 В продолжение рассказа об опасностях коммуникации
В продолжение рассказа об опасностях коммуникации
Введение
Конфликты за ограниченные ресурсы исследовали у широкого круга видов (Huntingford & Turner 1987). Обычно рассматриваются два вида «платы» за них — это травмирование и/или потери времени и энергии. Неучтенным остаётся повышенный риск попасться хищнику, будучи «занятым» в конфликте (Jakobsson 1987; Hammerstein & Reichert 1988). Авторы изучали охрану территорий самцами двух видов, становящихся жертвой хищничества, — южноамериканской цихлиды Nannacara anomala и пеночки-веснички Phylloscopus trochilus из Западной Палеарктики. У обоих бдительность понижается по мере развития конфликта, поэтому риск хищничества важен для понимания как эволюции демонстраций ритуализированной угрозы, так и «нормативных» последовательностей смены демонстраций и действий по ходу конфликта, его «синтаксис».
Материал и методы, результаты
Самцы весничек прилетают с африканских зимовок в район наших исследований (зоологическая исследовательская станция Товеторп, 58”56′ с.ш., 17”08″ в. д.) в конце апреля -начале мая (Jakobsson, 1988).
Самки прилетят примерно через две недели; систему спаривания лучше всего охарактеризовать как социальную и генетическую моногамию (Gyllensten et al., 1990). Самцы активно конфликтуют за территории и легко провоцируются [на проявление территориальной агрессии]. Воспроизведение песни веснички и подстановка чучела самца выявляют весь спектр агонистического поведения вида (например, сближение, трепетание крыльев и атака) (May 1949; Jakobsson 1987). Вторжение захватчика имитировали, выставив чучело веснички и проигрывая песню в границах самцовых территорий. Обострение конфликта определялось моментами, когда хозяин территории [после всех демонстраций, пения и перелётов] доходил до того что садился на чучело и клевал его в голову и глаза, как это случается и в настоящих конфликтах за территорию. Конфликт без обострения определяли как демонстрации и действия самца, сидящего в радиусе до 1 м от чучела и на сопоставимой высоте (0,5 м над землей).
Для проверки, действительно ли эскалация конфликта снижает способность весничек отследить внезапное появление хищника, авторы пользовались чучелом воробьиного сыча Glaucidium passerinum, обычного хищника в нашем районе исследований. Его устанавливали на высоте 2 над землей в примерно 2,5 м от места проигрывания песни, и держали спрятанным в коричневой пластиковой трубе, подобной стволу дерева. Экспериментаторы могли быстро вытащить чучело за моноволоконную леску в разные моменты реакции владельцев участка на провокации, с эскалацией угроз иди без. Охота сыча на птиц включает поиск и посадку повыше потенциальной жертвы прежде чем атаковать из этой засады (данные > 100 часов наблюдений за радиопрослеживаемыми совами, С. Куллберг, неопубликованные данные). Внезапное появление воробьиного сыча на небольшом расстоянии сверху — смертельный риск для всех мелких птиц. Все опыты провели утром и днём, с 06:00 до 12:00 мая 1993 г., до появления самок весничек, их ставили при освещённости, облегчающей выявление. Весничек протестировали по 10 раз на этапе эскалации конфликта и в ответе без него. Момент первого опыта в каждый день выбирали случайно, если дело доходило до драки, следующую птицу тестировали пока не дошло и пр. Всех их тестировали лишь однажды: если это было при эскалации конфликта, в следующий раз сову предъявляли в отсутствие эскалации и т.д.
Каждая птица подвергалась испытанию только один раз: лишь один 6-летний самец до того слышал проигрывание, будучи пойман так в 1988 году. Владельцев соседних участков в один день никогда не тестировали. Контролем было предъявление 6 самцам чучела поползня Sitta europea вместо сыча до обострения конфликта, все разы без какой-то реакции. Ещё 6 тестирований исключили из анализа, так как 2 раза веснички напали на чучело до обострения, 4 раза — вообще не обращали внимание, не присаживались ближе 1 м от чучела.
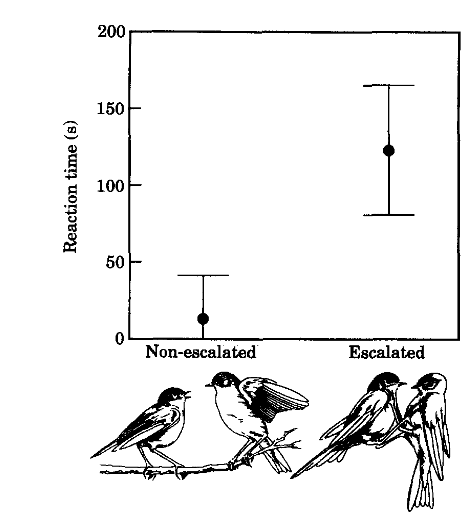
Рисунок 1. Латентный период реагирования самцов весничек на неожиданную демонстрацию чучела сыча вблизи. Справа — эскалация конфликта, слева — её нет (оба N= 10).
В случаях эскалации конфликта самцы веснички реагировали на опасность значимо позже, чем в отсутствие эскалации: среднее время реакции±стандартная ошибка = 123,1±42,0 с (диапазон 28-150 с, медиана= 150 с) vs 12,7±28, 2 (l-90 с, медиана = 1 с). Различия значимы по критерию Манна-Уитни, U = -3,76, P=09002, n=10. Сравнение всех других форм поведения между эскалацией территориальных конфликтов и её отсутствием не выявило различий ни по одной из них. Значимость различий по латентному времени реагирования на воспроизведение песни z = -0,53, P=0,60; по времени, необходимому для приближения к чучелу <5 м z= -1,74, P=0,08; то же, на дистанцию <1 м: z= -1,10, P=27,000; по времени обнаружения чучела сыча z= -0,83, P=41,0000; везде использовали тест Манна-Уитни U. В 7 из 10 предъявлений сыча при эскалации конфликта участники так и не среагировали на опасность на всём протяжении тестирования (150 с). Напротив, при отсутствии эскалации сову за период тестирования все веснички среагировали на сыча, у 7 из 10 это длилось секунду и более. Обнаружив сыча, они сразу слетали с тревожными криками.
Другую серию опытов авторы ставили с половозрелыми самцами Nаnnacara anomalа, небольшой южноамериканской цихлиды, чтобы выяснить восприятие риска нападения хищника при разной интенсивности территориальной агрессии: максимальной («борьба ртов», где самцы пробуют вытеснить оппонента), низкой (удары хвостом, направляющие волну на органы боковой линии оппонента, болевое воздействие) или же в одиночку. Критерием реактивности была дистанция, с которой рыбы заметят быстрое приближение модели хищника, установленной на электропоезд, проход которого был одновременно масштабом.
Самцы этого вида конфликтуют за ранг в системе агонистического доминирования. Конфликты имеют постоянную временную структуру, заданную «повышением планки» ритуализованной агрессии, и начинается с её низкоинтенсивных форм: боковой демонстрации и ударов хвостом (Jakobsson et al. 1987). Соперники здесь поворачиваются боком, и ударами хвостов направляют струи воды на боковую линию друг друга (рис.2). После ударов самцы поворачиваются фронтально, стараясь захватить пасть противника и крепко удерживать. Если это выходит, начинается «борьба ртов». Её продолжительность варьирует, будучи в среднем около 10 с (диапазон 1-144 с, см. Enquist et al., 1990). Между раундами собственно схватки вкраплены демонстрации: боковая, удары хвостом и ритуализованное кусание оппонента.
Аквариум (30 * 30 * 30 см) делила надвое непрозрачная перегородка. В каждой половине была коробка с самцом цихлиды. За 15 минут до начала опыта убрали перегородку и дистанционно открыли обе коробки, пробудив рыбок взаимодействовать (Enquist et al., 1990). Когда конфликт достигал стадии, требующей тестирования (раунды «борьбы ртов» или ударов хвостом), модель хищника, изначально спрятанную за ширмой, выдвигали наружу и двигали в сторону рыб. Модель хищника делалась видимой на максимальной дистанции — 3,65 м и двигалась к аквариуму со скоростью около 72 м/с. Все опыты снимали на видео, что позволяло для каждого из них рассчитать дистанцию приближения, с которой начинается бегство.
Рыбок покупали у местного дилера и содержали в 250-литровых аквариумах до начала появления у них признаков самцов, далее пересаживали в отдельный аквариум (30 * 30 * 30 см) минимум на 2 недели перед опытами; за 24 ч до них рыбок сажали в аквариум для тестирования. В «борьбу ртов» перерастают конфликты рыб одного веса (Enquist et al., 1990), пары соперников подбирали по весу. Возраст цихлид в это время был 5-6 месяцев, вес 0,5-1,5 г; каждую рыбку тестировали лишь раз.
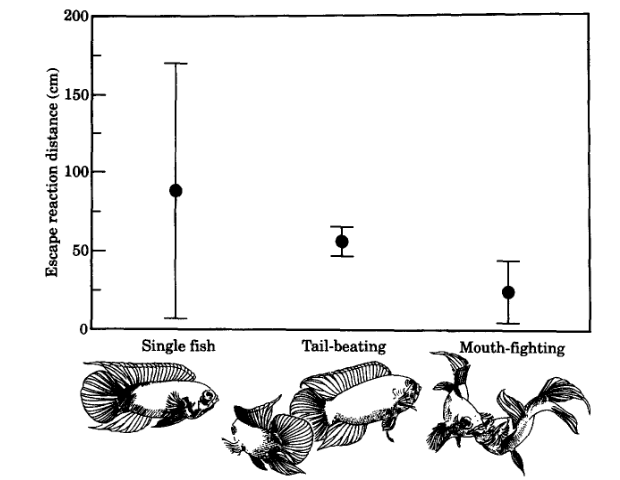
Рисунок 2. Дистанция обнаружения приближающегося хищника самцами N. anomalа (в см, ордината). Абсцисса — ситуация, в которой распознаётся хищник: слева направо вне конфликтов, самцы одиночны; самцы конфликтуют, раунд «ударов хвостом»; то же, раунд » борьба ртов».
Во всех аквариумах был гравийный субстрат, поддерживалась постоянная температура температура 26°C при световом дне 12 ч. Раз в день рыб кормили живым или мороженым мотылем либо артемией. Ни одна рыба не травмировалась во время конфликтов; они различались длительностью, но модель хищника всегда предъявляли в их начале (также по 10). Среднее время между началом «борьбы ртов» и демонстрацией хищника 23,6 с ± SE 8,6 с (2-92 с). В каждом конфликте до «борьбы ртов» прошло в среднем по 2 удара; ни разу не не отмечали кружения, описанного ранее у этой и прочих цихлид (см. Enquist et al., 1990).
Самцы, вовлечённые в «борьбу ртов», подпускают модель значимо ближе и бегут позже (рис. 2), чем участвующие в «ударах хвостом» и тем более чем одиночные (на 24,9 ± SD 20,0 см (пределы O-54,2 см, медиана=245 см, N=l0 vs 56,8 ± 9,3 см (36,9-69,8), медиана 55 см, N= 10 vs 88,9±81,1 см (30,1 — 357,5) медиана 51,2 см, N=19.
Различия по дистанции бегства от хищника значимы между двумя случаями : «борьба ртов» vs «удары хвостом», z= — P=O,034; » борьба ртов» vs одиночки z= -2,98, P= -0,029; везде тест U Манна-Уитни. Различия в дистанции бегства незначимы между одиночными рыбами и фазой ударов хвостом, z= — 0,35; P=O,85. Бегство от модели представляло собой, в основном, быстрый бросок прочь с дальнейшим уплыванием. В 3 из 10 тестирований во время «борьбы ртов» участники начали движение, когда модель хищника уже запустили, но она все еще невидима за ширмой, их исключили из анализа ибо условием было, что рыбы заняты «борьбой ртов» в момент, когда модель делалась видимой.
Исключенные дистанции бегства — 91,6, 61,4 и 52,6 см; при их учёте различия остаются значимы: «борьба ртов» vs «удары хвостом» z= — 2,04, P=0, 041; «борьба ртов» vs одиночки: z= — 2,16, P=O,O31. Интересно, что в этих трех случаях, когда самцы уже расплывались, прекратив «борьбу ртом», невидимую, рыбы замечали на большей дистанции, чем оставшиеся 7, занятые » борьбой ртов» (z= -2,17, P=0,03).
Обсуждение
Авторы разделяют теорию выбора действий в агонистических контактах, основанную на подходе эволюционно-стабильных стратегий ESS, утверждая их эффективность в объяснении эволюции данного поведения (Maynard Smith, Price 1973; Maynard Smith 1982) в оценке способности бороться (Parker 1974; Maynard Smith, Parker 1976), или в понимании правил принятия решений в поединках (Enquist, Leimar 1983). [Правда, работы данных авторов по агонистическому поведению глупыша и этого же вида цихлид показывает необходимость учесть, что вовсе не все формы поведения здесь — силовые действия одной особи против другой. Часть из них, и самая важная во всех трёх названных выше аспектах — сигналы, материальный носитель которых — ритуализованные демонстрации. В оценке платы и выигрыша от их использования, обязательной для понимания того, какие модели поведения в конфликтах могут быть ESS, какие нет, нужно учесть их сигнальную эффективность и риск демонстрирования. Снижение реактивности при появлении угрозы гибели, отмеченное в данной работе в связи с эскалацией конфликта, т. е. использованием более ритуализованных форм поведения, не только более ожесточённой борьбы — важная часть риска демонстрирования. Последний включает ещё 2 составляющие (риск травмирования оппонентом и риск проигрыша взаимодействия, если оппонент точней скорректирует будущее поведение по информации, содержащейся в демонстрациях данной особи), и неизменно пропорционален риску демонстрирования, как выяснили эти авторы для глупышей, J. Popp для американских чижей и мексиканских чечевичников и пр. См.подробней «Риск демонстрирования как подтверждение сигнальности демонстраций«. Прим.публ.]
Описанный выше подход раньше никак не учитывал рост риска гибели от хищника при эскалации борьбы [я добавлю, что как только мы отделяем силовые действия от сигнальных и вторые характеризуем эффективностью и риском демонстрирования, подходы сравнительных этологов и социобиологов делаются не только вполне совместимыми, но и взаимно обогащающими. Прим.публ.].
Отсутствие исследований роста риска хищничества при демонстрировании удивительно, поскольку поведенческие решения, принимаемые в условиях риска нападения хищников (обзор см. в Lima & Dill, 1990), уже вызвали большой интерес в контексте поведения ухаживания (Ryan, 1985; Magurran & Seghers, 1990), выбора партнера (Forsgren, 1992), заботы о потомстве (Magnhagen, 1991), исследования возможного наличия хищников (Milinski, 1987). и кормодобывания (Milinski & Heller, 1978). Недавно показано, что молодь кижуча Onchorynchus kisutch значимо менее агрессивна к зеркалу, когда к воде примешаны запахи хищника.
Считая больший риск гибели от хищника «ценой» эскалации боевых действий, возможно иначе взглянуть на недоизученные аспекты агонистического поведения, вроде ритуализации отдельных элементов репертуара и [правил] выбора действий в проблемной ситуации. Скажем, агонистическое поведение временами включает совместные действия участников («удары хвостом» и «борьба ртов» этого вида именно таковы), когда корректировка демонстраций или отказ от демонстрирования полезны обоим. Особенно интересен здесь «разворот», когда от эскалации конфликта участники возвращаются к менее интенсивному демонстрированию, присущему начальной и срединным стадиям взаимодействия.
Возможно повышение бдительности участников (потенциальных жертв), что увеличивает их шанс обнаружить затаившихся хищников и спастись от гибели.
Структура конфликта, увеличивающая риск демонстрирования и смелость участников по ходу взаимодействия, даёт им обоим информацию о «качестве» используемых моделей поведения [их эффективности и риске, прежде всего для специфических единиц поведения, демонстраций] при минимуме «платы за вход». Риск, созданный травматизмом, и риск гибели от хищника здесь идут рука об руку. [Отмеченная авторами зависимость вполне подтвердилась у всех видов, изученных в данном отношении, скажем у другого вида цихлид, Neolamprologus pulcher (Hess et al., 2016). Зарянки Erithacus rubecula, вовлечённые в территориальный конфликт, намного медленней реагировали на модель перепелятника Aciipiter nisus (Dunn et al., 2004). Королевские пингвины Aptenodytes patagonicus в гнездовой колонии при приближении наблюдателя показывали тем меньшую дистанцию бегства, чем чаще вступали в агрессивные взаимодействия с соседями (Hammer et al., 2023) и т.д.]
Animal Behaviour. 1995. V.49. P.234-239.
Рекомендуем прочесть
 Что не так с критикой Ильенкова
Что не так с критикой Ильенкова Пение самок воробьиных птиц: исключение или закономерность?
Пение самок воробьиных птиц: исключение или закономерность? Быстрая перестройка фенологии в новых местах гнездования ласточек
Быстрая перестройка фенологии в новых местах гнездования ласточек Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных Восприятие шумов
Восприятие шумов Социальная сегрегация в городах
Социальная сегрегация в городах Культ книги в РККА: как красноармеец стал главным пропагандистом грамоты в стране
Культ книги в РККА: как красноармеец стал главным пропагандистом грамоты в стране Как соотносятся прямое приспособление и эволюционные изменения?
Как соотносятся прямое приспособление и эволюционные изменения?
