
Джон Мэйнард Смит, начавший применять теорию игр к агонистическим взаимодействиям и, шире, к коммуникации. Наряду с Уильямом Дональдом Гамильтоном и Робертом Лудлоу Трайверсом стал «дедушкой» социобиологии: обосновал концепты, легшие в её основу
Плюсы социобиологии
Содержание
Я много и, надеюсь, по делу, ругал социобиологию1, но у неё есть несомненное достижение. Именно социобиологи показали, что
а) демонстрирование неизменно сопровождается пропорциональным ему риском;
б) эффективность действия единиц поведения и риски их предъявления во взаимодействиях вида Х в контексте У положительно коррелируют между собой (найдено во всех Х и У, специально исследованных в этом плане, Фридман, 2013, 2017аб).
Они же впервые выяснили, что с точки зрения риска коммуникация и её «материальная часть» — демонстрирование, сравнимы с риском радиста разведгруппы, разыскиваемой функабвером:
«Разведывательная сеть — это своего рода подводная лодка, которой всегда приходится плавать с перископом над водой. Этим перископом является антенна передатчика.
Жестокая охота, поскольку травят человека. Каждый раз, когда радист садится за передатчик, он в большом напряжении — ведь он чувствует погоню, чувствует, как сжимаются тиски. Радист также беззащитен, как и солдат, поднявшийся из окопа под обстрелом вражеских пулемётов. Он знает, что свора напала на его след. Он представляет себе машины, движущиеся на малой скорости, мысленно видит притворно безучастных «прохожих», приближающихся к дому, слышит выстрелы по замочной скважине двери.
Мужество его состоит в том, чтобы не встать с места и продолжать передачу. Это, несомненно, труднее штыковой атаки. Возвышенная охота, потому что героизм без мишуры и фанфар достигает здесь абсолютной чистоты2».

Точный изоморфизм готовности радиста вести передачу вопреки растущей опасности фиксируется в коммуникации животных, даже в трёх формах: все они связаны с опосредованием действий демонстрациями.
Во-первых, это «парадоксальная сдержанность» особей, дальше выигрывающих взаимодействие: в ответ на прямые действия оппонента, включая чреватые травмой и даже гибелью (клевки, вытеснения, остановка силой партнёра для спаривания) они лишь сильней и стереотипнее демонстрируют. Будущие проигравшие, наоборот, показывают больше открытой агрессии, чем демонстраций, и тем больше, чем сильнее определился исход конфликта.
Во-вторых, данная «сдержанность» делает демонстратора особенно уязвимым к нападению хищника, вроде бахромчатогубых листоносов Trachops cirrhosus, ловящих тунгарских лягушек Engystomops pustulosus (сем. cвистунов Leptodactylidae), ориентируясь по брачным крикам самцов. И, шире, всякое демонстрирование ухудшает возможность бегства в укрытие, тем сильней увеличивая угрозу гибели, чем дальше зашло взаимодействие и/или ритуализованней формы используемых демонстраций3. Последующий анализ показывает, что позитивная связь уязвимости к хищничеству с ритуализованностью форм поведения имеет ту же природу, что связь с нею же риска ранений и травмы от оппонента при демонстрировании.
В третьих, у более эффективных демонстраций выше стереотипность исполнения комплекса телодвижений и действий, образующих демонстрацию4 в связи с передачей более ценной сигнальной информацией обоим участникам. Всегда есть возможность, что оппонент выиграет взаимодействие, использовав её лучше самого демонстратора.
Все три фактора риска неизменно присутствуют в коммуникации, и дальше рассматриваются последовательно. Самое важное во всех трёх — как чувства радиста (волнение, страх, ненависть ко врагу) не должны снизить точности его профессиональных действий («автоматизм» и стереотипность исполнения отличают их от обычного поведения), так у животных-участников коммуникации созданный ею рост рискованности демонстрирования не снизит ни готовности к выбору более эффективных демонстраций на «следующем шаге» процесса, ни стереотипности их исполнения. Что вполне изоморфно молчаливому героизму радиста и отлично от поведения особей, ещё не начавших коммуникацию или не втянутых в неё как следует — здесь увеличение риска подавляет эту готовность, ограничивая выбор демонстраций или вовсе прерывая процесс (см. также факт 24).
Риск открытой агрессии как «цена» демонстрирования и «плата» за информацию
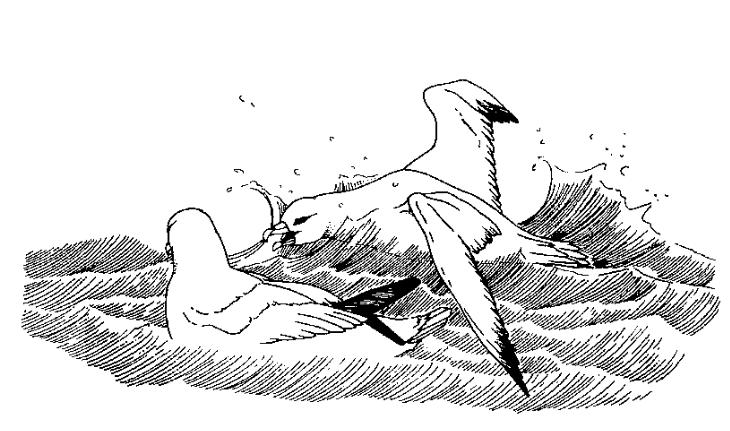
Так, репертуар агонистических реакций серебрянных белоглазок Zosterops lateralis включает демонстрации (вокализация, широкое раскрывание клюва с выпадами в сторону противника, трепыхание крыльев и пр., всего 9 поз) и прямые действия (клевки, погони, подскоки, замещающие прыжки), из которых состоят открытые драки (рис.1). Большинство птиц, инициирующих конфликты и все победители (это не только инициаторы), начинают контакт с демонстрации, а не с действия. Cравнительно с проигравшими они значимо больше демонстрируют, чем действуют по ходу конфликта.
 А.
А.
 Б.
Б.
 В.
В.
Г.
Д.
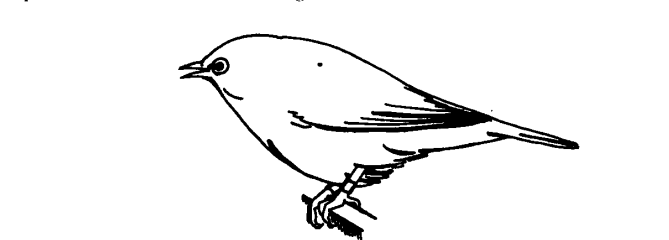
Рисунок 1. Агонистические демонстрации серебрянных белоглазок с организацией обмена ими в конфликтах. А. Белоглазка в Виктории, Австралия. Б. Ритуализованная угроза «наклон вперёд». а. Внешний облик: тело вытянуто, крылья опущены, ноги согнуты, клюв открыт, b. Как это видится атакуемой птице: обратите внимание на «подчёркнутость» позой белых нижних кроющих перьев крыльев и поднятых вверх пятен на боках.
В. «Распушённая» поза, выражающая отступление и подчинение, из признаков агрессивных побуждений остался только раскрытый клюв.
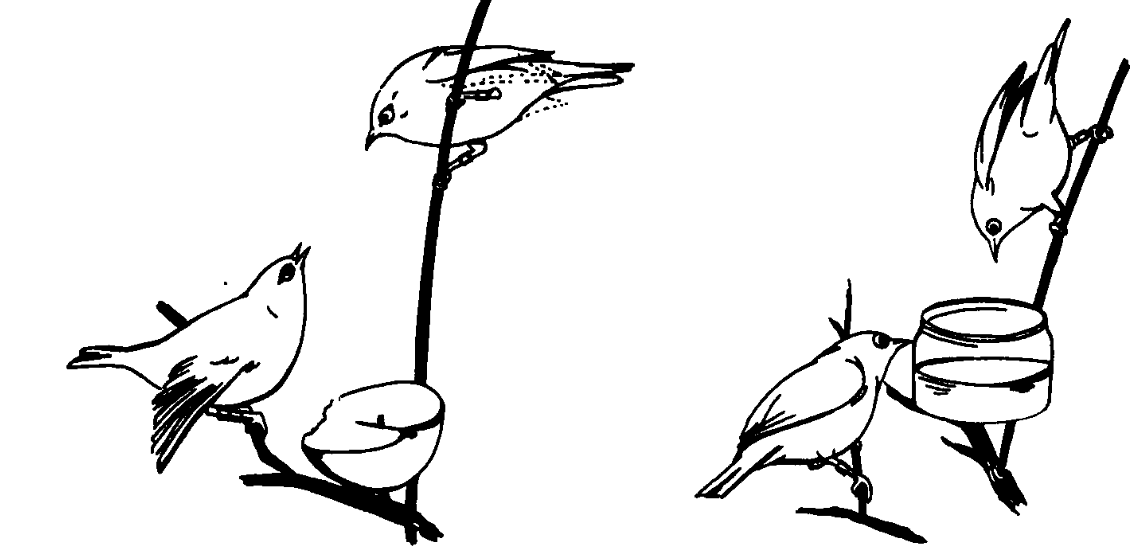
Г. Кадры агонистических взаимодействий. Слева: ответная поза угрозы нижней особи прекращает у верхней демонстрирование и «включает» прямую атаку, повторение этого ведёт к проигрышу. Справа: подчинённая (т. е. много проигрывавшая в предыдущих конфликтах, см. Kikkawa, 1961, fig.8) самка внизу пьёт капли сладкой воды, разбрызганные по стенкам поилки, доминирующий самец уверенно движется к самой удобной позиции для питья (вверху).
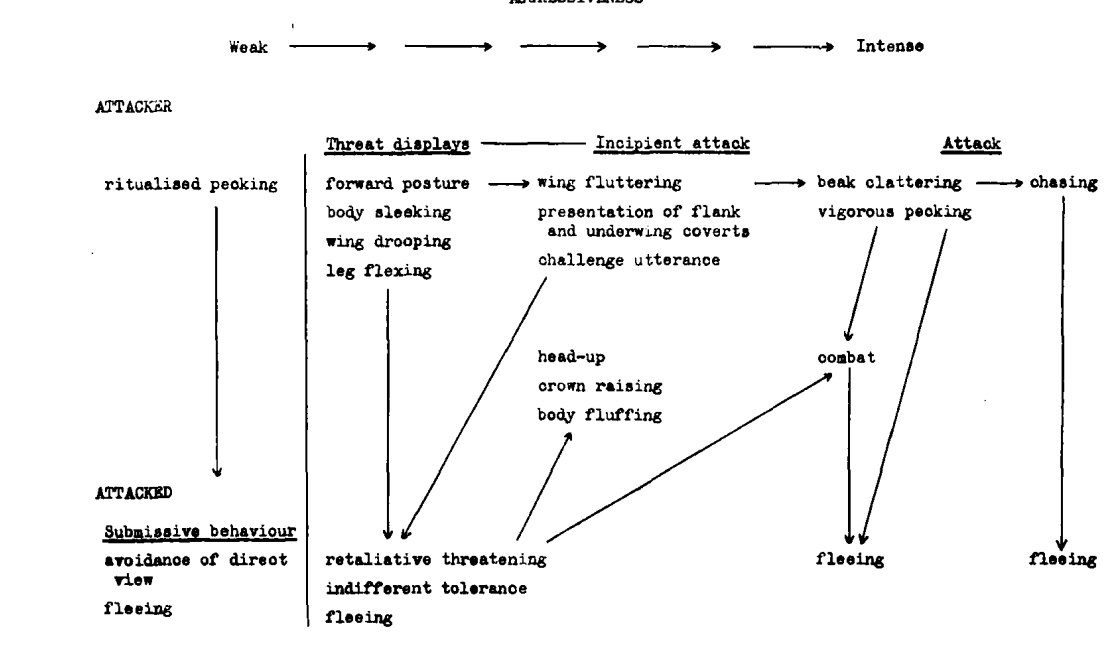
Д. Ординация агонистических демонстраций Z.lateralis по осям конфликта побуждений нападение — бегство (У, агрессия растёт вверх, страх с отступлением вниз, сверху атакующий, ему свойственны ритуализованные клевки, снизу атакуемый, ему — избегание прямого взгляда и слёт) и уровень возбуждения (Х, растёт слева направо, разделяясь на 3 категории, ритуализованные угрозы — движения намерения атаки/бегства — открытая атака/бегство). Стрелки — направления «перебора» демонстраций во взаимодействиях, их описания слева направо и сверху вниз; 1. Вытянутая поза, прижатое оперение, опущенные крылья, ноги согнуты. 2. «Брошенный вызов»: взмахивания крыльями c демонстрацией нижних кроющих крыла и боков. 3. Цоканье клювом, энергичные клевки. 4. Преследование. 5. Поза «голова вверх», перья темени подняты, оперение тела распушено. 6. Драка. 7. Ответная (ритуализованная) угроза, затем безразличная терпимость и далее слёт. 8 и 9. Слёт.
Источник: Kikkawa, 1961.
Напротив, проигравшие белоглазки выказывают больше открытой агрессии, чаще нападают и устраивают погони, но в обменах демонстрациями неизменно проигрывают. Побеждающие — отвечают на открытую агрессию демонстрированием: переходят к более эффективным (сильнее ритуализованным) демонстрациям репертуара5 или подчёркнуто стереотипно удерживают текущую. Во-вторых, по мере развития конфликта и увеличения уровня агрессивности участников (что видно по учащению ударов и погонь в паузах между демонстрациями) поведение будущих победителей ритуализуется всё сильней: обмен демонстрациями теснит открытую агрессию по времени, её акты приурочиваются только к моментам выбора и смены у первых. Будущие побеждённые, наоборот, интенсифицируют проявления открытой агрессии параллельно росту эффективности сигналов и, в конце концов, «проигрывают гонку», принимая позу подчинения или спасаясь бегством (Kikkawa, Wilson, 2002).
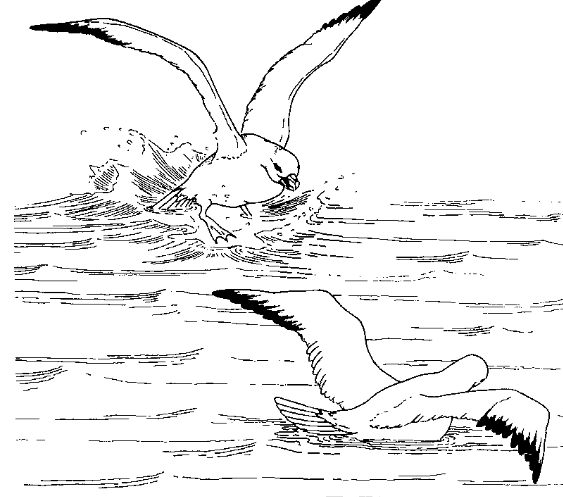
Действительно, анализ агонистических взаимодействий глупышей Fulmarius glacialis (Enquist et al., 1985), конкурирующих за пищу, показал, что больший или меньший уровень риска — следствие выбора и стереотипного исполнения одной из возможных демонстраций репертуара, а вовсе не «силового» давления особи на противника или наоборот6.
Авторы анализировали роль агонистических демонстраций вида (рис.2) как сигналов-посредников в столкновениях птиц, скапливающихся вокруг выброшенных внутренностей пинагора Cyclopterus lumpus. Каждому куску находится свой «бесспорный владелец»: поедая, он одновременно угрожает остальным, крутящимся вокруг и пытающимся вырвать часть или завладеть всем. В 22% наблюдений владелец сменялся другим, не отстояв пищу, что почти не зависело от размера / пищевой ценности куска.
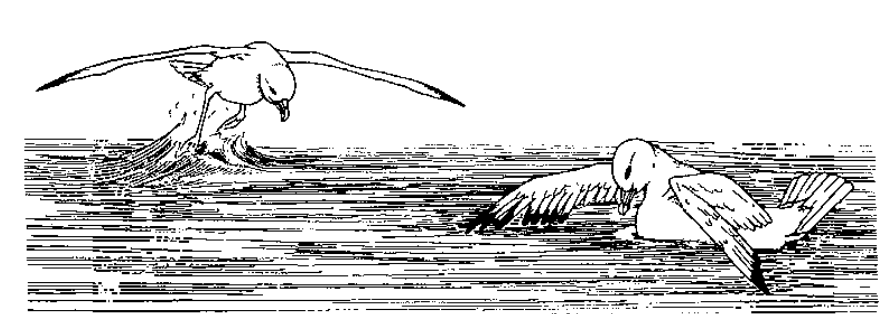
Угрозы глупышей ритуализованы, прямые действия агрессии опосредованы 8 демонстрациями, часть их показана на рис.2:
1. Крылья подняты и распростёрты, иногда с опущенными первостепенными маховыми (рис.2А). Обязательная часть всех других демонстраций, за исключением трёх случаев демонстрации №4. Помимо угрозы конкретной особи, №1 постоянно используется для угрозы «в пространство»: глупыши поднимают крылья, завладев куском и поедая его, чем пресекают часть попыток напасть. Здесь №1 не сочетается с прочими телодвижениями, исключая движения хвоста.
2. Крылья частично подняты, их верхушка опущена, создавая впечатление амбивалентности (конфликта побуждений нападения и бегства). Визуально легко отличается от №1 (рис.2В).
3. Раскрывание хвоста. Обязательный компонент №1 и №2 при угрозе конкретной особи (рис.2А), не отмечено как отдельная демонстрация.
4. Поза изгиба: шея изогнута, голова и клюв направлены вниз, к воде, при поднятой передней части корпуса. Почти всегда в сопряжении с №1, кроме трех случаев её предъявления захватчиком.
5. «Указание клювом» на противника (разной интенсвности, от поворота головы в его сторону до попытки клевка, явно без цели попасть, т. е. демонстративно). Всегда сочетается с №1, птица часто ещё разевает клюв и издаёт резкий звук. Также используется в угрозе летящим птицам.
 А.
А.
 Б.
Б.
 В.
В.
 Г.
Г.
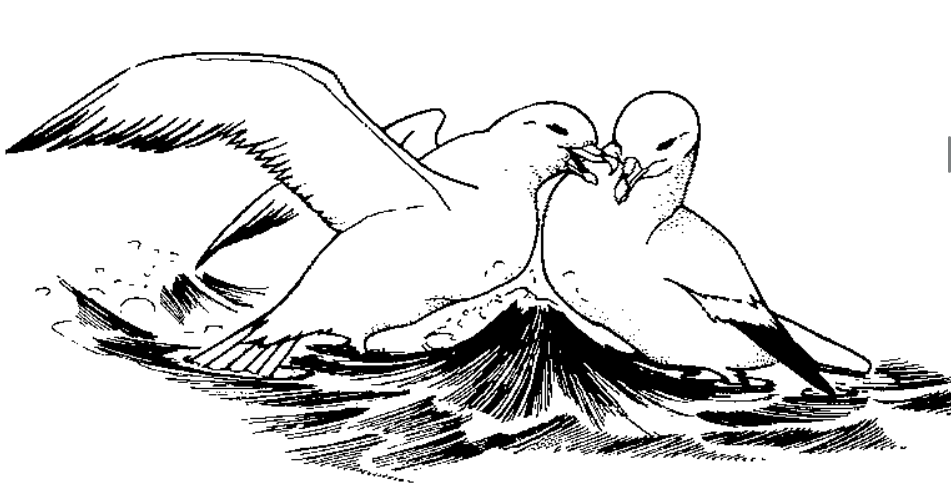
Рисунок 2. Единицы ритуализованной агрессии глупышей. А. Взаимная угроза распростёртыми крыльями (у обороняющейся птицы с опущенными первостепенными маховыми), захватчик налетел слева. Б. «Указание клювом» (верхняя птица). В. Натиск на противника (в позах угрозы), здесь осуществляется левой птицей, права отвечает №1. Г. Противостояние «грудь-в-грудь». Источник. Enquist et al., 1985.
6. Щипки. Успешная попытка щипнуть оппонента клювом. Не отмечены непосредственно при физической борьбе, почему их можно считать ритуализованной формой агрессии.
7. Ритуализованный натиск. Бег по воде на противника с короткой дистанции (<3 м), с шумом и брызгами, в позах №1 или №4, по мере сближения клюв направляется на оппонента, показывая рост уровня агрессивности. У одних птиц натиск реализован независимо от позиции относительно оппонента, другие неизменно заходят сзади и осуществляют натиск оттуда.
8. Глупыш выпускает рыбу, что не значит «сдаётся»: часто возобновляет атаки на нового владельца. «Переход» куска пищи от одного участника конфликта к другому — обычное дело.
9. Противостояние «грудь-в-грудь». Находясь напротив друг друга и приподняв переднюю часть корпуса, глупыши сталкиваются грудью, часто с полураскрытыми крыльями. Неизменно предшествует №7 в исполнении одной птицы или обоих.
10. В части конфликтов обмен демонстрациями №№1-9 завершался открытой дракой (или «срывался» в неё, сравни c поведением при подстановке чучела). Птицы бьют и/или хватают противника клювом (иногда прочно сцепляются клювами), бьют крыльями, тело держат почти вертикально, с поднятыми крыльями, часто соприкасаясь между собой. В смысле причинения боли клюв всего эффективней, крылья второстепенны.
Шансы такого развития после использования конкретных демонстраций авторы полагали их риском предъявления: тогда время удержания рыбы значимо короче, а её владельцы менее успешны в конфликте (при прочих равных), чем когда всё ограничивалось обменом демонстрациями. Характер течения (продолжительность конфликта, число сменившихся элементов №№1-11) и специфический результат взаимодействия целиком задан выбором участниками демонстраций с присущей им эффективностью и риском (Enquist et al., 1985).
Уровень агрессивности определяли по времени удержания пищи: голодный глупыш обороняет рыбу дольше, чем сытый. Владельцы рыбы выигрывали 78% конфликтов, особенно если они их инициировали (98%), захватчики 22% (если инициировали — 27%). Успешность захватчиков возрастает с ростом числа смен поведенческих актов во взаимодействии, владельцев — падает, если конфликт завершён дракой (63% vs 80% в её отсутствие). Все различия высоко значимы, Р<0,001 (Enquist et al., 1985, table 2).
11. Бросание вызова обладателю рыбы с воздуха. Полёт и приземление около владельца рыбы, инициирующее конфликт (другой вариант — прямо на него). Не включая каких-либо демонстраций, несут информацию о претензиях: это не просто приближение к оппоненту, сидящие на воде птицы взлетают, делают широкий круг и опускаются.
Увы, в описании авторы не различают собственно демонстрации и их составляющие — отдельные телодвижения, более или менее «вычурные» и «неестественные», чьё сопряжённое исполнение даёт собственно демонстрацию. Однако все они ритуализованы, используются для угрозы, «подкрепляемой» «выплеском» прямой агрессии (рис.2). Особая ценность работы — в определениях эффективности и платы/риска демонстрирования, исходящих напрямую из наблюдаемого поведения, его измеряемых характеристик, вместо, как это бывает обычно, вытекающих из теории авторов7.
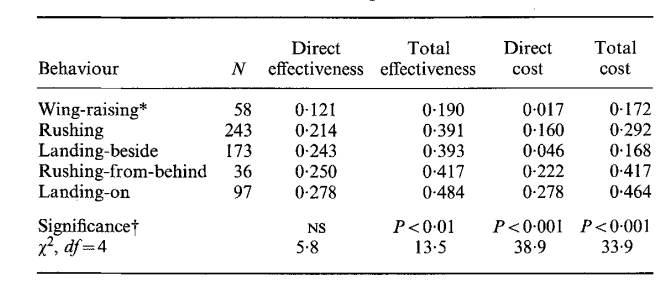
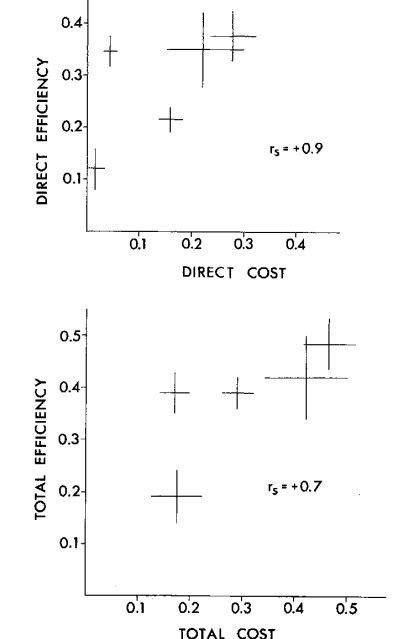
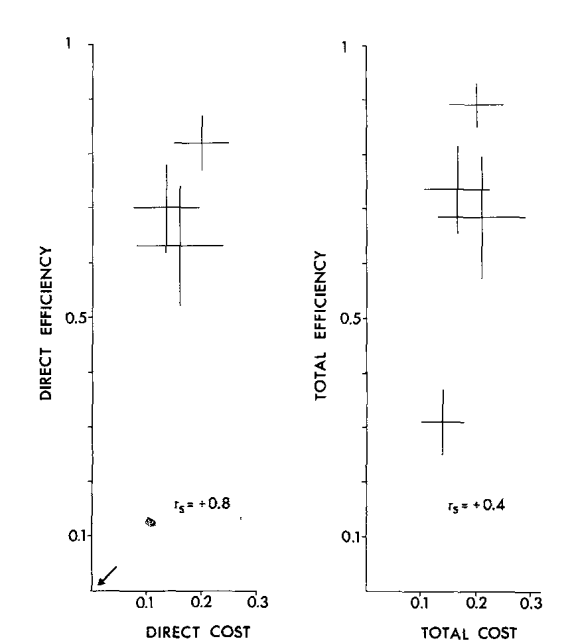
Общая эффективность (total effectiveness) угрожающих демонстраций определялась шансом победы демонстратора, прямая эффективность (direct eff.) — шансом победы сразу после принятия данной позы (что оппонент не «продолжит» взаимодействие следующей демонстрацией), непрямая (indirect eff.) — вероятностью победы в конечном итоге через некоторое число смен поведенческих актов; влияние выбора данной демонстрации на данный исход устанавливали при помощи техник непараметрического дисперсионного анализа. Плату определяли шансом столкнуться с опасными действиями оппонента, разделёнными на более и менее травмогенные (№6, №№9-10 vs все остальные).
«Плата», она же риск (cost) — вероятность перерастания в физическую борьбу. Её разделили на прямую (direct cost) и общую (total cost) платы демонстрирования, т. е. шансы столкнуться с травмогенным действием сразу после предъявления данной позы и шанс на перерастание обмена демонстрациями в физическую борьбу «в конечном счёте», после нескольких смен (рис.3).
Для всех выделенных демонстраций показана линейная зависимость эффективности и риска (Enquist et al., 1985). Важно подчеркнуть: уровень агрессивной мотивации глупышей определяется именно готовностью пойти на бόльший (меньший) риск демонстрирования, а не бόльшим (меньшим) проявлением открытой агрессии. Это означает ритуализованность выражения агрессии и «доверие» всех собравшихся птиц к эффективности демонстраций как специфических средств противостоять отбиранию пищи, причем непрямым образом — за счёт их ритуализованной формы и сигнального действия.
Действительно, взаимодействия, выигранные захватчиком, составлены большим числом смен демонстраций, чем выигранные владельцем рыбы (~6,1 vs ~3,5); рост их числа понижает шансы победы владельца (коэффициент ранговой корреляции Спирмена rs = 0,64, N = 1383, P < 0,001). Частота взаимодействий высока: от 0,8 до 22 конфликтов/мин, в среднем 2,2, включающих 4,1 смены (пределы 1-22 смены). В среднем владельцу 3,97 раза бросают вызов захватчики, прежде чем рыба перейдёт к другому (пределы 1-31 раза); среднее время её удержания 88 с (5-990).
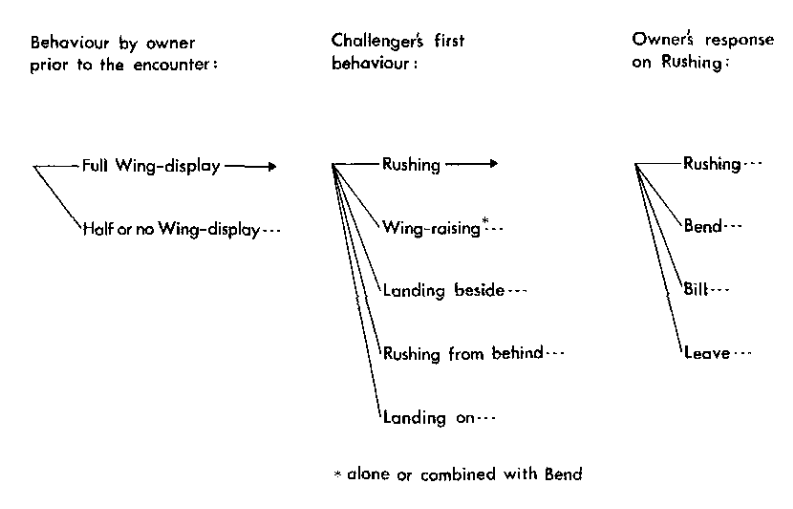
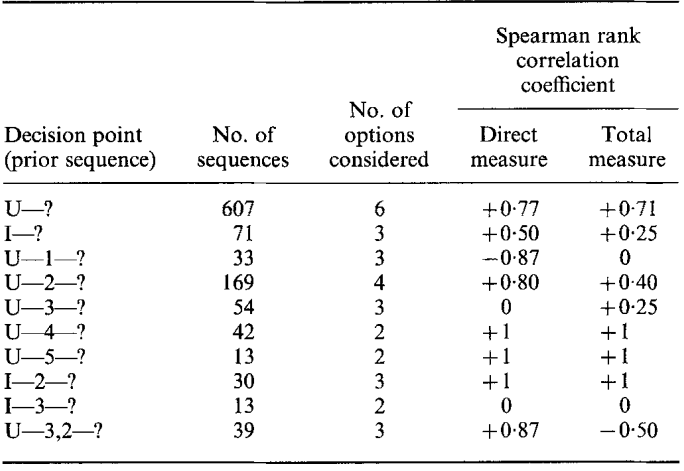
Анализ Enquist et al., 1985 подтвердил как зависимость специфических результатов взаимодействия (удержит глупыш рыбу при данном уровне притязаний? как долго? какой ценой?) от выборов захватчиком и владельцем конкретных демонстраций репертуара с присущими им эффективностью и риском на разных «шагах» развития конфликта, особенно самом первом. См. дерево решений на рис.3; для репрезентативности выводов в анализ включили лишь точки принятия решений с частотой выбора самого редкого поведения >30 раз, почему их только 4. Напротив, не выявлено зависимости от интенсивности «силового» воздействия птиц друг на друга затянутости взаимодействий и т. д. обстоятельств контекста. Так исследования, выполненные в прокрустовом ложе социобиологической доктрины, снова и снова подтверждают концепты её конкурентки, сравнительной этологии.
 А.
А.
 Б.
Б.
 В.
В.
 Г.
Г.
 Д.
Д.
 Е.
Е.
Рисунок 3. А. «Дерево решений» в агрессивных взаимодействиях глупышей. Слева возможное поведение владельца до конфликта (№1, №2 и т.д), в центре — первая демонстрация захватчика (№7, №1, в одиночку или в комбинации с №4, №11 с приземлением рядом с владельцем, №7 с приземлением не на него, а рядом, №11 на него и т. д.), справа возможные выборы владельцем ответной демонстрации на №7: №7, №4, №5, №8 и т. д.
Б. Эффективность и плата (риск употребления) первых демонстраций захватчика (строчки сверху вниз — №1, в одиночку или в комбинации с №4; №7; №11, с приземлением рядом с владельцем; №7, с тыла; №11, на владельца). Столбцы слева направо: прямая и общая эффективности, прямая и общая платы демонстрирования. Везде * поза одиночная или в комбинации с позой изгиба; † — значимость влияния фактора на отклик, опровержение нуль-гипотезы независимости выбора поведения в строках и наблюдаемых вероятностей эффективности и риска в таблице 5*2, тест Краскелл-Уоллеса. В. Среднее время владения рыбой (t, c ± стандартная ошибка, SE) как функция первого выбора демонстрации (строки, см. Б) при захвате рыбы. Влияния фактора на отклик высоко значимы (тест Краскелл-Уоллеса).
Г. Положительная связь прямых эффективности и платы (ранговая корреляция Спирмена rs) для первой демонстрации захватчика (сверху); то же для общих эффективности и платы (внизу); средние значения вероятностей ± SE. Д. Положительная связь прямых эффективности и платы для демонстраций-ответов владельца на ритуализованный натиск захватчика (слева); то же для общих эффективности и платы (справа), средние значения вероятностей ± SE.
Е. Положительная связь эффективности и риска демонстрирования (значения rs) неизменно присутствует в разных цепочках выборов поведения (строки). Включены все такие цепочки с >2 смен демонстраций и частотой >5 случаев каждая. Обозначения. U — демонстрация №2; I — её исполнение наполовину или менее; 1 — №1 в одиночку или скомбинирован с №4, 2 — демонстрация №7; 3 — №11, рядом с фокальной особью, 4 — №11, непосредственно на неё, 5 — №7, сзади фокальной особи. Буквы относятся к владельцу рыбы, цифры — к захватчику. Столбцы слева направо: n; число смен демонстраций; rs для прямых эффективности и платы демонстрирования; то же для общих.
Данные Enquist et al. (1985) напрочь разбили исходное представление социобиологов, что «честное» информирование о намерениях в социальных взаимодействиях не является эволюционно-стабильной стратегией8, почему попросту не могло быть создано эволюцией. Отталкиваясь от этой теории, социобиологи 15-20 лет подряд игнорировали, что передача информации о намерениях и/или потенциальных возможностях разрешения данного взаимодействия ритуализированными демонстрациями репертуара установлена более чем надёжно, подтверждена независимыми исследованиями не раз и не два, а множество (см. «20 фактов об инстинктах», факты 15 и 18), как и её «кодирование» специфическими параметрами форм демонстраций.
Обнаружив то и другое в работах, выполненных в рамках своей парадигмы, социобиологи «изобрели велосипед»: выявили качественную неоднородность поведенческих актов, составляющих агонистические, потом и все прочие взаимодействия, включая ухаживания. Наряду со «средствами силовой борьбы», передающими воздействие, возбуждающими и/или подавляющими активность оппонента сугубо неспецифически, репертуар поведенческих актов включает в себя видоспецифические демонстрации, передающие информацию
а) о возможностях разрешения данного конфликта «в пользу» того из участников9, кто ей лучше «воспользуется», скорректировав прежнюю модель поведения точнее и/или раньше, и
б) о «цене» демонстрирования — уровне риска, на который идёт животное ради такой передачи.
После всего перечисленного социобиологическое «обоснование» неустойчивости «честной» коммуникации (как ЭСС), маловероятности и/или невозможности типологически определённой формы, сигнальной функции демонстраций и пр. концептов сравнительной этологии должны бы пополнить перечни красивых теорий, «убитых» некрасивыми фактами. Но увы! Всё это держится ненаучными средствами, вместе с прочими социобиологическими концептами, уж слишком они хорошо подыгрывают предрассудкам публики; хочется верить, что книга всё же заставит задуматься и понизит их популярность.
Превосходство будущих проигравших в открытой агрессии контринтуитивно настолько, что кажется невозможным, но неизменно присутствует везде в агонистических взаимодействиях (в территоральных конфликтах, в агонистическом доминировании и пр.). Как и его сопряжённость с «повышением планки» агрессии ритуализованной, т. е. выраженной демонстрациями видового репертуара у обоих противников: она возникает именно вследствие положительной связи эффективности и риска демонстрирования. Агрессия, образующая последний, создана демонстрированием побеждающей особи и реализуется инстинктивным реагированием проигрывающей на её демонстрации при таком повышении, вовсе не персональным уровнем её агрессивности.
Больше того, правило «выигрывающие участники взаимодействия больше демонстрируют, чем действуют, «сдерживая» прямую активность, у проигрывающих наоборот, она превалирует над демонстрациями, и чем дальше, тем больше», верно и для прочих контекстов — ухаживания, «тестирования» потенциальных партнёров при образовании пары, информирования о корме или предупреждения об опасности (Фридман, 2013, табл.10). Отсюда «автоматически» следует возрастание всех вышеописанных рисков: более эффективные демонстрации более ритуализованы и стереотипнее исполняются, то и другое — непосредственные источники риска (действующая причина по Аристотелю).
Риск демонстрирования рождается из стереотипности
Изоморфизм положения участников коммуникации относительно всех других компаньонов, от хищников до «своих» брачных партнёров и противников в территориальных конфликтах, и работы радиста разведгруппы вполне точен, при всей литературности. В обществе подобная доблесть довольно редка, её выработка требует специального воспитания (включая умение подавить рост непродуктивных эмоций, вроде агресси, гнева и ненавистик к врагу, сделать что должен профессионально и точно вопреки всем «возмущениям»), почему почитается. В природе готовность животного продолжать коммуникацию дальше и дальше, пока не достигнет выигрыша (т.е. в сторону “повышения планки” ритуализированного ухаживания или угрозы = перехода ко всё более эффективным и рискованным сигналам) требует такой же готовности преодолеть каждый раз всё более высокий барьер страха, стресса и т. д. негативного состояния “внутри” действующего индивида, подавляющий любую активность, а в особенности телодвижения столь вычурные, неестественные, разнородные и при этом так точно сопрягающиеся между собой, как более эффективные демонстрации в сравнении с менее эффективными.
Скажем, у бурого анолиса A.sagrei описаны три угрожающие демонстрации10 – кивки головой (head-bob display), отжимания на лапах (push-up) и раздувание горлового мешка (dewlap). При моделировании ситуации нападения хищника показом модели пустельги значимо сокращается частота использования «отжиманий» и «раздуваний», но не более эффективных «кивков» (Simon, 2007). Бόльшая устойчивость более эффективных демонстраций к риску хищничества, внешнему беспокойству и другим дестабилизирующим факторам также зафиксирована в социальной коммуникации рыб, вроде подробно исследованной «борьбы ртов» у южноамериканской цихлиды Nannacara anomala, и других групп позвоночных (Фридман, 2013).
Перечисленное — продукт стереотипности исполнения демонстраций, пропорциональной их эффективности, поскольку её порождает. Чем эффективнее демонстрация, тем “вычурней”, неестественней разнородные телодвижения, “вычерчивающие” её характерный образ, их исполнение жёстче сопряжено во времени (т. е. оно делается одновременей), контрастируя с разнородностью происхождения данных движений. Т.е. сильней и устойчивей корреляции, образующие демонстрацию: в создании её инвариантного образа и, главное, его устойчивом предъявлении вопреки «возмущениям» они играют роль обручей в бочке. Коммуникативное значение имеет лишь цельная комбинация элементарных движений (по отдельности каждое из них незначимо), и только при скоррелированном исполнении разнородных движений и достаточной устойчивости корреляций к изменениям контекста (Фридман, 2013).
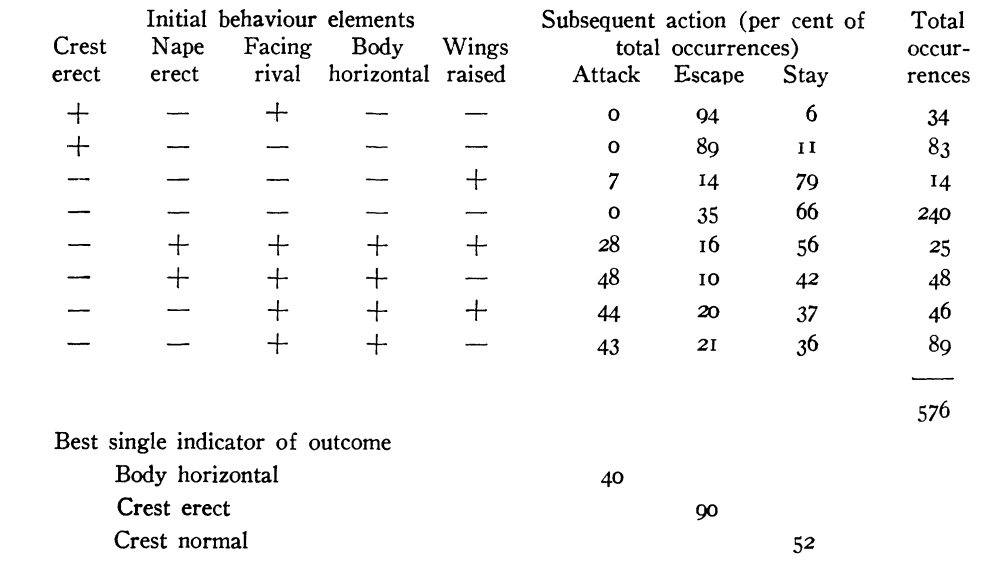
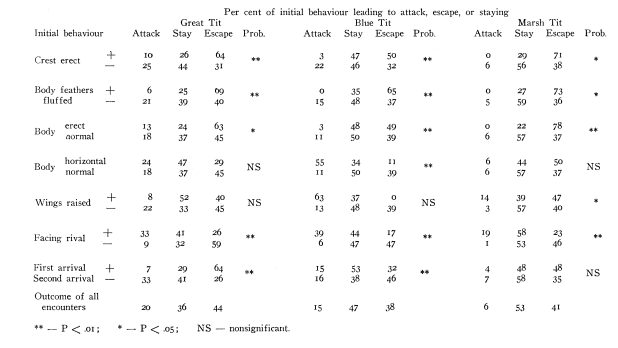
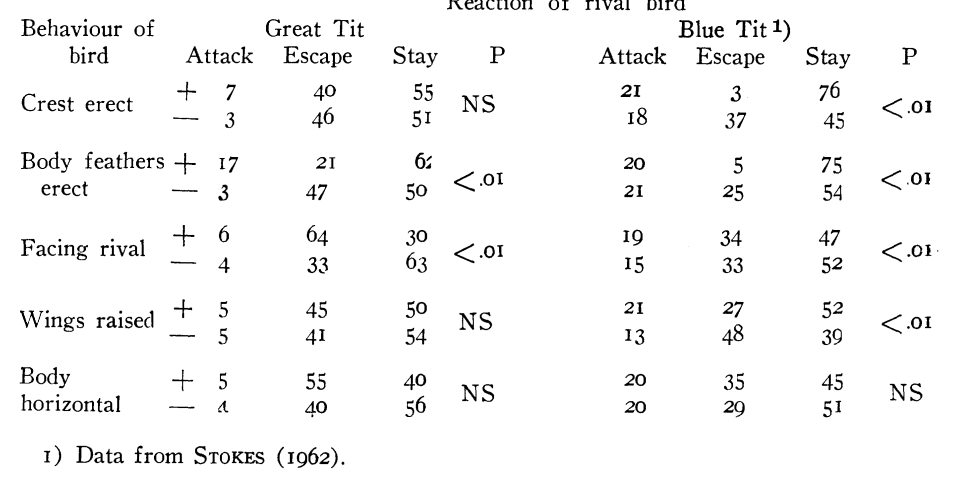
Впервые это показано A.W.Stokes (1962ab) в анализе образования агонистических демонстраций из выразительных движений у синиц: последние более или менее гомологичны у разных видов, но, будучи «собраны» в видоспецифичные комбинации, они образуют разные формы демонстраций (рис.4А-Г). Автор сперва изучил скоррелированность предъявления элементов в составе комбинаций, дальше сигнальные эффекты целого — демонстраций — сравнивали с таковыми их субъединиц. Так, в агонистических демонстрациях лазоревки Parus caeruleus выявили корреляции между 9 компонентами демонстраций, из которых важнее всего положение хохолка, тела, затылка, крыльев и ориентация относительно партнёра: значимы 31 парная корреляция из 36 возможных (Р<0,01). Часть из них положительна (хвост веером обычно связан с подъёмом крыльев), часть отрицательна: взъерошив оперение на голове, синица не поднимала ни крыльев, ни клюва, и не распускала хвоста. Более подробный анализ пяти компонент (положение хохолка, тела, затылка, крыльев, ориентация относительно другой синицы) показал значимость лишь 20 из 47 возможных комбинаций, 8 из них отмечены 576 раз (всего n=629). (Stokes, 1962a-b).

А.

Б.
 В.
В.
 Г.
Г.
Д.
Рисунок 4. Гомология выразительных движений, образующие позы угрозы у разных видов синиц (А), при видоспецифичности самих поз (Б, позы угрозы больших синиц слева и лазоревок справа). Приведены данные о скоррелированности исполнения выразительных движений при образовании данных поз (В).
Обозначения. В. Показаны комбинации 5 выразительных движений лазоревки (слева направо: поднятие хохолка, выпрямление линии затылка, позиция лицом к противнику, тело горизонтально, раскрываете крыльев) как предикторы исхода встречи двух лазоревок на кормушке. + телодвижение присутствует в комбинации, — отсутствует, следующие 3 столбца справа частоты последующих действий демонстратора: атака, бегство, оставаться на месте, % от n. Три строки под таблицей: телодвижения — наилучшие индикаторы для исходов: тело горизонтально — для атаки (0.4), хохолок поднят — бегства (0, 9), хохолок опущен — оставаться на месте (0, 52). Г. Аналогичная таблица для трёх видов синиц — большой слева, лазоревки (центр), болотной гаички Parus palustris (справа). Д. Сигнальное значение гомологичных поз у большой синицы (центральный столбец) и лазоревки (правый столбец). Строки слева — состав телодвижений в комбинации (строки сверху вниз, их перечень см. В):, + присутствует, — нет. В каждом столбце даны частоты альтернативных ответов второй особи: атаки, бегства, оставаться на месте (слева направо).
Источники: А. Петрова, 2003. Б-Д. Stokes, 1962ab.
Отдельные выразительные движения — плохие предикторы последующих атак или бегства, корреляции здесь крайне слабы, хороший предиктор — устойчивые комбинации элементов. Скажем, взъерошивание хохолка на затылке повышает шанс нападения в ситуации, обычно не связанной с агрессией, но понижает, если птица уже приняла агрессивную позу. Подобные составляющие демонстраций в агонистических взаимодействиях синиц появляются, как правило, в определённых комбинациях, и связаны с последующим поведением намного жёстче. Одна комбинация в 94% случаев ведёт к бегству, после другой птица в 79% случаев остаётся на месте, за третьей в 49% случаев следует нападение. Далее было показано, что устойчивые корреляции элементарных действий как раз и образуют те агонистические демонстрации, предъявление которых обеспечивает предсказуемость и специфичность ответа партнёра, позволяет обоим участникам конфликта чётко прогнозировать поведение друг друга и принимать решение о двигательном ответе на сигнал, будь то бегство, замирание, ответ другой демонстрацией и пр. (Stokes, 1962a-b).
Из этих работ выясняется, что
а) для партнёров животного и для самого демонстратора информативны именно комбинации элементарных движений,
б) лишь когда последние предъявляются скоррелировано во времени, они обладают специфическим образом, превращающим их в демонстрацию (Stokes, 1962a-b).
в) Лишь тогда данные комбинации информируют о намерениях демонстратора, соответствующая информация не содержится ни в какой из их составных частей, даже в происходящих из движений намерения животного (они, казалось бы, могли быть «значимы»). Каждое из выразительных движений, образующих демонстрацию, по отдельности лишь неспецифически возбуждает оппонента и самого демонстратора. Хотя все они в той же степени детерминированы специфическим конфликтом мотиваций «внутри» особи, что и самоё демонстрация, информирует о намерениях и действиях животного именно и только её целостный образ, созданный комбинированием выразительных движений по ходу взаимодействия и дальше преобразуемый в другую целостность при смене демонстраций.
Иными словами, специфические средства коммуникации животных, сигналы и социальные релизеры обнаруживают «двойное членение» точно так же, как и знаки семиотических систем человека, причём оно равно присутствует в материальной (демонстрирование, «уколы» или «давления» релизеров) и в информационной составляющей происходящего (семантика специфических форм демонстраций). Везде
1) значимые элементы (формы демонстраций) образованы из незначащих субъединиц (выразительных движений, ЭДА, факты 11-12);
2) действие целого, сигнальное или релизерное, не редуцируется к эффектам воздействия составляющих (предельное нарушение закона гетерогенной суммации А.Зайца11);
3) действенность каждого обеспечена инвариантностью форм, «вычерченных» при демонстрировании, т.е. сопряжённостью исполнения телодвижений и действий, образующих данное целое, т.е. корреляциями, изученными A.W.Stokes и последовавшими за ним этологами, включая меня самого (Фридман, 2013), и никогда — прямых изменением состояния реципиента от «уколов» или «давлений» релизеров данной особи. Каждое выразительное движение (вроде ЭДА, обсуждавшихся в факте 11) по отдельности лишь неспецифически возбуждает животное, воздействие целостных форм «составленных» из них демонстраций наоборот, имеет специфический и информационный характер.
Всё это делает демонстратора максимально уязвимым ко всяким опасностям. Жёсткое сопряжение разнородных телодвижений (моторно различных, «движущих» разные части тела) при демонстрировании одновременно мешает спасению жизни от нападения хищника (“поза враскоряку” увеличивает заметность и задерживает переход к броску в убежище, снижая начальную скорость последнего, а вероятность быть схваченным обратно пропорциональна ей) и понижает ловкость прямых действий агрессии, бегства, спаривания, сопровождающих демонстрации. Трудно точно клевать противника наряду с исполнением демонстраций, ещё трудней уворачиваться от ответных клевков, то же верно для насильных попыток сближения и спаривания. Оба момента увеличивают риск демонстрирования пропорционально эффективности выбранных демонстраций, первый за счёт риска гибели, второй — риска стресса, ранений и травм от прямого действия оппонента.
Биологический смысл риска демонстрирования здесь в «цене» за употребление данных средств, установленной отбором за употребление каждой из демонстраций в данных взаимодействиях, характеризующая данные единицы репертуара не меньше их эффективности, а вовсе не функция агрессивности оппонента. «Доверие» существует несмотря на случайность появления каждого из скоплений, его анонимность и незнакомство друг с другом птиц: не проходит обычное объяснение такого «доверия» персонализацией отношений у членов сообщества.
Будь социобиологические теории верны, «обманщикам» было б раздолье! Но видим противоположное: «доверие» к видовым сигналам абсолютно, «обман» отсутствует (Enquist et al., 1985). Вообще, любая неточность соответствия сигнала и ситуации, о которой сигнализируется, сигнала и ответа раз за разом оказывалась «ошибкой» или «сбоем» коммуникации, отнюдь не «обманом». Своеобразная ирония истории — всё это выяснилось в последние 35-40 лет монополии социобиологии в области поведенческих исследований и авторами, работающими в данной парадигме. Нарочно не придумаешь надежней опровержения социобиологической критики сравнительной этологии12.
Важность конкуренции за малоценный ресурс в исследовании сигнальной функции демонстраций связана именно с его несущественностью для выживания «здесь и сейчас», тем, что от него «всегда можно отказаться», и пр. Сам по себе он недостаточен как «награда» победителю (каким был бы, будь эта конкуренция чисто эксплуатационной). Однако его «розыгрыш» в коммуникации делается пробным камнем для выявления социально обусловленной разнокачественности индивидов, по результатам которой в сообществе устанавливаются уровни социально-обусловленной доступности всех ресурсов такого рода для всех особей, проранжированных соответствующими взаимодействиями.
Иными словами, реально «разыгрывается» не ресурс, а «место» индивида в социальной дифференциации группы, конкуренция происходит за социальный ресурс, выявленные M. Enquist et al. (1985) демонстрации суть посредники между действием одного и контрдействием другого индивида. Действительно, коммуникация считалась продемонстрированной, если в условиях конкуренции фиксируется значимая корректировка модели поведения реципиента под воздействием сигналов партнера. А успех (неуспех) коммуникации состоит не в ресурсе как таковом, но в выстраивании оптимальных отношений13 с определенными компаньонами по результатам информационного обмена в системе взаимодействий, где «оспаривание обладания» (статусом в системе иерархии, половым партнером в системе брачных отношений, территорией в лучшей или худшей части анизотропного пространства группировки) лишь повод.
Только оно делает доступ к экологическим ресурсам предсказуемым и гарантированным, будь то пища, самка (самец) в рамках брачных связей, безопасность и корм, доступность которых зависима от системы иерархии группы. По той же причине предсказуемость поведенческих изменений, наступивших под действием стимуляции от конкурента, для индивидов важней силы последней, независимо от того, кто выиграл и проиграл (Фридман, 2013).
Видим, что в «повышении планки» агрессии ритуализованной участвуют оба глупыша, т.е. эскалации открытой агрессии нет даже у будущего проигравшего, где она всё больше и больше превалирует над её ритуализованным выражением. Опосредование демонстрациями делает агрессивные действия в отношении оппонента (образующие риск демонстрирования) ещё одним способом — через прямое воздействие — «подчеркнуть» семантику каждой демонстрации, как бы указав «цену», которую обе особи «платят» за продолжение коммуникации, т.е. за информацию от сигналов друг друга, ценную даже при проигрыше, не только за собственно выигрыш.
Биологический смысл готовности рисковать ради успешного демонстрирования
Неслучайно «парадоксальная сдержанность» именно тех участников, которым достанется выигрыш, найдена у самых разных видов позвоночных каждый раз, когда их коммуникацию исследовали специально. Эти особи предпочитают стереотипное демонстрирование прямым действиям даже тогда, когда их преимущество уже выявилось в полной мере. Поэтому она может рассматриваться как универсалия, позволяющая утверждать, что успешные участники взаимодействия подвергают себя риску открытой агрессии со стороны менее успешных партнёров именно ради устойчивости приёма-передачи информации о необходимых следующих действиях через демонстрации – знаки-посредники в информационном обмене. Такой расчёт оправдывается у всех видов, где вообще возможен устойчивый и направленный обмен демонстрациями между особями, — даже при существенных различиях в физической силе, уровне мотивации, социальном ранге и статусе участников коммуникации, причём часто не в пользу особи, выбравшей демонстрирование вместо действия.
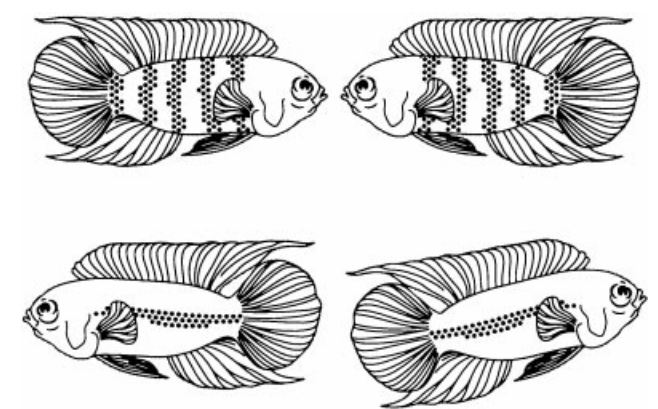
Поэтому опасность и неспецифическое беспокойство эффективно подавляют демонстрирование, будь то риск нападения хищника или общее беспокойство проходящими людьми, другими животными и т.п. За единственным исключением – коли организованный обмен демонстрациями уже начат, он будет доведён до конца, вопреки эскалации порождаемых им опасностей, как в примере с радистом. Подробней всего это исследовали (опять же!) социобиологи у южноамериканской цихлиды Nannacara anomala.
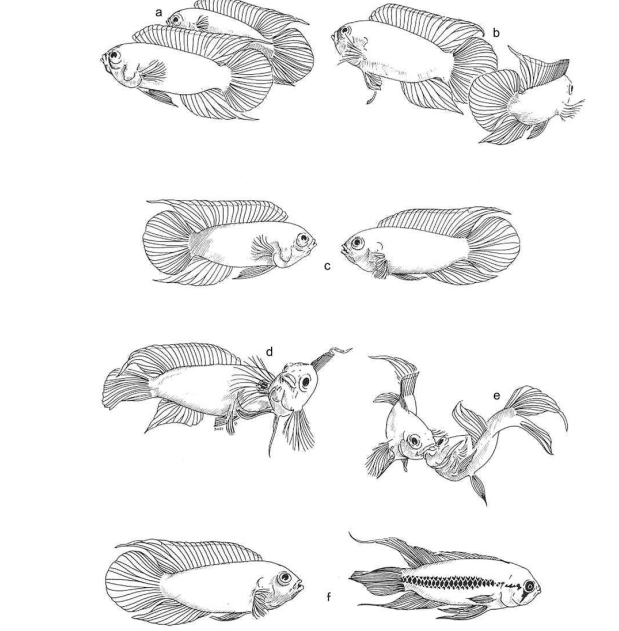
Авторы фиксировали обмены демонстраций в агрессивных взаимодействиях между сексуально активными самцами N.anomala, уже приобретшими самку (Hurd, 1997; Brick, 1998). Обмен демонстрациями всегда начинается с реакций низкой интенсивности (изменения окраски). Затем следуют ритуализированные удары хвостом, во время которых ярче всего выделяется продольная полоса медного цвета. Далее рыбы переходят ко всё более интенсивным и «экстравагантным» реакциям, таким как толчки рылом и «борьба ртов», когда выделены вертикальные полосы14. Сцепившись широко раскрытыми ртами, рыбы толкают друг друга рылом, как борцы сумо, пытаясь выкинуть противника за круг, очерченный движениями их тел. Максимум эскалации — «кружение» (англ. circling) рыб, стремящихся вытолкнуть друг друга (рис.5). При этом переходе оба противника с определённой скоростью «перебирают» все демонстрации ряда, без пропусков и без «забегания вперёд». Реакции высокой интенсивности более ритуализированные и исполняются с большей стереотипностью. Они более эффективны как релизеры, дающие значимый шанс подчинения и бегства оппонента, но и более рискованны: шанс спровоцировать прямую агрессию партнёра в ответ также растёт.
Анализ влияния риска хищничества на агонистическую коммуникацию показал, что у N.anomala, как и других позвоночных, риск поимки во время демонстрирования положительно коррелирует с прочими — риском столкнуться с прямой агрессией вместо ритуализованного ответа на демонстрацию и риском значительных затрат времени и энергии на безрезультатное взаимодействие. Неслучайно и выразительные движения, составляющие демонстрации — одни из самых энергоёмких видов движений; образованные ими устойчивые комбинации (=самоё демонстрации) требуют значительных затрат времени на развёртывание «типичных» последовательностей единиц в «типических» ситуациях взаимодействия.

А.
Б.
 В.
В.
Рисунок 5. А. Самцы N.anomala, вовлечённые в «борьбу ртов». Из: курс этологии в Aurora University, Иллинойс, США. Б. Изменения цвета в агонистических взаимодействиях N.anomala. Верхняя пара рыб демонстрирует оппоненту вертикальные полосы, нижняя — медиальную линию. Из: Hurd, 1997 В. Развитие агонистического взаимодействия N.anomala, стадии эскалации ритуализованной агресси от (a) к (f). Чем более сходны соперники по мотивации и/или способности бороться, тем дольше и напряжённей конфликт. (a) Демонстрации боком. (b) Удары хвостом. (c) Фронтальные демонстрации. (d) Укусы противника. (e) «Борьба ртом». (f) Успешное завершение конфликта: проигравшая рыбка опускает плавники, с потемнением средней линии вдоль тела. Из: Sloman et al., 2005, fig.5.1, цит.по Jacobsson, 1987.
Как везде, более эффективные демонстрации N.anomala и более рискованны: мера риска здесь — минимальная дистанция, с которой предъявление модели хищника вызывает бегство демонстрирующих особей. Выяснилось, что у обоих рыб (включая проигрывающую!) эта дистанция прогрессивно снижается по ходу коммуникации: максимальная в начале процесса, она падает в последующем, при переходе ко всё более интенсивным демонстрациями. Специальные исследования демонстраций, полярных по эффективности, «борьбы ртов» и ритуализированных ударов хвостом, также показывают что при дистанция бегства от хищника в первом случае много ниже, чем во втором.
Само удержание демонстраций во время конфликта не непрерывно: время от времени процесс прерывается раундами возврата к повседневной активности или к низкоинтенсивным реакциям. Предъявление модели хищника рыбкам, уже вступившим в обмен демонстрациями, вызывает реверсию процесса к демонстрациям низкой интенсивности и даже к открытой агрессии. В присутствии модели хищника даже «борьба ртов» длится в среднем значимо меньше, чем в отсутствие опасности. Однако более интенсивные демонстрации и более устойчивы к подобным реверсиям при предъявлении модели хищника: реверсия к открытой агрессии при показе модели до исполнения «толчков ртом» в 4-5 раз выше, чем когда демонстрация уже началась.
Между прочим, подобная логика относится не только к визуальным или акустическим, но к любым демонстрациям, включая ольфакторные. Метки на территории следует регулярно обновлять, иначе они деградируют и теряют действенность. Периодическое маркирование участка придаёт поведению владельца заметную дополнительную регулярность, которой легко воспользоваться хищнику или эффективному конкуренту, и создаёт ему дополнительные затраты времени или энергии, отклоняя от оптимума, соответствующего схеме обследования пространства только для целей кормодобывания и отдыха. Поэтому чем более ритуализовано поведение, и чем интенсивней демонстрирование, тем выше селективная смертность (и стресс), связанные с соответствующей формой активности.
Дальнейшие наблюдения авторов, опять же работающих в парадигме социобиологии, показали, что описанное в агонистической коммуникации глупышей, белоглазок, цихлид и т.д. справедливо для всех демонстраций, не только агонистических, не только для конкуренции за пищу, но за территорию, брачного партнёра и пр. На разных видах и в разных ситуациях конфликта — белоголовых орланов Haliaetus leucocephalus, кормящихся на трупах лососей, европейских Spinus spinus или американских чижей Spinus tristis в стаях (конкуренция за ранг в системе агонистического доминирования), мексиканских чечевичников на кормушках (место для кормления как функция ранга) — неизменно показывалось, что ритуализованные демонстрации несут информацию о намерениях реципиента. Благодаря этому они выступают предикторами набора допустимых ответов данному противнику (чаще всего также демонстраций, см. Enquist et al., 1985 и «20 фактов об инстинктах«), эффективней всего противодействующих ему как стимулы прямо сейчас, но не обрывающих взаимодействие, а продолжающих его, информируя (уже как сигналы и долговременно) о следующих «шагах» развития взаимодействия, в том числе демонстрациях, чей выбор и удержание максимизирует будущий шанс направить это развитие к выигрышу данной особи.
То и другое определяет возможности разрешения конфликта между обоими и снижает риск «срыва» последнего в драку, увеличивая устойчивость продолжения обмена демонстрациями, включая снижение частоты поранений и травм сравнительно с обычной дракой, где силовые действия особей друг в отношении друга не опосредованы демонстрациями. Из 541 агрессивных стычек белоголовых орланов Haliaetus leucocephalus на трупах лососей лишь в 10 наблюдали соприкосновение когтей с телом оппонента, в отсутствие опосредования действий демонстрациями это бывает намного чаще.
Резюмируя: дополнительные затраты на коммуникацию и дополнительный риск, вызванные участием в обмене ритуализированными демонстрациями, окупаются «качеством» информации, переданной-принятой с их посредничеством, т. е. развитостью предикторных свойств и долгосрочностью «прогнозов» проспективных потенций развития взаимодействия, выполненных его участниками (и, что особенно важно, «зрителями» процесса), с опорой на данные свойства, по результатам которых те и другие успешно корректируют модели поведения.
Для агонистической коммуникации чижей Spinus spinus J.C.Senar показано, что угрожающие демонстрации, которыми обмениваются птицы, это не аукцион, на котором распределяются ресурсы, и не экспрессивные жесты аукциониста. Это знаки возможностей разрешения конфликтной ситуации, в которую индивида ставит угроза особи-инициатора конфликта, и сигнал о возможностях выбора ответных действий, допустимых по отношению к данному оппоненту (в зависимости от того, выше рангом он или ниже), и эффективных против него. Поскольку эти ответные действия также представляют собой демонстрации и, следовательно, выбираются из элементов того же ряда угрожающих сигналов S.spinus, они продолжают процесс общения, а не обрывают его (Senar et al., 1989; Senar, 1990).
В развитие подхода J.C.Senar я доказываю, что появление этих специализированных «языков» для организации устойчивого информационного обмена внутри популяции – не редкое исключение, а общее правило для разных групп позвоночных. На определённом этапе эволюционной оптимизации системы отношений особей в группировках соответствующий «язык» появляется всегда и, раз появившись, направленно совершенствуется под действием тех же самых давлений отбора.
Видим: коммуникация тем «прибыльней» для особи и важней для сообщества, чем выше эффективность сигналов и наоборот: возникает положительная обратная связь, способствующая направленному совершенствованию систем сигнализации каждой отдельной группы после того, как в эволюционном прошлом этого филума действием тех же факторов из набора воздействий они превратились в семиотическую систему.
Заключение
Действительно, с эволюционной точки зрения разумен вопрос №1: ради чего участники коммуникации, демонстрируя, навлекают на себя дополнительный риск одновременно извне (хищник) и внутри коммуникативной системы (стресс, травма, ранение), ведь напряжённое положение тела, статистически невероятное в повседневной активности (факт 14) увеличивает то и другое одновременно? Единственно возможный ответ: ради ценности информации, передаваемой инвариантами форм данных демонстраций. Лучшая (более быстрая и точная чем, у оппонента) корректировка собственной модели поведения на основании сигнальной информации — единственная возможность выиграть взаимодействие, одной только силой “давления” и/или интенсивностью “уколов” релизеров здесь не обойтись.
В том числе потому, что “хороший стимул — плохой сигнал, и наоборот”, а нужна именно сигнальная информация о потенциальных возможностях (как сказал бы эмбриолог, проспективных потенциях) развития взаимодействия на следующих “шагах” процесса. Релизеры могут только “сбивать” оппонента, снижая стереотипность демонстрирования и точность восприятия сигнальной информации в демонстрациях другой особи, но одним “силовым воздействием”, ритуализованным или нет, ни конфликта не выиграешь, ни ухаживанию не дойти до биологически осмысленного результата, телесного контакта и спаривания партнёров.
Отсюда же следует: риск — специфическая характеристика самой демонстрации, а не обстановки вокруг, такая же, как её эффективность как релизера и/или сигнала, риск порождён самим демонстрированием для задач коммуникации с должной стереотипнотстью и т. д., а не обстоятельствами контекста.
Вопрос №2: почему, уже вступив в этот обмен, животное вопреки социобиологической логике всегда наращивает рискованность поведения, переходя ко всё более эффективным демонстрациям, «повышая планку» ритуализированного ухаживания или угрозы, вместо того, чтобы остановиться на некотором оптимальном уровне хотя бы после того, как победитель уже определился? Но нет, коммуникативные взаимодействия животных продолжаются много дальше этого момента, и риск продолжает увеличиваться сверх того уровня, который оптимален для индивида «здесь и сейчас» (в смысле, чтобы одолеть противника и зафиксировать своё доминирование над ним).
Ответ прост: демонстратор дольше готов рисковать ради получения информации, последовательные «порции» которой (считываемые с демонстраций противника и отправляемые с собственными демонстрациями) заблаговременно оптимизируют модель поведения не только на «следующем шаге» процесса, но и на более долгое будущее. Это позволяет животному структурировать последовательности демонстраций следующего этапа взаимодействия так, чтобы максимизировать вероятность выигрыша у данного противника при минимизации вышеназванных рисков. Всё перечисленное увеличивает предсказуемость поведения участников друг для друга и даже для «зрителей» процесса: в подобной системе честно передавать информацию о возможностях разрешения конфликта так же выгодно, как получать её от партнёра. Последнее эффективным образом исключает «обман» или, точней, сдерживает его в неких приемлемых рамках, заданных необходимым уровнем точности / однозначности распознавания сигналов, точностью осуществления сигнальной информации в поведенческих изменениях участников и «зрителей» процесса и интегральным критерием необходимого уровня эффективности коммуникации.
Отсюда единственно возможная «плата» за риск — сигнальная ценность специфических форм демонстраций, позволяющих действовать в проблемной ситуации не только с наибольшей возможной успешностью, но и инстинктивно (в том числе в смысле «автоматически», без собственного отслеживания поведения источника проблемы, будь то появившийся хищник или захватчик, навязавший конфликт). Стереотипное воспроизведение этих форм связано со значительным риском, но не удовлетворяет немедленно никаких внутренних побуждений животного (в отличие от прямых действий агрессии или спаривания), «давление» или «уколы» их как релизеров «проявятся» в виде долговременных изменений активности оппонента только при положительном исходе всего взаимодействия, без «срыва» в обоюдный конфликт или разлёта из-за потери интереса друг к другу.
Анализ рисков и затрат демонстрирования ясно показывает, что вопреки мнению сравнительных этологов, биологический смысл демонстраций как комплексов фиксированных действий (англ. FAP) не в самих действиях, усиливающих выразительность/экспрессию пропорционально усилению побуждений, не в эффектах воздействия демонстраций как социальных релизеров, удовлетворяющих данные побуждения, но в «сигнальности» форм демонстраций видового репертуара, эволюционировавших как специфические средства передачи информации, разрешающей проблемные ситуации в определённых типах «конфликтов интересов» особей, значимых для естественной истории каждого вида, вроде «патрулирования» у разных видов воробьёв («20 фактов об инстинктах«, факт 17) или внутри- и межвидовой охраны территорий у трёх видов поморников Strecorarius. Отсюда закономерная реализация специфических «значащих структур» и специфических образов, репрезентирующих эти структуры партнёру по ходу взаимодействия, позволяющая говорить об организованном обмене демонстрациями, описывать временную организацию данного процесса как его «синтаксис» и т. д. Факторы и механизмы эволюции, превращающей прямые действия особей в отношении друг друга, детерминированные побуждениями, в специфические средства коммуникации, сигнализирующие о потенциальных возможностях разрешения конфликтов, рождённых побуждений особей, в социальной организации вида, анализируются в нашей работе «Как социобиология сама себя отрицает«.
Поэтому увеличение опасности для вида, уже обладающего развитой системой сигнализации, вместо ожидаемого ослабления демонстрирования, или полного отказа от него, чаще всего контринтуитивно увеличивает сигнальность и знаковость демонстраций. Парадоксальный ответ коммуникативных систем самых разных видов на рост рисков коммуникации в их среде обитания состоит в «совершенствовании сигнальности» демонстраций15.
Следовательно, биологический смысл демонстраций как комплексов фиксированных действий животного состоит не в (1) совершении самих действий с увеличенной выразительностью/экспрессией сообразно усилению побуждений, и не (2) в специфическом релизерном эффекте воздействия демонстраций, удовлетворяющем последние, как считали классические этологи, но в (3) закономерной реализации специфических «значащих структур» и специфических образов, репрезентирующих эти структуры партнёру в специфических ситуациях взаимодействия, связанных с закономерным появлением дифференцированных проблемных ситуаций по ходу развёртывания процесса. Стереотипная репрезентация форм видовых демонстраций важна здесь не столько как стимул, сколько как сигнал: закономерная реализация «должного» образа демонстраций в «должные» моменты конфликта информирует обоих участников о дифференцированных проблемных ситуаций, закономерно сменяющих друг друга по ходу процесса, и возможностях их успешного разрешения на следующих «шагах» развёртывающегося процесса.
Непосредственный смысл поведения особи, выбранного и реализуемого сразу после компетентного участием в прошлом акте коммуникации – реализация полученной сигнальной информации в новой (изменённой) структуре деятельности животного, корректирующей прошлую модель поведения ради более эффективного участия в следующих актах той же коммуникации. Почти всегда эта реализация прошлой сигнальной информации в поведении «следующего шага» взаимодействия одновременно есть участие в следующем акте коммуникации, подача специфических сигналов о качественном изменении ситуации взаимодействия за период удержания данного сигнала с изменениями, произошедшими под его влиянием. Специфическое релизерное, возбуждающее или иное действие на партнёра здесь — лишь средство увеличения действенности сигнала, «поданного» предъявлением специфических форм демонстраций, например, подавляя сопротивление оппонента. Подобная «лесенка» (процесс предъявления каждого следующего сигнала реализует в поведении информацию прошлого и сам сигнализирует об изменениях ситуации взаимодействия, произошедших вследствие этой реализации) коммуникация во взаимодействии развивается автокаталитически, этот процесс обладает собственной устойчивостью и направленностью, несмотря на массу внешних и внутренних факторов, способных его остановить и сорвать (Фридман, 2013).
Примечания
1См. Фридман, 2017аб и «Что не так с социобиологией?» https://www.socialcompas.com/2017/08/23/chto-ne-tak-s-sotsiobiologiej/; «Преодоление социобиологии», https://www.socialcompas.com/2014/06/08/preodolenie-sotsiobiologii/ . Кроме неустранимых противоречий в теории и плохого соответствия эмпирическим данным, что мне не нравится в социобиологии философски или, скорей, гносеологически — она превращает исследования поведения в упражнение по части счастливых догадок. На что ни упадёт взор социобиолога — на внебрачные копуляции у моногамных видов, или систему расселения молодняка, или помощничество у птиц/млекопитающих, или обмен демонстрациями в социальном общении — немедля оказывается, что это чем-то адаптивно и выгодно, да ещё не для особей, худо-бедно способных к саморегуляции, но для самих «эгоистических генов», которым на окружение наплевать.
А ведь сложную систему нельзя оптимизировать сразу более чем по одному параметру: адаптивны изменения лишь обеспечивающей его структуры (-р). Все прочие трансформации представляют собой или подстройки по корреляциям, чтобы организм выжил, несмотря на изменения ключевых характеристик отбором (эффективный отбор неизменно снижает жизнеспособность, адаптивная эволюция везде и всегда неотделима от риска вымирания) или, ещё хуже, просто дестабилизацию и дезорганизацию «давлением» чересчур быстрого и/или жёсткого отбора, см.https://vk.com/wall-59576852_4655. Им приписывать адаптивность можно лишь через анализ тенденциозно собранных данных, т. е. делая нечто среднее между ошикой и фальсификацией.
Уместен вопрос: каковы шансы, что взгляд наблюдателя, в начале исследований заведомо незнакомого с «устройством» социальной жизни вида, упадёт именно на её ключевые моменты, дающие адаптивную выгоду (или «поправка» которых отбором даст оную выгоду)? Они ничтожно малы, тем более что социобиологи обычно не публикуют отрицательных результатов, мол, мы анализировали особенность поведения Х, но адаптивности не нашли, её специфика — следствие следствие чего-то другого или дезорганизация. Неудивительно что данные в пользу самых разных социобиологических построений — от родственного отбора до «неверные самки ищут лучшие гены» — сплошь и рядом или некачественны, или обусловлены контекстом, а не самим феноменом (Фридман, 2017аб). См., скажем «Социальный отбор против полового у чёрных лебедей и пеганок», https://www.socialcompas.com/2025/10/02/sotsialnyj-otbor-protiv-polovogo-u-chyornyh-lebedej-i-peganok/ ; «Взаимоотношения полов позвоночных и модели полового отбора», https://www.socialcompas.com/2019/12/15/vzaimootnosheniya-polov-u-pozvonochnyh-i-modeli-polovogo-otbora/ .
Панадаптационизм до добра не доводит, особенно в ситуации, когда генетиками надёжно показано, что отбору подвергаются фенотипы, а не отдельные гены, идея «эгоистичного гена» в ряде случаев (достаточно ограниченных) может быть полезной фикцией, но не онтологией и даже не универсальным объяснением.
2Жиль Перро «Красная капелла» (М.1990).
3См. подробней «Опасности коммуникации», https://www.socialcompas.com/2025/10/17/opasnosti-kommunikatsii/ ; «Риск гибели от хищника растёт с эскалацией социальных взаимодействий», https://www.socialcompas.com/2025/10/06/risk-gibeli-ot-hishhnika-rastyot-s-eskalatsiej-sotsialnyh-vzaimodejstvij/
4И жёстче скоррелированность исполнения телодвижений, моторно различных, не связанных между собой (факт 18 и «Вольфганг Шлейдт, Ноа Эшкол и концептуальный кризис сравнительной психологии»). Дальше увидим, что именно прямая зависимость эффективности демонстрирования от стереотипности исполнения делает демонстратора больше подверженным всем трём видам риска — в меру вычурности/неественности образов исполняемых телодвижений, созданной жёсткой сопряжённостью исполнения их во времени при разной природе (происхождении из разных видов повседневной активности — чисток, локомоции, кормодобывания, агрессии и пр.).
5Это «повышение планки» ритуализованной агрессии — основной тренд развития процесса коммуникации (факт 19-20).
6в противоположность неритуализованным дракам птиц и млекопитающих, где нет опосредования действий демонстрациями.
7хотя таковая у них имелась, социобиологическая и, как дальше увидим, неверная.
8Эволюционно-стабильная стратегия (англ. ESS, evolutionarily stable strategy) – модель поведения животного при конкуренции в определённом кругу социальных компаньонов (особенно агонистического, но также и полового, и прочих), которая, будучи принята достаточным числом особей, не вытеснима какой-то другой доступной им стратегией, благодаря лучшему соотношению эффективности и риска. Предложенное Дж.Мэйнардом Смитом в начале 1970-х при приложении теории игр к поведению животных, это понятие «биологизирует» известное равновесие Нэша и широко использовалось социобиологами в анализе агонистических взаимодействий и, шире, коммуникации животных (Maynard Smith, Proce, 1973; Maynard Smith, Parker, 1976).
Обычно они это делали «в лоб», со странными результатами (Maynard Smith, Harper, 2003; Searcy, Nowicki, 2005, анализ и критику см. Фридман, 2013, 2017аб), однако интересными и полезными, если это делать с умом, учитывая качественное различие трёх составляющих репертуара — ритуализованных демонстраций, чьё значение как сигналов и/или эффекты как стимулов (социальных релизеров) равно используются в коммуникации; прямых действий агрессии, спаривания, чистки и пр., непосредственно удовлетворяющих побуждения особи, особенно в поведении самообеспечения; экспрессивных реакций, выражающих уровень общего (неспецифического) возбуждения животного во всех перечисленных ситуациях.
Как и любые другие единицы поведения, наблюдатель имеет право “стратегию” выделить и описать как отдельность лишь в случае, если сами животные реализуют данное поведение так, что его форма (визуальный, акустический и т.д. образ) типологически определённая и дифференцированная от форм других особей, т.е. можно говорить о реальности её существования для самих особей.
9Или, что много важнее, из «зрителей» взаимодействия, в него непосредственно не вовлечённых, не «испытывающих на себе» ни стимуляции оппонента, ни собственного возбуждения от борьбы.
10 перечислены в порядке падения сигнальной эффективности и одновременно заметности демонстраций, их энергетической цены и т. п. «затрат» демонстрирования.
11См.подробней «Закон гетерогенной суммации Альфреда Зайца»
12как и её критики Е.Н.Пановым (1978, 2012), полагавшим эффективность и риск артефактом, статистически неустойчивым и зависимомым от контекста, хотя чисто a priori, без специальных исследований, чисто на вере в справедливость своих представлений о коммуникациии. Евгений Николаевич назвал их «солидно», системными, хотя верно «теория бесструктурного шума», поскольку отстаивала 1) специфический результат коммуникации достигается неспецифическими средствами, простым ростом неспецифического возбуждения участников, рано или поздно, через множество проб и ошибок, синхронизирующих эндогенные ритмы друг друга (хотя нельзя объяснять непонятное неизвестным — а данные ритмы, как и стимуляция, ненаблюдаемы для полевого этолога, они им додумываются); 2) единицы поведения, выделенные этологом, и дальше тестируемые как единицы коммуникации, стимулы и/или сигналы, просто кажимость, введённая им для упрощения процедуры описания поведения, отнюдь не реальность, которую можно установиться и проверить. Сегодня мы знаем: надежно установленные факты демонстрирования и/или использования демонстраций во взаимодействиях всецело опровергают 1) и 2).
13Оптимальных по степени социальной асимметрии в отношениях данного индивида с другими, которую ему при его «силах», «активности» и «способности бороться» удается поддерживать с максимальным соотношением «выигрыша» к «плате» (Enquist, Leimar,1987).
14Как и в прочих подобных взаимодействиях, демонстрации видового репертуара «перебираются» в направлении роста эффективности. См. подробней «Зачем большим пёстрым дятлам всю зиму охранять территории?», https://www.socialcompas.com/2020/04/26/zachem-bolshim-pyostrym-dyatlam-vsyu-zimu-ohranyat-territorii-2/ В этом «повышении ставок» эффективности и риска демонстрирования каждая рыбка старается опередить противника, но одновременно не потерять устойчивость собственной траектории перебора сигналов под влиянием сбоев, вызванных релизерным эффектом демонстраций противника с сопровождающими их «выплесками» агрессии. Для этого следует, постоянно опережая его «на полшага», не сильно «забегать вперёд». Сходно структурированы взаимодействия ухаживания и угрозы у других позвоночных: фактически это универсалия, детерминирующая «синтаксис» употребления ритуализованных демонстраций в социальных взаимодействиях.
15См.подробней «Опасности коммуникации»
Рекомендуем прочесть
 Пение самок воробьиных птиц: исключение или закономерность?
Пение самок воробьиных птиц: исключение или закономерность? Что не так с критикой Ильенкова
Что не так с критикой Ильенкова Глобальное потепление усиливает погодные экстремумы
Глобальное потепление усиливает погодные экстремумы Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных- Интеллектуальная акселерация (эффект Флинна): метаанализ
 Ревень Rheum palaestinum, самоорошающееся растение пустынь Израиля
Ревень Rheum palaestinum, самоорошающееся растение пустынь Израиля Быстрая перестройка фенологии в новых местах гнездования ласточек
Быстрая перестройка фенологии в новых местах гнездования ласточек Синдром темпа жизни (pace-of-life syndrome, POLS): эволюция концепции
Синдром темпа жизни (pace-of-life syndrome, POLS): эволюция концепции
