при максимальной температуре 400С.
Длина самого животного без хвоста ~5 см. Термостойкость оценивали, укладывая их на спину при постепенно растущей температуре. Её значение, при котором ящерицы не сразу «встают на ноги», и есть предел термостойкости.

Бурый анолис A.cristatellus, более теплолюбивый, чем упоминающий дальше зелёный
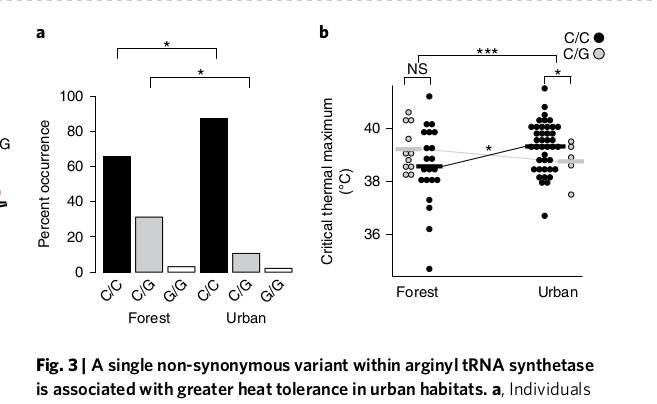
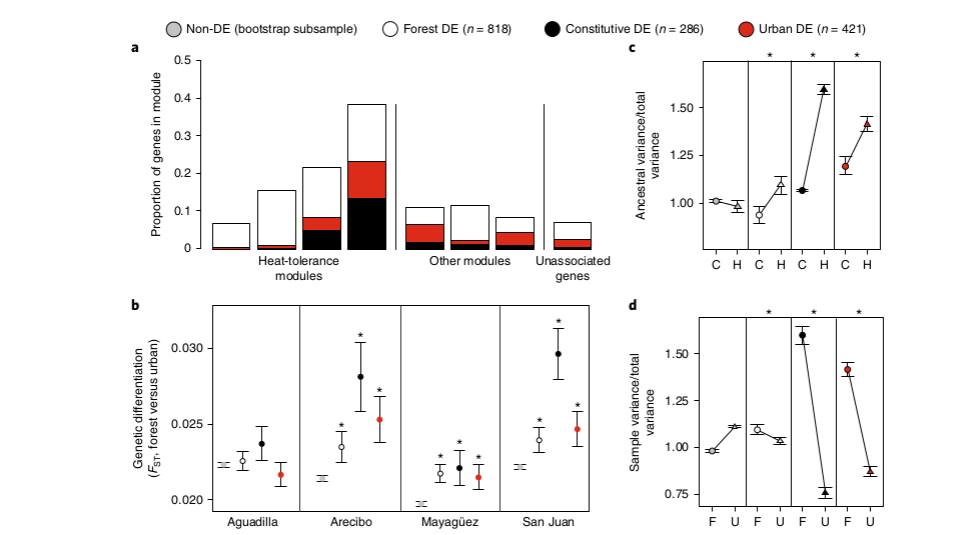 Урбанизация ведёт к накоплению 4-х несинонимичных замен в этом гене в позициях 1673, 1853, 595, 796. Первая из них наиболее значима функциональна: среди «городских» анолисов на 21,6% повышена доля особей с С-С там (это позиция 558 в белке, треонин заменён на серин), и понижена на 20,4% — с С-G. Только этот полиморфизм показывал значимые различия между популяциями во всех 4-х городах; именно у «городских» анолисов гомозиготы С-С более термоустойчивы, за счёт лучшей акклимации в ответ на тепловой стресс (fig.2-3).
Урбанизация ведёт к накоплению 4-х несинонимичных замен в этом гене в позициях 1673, 1853, 595, 796. Первая из них наиболее значима функциональна: среди «городских» анолисов на 21,6% повышена доля особей с С-С там (это позиция 558 в белке, треонин заменён на серин), и понижена на 20,4% — с С-G. Только этот полиморфизм показывал значимые различия между популяциями во всех 4-х городах; именно у «городских» анолисов гомозиготы С-С более термоустойчивы, за счёт лучшей акклимации в ответ на тепловой стресс (fig.2-3). 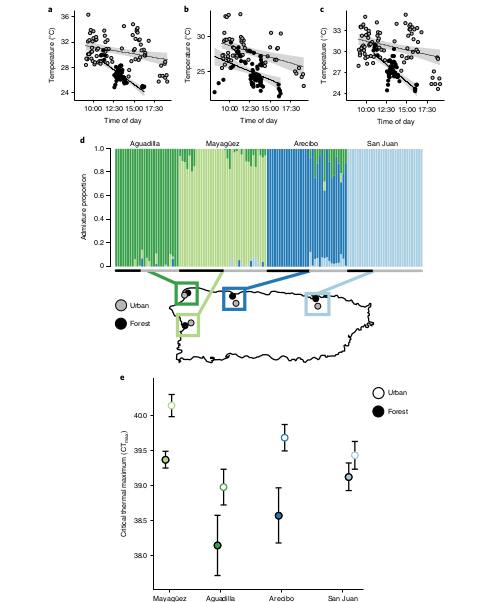
 Среди них выделили гены, предположительно связанные с термотолерантностью (у них была значительней разница частот полиморфизмов между «городскими» и «лесными» популяциями, чем у остального генома). Далее авторы из кладок «лесных» и «городских» анолисов в стандартных условиях искусственно вырастили потомство (n=7 и 9). Гены, чья экспрессия отличалась у «лесных» ящериц, также показывали большую дисперсию экспрессии при 320С, чем при 250С, создавая возможность дальнейшего роста термоустойчивости под действием тех же селективных давлений (fig.4).
Среди них выделили гены, предположительно связанные с термотолерантностью (у них была значительней разница частот полиморфизмов между «городскими» и «лесными» популяциями, чем у остального генома). Далее авторы из кладок «лесных» и «городских» анолисов в стандартных условиях искусственно вырастили потомство (n=7 и 9). Гены, чья экспрессия отличалась у «лесных» ящериц, также показывали большую дисперсию экспрессии при 320С, чем при 250С, создавая возможность дальнейшего роста термоустойчивости под действием тех же селективных давлений (fig.4). 
«Городские» анолисы бегают по вертикальным поверхностям
Осваивая города, анолисы удлинили конечности, чтоб спасаться вне леса, на открытых пространствах; потом они стали взбираться наверх. Но ответ на отбор в пользу лучшего лазанья, увеличив сцепление пальцев с субстратом, не максимизировал эту способность (укоротив ноги снова) ради интеграции обоих приспособлений: это т. н. адаптивный компромисс (Winchell et al., 2018).
Быстрая эволюция зелёных анолисов в ответ на неожиданные удары холода в Техасе
«Это убедительный пример быстрой эволюции»,

Бурый и зелёный анолисы, поза угрозы
Анолисы, перемещённые на остров с преобладанием древесной растительности над кустарниками, за 20 лет удлинили ноги. Жизнь на стволах деревьев требует проходить большие расстояния, жизнь в кустарниках – большей скорости и ловкости рывков – и переселенцы на закустаренные острова с единичными деревьями в те же 20 лет ноги укоротили. Интересно, что эти переселения ящериц на необитаемые острова Багамского архипелага исходно делались для того, чтобы изучать вымирание на архипелагах, предсказываемое моделями “островной биогеографии”.

Геккон Gymnodactylus amarali, быстро эволюционировавший на островах водохранилища в Бразилии

Зелёные анолисы из Браунсвилля, Техас, быстро повысили холодоустойчивость после аномальных зим
Укорочение крыльев скальной ласточки Petrochelidon pyrrhonota снизило гибель от столкновения с а/м
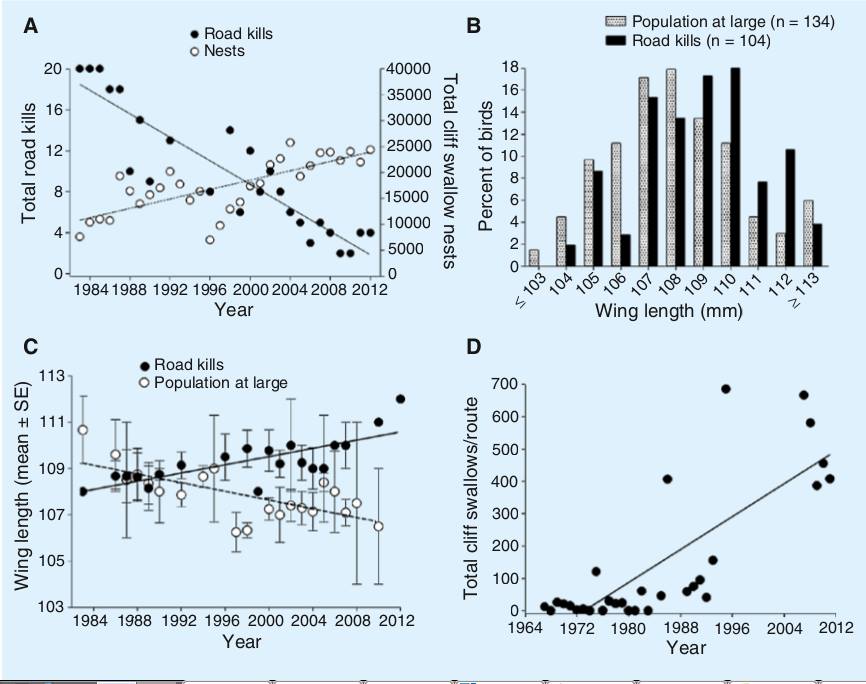 Рис. 5. Изменения числа погибших на дороге ласточек, и их длины крыла на юго-западе Небраски. За 30 лет это число неуклонно падает; наиболее длиннокрылые особи составляют непропорционально большую долю погибших во все годы после возникновения каждой из придорожных колоний ласточек. (A) Число потенциально предотвратимых смертей ласточек на дороге (темные кружки, сплошная линия) значительно снизилось за годы наблюдений (rs = 0,93, p <0,0001, n = 25 лет), несмотря на рост численности населения вокруг дорог в тот же период (белые кружки, пунктирная линия) (rs = 0,76, p <0,0001, n = 30 лет). Годы от появления колонии здесь — значимый предиктор числа столкновений ласточек с машинами (F1,22 = 38,8, p <0,0001, GLM), но не численности населения вокруг (F1,22 = 0,01, p = 0,93); (B) длины крыльев у птиц, погибших на дорогах (темные столбцы) в сравнении со всей популяцией (измерено у попадающих в паутинные сети; заштрихованные столбцы): видны значимые различия (F1,229 = 7,06, p = 0,007, ANCOVA); (C) у ласточек, погибающих на дорогах, средняя длина крыла (± SE; темные кружки, сплошная линия), с течением времени сильно выросла (rs = 0,78, p <0,0001, n = 20 лет), тогда как у популяции в целом (белые кружки, пунктир), за тот же период уменьшилась (rs = 0,63, p = 0,002, n = 21); (D) Общее количество ласточек в скалах, о которых сообщалось в ходе обследования гнездящихся птиц в округе Кейт, штат Небраска, за 1967–2011 гг., Указывает на то, что эти виды начали часто сталкиваться с транспортными средствами примерно в то время, когда началось это исследование. Линии указывают на регрессию методом наименьших квадратов наилучшего соответствия.
Рис. 5. Изменения числа погибших на дороге ласточек, и их длины крыла на юго-западе Небраски. За 30 лет это число неуклонно падает; наиболее длиннокрылые особи составляют непропорционально большую долю погибших во все годы после возникновения каждой из придорожных колоний ласточек. (A) Число потенциально предотвратимых смертей ласточек на дороге (темные кружки, сплошная линия) значительно снизилось за годы наблюдений (rs = 0,93, p <0,0001, n = 25 лет), несмотря на рост численности населения вокруг дорог в тот же период (белые кружки, пунктирная линия) (rs = 0,76, p <0,0001, n = 30 лет). Годы от появления колонии здесь — значимый предиктор числа столкновений ласточек с машинами (F1,22 = 38,8, p <0,0001, GLM), но не численности населения вокруг (F1,22 = 0,01, p = 0,93); (B) длины крыльев у птиц, погибших на дорогах (темные столбцы) в сравнении со всей популяцией (измерено у попадающих в паутинные сети; заштрихованные столбцы): видны значимые различия (F1,229 = 7,06, p = 0,007, ANCOVA); (C) у ласточек, погибающих на дорогах, средняя длина крыла (± SE; темные кружки, сплошная линия), с течением времени сильно выросла (rs = 0,78, p <0,0001, n = 20 лет), тогда как у популяции в целом (белые кружки, пунктир), за тот же период уменьшилась (rs = 0,63, p = 0,002, n = 21); (D) Общее количество ласточек в скалах, о которых сообщалось в ходе обследования гнездящихся птиц в округе Кейт, штат Небраска, за 1967–2011 гг., Указывает на то, что эти виды начали часто сталкиваться с транспортными средствами примерно в то время, когда началось это исследование. Линии указывают на регрессию методом наименьших квадратов наилучшего соответствия.
Около 80 млн птиц ежегодно гибнет от столкновений с машинами на дорогах США [1], и более миллиона — ежегодно в Европе [2] и других местах. Гибель от столкновений с а/м — серьёзная проблема для популяций, для разрешения которой были предложены различные изменения в конструкции и содержании а/д [3]. Однако, учитывая масштабы смертности некоторых видов [4], можно ожидать давления отбора в особей, учащихся избегать автомобилей, или чьи морфологические особенно снижают риск гибели от столкновений с а/м.
Если этот отбор идёт, число случаев гибели [у таких видов] должно падать со временем. В литературе мы не смогли найти информацию, действительно ли это происходит у каких-то видов [2]. Однако мы сами обнаружили это снижение в ходе 30-летнего исследования социального поведения и колониальности у скальных ласточек (Petrochelidon pyrrhonota) на юго-западе Небраски. Фиксируется а) снижение числа ласточек, гибнущих на дорогах, за годы, следующие после появления колоний на эстакадах и обочинах дорог, б) всех эти годы на дорогах избирательно гибнут особи, более длиннокрылые, чем в целом по популяции.
Мы работаем со скалистыми ласточками с 1982 года на юго-западе Небраски, с центром в округе Кейт на биологической станции Сидар-Пойнт (41° 12,591′ с.ш., 101° 38,969′ з.д.). Ласточки гнездятся колониально: свои глиняные тыквоподобные гнёзда они прикрепляют группами на вертикальной стене под горизонтальным выступом. Колонии размещаются в основном на автомобильных мостах, эстакадах, в бетонных колодцах, водопропускных трубах под дорогами или железнодорожными путями [5].
Исследования начинались после того, как птицы заняли места гнездования на обочинах дорог. Ежедневно передвигаясь между колониями, мы останавливались перед каждой погибшей на дороге ласточкой [для их подcчёта и измерения, а также препаровки для долговременного хранения. Это производили из года в год на одних и тех же дорогах, предпринимая одинаковые усилия для поиска трупов. Число ласточек, убитых автомобилями, сравнивали с числом случайно погибших в паутинных сетях, что отражало изменение влияния автомобилей как фактора смертности по годам. Оно было примерно постоянным, а вот число гибнущих на дороге значимо сократилось в 1983-2021 г. (рис.5а), тогда как численность вида в ближайших окрестностях дорог, наоборот, выросла (рис.1).
Уменьшение числа трупов на дорогах не объяснить ростом численности птиц-падальщиков в те же годы: ни один из их видов не показывает этого в данной местности. Прямая информация о местных падальщиках-млекопитающих, связанных с людьми, недоступна, хотя вряд ли менялась, ведь постоянное население округа Кейт не изменилось. Численность некоторых из них, вроде скунсов, даже упала в целом в штате Небраска [6]. Это не объяснить и сокращением трафика: на разных дорогах он или не изменился, или увеличился (в зависимости от используемых показателей оценки). По сравнению с 1980-х больше стала доля внедорожников и меньше — седанов, а первые опасней для птиц из-за большей поверхности для столкновения. Однако смертность ласточек на дорогах падала параллельно распространению более крупных а/м, с большей поверхностью, т. е. потенциально более опасных для столкновений.
Оригинал в Current Biology
Рекомендуем прочесть
 «Сортировка» вместо отбора в контексте урбанизации вида
«Сортировка» вместо отбора в контексте урбанизации вида Пресечение урбанизации птиц как зеркало московского экоцида
Пресечение урбанизации птиц как зеркало московского экоцида «Гены урбанизации» и приспособление к средовому стрессу
«Гены урбанизации» и приспособление к средовому стрессу Мини-рецензия на книгу Менно Схилтхёйзена «Дарвин в городе. Как эволюция продолжается в городских джунглях»
Мини-рецензия на книгу Менно Схилтхёйзена «Дарвин в городе. Как эволюция продолжается в городских джунглях» Теории биологической эволюции с позиций современного развития системной биологии
Теории биологической эволюции с позиций современного развития системной биологии Урбосреда как развивающая: сравнение с одомашниванием
Урбосреда как развивающая: сравнение с одомашниванием Дарвиновы вьюрки: расщепление и слияние форм
Дарвиновы вьюрки: расщепление и слияние форм Урбанизация «диких» видов птиц/млекопитающих как доместикация домашних
Урбанизация «диких» видов птиц/млекопитающих как доместикация домашних
