
Скромный бородастик Calorhamphus hayii
Кроме дятлов, гнездовые дупла выдалбливает ещё ряд видов птиц. В гнилой или сухой древесине дупла долбит дубоносый скворец Scissirostrum dubium с о.Сулавеси; даже череп его укреплён для долбления по типу дятлового.
Гнездится колониально, сотнями пар вместе.

Дубоносый скворец
Cамостоятельно изготавливают дупла гаички Poecile spp., выщипывая их в достаточно гнилой древесине (обычно берёзовых пнях). В тропиках в ней выщипывают гнёзда трогоны (группа названа по древнегреческому trogon «грызть»; движения работающей птицы похожи на выгрызание). Иногда это дупла — трубообразные, с длинным ходом, иногда просто ниши и полудупла. Птицы проверяют, «подходит» ли древесина, простукивая ствол дерева хвостом. Ряд видов также делает гнёзда в термитниках, а 3 вида «фиолетовых трогонов» (Trogon caligatus, T.violaceus, T.ramonianus) — в гнёздах ос, покончив с их обитателями, хотя могут гнездиться в гнилом дереве либо термитнике.
Иногда изготавливают дупла поползни — каролинский Sitta carolinensis, каштановогрудый S.castanea, обыкновенный S.europaea, хотя больше занимают чужие (иногда несколько доработав). Но главное — наряду с дятлами к первичным выдалбливателям дупел, «изготовляющим» их для других обитателей (англ. primary excavators) относят их более примитивных родственников, бородастиков. Их вклад, сравнительно с дятловым, в создание «фонда» дупел на разных континентах см.табл.11.
Бородастиков делят на 4 семейства: Capitonidae и Semnornithidae (тукановые бородастики, 2 вида) в Южной Америке, Lybiidae в Африке, Megalaimidae в Южной и Юго-восточной Азии. Первые (2 рода, 14 видов) это фактически те же туканы, вторые (7 родов, 43 вида) — родня медоуказчикам, третьи (2 рода, 34 вида) — собственно дятлам: их предки точно имели «бородастиковый» уровень организации и, возможно, их образ жизни. Да и долбление дупел только у них изучено сколько-нибудь подробно, поэтому рассказ почти полностью посвящён именно им.
Таблица 1. Обзор мировой фауны дуплогнёздников: «создатели» и «пользователи» дупел
|
Отношение к изготовлению дупел |
Число видов |
В том числе селятся в дятловых дуплах |
В том числе селятся в дуплах бородастиков |
В том числе селятся в дуплах, созданных ещё кем-то кроме дятлов и бородастиковc |
|
Первичные выдалбливателиb |
355 (19%) |
0 |
0 |
0 |
|
Факультативные выдалбливатели |
126 (7%) |
49 |
3 |
2 |
|
Вторично использующие |
1357 (73%) |
289 |
50 |
9 |
|
Не определено |
40 (2%) |
0 |
0 |
0 |
|
Всего |
1878 |
338 |
53 |
11 |
Примечание. % показаны от общего числа видов птиц-дуплогнёздников; bне учтено поселение одних «производителей» дупел в дуплах, созданных другими видами той же группы (скажем, дятлов в дуплах других дятлов), что регулярно случается, cимеются в виду трогоны, поползни и синицы.
Источник. Van der Hоek et al. (2017), table 1.
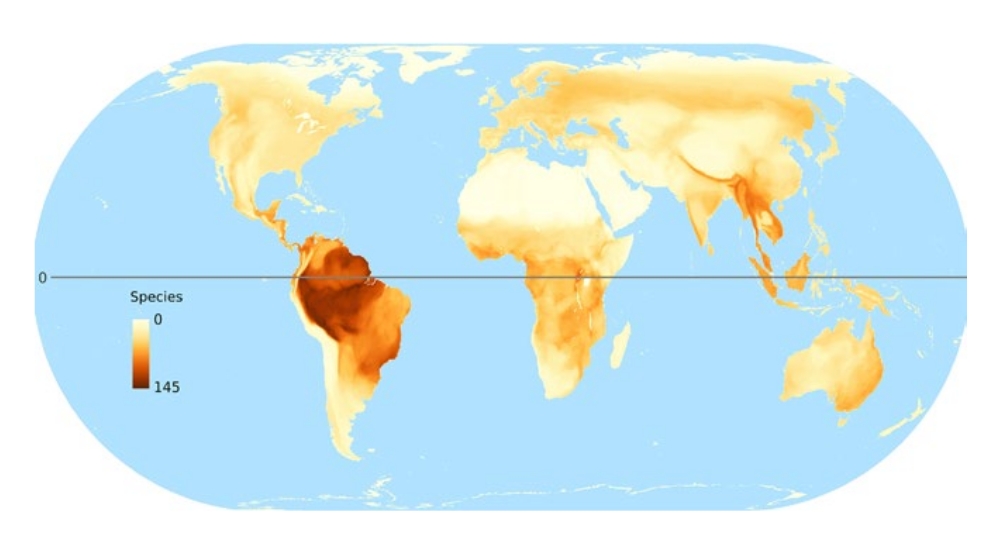
1
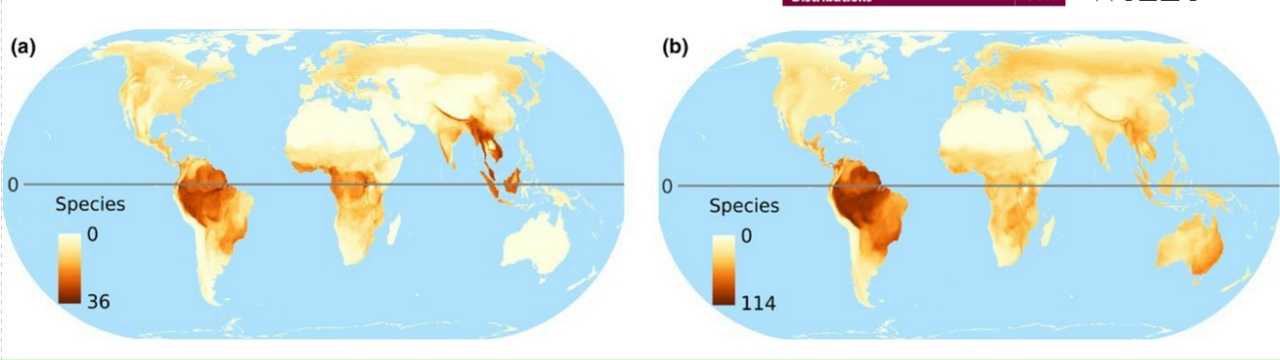
2.
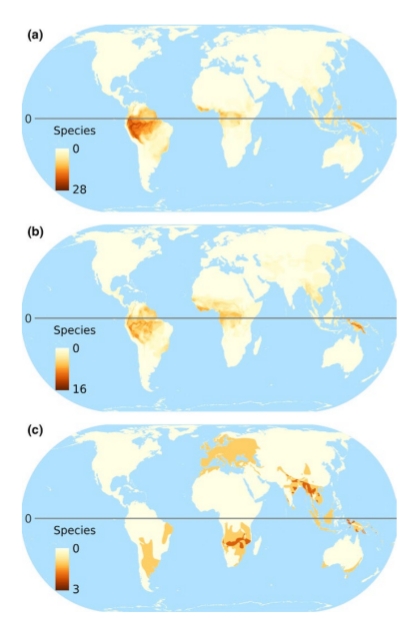
3.
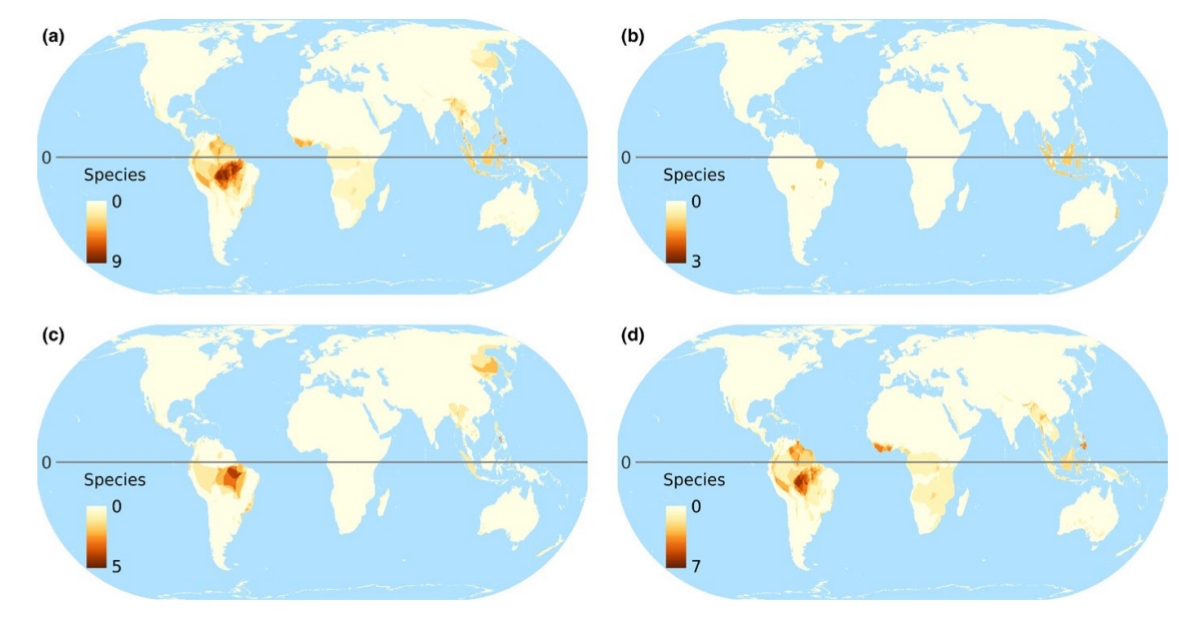
4.
5.
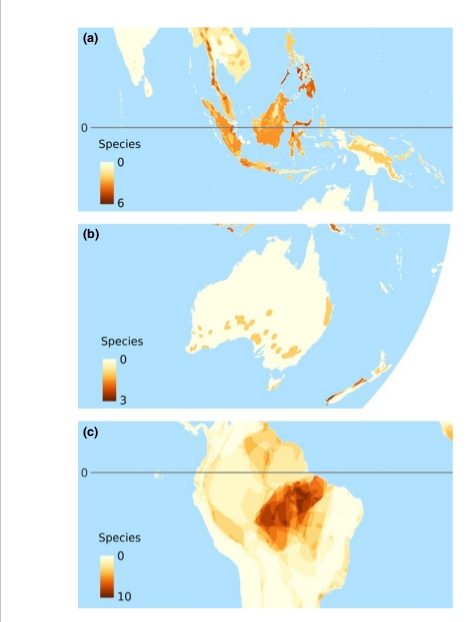
Карты мирового распределения птиц-дуплогнёздников, из van der Hoek et al. (2017). На картах: 1. Глобальное распределение числа видов дуплогнёздников всех категорий, 2. Глобальное распределение числа видов первичных (а) и вторичных (b) дуплогнёзников, 3. Глобальное распределение числа угрожаемых видов дуплогнёздников, а- общее число попавших в Красную книгу МСОП, b-d отдельно по категориям находящиеся на грани вымирания, угрожаемые и уязвимые. 4. Величина нашего незнания: глобальное распределение числа видов, для которых неизвестно, долбят они дупла постоянно или факультативно (а); неизвестно, гнездятся они в выдолбленных дуплах, в природных или в обоих типах (b); не определён природоохранный статус (с); 5. «Горячие точки» биоразнообразия краснокнижных дуплогнёздников отдельно для тех регионов, где их больше всего (сверху вниз) Юго-Восточная Азия, Австралия, Южная Америка.
Как «поставщики» дупел бородастики сравнимы с дятлами
Челюстной аппарат бородастиков своеобразно комбинирует адаптации к фруктоядности и к самостоятельному изготовлению дупел, в котором удары клювом, рвущие или размочаливающие древесные волокна, сочетаются с их выщипыванием (Трунов, 2012). На первый взгляд клюв бородастиков плохо приспособлен к долблению: кончик заострённый и часто изогнутый, конёк надклювья сильно изогнутый, сам клюв очень широкий. Однако они сами делают не только гнездовые, но и ночёвочные дупла. Последние немедля выдалбливаются на всякой вновь занятой территории, на открытом воздухе ночуют лишь расселяющиеся молодые; в пределах долговременных участков обитания, как и у дятлов, владелец имеет несколько ночёвочных дупел: будучи потревожен в одном из них, он скрывается в следующих.
Правда, бородастики больше дятлов зависят от мёртвой древесины, а подходящих мест для долбления дупел немного, их надо найти, и они определяются механическими свойствами субстрата2. Поэтому после успешного размножения в сделанном однажды дупле они стараются там же гнездиться повторно, новые долбят лишь при невозможности этого, и преимущественно на том же суку или дереве, со «знакомыми» механическими свойствами древесины.
Долбить предпочитают в сухих скелетных ветвях живых деревьев, в широком диапазоне углов ответвления, от полугоризонтального до почти прямого. Чаще всего бородатки не опираются на хвост при долблении, отсюда обычно расположение дупел на нижней стороне суков, отходящих под углами, близкими к горизонтали. Птицы их могут долбить, вися вниз головой (данные по Psilopogon henricii). Наилучшие гнездовые деревья для них крупноразмерные и достаточно старые, иначе не будет выбора «подходящих» суков. Бородастики предпочитают леса с их доминированием: при их наличии занимают даже участки лесов внутри урболандшафта, включая старые парки и ботанические сады.
Так, в приречных лесах юго-западной Индии3 «поставщики» дупел для 14 видов вторичных дуплогнёздников — три вида дятлов (большой султанский Chrysocolaptes guttacristatus, черноспинный султанский Ch. festivus, большой острокрылый Dendrocopos nanus) и три бородастиков (цейлонский Psilopogon zeylanica, красногрудый P. haemacephala и белоухий P.viridis). В 2013-2015 гг. прослеживали судьбу 376 созданных ими дупел, 1/3 из них использовалась повторно разными видами птиц (Manikandan, Balasubramanian, 2018).
Большинство таких дупел (69,28%) располагались в живых деревьях (т. е. более твёрдых и «долгоживущих»), в них появилась и большая часть естественных дупел. В 52% случаев вторичные дуплогнёздники (n=152) гнездились в дуплах, выдолбленных дятлами и бородастиками, 42% (n=108) в возникших естественно. Бородастики выдолбили большую часть выдолбленных дупел (73,05%), дятлы лишь 26,95%: наиболее значимые «поставщики» их для вторичных дуплогнёздников цейлонский бородастик и Ch.guttacristatus.
Активней всего заселяла такие дупла обыкновенная майна: из 71 гнезда 60 было в выдолбленных дуплах (34,04% принадлежали Chrysocolaptes guttacristatus, 29,78% Ch. festivus, по 10,29% цейлонским и белощёким бородастиками), и лишь 11 в естественных. Из 29 гнёзд бурой майны Acridotheres fuscus, найденных в выдолбленных дуплах, 28.12% принадлежали Chrysocolaptes guttacristatus, 21.87% цейлонскому, 18,75% белоухому бородастику, 15,62% дятлу Ch. festivus. Все гнёзда большой синицы Parus major найдены в дуплах бородастиков, 90,9% гнёзд бенгальской сизоворонки Coracias benghalensis в дуплах дятлов Chrysocolaptes guttacristatus и Ch. festivus, 10,1% — в дуплах цейлонского бородастика.
Попугай Крамера Psittacula krameri гнездился в дуплах, выдолбленных в 50% случаев дятлом Chrysocolaptes guttacristatus, в 33,33% случаев Ch. festivus и 16,66% — цейлонским бородастиком. У малабарского кольчатого попугая 94,6% гнёзд было в дуплах, выдолбленных первичными дуплогнёздниками (в том числе 52,94% Chrysocolaptes guttacristatus, 23,52% Ch. аestivus, 11,76% цейлонским и 5,88% белощёким бородастиками), и лишь 6% — в естественных. Только последние занимали 2 вида птиц-носорогов, удод, кольчатая совка Otus lettia, сорочий шама-дрозд Copsychus saularis, циорнис Тикеля Cyornis tickelliae, белопоясничный шама-дрозд Copsychus malabaricus (Manikandan, Balasubramanian, 2018).
5% дупел повторно использовалось другими животными. За лучшие дупла идёт конкуренция между разными видами птиц, главный конкурент — обыкновенная майна Acridotheres tristis. В марте и апреле 2013 г. отмечали захват майнами двух дупел, выдолбленных большимип султанскими дятлами и одного, заселённого бенгальской сизоворонкой. В апреле 2014 г. майна захватила дупло этого же вида дятлов. В 10 случаев обыкновенные и в 6 — бурые майны вмешивались в выдалбливание дятлами дупел, и в конце концов вытеснили их. Правда, конкуренция ограничена размерным классом «производителей» и «потребителей» дупел. Кластер-анализ выделяет их 5: более мелкие виды птиц сильно зависимы от первичных дуплогнёздников, более крупные также селятся в естественно возникших дуплах (при гниении древесины) (Manikandan, Balasubramanian, 2018).
Дупла отыскивали на всех стволах >50 см в обхвате, просматривая их в бинокль минимум с 3 сторон. Дупла определяли как полости в дереве, способные укрыть птицу полностью: их разделяли на выдолбленные (дятлами и бородастиками) и естественно появившиеся от выгнивания древесины, обычно распространяющихся от механических повреждений или выпавших суков. Гнездовые дупла, найденные в 2013 и 2014 гг., метили краской для дальнейшего наблюдения: их осматривали минимум дважды в неделю для отслеживания размножения, а также осматривали в следующие годы на предмет повторного заселения. Ряд «потребителей» дупел (кольчатая совка, оба вида шама-дроздов, циорнис Тикеля) исключён из анализа из-за использования атипичных полостей: неглубоких, типа ниши, или трубообразных, с широким входом и пр. (Manikandan, Balasubramanian, 2018)
Выявленная гнездовая сеть показана на рис.1 и проанализирована в табл.1.
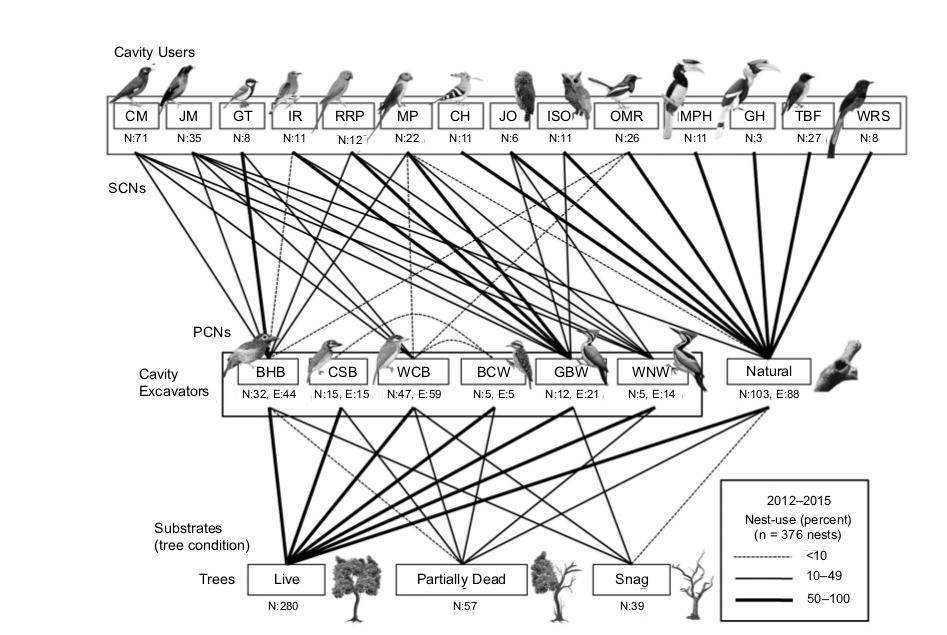 Рис.1. Гнездовая сеть показывает типы субстратов, в которых выдалбливают дупла дятлы и бородастики (нижний ряд слева направо — живые, частично усохшие и сухие). Средний ряд — число дупел, выдолбленных разными «производителями» (слева направо — цейлонским, красногрудым и белоухим бородастиком, дятлами D.nanus, Chrysocolaptes guttacristatus, Ch. festivus) или естественно (Natural). N — найдено гнездовых дупел данной категории, Е — выдолблено дупел данным видом. Верхний ряд — разные виды вторичных дуплогнёздников. Линии — число и % дупел разного происхождения, выдолбленных разными видами первичных дуплогнёздников (от нижнего ряда к среднему) или заселённых разными видами вторичных дуплогнёздников (от верхнего к среднему или внутри последнего — когда дятлы селятся в дуплах бородастиков). Толщина линий отражает число дупел каждой категории: <10, 10-49, 50-100.
Рис.1. Гнездовая сеть показывает типы субстратов, в которых выдалбливают дупла дятлы и бородастики (нижний ряд слева направо — живые, частично усохшие и сухие). Средний ряд — число дупел, выдолбленных разными «производителями» (слева направо — цейлонским, красногрудым и белоухим бородастиком, дятлами D.nanus, Chrysocolaptes guttacristatus, Ch. festivus) или естественно (Natural). N — найдено гнездовых дупел данной категории, Е — выдолблено дупел данным видом. Верхний ряд — разные виды вторичных дуплогнёздников. Линии — число и % дупел разного происхождения, выдолбленных разными видами первичных дуплогнёздников (от нижнего ряда к среднему) или заселённых разными видами вторичных дуплогнёздников (от верхнего к среднему или внутри последнего — когда дятлы селятся в дуплах бородастиков). Толщина линий отражает число дупел каждой категории: <10, 10-49, 50-100.
Источник. Manikandan, Balasubramanian, 2018.
Таблица 1. Сравнение дупел, используемых повторно или нет, в зависимости от состояния дерева и типа дупла.
|
Параметры, % |
Гнездовые дупла 2013 & 2014 года |
χ2 |
Р |
|
|
Повторно используемые (n=95) |
Не используемые (n=197) |
|||
|
Состояние дерева - живое -частично усохшее -мёртвое (сухостой) |
66,66 18,75 14,58 |
80,20 9,64 8,15 |
48,25 |
<0,000 |
|
Тип дупла -выдолблено дятлами -выдолблено бородастиками -естественно возникло |
40,62 44,79 14,68 |
23,86 41,12 35,03 |
15,43 |
<0,000 |
|
Тип субстрата -живой мёртвый |
62,50 37,50 |
42,13 57,87 |
6,00 |
<0,01 |
Рис.2. Цейлонский бородастик
Как бородастики освоили урболандшафт
Ряд видов бородастиков успешно урбанизировался в г.Сингапуре, на Тайване, в агломерациях Индии и в других местах. Увы, большинство наблюдений за долблением дупел у бородастиков выполнено в ботанических садах и городских парках, над небольшим числом легко урбанизирующихся видов. Строительство и использование дупел в коренных сообществах — влажных тропических лесах, плакорных или приречных, саваннах и пр. изучено много хуже.
 Рисунок 3. Красноголовый бородастик на священном фикусе
Рисунок 3. Красноголовый бородастик на священном фикусе
Красноголовый бородастик Psilopogon haemacephala indica в Сингапуре (рис.3) гнездится в сезон дождей, когда корма наиболее обильны. Долбление дупел отмечено с декабря по июль, молодые с июня по сентябрь. Во время ухаживания птицы много поют, топорщат «бородой» оперение подбородка, а головы — поднимают гребнем. Оно также включает сеансы демонстративного кормления самцом самки: он предлагает ей ягоды в дразнящий манере, а та возвращает их. Вне сезона размножения держится поодиночке, ночевочные дупла также использует одна птица. Начало сезона размножения маркировано появлением пар, совместно (по очереди) выдалбливающих дупло. Хотя львиную долю этой работы производит самец, это совместное долбление — важная часть ухаживания, присутствующая и у дятлов.

Рисунок 4. Взрослый кормит птенца соплодиями фикуса
P.h.indica при долбении использует хвост как опору. Ночует исключительно одиночно, и дупла для этого делает в одиночку: гнездовое же пара долбит совместно. Дупло с летком 4-5 см располагается на высоте 2-15 м, в кладке 3-4 яйца, обычно имеет 2 кладки за год. Вид часто использует старые дупла, проинспектировав их и очистив гнездовую камеру. В Сингапуре неизменно гнездится в сухих сучьях живых стволов, также выдалбливает дупла в мёртвых стволах пальм (рис.4). 16 апреля 2007 г. в Jalan Keli отмечен выдалбливающим дупло в сухом стволе красной сургучной пальмы Cyrtostachys renda, который казался слишком узким для этого вида (Loke, Lee, 2009). Внутри гнездового дупла ночуют оба партнёра.
Красноголовый бородастик заселил Сингапур самостоятельно, при расселении по Малайскому п-ову с севера на юг. Подвид indica избегает сомкнутых крупных массивов, при поселении в них держится у опушек или вырубок, предпочитает «парковые» насаждения близ населённых пунктов или в них. Его расселение было стимулировано преобразованием ландшафта, вид занимал фрагментированные леса, старые парки, сады и кладбища, коттеджную застройку, окружённую лесными микрофрагментами, вроде принадлежащей Совету по жилищному строительству. Сохранившихся участков первичных лесов избегает, также как старовозрастных вторичных. Эти же местообитания он занял и в Сингапуре, где гнездится с 1957 г. став самым массовым бородастиком. Поскольку город-государство вид заселил сам, его отнесли к аборигенной фауне, в отличие от P.lineata hodgsoni: его население в Сингапуре происходит от птиц, выпущенных из неволи (Lok, Lee, 2009).
Вид исключительно толерантен к людям и технике, спокойно кормится в их присутствии, гнездится недалеко от источников антропогенного беспокойства. Скажем, он часто кормится на низкорослых деревьях бодхи непосредственно на автостоянке (распространившихся из японского сада поблизости). В Сингапуре гнездится в разных деревьях и на разной высоте: в альбизии (Falcataria moluccana) на высоте 15 м в природном парке Букит Баток, в акации ушковидной (Acacia auriculiformis) (10 м) на кладбище Bukit Braun, в жакаранде туполистной (Jacaranda obtusifolia) (высота 10 м) в Китайском саду, в цезальпинии ржавой (жёлтое пламя) Peltophorum pterocarpum (5 м), в акации ушковидной на мусульманском кладбище Lim Chu Kang и во фагрее душистой Fragraea fragrans в Сингапурском ботаническом саду (20 м). Гнезда везде выдолблены в суках живых деревьев (Lok, Lee, 2009).
Известны случаи нетипичной ночёвки вида в городах, даже при наличии лесопарков и подходящих деревьев. В Ludhiana, Пенджаб, птица долгое время (с 15 августа по 20 ноября, пока шло наблюдение) ночевала в полости бамбукового шеста, поддерживающем провода кабельного ТВ, залезая туда сверху. Бородастик держался здесь, хотя в 150 футах были два городских парка. Его не пугали фейерверки и детские вопли на улице, особенно на праздниках Dusshera (2.10) и Divali (29.10) (Kler, 1994). Другой нетипичный способ ночлега отмечен в центре Цейлона, примерно в 2 м от второстепенной дороги близ г.Kambarawa. Взрослый самец ночевал в нише бетонного столба ЛЭП ~8 см в ширину и 15 см в высоту, на высоте 6 м от земли (рис.5). Местные жители сообщили, что птица ночует здесь уже 6 месяцев (Karunarathna, Amarasinghe, 2008).

Рисунок 5. Ниша в бетонном столбе, используемая для ночлега, а — общий вид, b — увеличено.
Залитые водой дупла бородастики используют для купания.
По наблюдениям в Сингапуре (Lok et al., 2009a), полосатый бородастик P.lineata также долбит и ночевочные, и гнездовые дупла дупла в мёртвой древесине сухих скелетных сучьев. Насиживание удлиняется в холодные и дождливые дни, иногда до 11 ч. Авторы считают, что от наличия соответствующих деревьев вид зависит не меньше, чем P.haemocephala. Однако Wells (2002) отмечал предпочтение живых стволов и сучьев для гнездования (видимо, с сердцевинной гнилью).
 Рисунок 6. Начало выдалбливания дупла полосатым бородастиком в суке альбизии4 (Falcataria moluccana), что важно, в месте, где от повреждения внутрь распространилась гниль.
Рисунок 6. Начало выдалбливания дупла полосатым бородастиком в суке альбизии4 (Falcataria moluccana), что важно, в месте, где от повреждения внутрь распространилась гниль.
Вид эвритопен. Его отмечали и в сомкнутых лесах (хвойных, включая сосновые, вечнозелёных и листопадных), и в редколесьях (перелесках, парковых вторичных лесах), на плантациях и в плодовых садах, особенно в достаточно старых, сомкнутых, обильно плодоносящих. В Индии, давно освоенной человеком, с терпимым отношением к дикой природе, многие исходно лесные бородастики, включая полосатого, перешли на плодовые деревья в городах и придомовые сады, где могут стать вредителями. Это же произошло в Сингапуре, где P.lineata сперва держался лишь во вторичных лесах, а теперь проникает в районы жилой застройки, на скверы и бульвары. Птиц привлекают плодоносящие декоративные деревья, посаженные Советом национальных парков.
Чаще всего гнездится в мёртвых ветвях альбизии (рис.6). Несколько необычное гнездо выстроено на высоте 3 м в коричнике вялом Cinnamomum iners, за общественным туалетом, немного вглубь леса. Птицы уже насиживали, делая это поочерёдно. Из-за обработок инсектицидами против комаров они приносили птенцам только фрукты, не насекомых, и смогли вывести лишь 1 птенца, тогда как обычно их 2-4. (Lok et al., 2009a)
Анализируя размножение собственно лесных видов бородастиков в Сингапуре, Lok et al. (2009b) отмечают, что многоцветный бородастик P.rafflesii также зависит от деревьев с обилием сухих скелетных сучьев, в которых выдалбливает дупла. Раньше еë размножение было малоизучено. Отмечали лишь предпочтение древесины, сильно размягченной гнилью, для долбления дупел (рис. 7), в том числе в стволах сухостоя на высоте 5–8 м. В выводке 1-2 птенца, размер кладки был неизвестен (Wells, 1999). В Сингапуре дупла выдалбливали оба партнёра, обычно работая более чем над одним дуплом одновременно. Способен подобно дятлу выдалбливать личинок ксилофагов в гнилой древесине.
 Рисунок 7. Многоцветный бородастик долбит дупло
Рисунок 7. Многоцветный бородастик долбит дупло
 Бурый бородастик посещает фиговое дерево на Майлайском п-ве
Бурый бородастик посещает фиговое дерево на Майлайском п-ве
Синеухий бородастик P.australis как правило, гнездится выше, также в стволах мертвых деревьев, но на высоте от 3 до 25 м, диаметр около 3 см, в кладке обычные 2–4 яйца (Shorts & Horne, 2002). Ухаживание включает в себя непрекращающееся пение обоих членов пары, сильное покачивание головой и движения хвоста из стороны в сторону с ухаживательным кормлением. В отличие от видов рода Psilopogon, бурый бородастик Calorhamphus fuliginosus полуколониален: три и более пар гнездятся вместе в стволах и или в суках мертвых деревьев, а также в древесных термитниках на высоте 1–20 м (Wells, 1999), а также в корневице папоротника костенца гнездового (Asplenium nidus). Его кладка 2-3 яйца, выводок — 2-3 птенца (Wells, 1999; Shorts & Horne, 2002).
Эти два вида исчезли, не сумев приспособиться ни к трансформации лесов растущим городом, ни к растительности последнего. Последние встречи относятся к 1921 г., так что P.rafflesii остался единственным видом коренных лесов в фауне Сингапура (рис.8).
 Рисунок 8. Смена родителей у гнезда многоцветного бородастика: один принёс корм, другой выносит помёт
Рисунок 8. Смена родителей у гнезда многоцветного бородастика: один принёс корм, другой выносит помёт
С.-N. Koh & F.-C. Lu изучили размножение тайваньского бородастика P.nuchalis с марта по сентябрь 2008 г. в ботаническом саду Тайбея. Найдено 27 дупел на 12 деревьях, т.е. данный вид также особо привязан к наиболее подходящим мёртвым сукам (реже стволам), почему их нельзя убирать из городских лесов. Гнездовые деревья относятся к 8 видам, в основном это камфора Cinnamomum camphora, среди них почти нет однодольных. Леток круглый, точно в размер птицы, из-за чего она пролезает с трудом, ориентирован с северо-запада на юго-восток, что обеспечивает хорошую вентиляцию и сухость в дупле [снижая риск заливания гнезда муссонами]. Выдабливание гнездового дупла P.nuchalis растягивается на 22 дня. Самцы также долбят больше самок, 17.6 ± 1.7 мин/ч в сравнении с 6.9 ± 0.8 мин/ч, данные по 4 гнёздам (Lin et al., 2010).
В тропических лесах юга Вьетнама (лес Ма Da, провинция Dong Nai) синеухий бородастик P.australis гнездится в мёртвых, часто обломанных скелетных ветвях, угол отхождения которых варьирует от полугоризонтального (350) до почти вертикального (80-900). См. табл.1 в Трунов, Васильева, 2012. Дупла часто долбят их нижней стороне. Примерно у половины гнёзд рядом с действующим дуплом расположены 1-2 старых, возможно прошлогодних, т. е. каждый год птицы выдалбливают новое но поблизости. Гнездостроение происходит в сухой сезон, с января по март, выдалбливание дупла занимает 6-14 дней (Трунов, Васильева, 2012).
Токующий самец сидит вертикально; кричит, вытягивая шею высоко вверх. При криках становится виден на горле раздувающийся мешок тёмной кожи, лишённой перьев, и в этом момент песня звучит громче.
 Синеухий бородастик
Синеухий бородастик
Подобно дятлам, бородастики при необходимости резко интенсифицируют усилия по выдалбливанию, и время изготовления сокращается в разы (хотя всё же не до одного дня, как у дятлов). Делает это почти исключительно самец. Места для гнезда выбирают оба партнёра, опять же при активной роли самца. Перемещаясь по ветвям в дятловом стиле, он в каждом привлекшем его месте совершает 2-3 лёгких постукивания5. Далее совершается несколько пробных попыток долбления в разных, расположенных рядом местах одно из которых становится окончательным. Их следы в виде мелких воронок глубиной до 1 см. соответствуют инициальным дуплам дятлов. Самка при этом держится поблизости и наблюдает, управляя самцовым выбором тем что приближается или нет к привлекшим его местам (Трунов, Васильева, 2012).
Выдалбливание дупла проходит последовательно 4 стадии: начальной воронки (при неподходящей древесине долбление дальше не идет), обработки летка, выборки гнездовой камеры; завершающей обработки дупла. На первых стадиях опилки и щепки птицы выгребают клювом и бросают наземь прямо у места долбления, на поздних, набрав в клюв, отлетают на соседние ветви и бросают уже там (Трунов, Васильева, 2012).
Помимо гнездовых дупел, бородатки долбят также ночевочные, на каждого члена пары. Старое гнездовье дупло может использоваться в этом качестве, наоборот никогда. Долбление дупла, как и у дятлов, движется вперёд понемногу, но каждый день, с большими перерывами на отдых и кормёжку. К концу строительства ее интенсивность растёт, см. табл.3 из Трунов, Васильева, 2012.
 У P.australis оформление летка и выборку большей части внутренней камеры неизменно производил самец, как и у других видов семейства. Самка лишь держалась поблизости; на 3м этапе она заинтересовалась дуплом и, пока самец не долбил, по нескольку раз в день подлетала к летку, заглядывала туда, пробовала забраться внутрь. Это происходило несколько раз в день. Когда гнездовая камера увеличилась до размера самки, она закончила общую работу. Отбивая и выщипывая волокна, она расширяла дупло, обустраивала его изнутри: тогда же самка начала ночевать в дупле. Самец также участвовал в строительстве на этой стадии. Если самец сбрасывал щепу прямо под дуплом, самка относила её в кроны соседних деревьев и бросала там (Трунов, Васильева, 2012).
У P.australis оформление летка и выборку большей части внутренней камеры неизменно производил самец, как и у других видов семейства. Самка лишь держалась поблизости; на 3м этапе она заинтересовалась дуплом и, пока самец не долбил, по нескольку раз в день подлетала к летку, заглядывала туда, пробовала забраться внутрь. Это происходило несколько раз в день. Когда гнездовая камера увеличилась до размера самки, она закончила общую работу. Отбивая и выщипывая волокна, она расширяла дупло, обустраивала его изнутри: тогда же самка начала ночевать в дупле. Самец также участвовал в строительстве на этой стадии. Если самец сбрасывал щепу прямо под дуплом, самка относила её в кроны соседних деревьев и бросала там (Трунов, Васильева, 2012).
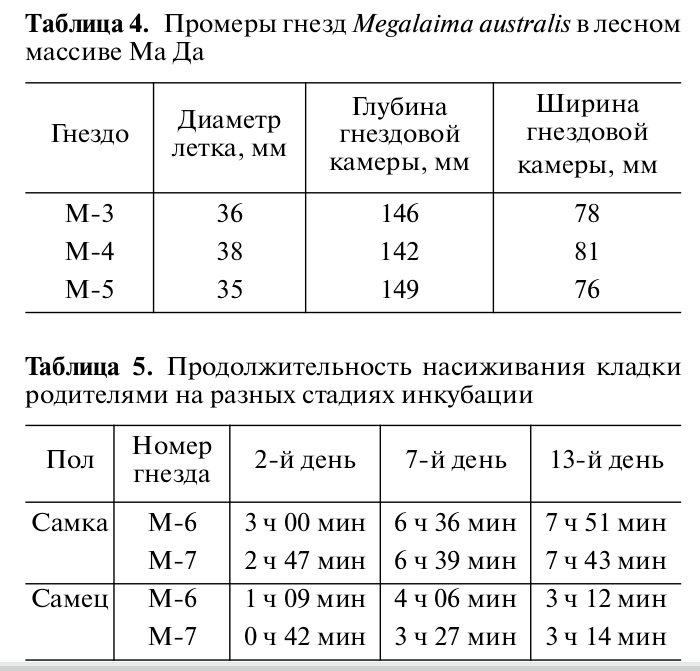
По данным обследования трех гнезд (табл.4), диаметр летка в среднем 39 ± 1 мм; гнездовая камера довольно правильной овально-вытянутой формы, ее глубина от нижнего края летка до самой глубокой точки дна 146 ± 2.4 мм, диаметр в самой широкой части 78 ± 1.8 мм. В отличие от дятлов, дно дупла чистое, без подстилки из слоя щепок, обдираемых со стенок дупла.
Ухаживание наблюдали только в течение 2х дней по завершению строительства. Самка преследовала самца, провоцируя короткие (15-25 с) погони с перелётами. Догнав, она мягкими, неагрессивными движениями хватала самца за клюв. В паузах самец издавал торопливые брачные крики, самка медленно покачивала из стороны в сторону поднятым вверх хвостом. Спаривание отмечено близ гнезда, на 5й день ночёвки самки в готовом дупле. Самец произвёл 6 садок по 3-7 с в течение 3 ч, с 8:08 до 11:12. После каждой самец отлетал, приносил плоды фикуса и кормил ими самку; 3 раза кормление отмечено вне связи с копуляциями. Весь этот день (с 6:07) до захода на ночёвку (16:23) самка держалась в одном секторе кроны (Трунов, Васильева, 2012).
Насиживают оба партнёра, сперва примерно с равным участием, на более поздних стадиях инкубации самка сидит заметно дольше (табл.5). Оба партнёра регулярно сгоняют чужих особей с деревьев, в которых потом построят дупло: видимо, они более привлекательны для всех особей данного вида, чем наугад выбранные, и даже в тропическом лесу они в дефиците, не только в городских парках или в антропогенном ландшафте. После начала выдалбливания чужаков на участках не отмечали (Трунов, Васильева, 2012).
В обзоре литературы авторы пишут, что у Psilopogon spp. дупла строят оба партнёра, при ведущей роли самца (Yahya, 2001; Lin et al., 2010). У P.australis он делает самое трудное – обработку летка и выборку основного объема гнездовой камеры, самка присоединяется только в конце, полностью отстраняя партнера.
В выборе места для гнезда также участвуют оба партнера, хотя самец более активен: основные критерии здесь толщина ветки и состояние древесины. Поскольку бородастики всегда строят дупла в мертвой древесине, видимо, значима степень ее разложения. По наблюдениям авторов, бородастики предпочитают не слишком старые мертвые стволы с прочным наружным слоем и более рыхлой сердцевинной частью, которая поддается несильным ударам и выщипыванию. Долбление пробных воронок позволяет оценить качество древесины и ее пригодность для строительства гнезда (Трунов, Васильева, 2012).
Продолжая обзор, авторы пишут, что мелкие бородастики долбят дупла в суках и стволах значимо тоньше, чем крупные (Yahya, 2001). Вообще, они предпочитают гнездится в ветвях с диаметром, минимально возможным для птиц их размера. Возможно, это снижает конкуренцию за дупло, ограничивая её участников лишь птицами меньше самих строителей, и не пускает вовнутрь белок, крупных дятлов и других потенциально опасных существ (Yahya, 2001). Средняя толщина ветвей с дуплом P. australis 16 см, при длине тела птицы 16–17 см. Возможно, бородатки испытывают дефицит наилучших деревьев для гнездостроения, по сравнению с дятлами им куда тяжелей долбить дупло в древесине, субоптимальной по качеству, поэтому они чаще используют одни и те же же суки несколько сезонов размножения подряд, так что на них видишь дупла разных лет постройки. [Подобная «многодупельность» наилучших гнездовых деревьев присуща и дятлам, как и стремление при первой возможности повторно использовать свои прошлогодние дупла. Прим.публикатора]
 Зеленоухие бородастики P.faiostrictus, смена на гнезде
Зеленоухие бородастики P.faiostrictus, смена на гнезде
Из бородастиков южного Вьетнама многодупельность наиболее подходящих сучьев присуща чаще всего P. haemacephala (Short, Horne, 2001; данные авторов) и P.faiostricta (данные авторов). У Р. australis трижды отмечены старые летки (1, 2 или более) близ жилого дупла. Везде дистанция между летками минимум вдвое превосходила среднюю глубину гнездовой камеры (табл. 4). Вероятнее всего, старые и новые дупла всегда были разобщены (Трунов, Васильева, 2012).
Размер кладки авторы не установили из-за слишком большой высоты дупел, не допускающей прямое обследование. По данным литературы, кладки Psilopogon spp. насчитывают от 2 до 4 яиц, чаще 2–3 (Wells, 1999; Short, Horne, 2002). Авторы отмечали 1– 2 слетка в каждом из 5 найденных гнезд. Однако кладки и выводки бородастиков часто гибнут от хищников или других причин (Yahya, 2001; Lin et al., 2010). С учётом этого кладки P. australis на юге Вьетнама могут быть больше (Трунов, Васильева, 2012).
Бородастики охраняют гнездо, избирательно реагируя на появление поблизости разных видов птиц. Тревога и/или агрессия возникает лишь при появлении других дуплогнездников: скворцов, дятлов (Yahya, 2001).
 Белоухий бородастик
Белоухий бородастик
Биология белоухого бородастика Psilopogon viridis изучена близ г.Kollam в индийском штате Керала в 1995-2018 гг.; всего найдено 18 гнёзд (Roopa, 2018). На исследуемой площадке в пол-акра гнездилось не более 1 пары этого вида, но его гнёзда находили на соседних участках (их не включали в анализ). Минимальное расстояние между гнёздами 50 м. Дупла выдалбливали в основном в сухих стволах кокосовых пальм Cocos nucifera, также как в усыхающих стволах бетелевой пальмы Areca catechu. Гнездовые дупла также выдабливали в сухих ветвях живых деревьев альбизии Albizia saman, кэшью Anacardium occidentale, мелкоплодного фикуса Ficus microcarpa и т. д., а также в живом стволе дерева кешью с гнилой сердцевиной (рис.9).
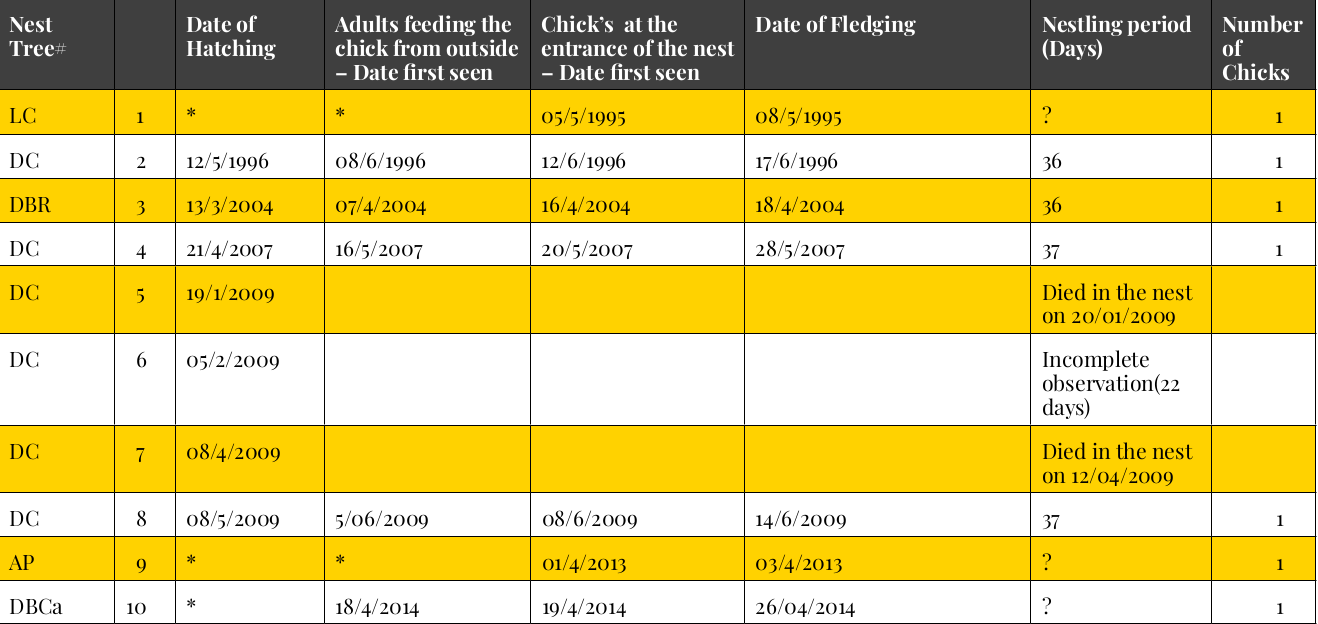
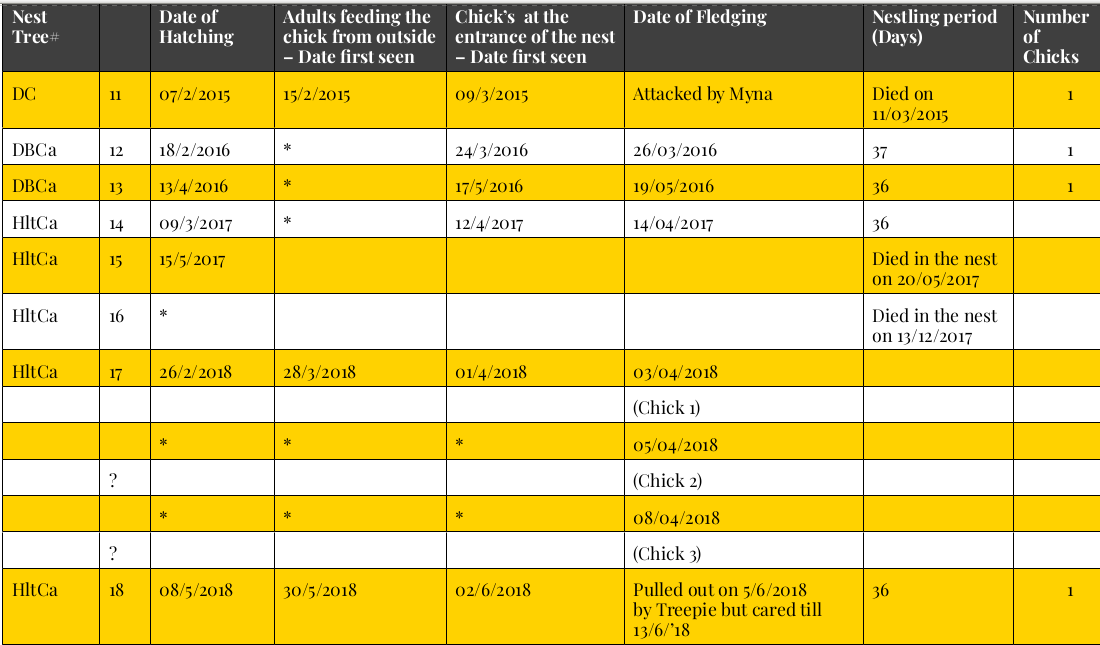 Рисунок 9. Деревья, использованные для строительства дупел, и фенология гнездования в них.
Рисунок 9. Деревья, использованные для строительства дупел, и фенология гнездования в них.
Обозначения. # LC= живая кокосовая пальма; DC= сухая кокосовая пальма; DBR= мёртвый сук альбизии; DBCa= мёртвый сук кэшью; AP= дерево бетелевой пальмы; Hlt Ca= живой ствол кэшью. *= точная дата неизвестна. Столбцы слева направо: вид и состояние гнездового дерева, его порядковый №, дата вылупления, дата, когда впервые отметили кормление птенцов вне летка, даты первого появления птенцов в летке, дата вылета, длительность гнездования, дней, число птенцов.
Брачные партнёры постоянно обмениваются двумя типами криков, звучащих как kotroo kotroo или kuroo kuroo (второй начинается со взрывного kurr), в особенности утром и вечером. Второй крик чаще всего слышен в ответ на первый и у самцов, и у самок; реже вторая птица повторяет крик первой. Крики гнездящихся пар стимулируют кричать их соседей. После вылупления птенцов родители молчаливы, кричат лишь изредка: крики kotroo возобновились лишь после вылета слётка. Крики kirr kirr отмечены при взаимодействии со своим партнёром или при изгнании от дупла других бородаток, майн и коэлей. Во время насиживания оставляющая гнездо птица иногда издавала звуки kueee kueee. Звуком khe khe родитель предупреждает птенцов об опасности (Roopa, 2018).
Найденные дупла располагались на высоте 6,6-10 м (20-30 футов). Их выдолбили в сухих или усыхающих стволах, либо в сухих скелетных сучьях, или в живом дереве с сердцевинной гнилью. Обычно у этого вида оба партнера долбят на равных; при смене приходящая птица кормит работающую. Это отметил и Hafiz Yahya (1989), чьи работы по биологии бородастиков в Индии стали классическими. Если выбранное птицами дерево было срублено, повторно долбит дупло в основном самец. Работа тогда весьма интенсивна, без перерыва по часу и более, а отдых между долблением составляет минуты.
Рисунок 10. Гнездо белоухого бородастика в кокосовой пальме
Белоухому бородастику (как и другим видам) явно не хватает подходящих гнездовых деревьев, подходящих для выдалбливания. Им, как и дятлам, особенно трудно долбить недавно усохший ствол кокосовой пальмы: он не используется 2 года и более, и только потом там появляются дупла (рис.8). [В подобных стволах] они долбят дупла сперва на верхушке дерева, где древесина помягче (высота 16,6-20 м, 50-60 футов), однако используют их как ночёвочные (и никогда в них не размножаются). Гнездовое дупло строится в середине ствола, где он толще (Roopa, 2018).
При отсутствии подходящих деревьев на своей территории, пары ищут их у соседей, пробуют выдолбить там, рискуя навлечь на себя территориальную агрессию хозяев, или отнять дупла соседей либо ждут вылета их птенцов, после чего сразу занимают дупло — как пара №1 в 2016 г. (рис.9). Если гнездовое дупло [после размножения осталось], бородастики там гнездятся повторно на слелдующий год, реже делают так и на третий. Размножение три сезона подряд (2016, 2017 и 2018 гг.) отмечали в дупле, выдолбленном в стволе кешью. Сперва его выстроили в небольшом сухом суке: когда тот в сезон дождей сгнил и отвалился, обнажилась гнилая сердцевина ствола [и возможность расширить туда дупло]. Начав долбить с этого места, птицы в 2017 году выстроили дупло в стволе, его же использовали в 2018 г, однако оставили в следующем, после расширения летка дятлами (Roopa, 2018).
Старых ночёвочных дупел на контролируемой территории больше, чем гнездовых. Когда при заходе на ночлег поздним вечером бородастик обнаруживает, что дупло уже занято (другим бородастиком, птицей или белкой), то не пробует отобрать, а летит прочь, в другое незанятое дупло [видимо, при постоянном обитании существует «фонд» незанятых дупел на территории пары, их расположение владельцам известно, почему они могут быстро добраться до них в темноте, не тратя время на поиски. Если нет, по необходимости ночуют открыто как в примере дальше. Прим.публикатора]. Автор однажды ночью нашёл бородастика, сидящего на оконнной решётке его дома: потревоженный, он улетел в темноту. У молодых ночёвочные дупла очень мелкие (Roopa, 2018).
Помимо защиты от конспецификов, P.viridis яростно защищают гнездо от других дуплогнёздников: майн Acridotheres tristis, джунглевых воробьиных сычей Glaucidium radiatum и малых индомалайских дятлов Dinopium benghalense. Дятлы используют дупла для ночлега, расширив леток: за них дятлы постоянно дрались в сезон юго-западного муссона. Гнездовое дупло они, как правило, строят сами. Гибел гнёзд заставляет пару делать повторную кладку, часто — ещё одну. Также бывают вторые кладки у рано гнездящихся пар (Roopa, 2018).
Кормление строящего гнездо партнёра при смене показывает, что у бородаток, как и у дятлов, выдалбливание дупла — часть процесса ухаживания, а не собственно размножения, сперва за потенциальным партнёром, а потом в образовавшейся паре для её консолидации. Такое кормление важная часть ухаживания у бородаток, у дятлов отсутствует, возможно заместившись совместным долблением. Как и у дятлов, успешное продвижение строительства связано положительной обратной связью с перерывами на ухаживание, позже и спаривание.
Гнездовые деревья P.viridis выбирает на участках с сомкнутыми пологом, видимо, там ниже заметность хищников (самого гнезда или слётков в момент вылета). Они не гнездятся ни изолированных сушинах кокосовой пальмы на бедных почвах, ни в рощах, где растут только они [там просматриваемость чересчур высока], но иногда долбят там ночёвочные дупла. Долбление гнездовых дупел в кокосовых пальмах отмечено только на тех участках, где они перемешаны с другими деревьями. Их летки обращались на юг, юго-запад, либо на юго-восток, возможно, для избегания ветров в период муссонных дождей (вначале они дуют с запада и севера). В сухом суку, отходящем наклонно, дупло долбят с нижней стороны. Тогда он бывает направлен на север, ибо сук прикрывает дупло от дождя. Летки у ночёвочных дупел не ориентированы в какую-то определённую сторону (Roopa, 2018).
В начале строительства гнезда птицы бросают опилки и щепу прямо у входа (боковым движением головы). На поздних стадиях, при выборке гнездовой камеры, они с ними в клюве слетают на ближайшую ветвь дерева и бросают там. В 3-х случаях гнездовое дупло и ночёвочное второго партнёра находилось на той же кокосовой пальме. В парах, где получилось определить пол, самка ночевала в гнезде, самец в ночёвочном дупле [у дятлов обычно наоборот. Прим.публ].
Найдено 14 ночёвочных дупел, 10 в кокосовых пальмах, по одному на фиговом дереве, индийском хлебном дереве Artocarpus heterophyllus, манго Mangifera indica и светлоголовчатой леуцене Leucaena leucocephala. 5 из 14 дупел в кокосовой пальме сильно прогнили из-за множественного использования ночующими майнами, сычиками или дятлами.
Ночёвочные дупла в кокосовой пальме внутри имеют форму фляги глубиной от летка до дн) 30 см, внутри они очень гладкие. Передняя стенка тонкая, отделена от внешнего мира только корой пальмы, задняя толще (со слоем древесины) и сужается кпереди. Входное отверстие камеры располагалось в верхней части камеры, где невыдолбленный ствол пальмы контактирует с раздолбленной камеры. Леток ночёвочного дупла мал (4,5 см диаметр) сравнительно с гнездовым, и открывался прямо во внутреннюю камеру (не было ведущей туда трубки). Ширина дупла была наибольшей непосредственно надо дном (16 см), внизу оно имело форму чаши или блюдца (Roopa, 2018).
Найдено ночёвочное дупло в сухом суку фикуса мелкоплодного (Ficus microcarpa) толщиной 32,5 см в окружности. Леток 4,5 см в диаметре — много меньше, чем у гнездового дупла — направлен на север, он открывался во внутреннюю камеру напрямую (не было трубковидного хода). Камера колбовидная глубиной 30 см, её передняя стенка толщиной 1,5, задняя 2,5 см, ширина в наиболее широкой части 9 см, низ внутренней камеры чашеобразен.
Гнездовое дупло имело вид фляги (реторты, как упоминает Юм, 1890) и окружено толстыми стенками (наиболее толстыми вокруг туннеля от летка в камеру, наиболее тонкими непосредственно над её нижней частью). Его внутренняя камера меньше, чем у ночёвочного, а стенки более гладкие; промеры показаны на рис.11. Входное отверстие вело к туннелю в его центре через толстую стену и переходило в камеру. Стенка камеры была самой толстой вокруг туннельной части и самой тонкой непосредственно над нижней частью камеры в форме чаши/блюдца. Задняя часть патронника была толще передней. Туннельный ход был воронковидным в разрезе, он расширялся в направлении от летка к гнездовой камере (Roopa, 2018).
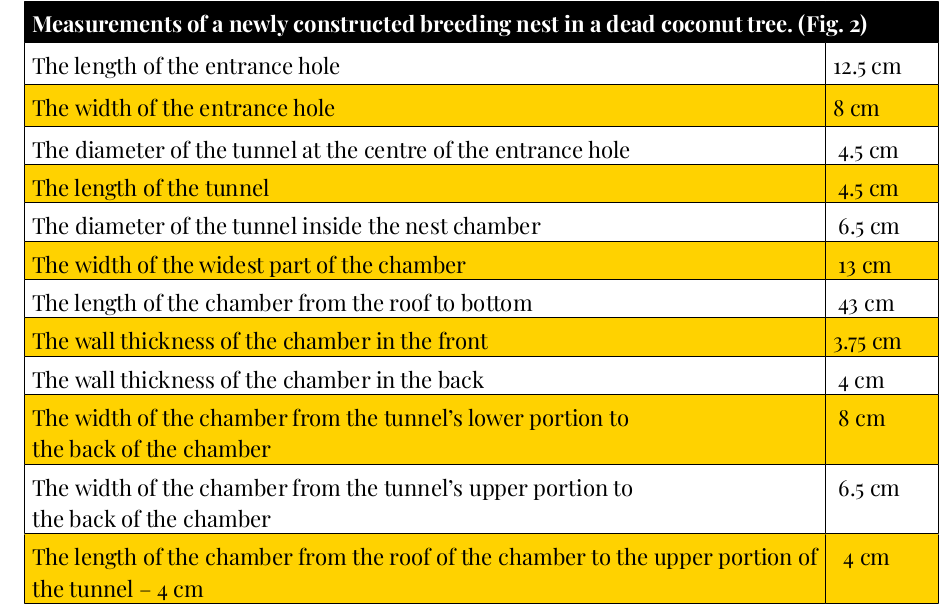 Рисунок 11. Параметры только что выдолбленного гнездового дупла в сухой кокосовой пальме. Строчки сверху вниз: длина входного отвествия, см.; его ширина, диаметр туннельного хода в месте входа во внутреннюю камеру, его общая длина; его диаметр за пределами внутренней камеры; ширина самой широкой части камеры; наибольшая длина внутренней камеры сверху вниз; толщина передней стенки дупла; толщина задней части дупла; ширина верхней части внутренней камеры, от отверстия туннельного хода до противоположной стенки; длина верхушки камеры, от её крыши до отверстия туннельного хода.
Рисунок 11. Параметры только что выдолбленного гнездового дупла в сухой кокосовой пальме. Строчки сверху вниз: длина входного отвествия, см.; его ширина, диаметр туннельного хода в месте входа во внутреннюю камеру, его общая длина; его диаметр за пределами внутренней камеры; ширина самой широкой части камеры; наибольшая длина внутренней камеры сверху вниз; толщина передней стенки дупла; толщина задней части дупла; ширина верхней части внутренней камеры, от отверстия туннельного хода до противоположной стенки; длина верхушки камеры, от её крыши до отверстия туннельного хода.
Пары сохраняются и на внегнездовое время. В 2009 г. в одной из них автор мог идентифицировать самца и самку, благодаря чему наблюдал их особенности захода на ночлег. Перед заходом в дупло оба издавали серии криков. После этого самец сразу заходит на ночлег (обычно уже совсем темно), самка ещё ждёт, и уходит в дупло сильно позже него (18.30 в январе, 19.00 в мае), когда её точно не видит никто. В любой ситуации, когда можно думать, что птица или зверёк это заметили, она вылезает наружу, и издаёт крики kuk-kuk, выживая, пока всё не успокоится. Залезает в дупло самка медленно, многократно выглядывает наружу. Самец и самка из гнездовой пары иногда спали в разных гнездовых дуплах, когда те находились на одном дереве. Утром партнёры покидают дупло также затемно — в 5.45 в мае, в 6.30 в январе, когда едва-едва рассветает, и сразу обмениваются криками (Roopa, 2018).
В паре, наблюдавшейся в 2009 г., самец кормил самку во время ухаживания и насиживания. Он протягивал ей насекомое или плод в клюве, она его схватывала (корм никогда не вкладывал в клюв самке). Перед или во время кормления самка никак не демонстрировала. При ухаживании самец времая от времени тёрся о самку телом и головой, чистил ей перья. Trounov & Vasilieva (2014) отметили эти же формы поведения у красногузых бородастиков P.lagrandieri, наблюдавшихся ими в южном Вьетнаме (лес Loc Bac, провинция Lam Dong).
Всего отмечали 9 копуляций, происходивших без каким-либо предшествующих демонстраций. Кормления партнёра не отмечено ни до, ни после копуляций. При садке птицы не трепетали крыльями и не кричали, хотя Yahya (1989) отмечал трепетание. Через несколько с после садки самец слетел на соседнюю ветку, самка осталась на месте, через короткое время самец вернул ся и спарился ещё раз (Roopa, 2018).
Вторую попытку у этого вида отмечали всегда, как и Trounov & Vasilieva (2014). После второй самки самка слетает первой, сбрасывая самца. Один раз отметили копуляцию, когда птенцы уже были в гнезде (№3); такое отметил и Yahya (1989).
Ночью насиживала и в гнезде ночевала только самка. За гнездами №№3, 5, 6, 7, 8 можно было наблюдать прямо с террасы дома, там удалось зафиксировать начало насиживания самцом, ещё затемно: в январе-декабре в 6.30, в апреле-мае в 5.45. Вылезли из ночёвочного дупла, он громко кричал kutroo – kutroo, затем умолкал и летел к гнезду. Подобравшись молча к летку, он издавал тихое kirr kirr: самка вылезала и отправлялась кормиться, чтобы сменить самца через 20-30 мин. При более поздних сроках насиживания самец «объявлял» о подлёте: сев на соседнее дерево, издавал kutroo kutroo. В ответ самка покидала гнездо с криком kuee kuee и летела на соседнее дерево, где кричала kuroo kuroo без начального слога kurr. По вечерам партнёры обменивались криками kuroo kuroo перед заходом на ночлег (Roopa, 2018). После вылупления оба партнёра стали совсем молчаливы, голос слышался только случайно. Каждый следующий день птицы заходили на ночлег позже, чем в предыдущий: соответственно при выкармливании это сильно после 19 ч., родители продолжали кормить и в темноте.
Гнездование красногузого бородастика P.lagrandieri изучено в апреле 2012 и апреле 2013 гг. в его естественных местообитаниях в лесу Loc Bac провинции Lam Dong, южный Вьетнам (Trounov, Vasilieva, 2014). Найдено 2 дупла с птенцам и 1 в процессе постройки. Все найденные дупла были выдолблены в сухих сучьях широколиственных деревьев, не определённых до вида, в ландшафтных выделах с относительно открытой экспозицией (склон холма, поляну в лесу или речной поток). Леток везде «смотрит» на открытое пространство. Дупло в разрезе бутылкообразное, с изогнутой входной трубкой (рис.12, сверху продольный, снизу поперечный разрез). На дне его совсем нет выстилки (в отличие от дятлов, засыпающих дно щепочками), еë стенки сухие, гладкие и чистые (Trounov, Vasilieva, 2014).
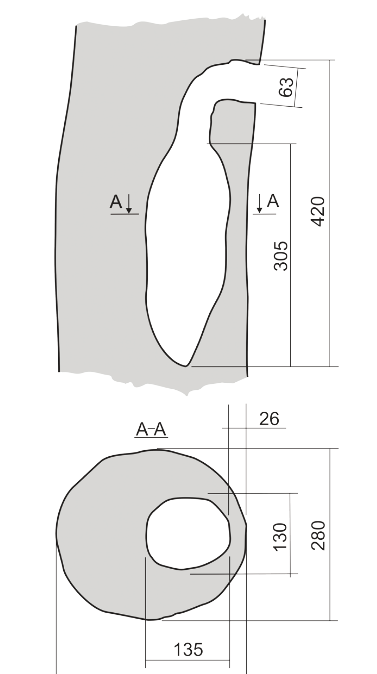
Рисунок 12. Дупло красногузого бородастика. Все расстояния даны в мм, «А» — уровень поперечного сечения
Общий объем гнезда был примерно 2783 см3, собственно гнездовой камеры — 2395 см3. Твердость вмещающей её древесины в среднем 0,4 кгc/мм2, её плотность 31,2 кг/м3.
Ранее форма гнездового дупла описывалась только для трёх видов: белоухого, цейлонского P.rubricapillus (Yahya, 1988, 2001) и синеухого бородастиков (Трунов, Васильева, 2012). Дупло красногузого бородастика очень похоже бутылковидной внутренней камерой и ведущим туда узким изогнутым туннелем. Решающие факторы в размещении гнезда — относительно открытая экспозиция, в выборе гнездового дерева — мягкая древесина, и диаметр ствола, коррелирующий с величиной птицы (Yahya, 2001; Трунов, Васильева, 2012). Для гнездовых деревьев мелкого синеухого бородастика (Трунов, Васильева, 2012) на юге Вьетнама он составляет 162 мм (n=9) и 375 мм (n=6)/360 мм (n=4) для более крупных зеленоухого P.faiostrictus и полосатого бородастиков (Трунов, личн. сообщ.).
Гнездовые деревья относительно крупного P.lagrandieri имели средний диаметр 387 мм (n = 3) и сильно более мягкую древесину, чем у всех местных живых деревьев, в три-четыре раза ниже твердости бальсы (максимум 1,6 кгс/мм2), самой легкой и мягкой из известных пород древесины. Будучи менее специализированными к долблению, чем дятлы, бородастики, видимо, могут долбить дупло только в мягких субстратах.
Два дупла авторы нашли готовыми, третье строилось примерно в 2,5 км от второго. Минимум 3 дня подряд трижды в день птица прилетала его долбить: между 10 и 10 ч. 25 мин (в среднем 9 мин долбления), между 13 ч и 13. 35 (9 мин), а также между 14.45 и 15.30 (14 мин). Птица не полностью скрывалась в летке, только по плечи, так что долбление было в самом начале (рис.13). Выдалбливание шло медленно, к концу наблюдений 16 апреля дупло было ещё не закончено (Trounov, Vasilieva, 2014).
Рис.13. Общие характеристики найденных дупел P.lagrandieri. Столбцы слева направо № гнезда, год наблюдений, состояние гнезда, диаметр ствола, в котором выдолблено, мм; высота над землёй, м; диаметр летка, мм; размер выводка.
Оба партнёра мономорфны, поэтому пол определим лишь во время совокупления. У пары, построившей гнездо №1, его отметили трижды: в 06:53–07:07 25 апреля 2012 г. и в 14:42– 14:52 и 17:20 28 апреля. Всегда перед этим птицы ~10–20 минут провели рядом друг с другом около гнезда: они сидели сплокойно, прихорашиваясь или двигаясь короткими прыжками вверх, иногда с короткими резкими криками и вздёргиванием хвоста. Однажды отметили, как самец перебирал перья на спине самки. Она, видимо, инциировала копуляцию: подлетела к самцу с тихим стрекотанием кричит, горизонтально поводила хвостом, распушив перья и слегка опустив крылья, после чего самец сделал садку. Каждая садка включала в себя 2 копуляции примерно за 5–6 с. Кормления самки самцом перед спариванием не отметили, однако наблюдали, как птицы из этой же пары соприкасаются клювами и передают друг другу фруктовую косточку (но не пищу) (Trounov, Vasilieva, 2014).
Красногузый бородастик у гнезда (фото из статьи)
Коммуникация между партнёрами была богатой. Самец и самка в парах, заботившихся о гнезде №1 и №2, проводили по 30-40 мин друг c другом, особенно во 2й половине дня, прежде чем идти на ночлег. Обычно они сидели бок о бок или двигались вместе на небольшой дистанции в кроне, после чего один бородастик шёл в ночёвочное дупло, другой в гнездовое. Что случалось между 17 ч 40 мин и 18 ч 25 мин в зависимости от погоды и условий освещения (Trounov, Vasilieva, 2014).
После входа в гнездо родитель сидел ещё 5-10 мин, высунувшись из летка, и осматривая окрестности. Утром он покидал гнездо в 5:20-5:25. При кормлении самца и самку различить невозможно, но оба они в этом участвуют. Иногда они встречались у летка, чаще одна птица ждала с кормом, пока партнёр покинет гнездо. Принесший пищу родитель сидел 5-7 мин в кроне гнездового дерева, смотрел вокруг, иногда манипулировал с кормом, растирая его. Лишьт изредка птицы прямо летели к гнезду.
1 Африканские виды рода Trachyphonus часто долбят дупла в термитниках, а то и выкапывают — в русле ручья или прямо в земле на плоской равнине. Периодически гнездятся в подобных местах и прочие бородастики.
2 Всё это верно для дятлов, пусть в меньшей степени. В выдалбливании гнездовых, да и ночёвочных дупел (к ним требования не так строги) они отнюдь не независимы и от состояния древесины: её твёрдости, особенностей строения, вроде внутренних сучьев и свилеватостей. Это ведёт к предпочтению повторного гнездования в старых дуплах везде, где для этого создана возможность устойчивым обитанием птиц на многолетне-постоянных участках, т. е. при высокой плотности населения, обычно достигаемой в малонарушенных старовозрастных лесах с излюбленным данным видом породным составом.
3Долина Атикадаву (11°11’26»-11°16’26» с.ш, 76°43’49»-76°50’25» в.д.), леса резервата Melur Slopes в Западных Гатах, 500-600 м.н.у м. Эта долина, прилегающая к юго-восточным склонам Нилгири, образует экологическую важный регион благодаря сохранившимся полулистопадным лесам вдоль р. Бхавани.
4 Она же батай, быстрорастущее дерево сем.Бобовых
5 Птицы также «простукивают» сучья и пни при кормлении, обнаруживая движение потенциальных жертв в древесине
Рекомендуем прочесть
 Зачем большим пёстрым дятлам всю зиму охранять территории?
Зачем большим пёстрым дятлам всю зиму охранять территории? О гнездовых дуплах и эволюции гнездостроения у дятлов Picidae:
О гнездовых дуплах и эволюции гнездостроения у дятлов Picidae: Болит ли голова у дятла?
Болит ли голова у дятла? Важность повторного использования дятлами дупел
Важность повторного использования дятлами дупел Сколько дятлов лесу нужно?
Сколько дятлов лесу нужно? Проблемы и риски выдалбливания дупел у дятлов
Проблемы и риски выдалбливания дупел у дятлов Исследование маскировочности окраски дятлов
Исследование маскировочности окраски дятлов В эпоху глобального экологического кризиса города — не только «проблема», но и “решение”
В эпоху глобального экологического кризиса города — не только «проблема», но и “решение”
