

1. Проблемы теории
Содержание
- 1 1. Проблемы теории
- 2 2. Инфантицид – дезорганизация, а не адаптация
- 3 3. Игра вето против дилеммы узника
- 4 4. Синицы «не сачкуют» при выкармливании птенцов
- 5 5. Плюсы социобиологии
- 6 6. Взаимоотношение полов в сексе и в заботе о потомстве не соответствуют социобиологическим предсказаниям
- 7 7. Про «избегание инбридинга» и логику системы
Возьмём социобиологическую идеализацию «эгоистических индивидов» (все силы и побуждения которых направлены именно и только на максимизацию итоговой приспособленности). Она полезна в той же степени, в какой полезна модель «идеального газа», показывает некое нулевое состояние полного отсутствия взаимосвязей в системе, отталкиваясь от которого мы должны объяснить появление и прогрессивное развитие таких связей из взаимодействия между элементами. В таком случае сразу же возникает вопрос – откуда в системе, построенной на конкурентных столкновениях между «эгоистическими индивидами», появляется «общий интерес» в виде устойчивого воспроизводства системного целого в череде поколений, в которое каждый индивид, реализуя собственные активности, должен «вкладываться» во взаимодействиях с другими индивидами до того, как может рассчитывать на соответствующий выигрыш.
И очень задолго до того: скажем, самцы территориальных воробьиных «вкладываются» в поддержание пространственно-этологической структуры группировок в апреле – мае, а «выигрыш» в виде репродуктивного выхода получается только в июле-августе. Причём на размеры выигрыша сильней влияет благоприятная погода в апреле – мае, то есть в период коллективного построения структуры системы, а не в июне-июле, в период выкармливания потомства каждой парой индивидуально. Об этом есть интересная статья Л.В.Соколова в «Зоологическом журнале», где также разобрано влияние глобального потепления климата на миграцию и гнездование птиц в ХХ веке, очень рекомендую.
Понятно, что «натягивание» пространственно- этологической структуры популяций на определённой территории и есть тот «общий интерес», который даже эгоистические индивиды должны поддерживать, как минимум месяц-два,0 не рассчитывая на немедленный выигрыш. Так откуда берётся способность к этому в данной системе?
Опять же, максимизировать итоговую приспособленность нельзя в одиночку, надо объединяться с другими особями, для того чтобы долговременно и устойчиво эксплуатировать их как социальный ресурс. Так самец в рамках социобиологической модели «войны полов» эксплуатирует самок для максимизации распространения собственных генов, давая им взаимен не столько лучшие гены, сколько лучшие родительские качества, а доминант в системе агрессивного доминирования эксплуатирует готовность подчиняться подчинённых особей, чтобы по их склоненным спинам забраться повыше самому. Откуда берётся эта способность к кооперативному поведению с другими особями, если каждая из них исходно нацелена на конкуренцию и противостояние усилиям своих контрагентов?
Наконец, откуда берутся сигналы, позволяющие «эгоистическим индивидам» ориентировать поведение на этот «общий интерес», чтобы «рассчитать заранее» оптимальный размер собственных инвестиций в коллективное воспроизводство структуры системы – чтобы и не остаться вне её, и чтобы остались силы на занятие «лучшей позиции». И чтобы ориентировать поведение на долговременный выигрыш в виде лучшей «позиции» в структуре системы, в противоположность постоянному соблазну немедленного удовлетворения побуждений, которое устойчивость социальной «позиции», наоборот, разрушает. Понятно, что подобные системы сигналов, используемые особями для разрешения конкурентных взаимодействий агонистического или сексуального характера, и есть те самые групповые адаптации, появление и прогрессивное развитие которых социобиология объяснить не может и вынуждена отступать к намного более уязвимым теориям группового отбора.
Вместе с тем объяснить появление групповых адаптаций довольно легко, начав с чисто социобиологического рассмотрения ситуации. Надо только не бояться выйти за пределы, где кончается социобиология как научная теория и начинается уже догматический взгляд.
Если вовлечённость особи в некую проблемную ситуацию обязательно влечёт за собой мотивационный конфликт, то простой анализ показывает, что соответствующий конфликт в принципе неразрешим без введения системы сигналов-посредников, информирующих животное о возможности и эффективности выбора определённых моделей поведения, соответствующих ситуации. Такие сигналы лишь образуются комбинированием выразительных движений животного, определённое сочетание которых выражает определённые состояния и намерения подобно пантомиме.
Но, функционируя как сигналы-посредники, информирующие «всех» заинтересованных особей о выборе специфических моделей поведения в специфических проблемных ситуациях взаимодействия, эти самые комплексы действий уже «относятся» не к тому индивиду, движениями которого нужные демонстрации воспроизводятся в нужные моменты процесса. Они «относятся» к виду в целом и к видоспецифической социальной организации, в которой обмен соответствующими структурами поведения поддерживает и обсуживает информационный обмен. Отдельные особи только «пользуются» сигнальной информацией, воспроизводя соответствующие формы сигналов в нужные момент процесса, как бы берут видовые сигналы в лизинг.
Общая схема рассуждения заимствована из книги Докинза «Эгоистичный ген» (1993), но цель логического анализа прямо противоположна. Я хочу показать необходимость появления групповых адаптаций (в частности, общих сигналов, «честно» ретранслирующих «позиционную информацию» всем участникам взаимодействия) в системе, основанной на свободной конкуренции «всех со всеми», для того чтобы поведение каждого участника – эгоистического индивида – было закономерным и предсказуемым для всех, в том числе для него самого. В этих условиях «честно» передавать информацию оказывается также выгодно, как её получать, ведь переданная информация делает поведение партнёра более предсказуемым в той же степени, что полученная – поведение самого индивида, и вполне очевидны выгоды от возможности прогнозировать поведение партнёров, не утруждая себя дополнительным отслеживанием ситуации внутри сообщества, лишь на основании сигналов.
Представим себе животное — «эгоистического индивида» — поставленное в условия острой конкуренции за территорию или партнёра виду необходимости максимизировать итоговую приспособленность. Например, морского льва на лежбище или тетерева на току.
Готовность ко включению в структуру группировки требует решения проблемы, какое место в составе группы именно мне необходимо занять, с какой именно особью вступить в конкуренцию, чтобы его собственный баланс платы и выигрыша сошёлся с максимальным плюсом. С одной стороны, если особь — потенциальный противник уверенно вторгается на мою территорию, сразу начиная её рекламировать, или если он обладает наибольшим гаремом на участке, расположенном в центре колонии, то мне прямой смысл оказывать максимальное противодействие, оспаривать у данной особи её ресурсы. Ведь само поведение данных индивидов «подсказывает», что именно «их» ресурсы — лучше, а их стратегия поведения весьма эффективна, и моей стратегии поведения следует её превзойти.
Следуя этим побуждениям, индивид должен следовать самой агрессивной и самой конкурентной из всего диапазона поведенческих стратегий, вообще возможных в данной ситуации. Этот диапазон можно проградуировать по шкале «максимальное давление на противника, подчинение противника своим собственным амбициям ↔ максимальная «уступчивость» амбициям противника, подчинение им собственного поведения»; соответственно, на основании вышеприведённого индивид должен осуществлять максимальное давление на наиболее мощного противника и тем самым реализовывать наибольшую мощность поведения, какую он может реализовать.
С другой стороны, то же самый чувственный опыт столкновений с противником в то же самое время у той же особи вызывает побуждения прямо противоположного рода. Ведь если оппонент так уверенно действует и /или владеет столь ценным ресурсом. значит он его в состоянии защитить от покушений аналогичных моим собственным и уверенно побеждает противников, аналогичных мне [так могло бы рассуждать животное, если соответствующие побуждения у него бы отображались в мыслях].
Этот ряд побуждений требует минимизировать риск поражения и сразу занять подчинённое положение относительно реализации поведения другой особи. Этот «вывод» следует из непосредственного чувственного опыта индивида с той же определённостью, что и прямо противоположное «решение» активно оспаривать соответствующий вожделенный ресурс. Очевидно, индивидам, постоянно оказывающимся в проблемной ситуации подобного типа, соответствующий конфликт нельзя разрешить, действуя на основании лишь своих побуждений и своих наблюдений за ситуацией – побуждения противоположны и взаимно несовместимы, наблюдения приводят к двойственным выводам. Необходима информация, передаваемая сигналами извне, о том, какую пропорцию «наступательной» и «уступательной» компонент животному оптимальней всего выдерживать в столкновении с данным противником.
Почему именно информация, не воздействие стимула? Ведь оно тоже может сместить равновесие в пользу доминирования одного из двух побуждений и таким образом разрешить мотивационный конфликт.
Дело в том, что обоюдный мотивационный конфликт, описанный выше, вызван столкновением стратегий поведения соответствующих индивидов в конкуренции за соответствующий социальный ресурс (повторив рассуждения, приведённые выше, легко видеть, что и противник «нашей» особи испытывает тот же самый конфликт).
Разрешить внешний «конфликт интересов» (а конфликт мотиваций служит просто внутренним отражением внешнего конфликта) можно единственным способом — через дифференциацию поведенческих ролей. Тогда в каком-то спектре мест, участков, территорий, и/или в неком спектре ситуаций животное реализует поведение, соответствующее максимальному давлению и максимальным притязаниям на соответствующий ресурс, а в других, альтернативных первым – поведение, соответствующее максимальному подчинению притязаниям партнёра. Его оппонент в тех же случаях поступает, очевидно, прямо противоположным образом, так что поведение обоих противников в прошлом конфликте, конкурировавших за общий социальный ресурс, оказывается взаимно-комплементарным, так что между ними возникает социальное связь, делающая их активность взаимозависимой не хуже чем между А и Т в ДНК-РНК.
Прочная социальная связь обязательна, чтобы соответствующее разделение ролей было устойчивым и обе особи в полной мере могли получать выгоды, связанные с подобной информацией в виде меньших затрат на взаимодействие и большей лёгкости достижения результата в каждой из ситуаций, где особи с дифференцированными ролями встречаются для взаимодействия. Очевидно, набор типологически отдельных «ситуаций», в которых одна «особь» реализует амбициозное, а другая – «уступчивое» поведение, должен быть общим для обеих особей и, шире, для всех потенциальных конкурентов за соответствующий социальный ресурс – «лучшую» территорию, «подходящую» самку и пр. Общим должен быть (или, точнее, стать) и набор градаций «давления» на соседа и реализация собственного поведения «в уступку» ему.
А, значит, нужна информация, где и как оптимальней всего обоим дифференцировать активности, в каком хронотопе один будет реализовывать доминантную модель поведения, в каком – уступительную, а где то же самое будет делать его социальный партнёр.
Более того, соответствующие градации ситуации и градации моделей поведения, реализуемых в соответствующей ситуации, должны быть дискретны и типологически разделены. Ведь переход от одной градации к другой требует переключения особей на запуск противоположных аспектов стратегии, реализованных элементами поведения с противоположными свойствами, а не плавного перехода от одних элементов к другим, смежным с ними.
Из такой организации конкурентной системы очевидно, где именно возникают знаки-посредники, определённая форма которых указывает на тип проблемной ситуации, рождённой активностью данного компаньона и одновременно на некую программу поведения в следующий период взаимодействия. Причём допустимой именно по отношению к данному партнёру, и такую, которая будет конструктивным решением проблемы в смысле, эффективным шагом к созданию асимметричной социальной связи, «снимающей» исходный мотивационный конфликт. Этот «шаг» в виде выбора определённой демонстрации в ответ на демонстрацию партнёра, сам изменяет характер проблемной ситуации в соответствующем направлении, так как каждая демонстрация одновременно является сигналом о характере проблемной ситуации и стимулом, воздействующим на партнёра в соответствующем направлении. Своей реакцией на сигнал первый партнёр делает свой следующий «ход» и т.д., замыкая положительную обратную связь сигнального обмена (сигнал→ответ→следующий сигнал) и тем самым делая следующий «шаг» к разрешению исходного конфликта через создание асимметричной социальной связи.
Итак, перманентное столкновение «эгоистических» индивидов требует конституирования в конкурентной системе некой типологии ситуаций, в которых возникают разные варианты конфликта интересов, и параллельно с ней — типологии программ поведения, способных конструктивно «снять» соответствующий конфликт и ординированных по шкале «максимальное давление на противника» — «максимальная уступчивость его амбициям. Во-вторых, внутри каждого отдельного индивида «конфликт интересов» вовне конституируется в его внутреннем мире в виде конфликта двух противоположных интенций реагирования на поведение компаньона (давление – уступка, при установлении отдельных градаций каждой из них).
Как только конституируются определённые градации существенных [для системы] проблемных ситуаций и градации поведенческих программ, эффективно разрешающих проблемные ситуации (и в этом смысле адекватные им), они ставятся во взаимно-однозначное соответствие друг с другом, а также с соответствующими уровнями «конфликта мотиваций» внутри индивидов. Причём ставятся общеупотребительным способом, признанным и понятным как для резидентов, уже включённых в систему, так и для «активных вселенцев», старающихся включиться в неё.
В этом случае совершенно неизбежно появление сигналов-посредников между типовой «проблемной ситуацией» в системе и «типичной» категорией конкурентных усилий, разрешающих соответствующую проблему, для того, чтобы информировать особей о тех и о других. Соответственно, некоторая определённая часть движений и действий животных, связанных соответствующими конкурентными отношениями, должна быть отдана в распоряжение социальной системе для обмена информацией системного характера и частично выведена (эмансипирована) из-под силового управления (термин Л.Б.Белоусова) стимуляцией извне, чтобы они могли осуществлять параметрическое управление в системе-социум.
Дальше процесс ритуализации уже «доводит» соответствующие движения и действия до максимальной эффективности функционирования в новой роли сигналов и тем самым превращает их в «демонстрации» (в понимании классических этологов). Однако при этом не прекращается их действие как специфических или неспецифических стимулов в тех довольно обычных случаях, когда коммуникативный режим выдержать не удаётся и взаимодействие приобретает некоммуникативный характер (по крайней мере, не прекращается сразу).
Таким образом, знаки системы, передающие информацию о выборе действий в проблемной ситуации, берут начало из признаков, специфически связанных с этой ситуацией и этим действием в силу самой природы вещей. Отсюда знаковые системы в социальной коммуникации животных развиваются на основе отличительных признаков и характерных черт тех самых действий и программ поведения, которые связаны с разрешением соответствующих ситуаций путём ритуализации, типизации и подчёркивания их наиболее отличительных черт.
Всякое прямое действие имеет потенцию превращения в демонстрацию как только с точки зрения индивида его связь с определённой ситуацией вовне и мотивацией внутри особи становится устойчивой и регулярной (законосообразной). Точно также всякая уже преобразованная демонстрация имеет потенцию к превращению в знак, если только с точки зрения системы остановлены взаимно-однозначные соответствия между проблемной ситуацией в социальной системе и стратегиями действия особей – её элементов по разрешению соответствующей ситуации, где эта демонстрация одновременно является и знаком, и действием.
По мысли В.Г.Черданцева, естественный отбор вообще оперирует признаками, а не структурами. Удачный выбор признаков позволяет различать объекты независимо от структуры. Паттерн наиболее существенных отношений между особями, образующий социальную организацию, должен контролироваться отбором с особенной жёсткостью. Причём это стабилизирующий отбор, поддерживающий устойчивость воспроизводства «идеальных форм» социальной организации вида вопреки всему множеству «возмущений», созданных нестабильностью экологической среды, демографического состава, переменностью статуса и состояния особей и пр. Чтобы соответствующий отбор был действенным, нужны признаки, «указывающие» на возможности эффективного воспроизводства соответствующих отношений в системе.
А поскольку воспроизводство отношений в системе-социум осуществляется динамически, через их разрушение в столкновении индивидуальных амбиций, создающих «конфликт интересов», проблемную ситуацию определённого типа, то признаки, с которыми «работает» соответствующий отбор – это сигналы возможности разрешения конфликта одним из n «предусмотренных системой» исходов процесса. Поскольку последние в момент столкновения индивидов ещё не реализованы и находятся только в потенции, возможность реализации этих возможностей (сорри за тавтологию) индивиды не могут непосредственно ощутить и с трудом ощущают, отслеживая изменения ситуации, так что вынуждены ориентироваться преимущественно на сигналы, «доверять» им более, чем непосредственным ощущениям от ситуации, мотивационным изменениям под воздействием ситуации и т.п. несигнальным воздействиям.
Поэтому сигналы животных, возникающие под действием отбора на оптимизацию системы, построенной на отношениях конкуренции за социальный ресурс, имеют значение «указателей» на реляционные характеристики индивидов (большая или меньшая способность бороться за социальный ресурс и возможность вести борьбу на данном уровне успешности именно в данном конфликте), а не на собственные характеристики вроде силы, агрессивности и т.п. «показателей качества». Соответственно, система сигналов этого рода оказывается в большей или меньшей степени символической и, главное, развивается в сторону всё большей и большей символизации или не развивается вообще.
Откуда берутся такие сигналы, тоже понятно — из экспрессивных телодвижений, связанных с возбуждением (или стрессированным состоянием) особи в связи с данной проблемной ситуацией. Для n дифференцированных ситуаций взаимодействия, отражающих разные градации изменения того, «о чём сообщается» в значениях сигналов, уровни возбуждения или стресса, или иной неспецифичной реакции также будут различны, что отразится на внешнем облике, интенсивности и других параметрах экспрессий. Дальше эти первичные различия между протосигналами усиливаются в процессе ритуализации, превращающем экспрессивные телодвижения в демонстрации, и в процессе означивания означивания, трансформирующего ряды демонстраций, выстроенные вдоль мотивационно-типологических градиентов агрессии, сексуальности и т.п. в специализированные знаковые системы. Тем уже формы разных сигналов ряда «строятся» в соответствии с некой системой оппозиций, разные комбинации которых задают разные элементы ряда, также как фонологическая система языка строит модели звуков – фонемы в соответствии с системой оппозиций присущей именно данному языку.
Во-вторых, необходимо отделить «типические» проблемные ситуации, закономерно возникающие в социуме с определённой организацией, в результате поведения особей, регулярно и направленно ищущих соответствующих контактов друг с другом, от случайных или вынужденных взаимодействий, возникающих нерегулярно в силу действия внешней причины, а не благодаря соответствующему направлению социальной активности самих особей («социальное притяжение»).
В третьих, те адекватные стратегии поведения для следующего периода взаимодействия, которые выбраны особью в ответ на сигнал и сами являются сигналами, кодирующими начатый процесс общения и доводящими его до осмысленного результата, следует отличать от просто экспрессивных реакций на внешний стимул или изменения внутреннего состояния. Отличия состоят в том, что первые обладают типологически определённой формой, дифференцированной от формы других сигналов и от «фона» в виде континуума несигнальных реакций участников взаимодействия, а вторые – нет, хотя столь же «ярки» и демонстративны.
Соответственно, главная проблема этологического анализа коммуникации – как воспроизводимо и точно осуществить это «разделение» во всех трёх случаях.
2. Инфантицид – дезорганизация, а не адаптация
Социобиологическая идея эволюции альтруизма посредством отбора родичей требует, чтобы близкие родственники всё время находились рядом, «под рукой», в пространстве индивидуальной активности более старших и более сильных (более высокоранговых) индивидов, которые, как предполагается, окажут помощь. Легче всего этого достичь, затруднив расселение потомства – вокруг размножающейся пары основателей возникают поселения из её потомков, и детей потомков, с некоторыми вкраплениями прибившихся «чужих особей». такая колония может закладываться, скажем, также группой братьев или сестёр, как сообщество шимпанзе или прайд львов. Если в такой разросшейся семье размножение молодняка / полувзрослых особей подавляется (они могут лишь занимать «вакансии» взрослых птиц по мере их выбытия), и они в охраняют общую территорию и выкармливают потомство старших возрастов, получаем коммунальные ячейки разной организации, которые типологически сходный с семьёй общественных насекомых.
Правда, постоянное присутствие родственников другого пола в сфере досягаемости создаёт опасность инбридинга, особенно у млекопитающих. Соответственно (объясняют социобиологи) надо расселение одного пола стимулировать, а другого задержать, тогда получаются плюсы родственного отбора без минусов. И действительно, у млекопитающих обычно расселяются самцы, остаются самки (у видов, где расселяются оба пола, самцы в среднем диспергируют активней, дольше и дальше), у птиц, наоборот.
Но есть и исключения: у части социальных видов млекопитающих в период полового созревания покидают группу самки, а самцы остаются в ней, в то время как у большинства социальных видов с развитым «альтруистическим» поведением – наоборот). Предложена следующая эмпирическая закономерность, удовлетворительно объясняющая это явление. У видов, у которых длительность пребывания в группе одного самца меньше, чем возраст самки к моменту первого зачатия, самки остаются в группе (поскольку менее «скороспелы», чем самцы). У видов, у которых самец и его ближайшие родственники находятся в группе дольше, чем их возможные дочери делаются половозрелыми и могут зачать потомство, из группы уходят самки. Естественно, это объяснение связано с идеей избегания инбридинга и сохранения всех выгод от отбора родичей[1].
Правда, более детальный анализ, показывает, что эти расселительные различия как-то связаны с правилом Холдейна: в обоих случаях расселительная активность больше у гетерогаметного пола и больше у гомогаметного, а от типа социальной организации (есть эусоциальность с репродуктивным разделением труда или нет) это в принципе не зависит. Среди прочего определяет бОльшую уязвимость гетерогаметного пола по сравнению со гомогаметным по правилу Геодакяна.
И если мы сравним близкие виды одного рода или семейства, у части которых развивается коммунальное гнездование с помощничеством или иные типы социальной организации с существенной альтруистической компонентой [вроде поведения сторожей у сурикат или взаимопомощь у вампиров, когда сытые особи делятся с голодными высосанной кровью], а у части нет, то мы увидим, что разница в дисперсии самцов и самок у первых не выше, чем у вторых.
Более того, межвидовое сравнение чётко показывает, что определённая степень различий в дисперсии самцов и самок сформировалась ещё при образовании соответствующего надтаксона (скажем, рода, включающего сравниваемые виды, или семейства) вне связи с эусоциальностью, возникшей позже в некоторых филогенетических ветвях, и её появление несильно модифицирует этот параметр. И даже если модифицирует, то здесь эволюция «шьёт из готового», а не создаёт новые характеристики популяционной системы.
Но даже в этом случае плюсы, связанные с возможностью развития альтруизма в группе размножающихся родственников одного пола [развитие общей системы оповещения об опасности, как наземных беличьих, лемуров, и некоторых обезьян] полностью нивелируется ущербом от инфантицида, уничтожения детёнышей соседних особей, когда они забегают на чужие участки, вмешиваются во взаимодействия и вообще путаются под ногами. Чужих детёнышей атакуют при попытках выпрашивания или при попытках вмешаться во взаимодействия взрослых особей, для безопасного участия в которых с минимальным риском ранения и гибели нужно компетентно участвовать в обмене агонистическими или брачными демонстрациями, а они у детёнышей, конечно, не развиты. Риск ранения и гибели особенно велик
— когда детёныш достаточно подрос, чтобы в силу высокой подвижности вероятность ухода к соседям по колонии была достаточно высока,
— когда он в силу взрослости уже достаточно мотивирован, чтоб вмешаться в территориальные или брачные взаимодействия у соседей (или во взаимодействия своей матери с пришлым самцом), но специфические демонстрации ещё не развиты настолько, чтобы вмешательство перестало быть опасным для жизни.
Поэтому, рассчитывая выигрыш от задержанного расселения родственников и их размножения в зоне досягаемости друг для друга, следует из потенциального выигрыша от распространения альтруизма следует вычесть потери от детоубийства, которое происходит уже сейчас. Это как с невыгодностью жизни в долг — берёшь чужие деньги и на время, отдавать приходится свои и навсегда. Оказывается, у всех соответствующих видов взрослые особи убивают детёнышей близких родственников. Так, у черноногих луговых собачек Cynomys ludovicianus этим усердно занимаются лактирующие самки.
Поселение луговых собачек включает несколько группировок, каждая состоит из 3-4 взрослых самок – близких родственниц, взрослого самца их детёнышей, и годовалых молодых от прошлых помётов. В период вскармливания каждая самка занимает отдельную нору, где проводит большую часть времени. Детоубийства совершаются, когда самка ненадолго покидает нору.
Родственницы самки, которые обычно оказываются её ближайшими соседками и имеют детёнышей того же возраста (а родственность самок между собой в значительной степени гарантирует сходные сроки протекания разных стадий репродуктивного периода вот по этим причинам) забираются в её нору и убивают одного или нескольких детёнышей. Как правило, убитые частично или полностью съедаются «мародёркой». В наблюдениях 1981-1984 гг. зафиксировано 40 случаев эффективного детоубийства, 19% всех выводков уничтожено полностью, 11% частично. Более того, тактика мародёрства для индивида оказывается выгодной: самки, истребившие чужие выводки, лучше выращивают собственное потомство. У них выше доля детёнышей, оставшихся живыми к моменту отлучения, их выводки больше, детёныши крупнее и сами «мародёрки» в период вскармливания меньше теряют в весе, чем самки-недетоубийцы. Все остальные типы детоубийства (чужими самцами, годовиками и пр.) у луговых собачек носят случайный характер[2].
Легко видеть, что данное поведение полезно для особей, практикующих инфантицид, и крайне вредно для распространения «эгоистических генов» альтруизма, если бы таковые существовали и определяли наличие высокоразвитой социальности у луговых собачек, поскольку избирательно истребляются близкие родственники – потомство матерей, сестёр, теток по материнской линии и т.п.
Дело в том, что когда самка уходит мародёрствовать, её собственное потомство также подвергается риску быть съеденным, 5 из 70 самок, наблюдаемых J.L.Hoogland, потеряли потомство именно в этой ситуации. Поэтому она не уходит далеко и охотится в ближайшей окрестности от норы, где селятся именно близкие родственники. Чужие животные детоубийства не совершают, так как все члены этой элементарной колонии дружно объединяются против «чужака». Обзор показывает, что такого рода инфантицид присущ всем социальным видам млекопитающих, где близкородственные особи одного пола поселяются группами совместно и тесно взаимодействуют друг с другом[3].
Тем не менее социальность у этого вида развилась до очень высокого уровня по сравнению с другими родами наземных беличьих, что ещё раз доказывает, что не эгоистичный ген, а эгоистичный индивид является действительной единицей отбора. Эгоистичный ген – лишь тень индивида, проекция множества селективно-ценных признаков на множество потенциально детерминирующих их задатков, которая возникает из генетического детерминизма социобиологов (последний сам по себе ошибка, тогда как идея «эгоистичного гена» может быть полезной фикцией в целом ряде случаев, к которым г.Докинз умело привлекает внимание).
Впрочем, в коммунальных группировках обычны ещё и убийства детёнышей неродственных особей. Скажем, в прайдах львов или крупных приматов (гамадрилы, гульманы, гелады, шимпанзе) смена самцов в группировке ведёт к тому, что они убивают детёнышей, рождённых самками от прошлых самцов – часто но не всегда, и далеко не всех детёнышей (и не всяких). Исходя из панселекционизма и идеи «эгоистического гена», социобиологи легко придумали этому объяснение – мол новые самцы стремятся максимизировать собственный успех независимо от «интересов вида» и убивают детёнышей других отцов, чтобы самки могли вновь беременеть уже от них. Тем самым они увеличивают распространение своих генов и уменьшают распространение генов других самцов, что должно поддерживаться отбором.
Опять же такие объединения, как прайд львов, стадо гамадрилов или гульманов, представляют собой сообщества самок, достаточно родственных между собой (или несколько подобных сообществ), которые захватываются группой самцов (одиночным самцом), также родственных между собой и могут оспариваться другими, пришлыми самцами.
Действительно, на первый взгляд всё логично. У гамадрилов Papio hamadryas группа из трёх взрослых самцов была введена в стадо гамадрилов и гибридов гамадрил х анубис, где взрослых самцов не было. Все самцы нападали на маленьких детёнышей, с матерями которых они формировали гаремы. У всех самок, у которых по разным причинам не было циклики, после появления взрослых самцов циклика восстановилась, причём у лакирующих самок она восстанавливалась обычно после убийства детёнышей самцом[4]. У шимпанзе к подобному детоубийству могут присоединяться другие особи: когда молодой взрослый шимпанзе в парке Махале убил трёхмесячного детёныша у самки, которая перешла из другой группы и в период беременности и родов держалась на «нейтральной зоне» между ними, к нему присоединились ещё 4 взрослые самца и самка. Убийство закончилось поеданием жертвы[5].
Однако, более тщательный анализ показывает, что и гамадрилы, и гульманы, и гелады, и львы убивают только тех полувзрослых детёнышей, которые вмешиваются во взаимодействие самцов с обретёнными самками. До этого вмешательства самец-гамадрил не собирался нападать, и относился к детёнышу дружелюбно, а интенсивность преследований, от которых в конце концов гибнут детёныши, прямо пропорционально неспособности прекратить вмешательство, оценить опасность ситуации, перестать в неё попадать и т.п. В результате детёныши, родившиеся вскоре после помещения в группу чужих самцов, «усыновляются» ими, у них развиваются вполне дружественные отношения с самцами, которые им, конечно же, не отцы. Точно также уцелевают достаточно взрослые детёныши, которые в ответ на агрессию самцов принимали позы подчинения, и, соответственно, не вмешивались во взаимодействия с их матерью (Gomendo, Colmenares, 1989). То же самое было обнаружено в случаях детоубийства у гульманов[6], гелад, львов и т.д.
То есть выживают детёныши именно тех возрастных категорий, сохранение которых максимально препятствует эффективному распространению генов новых самцов через спаривание с самками группы. Слишком маленькие детёныши требуют заботы, грудного вскармливания, и снижают репродуктивную активность самки, слишком большие быстро вырастают и становятся конкурентами самца.
Ещё более детальный анализ сообществ приматов показывает, что от инфантицида больше всего погибает детёнышей у молодых самок, неловкость которых в ситуации мотивационного конфликта как бы подставляет детёнышей под агрессию самца. Наиболее склонны к детоубийству введённые в группу молодые самцы с не вполне ритуализованным поведением (а не старые, спариваниям которых детёныши мешают гораздо менее). В случае неустоявшихся ранговых отношений в группе такой же эффект в виде всплеска инфантицида наблюдается при введении в группу новых самок, а не самцов, и этому адаптивный эффект приписать уже решительно невозможно[7].
То есть подобное поведение – это типичная дезорганизация, а не адаптация, что и понятно: сложную систему вроде сообщества приматов или львиный прайд нельзя адаптировать по всем измерениям сразу, и увеличение адаптивности по одному параметру будет оплачиваться дезорганизацией по остальным.
Далее, кроме инфантицида от тесного взаимодействия близких родственников происходит такой явный минус, как месть. Наблюдения за группой японских макаков в Римском зоопарке показали, что многие атаки взрослых обезьян на своих сородичей или их детёнышей происходят в ответ на нападение на их собственных детёнышей. Они представляют собой «месть», жёсткость которой пропорциональна спровоцировавшей её агрессии. В течение часа, следующего за нападением родственника на взрослого макака или его детёныша, животное избирательно атакует именно детёныша противника (частота атак на эту обезьянку была значимо выше статистически ожидаемой при случайном выборе). Атака проходила как правило, в отсутствие родителей. «Месть» не наблюдали в группах, составленных из разных родительских линий[8].
Наконец, для самых разных видов показано, что в условиях самой жёсткой необходимости избежать инбридинга или правильно адресовать альтруизм нужным особей никогда не формируется способность распознавать родичей как таковых. В выборе кому помогать и с кем спариваться животные руководствуются не родством, а более простыми правилами, которые при всякой социальной дезорганизации ведут или к избыточному инбридингу, или к чрезмерному аутбридингу в группировке.
Так, в ходе 5-летних наблюдений за индивидуально распознаваемыми группировками одичавших лошадей-мустангов с известной генеалогией зафиксировано 244 копуляции, завершившиеся рождением 92 жеребят. Лошади живут постоянными группами, где спаривания самок контролируются единственным гаремным жеребцом. Однако его контроль не идеале – даже если кобылы не покидают группы, они нередко спариваются с пришлыми холостяками. Эмиграция молодых кобылиц и жеребчиков происходит с наступлением их половозрелости, в 13-50 мес. у первых (среднее 2,2±0,8 лет), в 11-52 мес. у вторых (в среднем 2,3±0,9 лет).
В момент наступления половозрелости самка может оказаться в группе, удерживаемой неродственным ей гаремным жеребцом. В этом случае он не спаривается с ней, также как и с собственными дочерьми. В то же время гаремный жеребец не сильно препятствует спариваниям своих кобылиц – будь то его дочери или неродственные особи – с пришлыми холостяками, препятствует только уводу. Таким образом, избегание инбридинга здесь если и имеет место, то не за счёт опознания родичей, а лишь за счёт гораздо более простого правила – отказа от спаривания с лично хорошо знакомыми молодыми самками[9].
В то же время для распространения альтруизма через накопление «генов альтруизма» и отбор родичей нужно точное определение последних и чёткое отличение родственников от неродственников со сходным поведением, связанным, скажем, с исполнением той же социальной роли, в которой обычно оказываются родственники, осуществляющие акт помощи (социальные отцы и биологические отцы и пр.).
Так что развитая социальность и альтруизм возникают не из «генетического эгоизма», а из социальной дезорганизации, связанной с переуплотнением. Переуплотнение, кроме дезорганизации, сильно усиливает дезорганизации системы, увеличивает вероятность появления более сложных, более «громоздских» и долгосрочных отношений, с большей кооперативностью поведения особей, которые при меньшей плотностью просто не могли бы появиться и стать объектом оценки отбором. Порядок возникает из хаоса через усиление флюктуаций, всё как обычно, и эгоизм индивидов здесь материал и бензин, а не мотор и руль.
3. Игра вето против дилеммы узника
Создав идеальную модель «эгоистических индивидов», в том числе для решения проблемы эволюции альтруизма, социобиологи, естественно, должны решить, в каких случаях эгоистичные индивиды будут кооперироваться, несмотря на свою эгоистичность, и почему естественный отбор в эволюционных масштабах времени будет эти условия накапливать, а не рассеивать. Для этой цели используется другая идеализация, названная дилеммой узника (prisoner’s dilemma): узники, запертые в камерах, или кооперируются, или отказываются, и от этого зависит, получает ли каждый своё наказание, или они в вместе могут скинуть весь срок, или один подставляет другого. То есть игра симметрична, оба игрока имеют одинаковые стратегические права, решения основаны только на информации, полученной в сходных взаимодействиях с тем же игроком в прошлом.
В изящных компьютерных имитациях Анатоля Раппопорта и теориях Роберта Аксельрода (см. докинзов «Эгоистичный ген») показано, что если выбор «узников» «кооперироваться» или «отказываться» целиком стохастичен и модифицируется лишь потерями или выигрышами прошлых решений, то «узники» начинают кооперироваться, если предстоящее им число итераций неизвестно.
Но оставим диллему узника социобиологам, для зоолога проблема в том, что она совершенно абиологична!
«Узники» полностью изолированные друг от друга, не пытающиеся прокоммуницировать, обменяться сигналами, чтобы выбрать собственную модель поведения по результатам коммуникации, а не «вслепую» — чушь и нонсенс. Реальные, живые позвоночные (да и беспозвоночные, хотя тут не уверен) прежде чем принять решение о кооперации/отказе, обязательно коммуницируют, и вступают и прочные социальные связи со всеми теми особями, с которыми возможна кооперация или отказ. Того рода связи, пусть мимолётные, возникают даже в анонимной стае, где малознакомые особи кормятся рядом друг с другом; тут связи слабее, но скоромней и возможности кооперации.
Сама возможность выбора — кооперация или отказ? в случае «узников», изолированных особей просто не существует, она следствие социальной связи, и коммуникации, поддерживающей эту связь. И вот тут уже выбор, кооперироваться или отказываться, животное делает не случайно, а направленно, на основании той модели поведения партнёра, которая у него «сложилась» по результатам прошлой коммуникации (фактически знание о Другом, особенно если речь идёт о высших позвоночных с развитой социальностью — врановых, ткачиковых, хищных, копытных, приматов).
То есть диллема узника плоха, поскольку игнорирует такую реальность социальной жизни животных в сообществах, социальная организация, в которой обязательны коммуникация и кооперация. Их результатами поддерживается социальная струткура группировки со следующей из неё поведенческих ролей, известная всем членам сообщества, благодаря чему поведение каждого члена делается высоко предсказуемым для остальных. Далее, ещё одна абиологичность диллемы узника — в фиксированном вознаграждении (или плате) для разных вариантов решений.
Понятно, что такого в природе просто не бывает, выигрыши и потери есть функция состояния среды, которое постоянно меняется, на семь жирных дней приходится семь тощих, и наоборот, так что в зависимости от средовой динамики выигрыши от кооперирования или отказа будут «плавать» и в среднем сильно пересекаться. Конечно, в таких условиях никакого прогресса в эволюции кооперирования «узников» не будет, что легко показать той же имитацией. А вот выбор кооперирования или отказа естественным способом, через социальную связь и коммуникацию (в этом случае кооперироваться значит создать альянс совместно действующих индивидов, направляющих условия к общей цели и преодолевающих сопротивление оппонентов и конкурентов, отказ от кооперирования значит действие в одиночку) в условиях нестабильности среды имеет тот явный плюс, что даже если искомый выигрыш не будет получен, то в результате социального взаимодействия представления животного о своих социальных партнёрах будут сильно уточнены.
И в следующий раз, когда встанет аналогичный выбор, животное сможет собрать куда лучшую «коалицию», из индивидов, способных действовать вместе с большим успехом, точней определит необходимый срок создания таких коалиций (если затягивать время её существования, она становится источником вражды и т.п.). А этот прогресс в «самопознании» особей, возможностей действия индивида в рамках разных социальных ролей, естественным образом развивает способность к кооперации (то бишь толкает вперёд ту самую «эволюцию альтруизма») не меньше, а больше, чем собственно выигрыш от кооперативного выбора
В чём здесь выигрыш? Без объединения в коалицию вероятность доступа к самке поодиночке у всех её членов 0 или близко к тому. После объединения она возрастает пропорционально силе и авторитету участников: у лидера, скажем, до 0,7, у его соратников до 0,2-0,3. При этом у них оказывается «блокирующий пакет»: если лидер «не делится доступом» к самкам, совместно отбитым у доминантам, или властью, к которой поднялся благодаря коалиции, миноритарии подрывают положение лидера просто выходя из неё.
Из этого исходит альтернативная дилемме узника модель вето, развитая Ronald Noё[10] (1989) на примере образования самцовых коалиций у павианов бабуинов Papio cynocephalus – вида, в сообществах которого доминирование и подчинение максимально выражены, а доминант наиболее ограничивает поведение подчинённых (по крайней мере в сравнении с другими видами обезьян).
Модель вето основана на коалиционных играх, где сделки не только возможны, но и эффективны, а права игроков принципиально неравны, как это и наблюдается у кооперирующихся обезьян. Соответственно, основная проблема слабого животного, нуждающегося в союзнике, состоит не в контроле за поведением партнёра, а выборе наилучшего партнёра из имеющегося лимитированного набора по результатам прежней коммуникации (каждый завершившийся акт которой уточняет и совершенствует представления о других игроках, имеющиеся у данной особи). Основная проблема сильных участников коалиции состоит в том, чтобы коалиция не распалась, то есть чтобы слабые получали большую часть выигрыша, чем та, на которую они могут рассчитывать самостоятельно, ведь длительная эксплуатация соответствующего ресурса возможна только в составе коалиции (Noё, 1989).
В таком случае понятно, что использование диллемы узника для доказательства невозможности коммунизма (как случая предельно широкой кооперации с людьми вообще, а не только с ближними, знакомыми и полезными) представляет собой попытку с негодными средствами.
Модель проверяли на случаях образования коалиций взрослых самцов бабуинов, которые образуются, чтобы «пробить» её членам доступ к эстральным самкам или к позиции доминанта. Такой ресурс исключает использование диллемы узника, так как в каждый момент неделим, и может достаться только одной особи, но в отличие от выигрышей в диллеме узника, он существует длительно во времени, то есть захватив доступ, коалиция может распределить эксплуатацию ресурса согласно «коэффициенту авторитетности» или «трудового участия» своих членов. Собственно, все социальные ресурсы такие.
Средняя группа бабуинов составляет 40-70 особей, из них 5-10 взрослых самцов. Обычно объединяются низко- и среднеранговые самцы против высокоранговых. Применение к ней модели вето предполагает что животные «договариваются о сотрудничестве» в рамках соответствующего социального взаимодействия, а необходимая информация об «уровне личной годности» партнёров передаётся самими актами социального взаимодействия. Действительно, когда партнёры активно ищут друг друга для образования коалиции, а не изолируются друг от друга, как «узники», состязательность между ними автоматически переходит в сделку, накопление опыта успешных «сделок» с данными компаньонами создаёт между ними эмоциональную привязанность уже вполне «бескорыстного» характера, что хорошо известно исследователям социальных сетей, при высоком уровне взаимной привязанности так что коалиция начинает действовать как нечто целое и не распадается на атомизированных индивидов даже в случае, если дела их не задались и т.п. (Noё, 1989).
Действительно, система брачных связей у бабуинов на первый взгляд кажется промискуитетом, а доминант выглядит настолько неотразимо и мощно, что кажется пользующимся неоспоримым преимуществом в доступе к самкам. Полевые исследования групп бабуинов в саваннах около Найроби, однако, показывают, что самки совсем не являются пассивным объектом для удовлетворения желаний самого сильного самца.
Самки активно предпочитают одних самцов другим, ограничивают спаривание одних самцов и способствуют приближению других, причём ведущим фактором выбора оказываются совсем не отношения доминирования между самцами, а связи коалиционного характера, такие как «дружба» и «брачное супружество». Причины таких предпочтений выявлены наблюдениями в период, когда самки нерецептивны. При перемещениях и на отдыхе разные самки группы держатся около кого-нибудь из самцов. В таких диадах очень высок процент взаимных чисток и других дружелюбных контактов, во время конфликтов самец поддерживает самку (-ок) и выступает на её стороне. Эти отношения были обозначены как «дружба».
С каждым самцом обычно «дружат» 8-9 самок: самцы обычно неродственны друг другу, а вот самки его свиты часто (хотя не всегда) оказываются близкими родственницами. Доминирующие самцы имеют не больше «подруг», чем подчинённые, но вот старые самцы независимо от ранга имеют «подруг» больше, чем молодые. Детёныши самок-«подруг» держатся рядом и контактируют с тем же самцом, с которым «дружат» их матери, во время спаривания самки в среднем предпочитают «друзей» другим самцам, хотя на их выбор влияет много факторов (Smuts, 1987[11]).
То есть заводить себе друзей выгодно самкам, а кооперироваться с самцами, популярными у самок, выгодно уже для самцов.
Дальше в тех же кенийских саваннах исследовали факторы, влияющие на репродуктивный успех самца бабуина. У взрослых самок бабуина набухание половой кожи, соответствующие фолликулярной стадии овариального цикла, длится 15-20 дней, спаривания происходят в последнюю неделю этого периода (то есть декада на сближение партнёров), вероятность оплодотворения максимальна в последние 3 дня. Хотя в среднем каждая самка спаривается с многими самцами, и каждый самец, с многими самками, в стаде существует чёткая тенденция к образованию временных брачных связей. Последние состоят в том, что в течение определённого времени от нескольких дней до нескольких часов партнёры монополизируют друг друга: держатся вместе и спариваются только между собой, и даже доминант не может нарушить это соединение. Благодаря этому примерно в 1/3 всех наблюдений рецептивного периода >60% спариваний совершал один самец – временный супруг, а остальное время делилось между его партнёрами по коалиции.
Тактика монополизации времени самок коалицией самцов по сравнению с гипотетически возможной альтернативой «силового» доступа к самкам доминирующих самцов скажем в те последние 3 дня. Частота спариваний самцов с рецептивной самкой примерно одинакова во все дни периода, а периоды, когда самке удалось забеременеть, и «пустые» периоды, не различаются между собой по параметрам поведения самцов или самок.
Репродуктивные успехи самцов могут быть описаны совокупностью 5 характеристик, которые сильно коррелируют друг с другом: число эйякуляций, число эйякуляций в период наибольшей вероятности зачатия, число и длительность пребывания во временной брачной связи, степень монополизации времени самок, на которых самец обращает внимание. Эти признаки связаны не столько с доминантным статусом самца в группировке, сколько с умением выбрать успешную коалицию в данный конкретный сезон (Berkovitch, 1987[12]).
Кроме бабуинов, вообще у многих приматов, между самцом и самкой устанавливаются отношения, которые обозначают как «половое супружество», в котором самка – не добыча сильного, а участник взаимовыгодных отношений с дружественным ей самцом. «Супружеские» отношения создают привязанность, и которые оба могут прервать, а третий не может нарушить, пока они существуют. В колонии макаков-резусов на о.Кайо-Сантьяго «супругами» считали особей, которые держатся вместе и спариваются только друг с другом, если это условие не выполнялось, связь считали разорванной, если через некоторое время партнёры снова были вместе, то это расценивалось как новое супружество.
За репродуктивный период время супружества длилось от 1 до 11 дней, в среднем 1,75. Общее число супружеств за сезон и продолжительность супружества одного самца позитивно коррелировали с его рангом. А вот связь между доминантными рангами партнёров и инициативой поддержания супружества отсутствовала, то есть возможность монополизации самки или самца зависит не от их собственных качеств, а определяется «эффективностью политики», проводимой каждым из них в отношении других особей в группе.
В 60% случаев инициатором супружества был самец, связи поддерживаемые самцами, были продолжительней, чем связи, поддерживаемые самками. Животное, инициирующее супружество, чаще чистило своего партнёра, чем тот его, что показывает подчинённое положение и зависимость от выбора другой особи. 41% всех супружеств образуются за неделю перед зачатием, 28% — в период зачатия и 30% после. В первом случае преобладают «супружества» с высокоранговыми самцами, во втором – с самцами из коалиций, в третьем – чёткой зависимости от ранга не найдено.
Так что животным в сообществах выгодно уметь оценивать друг друга и кооперироваться в меру эффективности этой оценки именно потому, что «единица вздор, единица – ноль» хорошо подобранная коалиция преодолевает силу любого доминанта и, в свою очередь, преодолевается коалицией, подобранной ещё лучше и в нужный момент. Естественно, это обеспечивает прогрессивную эволюцию кооперирования.
4. Синицы «не сачкуют» при выкармливании птенцов
У певчих птиц, дятлов и других групп с птенцовым типом развития, где выкармливают оба родителя последние часто конкурируют друг с другом за возможность кормить птенцов. Иногда дело доходит даже до стычек в случае совместного появления самца и самки у гнезда. Особенно это заметно у большого пёстрого дятла, где самки немного крупнее и много агрессивнее самцов. Последние часто боятся показываться у летка вместе с самкой или, в ответ на её агрессию, бывают вынуждены съесть корм и улететь за новой порцией, так и не отдав птенцам предыдущую. У певчих птиц это наблюдается тоже, только там чаще доминирует самец, а не самка (также как у белоспинного и трёхпалого дятлов).
Понятно, что развитые конкурентные отношения между родителями дают превосходную возможность проверить главнейшую исходную предпосылку социобиологии: что индивиды остаются «эгоистическими животными» всегда, даже находясь в социальной связи друг с другом (в данном случае в паре, но это могут быть отношения доминирования-подчинения, или отношения территориальных соседей, охраняющих каждый собственное пространство и старающихся расширить его за счёт других и т.д.). Соответственно, что бы ни происходило, индивиды выбирают такую схему поведения из нескольких возможных, которая увеличивает их выигрыш и уменьшает затраты, независимо от успеха и неудачи «общего дела», в данном случае выкармливания птенцов.

С точки зрения социобиологии, того самого «общего дела», в данном случае выкармливания птенцов, просто не существует, это эпифеномен отношений, удовлетворяющих «эгоизму» каждого из родителей и поэтому устойчивых (представляющих собой эволюционно-стабильную стратегию, ЭСС). С точки зрения оппонентов социобиологии, это самое «общее дело» и определяет структуру отношений родителей друг к другу, также как устойчивое воспроизводство видоспецифического паттерна социальных отношений в популяции определяет те демонстрации, которые животные адресуют друг другу в конкретных специфических ситуациях взаимодействия (чтобы разрешить их в свою пользу, но в рамках «типических отношений» системы, а не так, как выгодней с точки зрения «эгоистического расчёта», о котором твердят социобиологи).
Проверить, как это на самом деле, достаточно просто. Нужно в половине гнёзд в момент прилёта одного из родителей (скажем, самки) искусственно интенсифицировать её выкармливая, озвучивая её прилёт искусственной подачей криков выпрашивания птенцов, в дополнение к их собственной вокализации. А в другой половине гнёзд то же самое сделать в момент прилёта самцов, а не самок; вне прилётов, когда ни одного из родителей нет у гнезда, не делать ничего.
Эта озвучка интенсифицирует поведение выпрашивания одного из родителей и, если социобиологические представления верны, нагрузка на второго родителя уменьшится и он в меру уменьшения решит сачкануть. Если же неверны, то интенсификация поведения выкармливания одного из родителей побудит и второго больше вкладываться в «общее дело», хотя непосредственно он стимуляции не подвергался.
Соответствующие опыты были проведены с большими синицами в Великобритании; результаты изложены в работе С.А.Hinde (2006). Дополнительная стимуляция криками выпрашивания птенцов при прилёте реально действовала на фокальную особь, увеличивая число визитов с кормом к гнезду в час по сравнению с контролем (рис.2, чёрные столбики). И что самое важное, воздействие на одного родителя вело к изменению поведения второго в том же направлении (т.е. в сторону увеличения частоты визитов с провизией), хотя и в несколько меньшей степени (рис.2, штрихованные столбики). Эффект «сопряжения родительских усилий» был сопоставим в случаях, когда стимулировали самку или самца из пары.
Забавно, что искусственное усиление вокализации в моменты прилёта самок или, соответственно, самцов, интенсифицировало частоту подачи криков выпрашивания у самих птенцов, но только в эти моменты. Устойчивого подъёма интенсивности выпрашивания это не вызывало, и разницы в интенсивности вокализации между контролем и прилётом второго родителя найдено не было. Не менялись также поза кричащего птенца, структура серии криков выпрашивания и размер приносимой жертвы, даже у того родителя, который подвергался обработке (табл.3).
Видимо, долговременно-устойчивый уровень интенсивность вокализации птенцов пока зависит только от их собственных характеристик — уровня пищевой мотивации, двигательной активности и массы соответствующего птенца, то есть пока они маленькие, они асоциальные, и не находятся в социальной связи с родителями, а только выпрашивают и принимают пищу от них, в отличие от млекопитающих.
Следовательно, особей, связанных социальными связями в сообщество, уже нельзя рассматривать как «эгоистических индивидов», каждый из которых независимо оптимизирует своё собственное поведение и свой бюджет, пусть даже и за счёт партнёра или «общего дела» (биологически осмысленного результата взаимодействий, поддерживающих эту социальную связь). Вопреки представлениям социобиологов, особи, находящиеся в социальной связи, реагируют как нечто целое, и оптимизируют не индивидуальный бюджет, а устойчивость достижения общей цели вопреки разного рода «возмущениям среды», от плохой погоды, мешающей выкармливать, до циклических колебаний уровня мотивации, скачкообразно меняющих готовность вкладываться в выкармливание.
5. Плюсы социобиологии
Написав уже три поста с критикой социобиологии (Преодоление социобиологии (часть 1), Преодоление социобиологии: инфантицид как дезорганизация, а не адаптация (Часть 2) и Преодоление социобиологии: игра вето против диллемы узника (Часть 3)) я, как честный человек, должен рассказать, в чём несомненные плюсы этого направления. В чём социобиологи как продвинулись дальше, чем классические этологи, которых они критиковали и которых вытеснили? или социоэтологи, с которыми они конкурировали? Что нового о поведении животных мы смогли узнать только благодаря социобиологии, поскольку структура социобиологической теории предполагала возможность подобных явлений, а другие направления отрицали их или закрывали на это глаза?
Впрочем, все плюсы социобиологии имеют чисто научное значение, и будут малоинтересны тем, кто поддерживает социобиологию из идеологических соображений — как «научное обоснование» господствующего эгоизма, попытку «доказать», что сконцентрированность на себе любимом – не душевный дефект, а нечто прогрессивное и полезное. В самом деле, если из взаимодействия миллионов частных эгоизмов родится большая общественная польза, как доказывают популяризаторы социобиологии, эгоистом быть не стыдно, а как бы даже почётно, и понятно, кто и почему эти доказательства принимает на ура. Но бог с ними, я про научные достижения.
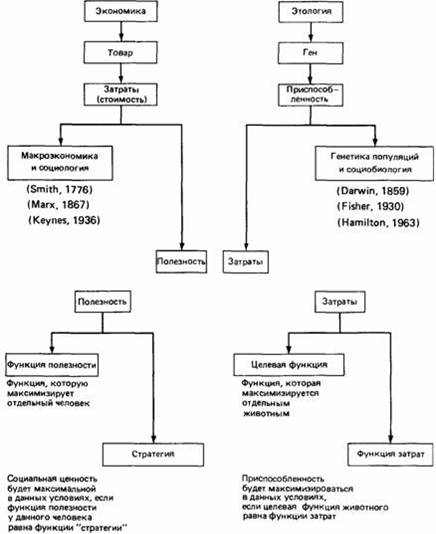
Параллель между понятиями экономики и этологии. Вверху. Общая схема, на которой указаны фамилии исследователей, внесших наибольший вклад в развитие этих областей науки, и даты публикации их основных работ. Внизу. Детализированная схема в приложении к отдельному человеку или животному. (McFarland, Houston, 1981.).
Благодаря социобиологам было строго показано, что животные в социальном общении – это именно «эгоистические индивиды», калькулирующие соотношение платы и выигрыша, всегда и везде поступающие так, чтобы это соотношение выросло за счёт противника, как вода течёт вверх, а не вниз. Это не добрые самаритяне, совместными усилиями реализующие видовой инстинкт, от которого польза всем (такими их представляли классические этологи). [«Калькулирующие» здесь, понятное дело, означает, что уже филогенетические предшественники этих форм эволюционно изменились так, чтобы при всяком действии животного в его типичной социальной среда «калькуляция» получалась автоматически].
Далее, было показано, что у всех «правильных» социальных отношений – территориальность, колониальность, ток, коммунальное гнездование – есть своя «оборотная сторона», образованная «неконвенциональным поведением» тех же особей, вроде рейдов резидентов, самцов и самок по чужим территориям, без территориальных претензий, но для внебрачных копуляций. Все процессы коммуникации, поддерживающие перечисленные отношения, помимо «конвенционального поведения», соответствующего «правилам игры» в качестве другой возможности всегда предполагают «обман», когда особи пытаются тот же результат «схватить» без организованного обмена демонстрациями и связанного с этим риска.
Соответственно, исполнение животными специфических элементов процесса взаимодействия – видовых демонстраций – может очень сильно варьировать по шаблонности/стереотипности, от точного воспроизведения «идеальных образцов» формы соответствующего сигнала до крайне «расшатанного» и «пластичного», когда сигнал крайне затруднительно распознать, а телодвижения особи больше напоминают прямые движения спаривания или агрессии, чем соответствующие демонстрации. Соответственно, «эгоистические индивиды», вступая в коммуникацию друг с другом, из этого разнообразия уровней стереотипности исполнения демонстраций выбирают тот, который выгоден и доступен именно им – а потом, по достижении результата взаимодействия, оказывается понятно, выиграла особь, совершив этот выбор, или будет расплачиваться за него.
Иными словами, благодаря социобиологическим концепциям, альтернативным концепциям классических этологов, мы не только узнали больше о социальной жизни конкретных видов, но внимание исследователей было привлечено к тому, что раньше считалось несущественным – «оборотная сторона социальности», «обман», неожиданная вариативность исполнения демонстраций. Это поставило новые вопросы – а как, собственно, сохраняется видовая норма социальной организации (в поддержание которой эгоистические индивиды делают инвестиции, которые оправдываются далеко не всегда), почему она на не деградирует в условиях обмана, по крайней мере, на длинных масштабах времени, если во многих случаях «обманывать» достаточно выгодно, по крайней мере, здесь и сейчас?
Почему видовые демонстрации остаются самими собой, сохраняют дифференцированные формы и нужную стереотипность исполнения, почему «честно сигнализирующих» особей не вытесняют обманщики, идущие на нарушение «правил игры», или заменяющие демонстрации пластичным действием? Ведь их поведение результативно, а рискуют они существенно меньше, они не должны накапливать заведомый избыток ресурсов, ведь коммуникация «по правилам» — это своего рода «предмет роскоши».
Далее, видовые демонстрации и социальная организация вида, которая поддерживается организованным обменом демонстраций в конкретных взаимодействиях особей – это классический пример групповой адаптации – приспособления, выгодного сообществу в целом, и невыгодного каждой отдельной особи. Особь «принуждена» инвестировать в устойчивое воспроизводство таких адаптаций, снова и снова, в каждом новом сообществе данного вида, образующегося из скопления особей, которая должна рисковать и «рвать когти» снова и снова, обеспечивая себе заведомый избыток ресурсов для успешной коммуникации и т.п. Так как же возникли подобные адаптации? почему они прогрессивно развиваются в большинстве филогенетических линий – социальная организация усложняется, территориальных или брачных сигналов становится больше, сами сигналы – дифференцированней, из стимулов они превращаются в знаки и т.п. И всё это совершенствование системы в целом идёт поперёк интересам «эгоистических индивидов» в каждой отдельной точке процесса!
Другое дело, что, верно поставив вопрос, социобиологи не могут дать на него ответ в силу внутренних противоречий концепции, которая для этого должна быть преодолена. Но хороший вопрос – это уже половина ответа.
Ещё один важный плюс – именно социобиологами было показано, что
1) ритуализированные демонстрации (и, вообще любые телодвижения и/или морфоструктуры, выступающие как сигналы), характеризуются не только эффективностью, но и риском,
3) для каждого отдельного ряда демонстраций (агонистических, брачных, умиротворения, предупреждения об опасности и пр.) риск и эффективность употребления положительно связаны, то есть ввод во взаимодействие более эффективных демонстраций связан с бОльшим риском, поэтому животные к ним переходят не сразу, а постепенно «поднимают планку» ритуализированной агрессии или ухаживания.
Правда, единственно возможного вывода из этой связи – что больший риск употребления более эффективных демонстраций есть плата за «считывание с них» большего количества информации, или более ценной информации, точная реализация которой в последующем поведении животного и есть причина большей эффективности данной демонстрации, — социобиологи так и не сделали. Ну и из этой же положительной зависимости эффективности и риска демонстрирования понятно, чем были демонстрации, до того как стать сигналами – барьером, препятствующим непосредственному сближению партнёров для ухаживания и агрессии на то время, что необходимо для того, чтобы демонстрации подействовали и будущее сближение, когда окажется возможным, было не травматичным.
6. Взаимоотношение полов в сексе и в заботе о потомстве не соответствуют социобиологическим предсказаниям
Как знает читатель, я противник социобиологических теорий, но сторонник её методов оценивания каждой из сопоставляемых стратегий или форм поведения в «единой валюте» соотношения платы и выигрыша, эффективности и риска. Почему против теорий? Потому что они превращают науку в историю счастливых догадок, как по сходному поводу удачно сказал г.kouprianov. Скажем, ещё в 9-м классе, лишь начав интересоваться поведением животных, я чисто умозрительно вывел для себя правило, «которым руководствуется половой отбор». Мол, самки производят мало гамет, но дорогих, самцы много, но дешёвых, поэтому женский пол заинтересован спариваться редко, но с лучшим партнёром, ещё лучше – сделать его постоянным, а самцы – во множестве партнёрш и непостоянных отношениях с ними.
Увы, единственный способ подсчитать, что «дешёвое», что «дорогое» — это определить сравнительные затраты самцов и самок на репродукцию, от распределения территорий и ухаживания до насиживания и выкармливания птенцов. Соответствующие цифры для разных видов птиц (с разными типами социальной организации) приведены в замечательном обзоре В.Р.Дольника (1995) «Ресурсы времени и энергии у птиц в природе» (Труды ЗИН, т.179):
Увы, они радикально несовместимы с этой гипотезой: затраты самцов на репродукцию не меньше, а то и больше затрат самок. Более того, социобиологический вариант теории полового отбора исходит из асимметричности отношений самцов и самок – один пол активно конкурирует за внимание противоположного, а другой пассивно выбирает победителя в этой борьбе. Фишка в том, что недавними работами T.Clutton-Brock показано, что у целого ряда видов конкуренция самок за внимание самцов может быть не менее жёсткой, чем аналогичная конкуренция самцов за внимание самок, что конкурентная система структурируется также и ведёт к тем же последствиям, что и в известных примерах жёсткого полового отбора у самцов, 1-2. Но, понятное дело, у одного вида интенсивный половой отбор «среди обоих полов» невозможен.
Соответственно, оптимизироваться должны энергозатраты только одного, «выигрывающего» пола. Однако приводимые Дольником данные показывают, что у всех исследованных видов затраты на репродукцию так распределяются между полами, что самцы и самки как бы компенсируют себе затраты, так что видовой бюджет приближается к наиболее экономному из возможных.
Пока самки тратят энергию на формирование крупных многожелтковых яиц и рыщут по территориям, выбирая лучшие с точки зрения прокормления не только себя, но и птенцов выводка (для самых разных певчих птиц при выборе территории показана ориентация самок на кормность летом для всего выводка, а не для самой себя весной), самцы тратят энергию на строительство гнёзд, а потом и насиживание с выкармливанием. И это не считая взаимодействий друг с другом, привлечения самок и пр. Понятно, что преимущественно самцовый вклад в гнездостроение-насиживание-выкармливание при значительных затратах на формирование яиц выгоден, что мы и наблюдаем в реальности – вопреки социобиологическим прогнозам.
Совсем не случайно повышенное участие самцов в заботе о потомстве у птиц мы видим именно в более примитивных группах каждого таксона. Среди воробьиных птиц это те, которым свойственна клаустрофилия (когда самцы тратят кучу сил, чтобы строить тёплые закрытые гнёзда, ну и много насиживают потом, и кормят птенцов), в противоположность стоящим открытые чашеобразные гнёзда. Среди ржанкообразных – всякие примитивные семейства с малым числом видов, вроде цветных бекасов. Среди птиц в целом максимум отцовской заботы на потомстве падает на более примитивные группы (бескилевые: страусы-нанду-эму-киви и тинаму), а у более продвинутых преобладает или парная, или исключительно самочья.
Я думаю, тут дело в том, что более примитивные группы в среднем хуже добывают пищу, у них затраты энергии на единицу добытого корма ниже. А репродукция — это ещё крупные дополнительные затраты, при ухудшении условий добывания корма. Соответственно, этот груз надо разделить: самец, скажем, гнездо строит и птенцов обихаживает, а самка тратится на много крупных многожелтковых яиц.
В более продвинутых группах с большей эффективностью кормодобывания это разделение не так насущно.
Второй минус социобиологических концепций связан с доверием к такого рода «счастливым догадкам» и долговременным следованием им, вместо того, чтобы рассмотреть вопрос системно. Показательна история с «пернатыми многожёнцами»: выгодны или нет внебрачные копуляции и если да, то чем?
Так, они почему-то думают, что именно те структуры или свойства поведения, на которые «упал глаз» учёного, и есть главное направление эволюции, что именно они эволюционируют, а не приспосабливаются к эволюции других структур той же организации, на которые мы в данный момент не обращаем внимания.
А ведь даже если он квалифицированный специалист, это может быть совсем не так: в самом начале изучения объекта все мы подходим к нему с обычной человеческой меркой, а она чаще всего бывает неадекватна объекту из-за радикальных различий в «напряжении жизни» у человека и животных. Поэтому «для нас» наиболее важны слова, символы, понимание и коммуникация, а «для них» — еда, хищники и паразиты; всё относящееся к коммуникации и социальности для животных – «предмет роскоши», который лишь в некоторых ситуациях бывает критическим.
Понятно, что сложную систему нельзя оптимизировать по нескольким параметрам одновременно, поэтому очень важно точно установить, какую именно структуру или характеристику непосредственно изменяет отбор, а какие изменения будут зависимыми и производными от этих ведущих: что летит из-за этого по корреляциям, и какие структуры, сопряжённые с ней, вынуждены меняться в контрапункте с происходящими изменениями ради того, чтобы не упала устойчивость воспроизводства организации индивида и популяции, иначе всё сдохнет, не выдержав эволюции.
Ведь эволюционирует не россыпь атомизированных признаков эгоистичных особей или генов, а целостная система, которую надо уметь выделить, которая «умеет» управлять элементами, особями или генами, преодолевая их «эгоистичность» через онтогенетические регуляции и связи в популяционной системе, обеспечивая кооперативное поведение элементов вкупе с управляемостью для удобства воспроизводства системного целого. Соответственно, для этого последнего характерны контринтуитивность поведения и пороговость реагирования.
Собственно, социобиологи с их позицией неограниченного эволюционизма потому раз за разом лажают в определении того, какие эволюционные изменения будут ведущими, какие – обеспечивающими, а какие вообще представляют собой энтропию процесса, социальную дезорганизацию и т.д., что «за деревьями не видят леса», за элементами – управляющей ими системы, будь то нормальный онтогенез (при эволюционных изменениях морфотипа) или популяция (вид как система популяций).
***
Красивых примеров здесь два: первый – интерпретация насильственных копуляций у речных и нырковых уток, а второй – ошибки гнездовых паразитов, разбивающих собственные яйца.
Когда утиные пары уже образовались, и самки начинают садиться на кладку, потихоньку всё более отдаляясь от самцов, те поодиночке и группами начинают преследовать чужих самок и насильственно спариваться с ними. Достаточно часто при этом присутствует «свой» самец, но даже не пытается этому помешать, причина чего довольно прозрачна исходя из «слепоты инстинкта».
«Насильственные копуляции речных уток – лучшая модель некоммуникативных взаимодействий, которые, тем не менее, входят обязательной составной частью в социальную жизнь соответствующих видов. Между самкой и самцом в паре происходят нормальные копуляции: им предшествует обычный обмен видоспецифическими демонстрациями ухаживания, опосредующими сближение партнёров, и позволяющими им на определённом этапе развития отношений координировано копулировать с частотой и эффективностью, достаточной для успешного оплодотворения (Lorenz, 1939b, 1941; Johnsgard, 1965).
Кроме того, все самцы совершают насильственные копуляции с самками соседних пар в присутствии их партнёров, которые не сопровождаются какими либо демонстрациями до, во время или после этого акта.
В вольерной группе из 8 пар чирков-свистунков Anas crecca насильственные копуляции всегда происходили с самками, уже готовыми к яйцекладке или приступившими к ней. Самое интересное, что партнёр самки, принуждаемой к копуляции, стремился держаться в непосредственной близости от неё и самца-преследователя, но нападал на последнего не раньше, чем тот начнёт садку. Эта неспособность сохраняется, хотя насильственные копуляции явно причиняют ущерб воспроизводству группы. У 7 из 8 наблюдавшихся самок свистунка зафиксировано от 2 до 15 насильственных копуляций. 2 самки погибли от травм и стресса, только 3 отложили яйца, полностью не закончила кладку ни одна (McKeeny, Stolen, 1982).
Причина неспособности «законного» самца дать отпор «насильнику» — именно в отсутствии специфических сигналов-посредников для разрешения «конфликта интересов» обоих самцов именно в этом специфическом контексте поздних этапов ухаживания, когда последнее почти завершилось, и утки переходят к яйцекладке. Если бы чужие самцы преследовали самок из пар «как обычно», брачными демонстрациями, или попытки «изнасилований» приурочивались к более ранним этапам ухаживания, скорее всего, выработались бы ответные сигналы, позволяющие самцам конкурировать за внимание таких самок.
То есть коммуникативный и некоммуникативный режим реализации социального поведения сосуществуют друг с другом у одних и тех же особей и в одних популяциях, как «лицевая» и «оборотная» сторона общего процесса коммуникации. Это заставляет признать, что не любой групповой образ жизни, не всякое «репродуктивное разделение труда», не всякая устойчивая система иерархии суть проявления социальности, но только те из них, где
— взаимодействия включают информационный обмен, «обслуживаемый» системой демонстраций-посредников между действием одних и контрдействием других индивидов,
— присутствует свобода выбора поведения на основании сигнальной информации».
То есть система взаимоотношений самцов и самок у уток (в отличие от гусей и полугусей – огарей, пеганок, магеллановых и нильских «гусей». Там самец остаётся с самкой до конца, чтобы охранять выводок и кладку от хищников, чем и мотивируется атака «насильников» как врагов) устроена так, что именно при переходе к «яйцекладке» имеет конструктивный дефект. Самцы ещё сексуально активны, но уже начали дезертировать от самок, с которыми образовали пару, благосклонности которых добились в процессе консолидации и пр., так что коммуникативный эффект демонстраций, опосредующих конкуренцию самцов за самок, уже не защищают последних от посягательств «чужака». А «силовое» воздействие, не опосредованное демонстрациями, в социальном поведении животных всегда неэффективно, в том смысле, что не даёт устойчивого эффекта: отогнать противника можно, но он будет возвращаться снова и снова, и гарантированного отсутствия вторжений на предсказуемое время вперёд можно достичь только успешно завершившимся обменом демонстрациями.
Сейчас конкурируют два объяснений «изнасилований» у уток. Первое социобиологическое: оно требует от своих приверженцев долго и тщательно искать выигрыши от «изнасилований» в виде увеличения приспособленности (ну хотя бы для самцов). Беда в том, что таковые не обнаруживаются даже под микроскопом. Вероятность оплодотворения при насильственных копуляциях ничтожна, самки от этого только травмируются психически и физически, иногда гибнут, а самцам – лишь затраты времени и энергии на очень дорогое поведение (ибо включает в себя полёт, а не только демонстрации на воде), явно не оправдывающие себя.
Второе было предложено ещё Лоренцем. Дело в том, что утиные – одна из немногих групп птиц, сохранивших пенис – гомолог копулятивных органов рептильных предков. У других птиц, не имеющих наружных половых органов, эффективность оплодотворения жёстко определяется синхронизацией действий самца и самки во время сближения перед и непосредственно во время копуляции, иначе всё дело сорвётся и усилия будут напрасны.
«Например, из 102 парящих приближений самца зеленушки только 25 окончились успешным спариванием. В 7 случаях самка даже перешла к агрессии, уже дав самцу спуститься на спину.
Адекватно реагируя на брачный сигнал, партнёры настолько синхронизируют свои двигательные реакции, что в определённый момент их успешные копуляции уже обходятся без предварительного обмена демонстрациями, а сами действия самца и самки при копуляции оказываются быстрыми, согласованными и точными (Хайнд, 1975: 394-396). Столь высокой синхронизации действий партнёра не достичь без продолжительного процесса эффективной брачной коммуникации, одним повышением уровня сексуальности обоих (а тем более только самца).
У пингвина адели Pygoscelis adeliae на о. Росса исследовали эффективность передачи спермы в процессе копуляции в успешно образовавшихся парах. Самка этого вида после копуляции некоторое время остаётся в позе спаривания с поднятым хвостом, поэтому легко увидеть, есть ли сперма в центре клоаки либо за её пределами или клоакальный контакт вообще происходил без передачи спермы (Hunter et al., 1996). Очевидно, что полностью успешным может быть только первый вариант, третий неуспешный, второй – промежуточен между ними. Далее самок отлавливали и у них брали мазки из клоаки, которые просматривали под микроскопом. За сезон в каждой паре происходило в среднем 34,4±11,9 клоакальных контакта, передача спермы зафиксирована только в 20,0±8,8. Ещё в 3,4±4,5 копуляциях сперма попадала мимо клоаки, в 4,4±4,0 копуляциях передача спермы не отмечена вовсе, в 6,6±3,5 — не удалось определить наличие или отсутствие передачи спермы. Таким образом, точность передачи спермы в акте копуляции здесь не более 58,8% (Hunter et al., 1996).
Иными словами, видоспецифические автоматизмы даже на стадии консуматорного акта работают не очень точно, поэтому всякое устойчивое увеличение синхронизации поведения партнёров на предшествующих стадиях консолидации пары, достигаемое в обмене демонстрациями, должно быть крайне существенно, а пары, более и менее успешные в спаривании – хорошо различаться по этому признаку.
Далее у пингвинов Адели исследовали эффективность переноса спермы при брачных и внебрачных копуляциях (Hunter et al., 2000). Точность помещения спермы в клоаку самки при внебрачной копуляции оказалась выше, чем при спаривании с собственным партнёром (притом, что у данного вида разные копуляции по данному параметру сильно варьируют, Hunter et al., 1996). Однако частота эякуляций при внебрачных копуляциях значительно ниже, так что вероятность оплодотворения партнёра в собственной паре в итоге оказывается выше, чем «чужой» самки. Далее, частота эякуляций самцов, вовлечённых в сорвавшиеся попытки внебрачных копуляций, не ниже частоты копуляций самцов, совершающих внебрачные копуляции успешно, то есть успех или неуспех данных актов — функция наличия/отсутствия «сбоев» поведения партнёров по ходу контакта, а не следствие различного «качества» разных самцов (Hunter et al., 2000).
Иными словами, эффективность переноса спермы при копуляциях у пингвина Адели есть следствие эффективности предшествующей брачной коммуникации (более длительной и успешной с собственным партнёром и отсутствующей при внебрачной копуляции), а не калькуляции «плат и выигрышей» индивида при выборе копуляций первого и второго типа».
Иными словами, птицам необходим некоторый уровень синхронизация прямых действий партнёров друг относительно друга. Он позволяет самцу сделать точную садку, самке – принять позу подставления и не дёргаться, не клевать самца по ходу процесса. Будет этот уровень сформирован или нет в нужные сроки, строго зависит от эффективности взаимного ухаживания партнёров в предшествующем периоде, когда они сближаются, обмениваясь брачными демонстрациями и переходят от менее эффективных сигналов репертуара ко всё более эффективным. Тогда ещё никаких копуляций нет, но успешный обмен демонстрациями, как это положено в подготовительной фазе развёртывания инстинкта, создаёт предпосылки для их успешности. Побуждает партнёров, ещё декаду назад бывших «эгоистическими индивидами», терпимо относиться друг к другу при всё более тесном сближении и не «дёргаться», реагируя агрессией или уходом, на резкие движения, связанные с демонстрациями (копуляторные действия менее резки и вычурны).
И даже несмотря на подобную подготовку, эффективность копуляций в «успешных парах» (=продемонстрировавших достаточный уровень эффективности копуляций) очень невысока – хотя всё же выше нуля. Как в примерах выше.
Иными словами, пары, превысившие некий видоспецифичный порог эффективности коммуникации в период консолидации пары, получают шанс на успешное оплодотворение, а недотянувшие до порога – шиш. Соответственно, вместо непрерывного распределения разных пар по успешности оплодотворения и, соответственно, возможностям репродукции в этом сезоне, коммуникативная система делит их на два кластера – успешных и размножающихся vs неуспешных, «пропускающих ход». Что соответствует трейд-оффу между выбором «инвестировать в размножение-рисковать» в данном сезоне и «беречься, не тратиться на размножение, не рисковать», чтобы в существенно лучшей форме размножиться в сезоне следующем, исключительно важном для эволюционной оптимизации жизненных стратегий всех позвоночных.
И этим классическая этология даёт наилучший ответ на интересный вопрос, поставленный именно социобиологами: почему у моногамных видов птиц и других позвоночных после образования пары самец и самка отнюдь не приступают к копуляциям сразу же, а ещё долго токуют друг с другом, постепенно сближаются, обмениваясь демонстрациями и т.п. К чему эти, на первый взгляд, непроизводительные затраты на стадии консолидации пары, охватывающей от нескольких дней до месяца, почему бы сразу не трахнуться?
Оказывается, это инвестиции в долговременный выигрыш: у моногамных видов образование пары само по себе гарантирует некую нечувствительность обоих партнёров к активному токованию холостых особей, обычно происходящему вокруг, укрепление устойчивости брачных связей по ходу консолидации ещё больше повышает этот порог, делая внебрачные копуляции исключением, связанным с дезорганизацией связей, а не нормой. Плюс, обмениваясь демонстрациями, оба партнёра инвестируют в долговременный выигрыш обоих, именно в «преодоление порога» эффективности копуляции, после которого можно начать яйцекладку. Хотя вроде бы каждому из участников процесса нарушение связи и измена могут быть выгодны, чем дальше они продвинулись по пути консолидации, чем устойчивей эта связь, тем сильнее канализировано развитие их поведения в следующий период, так что «дезертирование» и «обман» всё труднее и труднее осуществить.
“Устойчивость брачной пары – «продукта» эффективной коммуникации — проявляется в том, что оба партнёра отвергают ухаживание третьих особей и пара не распадается, даже если самец и самка практикуют внебрачные копуляции.
Например, у сорок Pica pica период наиболее частых копуляций самца и самки в паре, за два-три дня до откладки первого яйца — одновременно и время наиболее активных попыток внебрачных копуляций со стороны соседей. В этот период самки издают громкие крики выпрашивания и повсюду следуют за «своими» самцами, которые начинают их кормить. Защищаемая территория пары в среднем — 3 га, расстояние между соседними гнёздами 100-500 м., так что соседи имеют свободный доступ на участки друг друга (Buirton, 1983).
В период интенсивных спариваний партнёры постоянно держатся вместе на дистанции менее 5 м, перемещение партнёров по территории максимально скоординировано — самка инициирует 60% перемещений, в 94% случаев самец сразу следовал за ней. В остальных случаях инициатором был самец, самка следовала за ним в 65% случаев. То есть координация перемещений связана с сопровождением самцом самки, копуляциям с ней и одновременно пресечением внебрачных копуляций (это так называемое mate guarding).
Отмечено 92 случая, когда чужаки (всегда самцы) приближались к гнезду на расстояние менее 5 м. В 46 случаях они не взаимодействовали с самкой на гнезде, но в 44 случаях самка слетала с гнезда и вела себя по отношению к ним точно также как к собственному партнёру — выпрашивала корм, сопровождая это специфической вокализацией и демонстрациями. В 14 случаях у чужих самцов при этом зафиксированы элементы ухаживания и предкопуляции, что заставляет предполагать возможность внебрачной копуляции (Buirton, 1983).
Однако кормление самки чужим самцом в ответ на её выпрашивание не наблюдали ни разу. По-видимому, нормальное появление этого акта во взаимодействиях самца и самки требует накопления истории успешных токований в составе пары в рамках длительного развития отношений между партнёрами. Агрессивность самок к чужим самцам отмечена лишь в 6 случаях из 92, самцов — в 11 случаях из 59, когда во взаимодействиях с чужим самцом участвовал также хозяин территории «третьим» (Buirton, 1983).
Анализ реакций самок из пары на ухаживание чужих самцов показывает, что устойчивость брачных связей ограждает от внебрачной копуляции намного надёжней сопровождения партнёра и агрессии её собственного самца. Без них отношения и взаимодействия партнёров не будут скоординированы (включая координацию перемещений по территории), а без этой координации не развивается ритуал взаимного кормления, необходимый для успешной копуляции».
Уже в начале периода консолидации устойчивость брачных связей пересилила «эгоизм» индивидов (явственно видимый, когда они действуют в анонимных стаях, и не связаны социальными связями). Иначе бы пара не образовалась вообще, точнее, была бы, как у видов, спаривающихся на токах, кратковременной, нужной именно и только для копуляции. А по ходу взаимного ухаживания самца и самки в паре поведение обоих всё больше формируется требованиями, вытекающими из устойчивости брачной связи, и продиктованной ею дифференциацией поведенческих ролей, и всё меньше – «эгоистическими интересами» каждого отдельного индивида.
То же самое видим в сообществах животных, структурированных иными типами отношений, скажем, территориальностью или агрессивным доминированием: везде индивиды, вполне «эгоистические» вне сообщества и/или в анонимной стае, связываясь социальными связями в группировки с видоспецифической структурой, утрачивают любимый социобиологами «эгоизм», обретают кооперативность (синхронизованность) поведения и становятся репликами собственной социальной роли.
Вернёмся к пенису. Т.е., по версии классической этологии, в потерявших его группах птиц точность синхронизации телодвижений самца и самки критична. Поэтому надо виртуозно ухаживать, обмениваясь демонстрациями, пока самка должным образом не примет позу подставления, а самец не сделает точную садку (что ему затруднительно из-за сильного конфликта мотиваций, половая в нём борется с сильным страхом перед самкой, а обмен демонстрациями в т.ч. разрешает конфликт). А в случае уток, у которых пенис есть, мол, получится и насильственно.
Но увы – у бескилевых птиц пенис наличествует, а насильственных копуляций нет. Почему? Социальная система радикально иная, забота о кладке и потомстве переложена на самцов, самки лишь путешествуют между их участками и несут им яйца. Предпосылок к насильственным копуляциям не создаётся, ибо структура социальных отношений не создаёт исходных возможностей для развития этого поведения – самцы слишком заняты заботой о потомстве. И наоборот – у видов без пениса, но гнездящихся в плотных колониях, с обычной для них социальной дезорганизацией (тонкоклювая кайра, белолобая щурка, балтиморский цветной трупиал) насильственные копуляции присутствуют – и тоже менее успешны, чем обычные, когда самка не сопротивляется самцу, или даже привлекает его, принимая позу подставления. У тонкоклювых кайр принудительные внебрачные копуляции были успешными (то есть состоялся контакт клоак) в 6% сучаев, ненасильственные, но тоже внепарные – в 96% (Hatchwell, 1988).
Другой пример связан с подкладыванием яйц гнездовыми паразитами (кукушками, воловьими птицами и пр.) в гнёзда видов-хозяев. Традиционный социобиологический взгляд (целиком и полностью априорного характера) — что в этой системе обязательно начинается «гонка вооружений»: хозяева изо всех сил совершенствуют распознавание своих яиц, всё лучше отличая их от подложенного, а гнездовой паразит, в свою очередь, изощряется в сближении подкладываемых яиц с хозяйскими по размеру и форме.
При этом, естественно, паразит должен уметь точно распознавать свои яйца, не хуже чем яйца хозяина. Если вдруг в гнезде одно яйцо уже есть, то большинство видов кукушек должна искать новое (второго кукушонка мелкие певчие птицы не выкормят), а воловьи птицы, у которых возможно выкармливание нескольких птенцов паразита в одном гнезде — удалить ещё одно яйцо хозяина. Эта проблема неактуальна только для видов, вроде хохлатой кукушки, подкладывающей яйца сорокам, голубым сорокам и др. видам врановых, которые доставляют пищу в избытке. Однако таких видов немного, в подавляющем большинстве виды-паразиты крупнее хозяев, и, соответственно, откладывают яйца, удаляя яйца хозяина.
Но раз за разом оказывается, что не только хозяева, но и сами гнездовые паразиты не знают облика собственных яиц и, соответственно, не могут определить, есть ли уже в облюбованном ими гнезде яйца их собственного вида или нет. Самую забавную систему такого рода образуют аргентинский пересмешник Mimus saturninus и паразитирующая на нём блестящая воловья птица Molothrus bonariensis.
Пересмешник не отличает своих яиц от яиц паразита; если они отложены, покорно насиживает, хотя самих воловьих птиц изо всех сил старается отогнать от гнезда. А они изощряются, отслеживают момент, когда насиживающая птица кормится и т.д.; вот тут настоящая «гонка вооружений». Поэтому из 347 изученных гнёзд в 89% было ≥1 яйца, в 35% — >3 и в 16% — >5. В работе Ros Cloag et al. (2011) вели видеозапись у гнёзд пересмешника, отслеживая проникновение в них воловьих птиц, а также искусственно меняли соотношение яиц пересмешника и яиц паразита, чтобы оценить выживаемость тех и других при разном количестве. Оказывается, воловьи птицы не знают своих яиц, или как минимум не отличают их от яиц вида-хозяина.
А так как, откладывая новое, они удаляют или бьют одно яйцо из уже находящихся в гнезде, они с равной вероятностью губят что яйца пересмешника, что своего вида. Соответственно, чем больше яиц воловьей птицы уже есть в гнезде, тем больше вероятность для них быть разбитыми собственным видом.
Моделирование разных стратегий хозяина («принимающая» или «отвергающая») методом Монте-Карло показывает, что в этой ситуации принимать яйца паразита выгоднее, чем их отвергать, ибо при неразборчивости паразитов приём увеличивает число мишеней для следующих посетивших это гнездо и снижает риск уничтожения для яиц
хозяина.
Поскольку пересмешник начинает кладку чуть раньше воловьих птиц, несущихся в заведомо не пустое гнездо, то выживаемость их птенцов неожиданным образом даже несколько возрастает с увеличением общего числа отложенных в гнездо яиц паразита — ибо одновременно растёт риск самоуничтожения. Эта тенденция особо заметна начиная с 3-х отложенных яиц воловьей птицы и далее растёт к 5-6. Надо отметить, что в отличие от кукушек и медоуказчиков вылупившийся птенец воловьей птицы не губит других птенцов, но они тем не менее сильно объедают хозяина.
К слову, одно из неожиданных открытий, связанных с применением ДНК-фингерпринта в области исследования социальных отношений у птиц, касается как раз гнездовых паразитов. До эры использования молекулярных методов в оценке родства и индивидуальной идентификации особей все считали их сугубо промискуитетными (погинно-полиандрическими) видами, вроде желудёвых дятлов или страусов нанду.
А тут оказывается, что это строгие моногамы, живущие парой на постоянной территории, где они последовательно выявляют все гнезда потенциальных хозяев, и ежедневно контролируют их, следя, на какой стадии репродукции какое находится, чтобы подложить яйцо в самый удобный момент, — когда кладка уже сформирована и плотное насиживание только-только началось. Иными словами, кукушки с воловьими птицами отнюдь не освобождают себя от заботы о потомстве; они просто меняют форму последней. Если считать по энергозатратам, то расходы энергии пары на отслеживание всех гнёзд и «впаривание» по яйцу в каждое, с обманом родителей (что члены кукушечьей пары часто производят совместно, координируя действия друг с другом) сравнимы с таковыми хозяйского вида.
Фактически оказывается, что всякий реально возможный ущерб от паразита (при том соотношении численности обоих видов, которая устанавливается в данном местообитаний) слишком незначителен по сравнению с энергозатратами на формирование яиц и сбор корма для птенцов, особенно при периодических ухудшениях погоды. Поэтому главное направление адаптации связано с совершенствованием поведения самообеспечения особей (кормление, самосохранение, безопасный отдых и пр.), а не социальных взаимоотношений между ними и не поведения, связанного с взаимодействием с другими видами.
Выигрыш от совершенствования того и другого невелик по сравнению с главной проблемой и статьёй расходов, а усилий и затрат требует массу (тем более что всякое периодическое развёртывание дополнительной специфических видов социальной активности или активности, отслеживающей и пресекающей поползновения гнездовых паразитов). Опять же, запуск социальных взаимодействий, социальных демонстраций или взаимодействий с другими видами вреден тем, что постоянно прерывает кормление, не позволяя реализовывать самые специализированные методы кормодобывания, требующие много времени на подготовительные действия.
А они как раз самые эффективные! Отсюда птицы, увеличивающие интенсивность социальных взаимодействий с особями своего вида или с биоценотическими партнёрами в у раз, обрекают себя на снижение эффективности кормодобывания в уn раз.
Следовательно, социальная коммуникация у птиц и других позвоночных – «предмет роскоши»: она не запускается без наличия на индивидуальном участке заведомого избытка корма (или безопасности), превышающего некий предел. Поэтому на выборке моногамных видов птиц, обладающих разной социальной организацией, было показано, что ухудшение погодных условий и большая непредсказуемость изменений погоды в месте гнездования уже образовавшихся пар увеличивает частоту внебрачных копуляций их участников.
Это верно как для разных географических популяций одного вида, так и для разных социально моногамных видов, далёких друг от друга таксономически. Оно и понятно, чем хуже погода и менее предсказуем её ход (здесь надо учесть, что как минимум у моногамных воробьиных при образовании пары весной значима кормность участка для выводка летом и родительские качества самца, которые проявятся тоже только летом), тем трудней паре в период образования и консолидации обеспечить тот «минимально необходимый уровень избыточности» корма, безопасности и других витальных ресурсов, который необходим для запуска эффективной коммуникации с необходимой частотой и интенсивностью взаимодействий партнёров.
Следовательно, устойчивость брачных связей будет ниже, а партнёры – куда более благосклонны к ухаживаниям токующих третьих особей, чем сороки в примере выше. Т.е. наблюдаемая частота внебрачных копуляций — это не специфическая адаптация к нестабильному климату, как тщатся показать авторы, а пассивное следствие социальной дезорганизации, как рост заболеваемости гепатитом А – следствие плохого водопровода. Ведь в условиях плохой погоды участие во внебрачных копуляциях снижает приспособленность, а не повышает её по сравнению с «более верными» парами, ибо представляет собой неэффективное «распыление сил» (также как в случае с болезнью, изученном Е.А.Литвиновой с соавт.).
То же верно и для отслеживания гнездовых паразитов, распознаванию их яиц и пр.
Аналогичным образом весь XX век орнитологи считали, что разнообразие окрасок птичьих яиц связано с маскировкой от хищников, и лишь сейчас набираются доводы, что окраска фона и плотность пятен – это скорее специфические светофильтры, поддерживающие устойчивость стимуляции зародыша, особенно в момент, когда самка открыто гнездящихся птиц сходит с кладки. Чем извлекается из забвения соответствующая теория конца XIX века, оно и понятно: равномерно устойчивый прогрев и оптимизация микроклимата существенны для всех сделанных кладок, которые стали насиживать, а защита от хищника – лишь в том сильно меньшем % гнёзд, наличие которых заподозрят хищники данной местности и станут искать. При сколько-нибудь реальной численности хищников и эффективности их поиска первый фактор много больше влияет на выживаемость гнёзд и яиц, чем второй.
Отсюда помимо неадекватности собственно теоретических интерпретаций социобиологов, у исследователей этого направления практически нет шансов понять, что форма поведения или морфоструктура, на которую «первой упал их взгляд» — не главное направление эволюционных изменений, а обеспечивающие/подстраховочные преобразования организации, а то и просто пассивное следствие происходящего. Они упорно громоздят одно ah hoc предположение на другое, лишь бы спасти исходный тезис, не подтверждающийся уже 30 лет (случай адаптивности внебрачных копуляций, что «неверные самки ищут лучшие гены» или «неверные самцы их распространяют»). Плюс структуры или формы поведения, оставляющие впечатление «самых значимых» в начале исследований, когда мы знаем биологию вида крайне поверхностно, почти всегда оказываются зависимыми и второстепенными, когда мы её изучим как следует.
7. Про «избегание инбридинга» и логику системы
В социобиологической концепции эволюции альтруизма путём родственного отбора есть неустранимое внутреннее противоречие (и не одно…). С одной стороны, чтобы альтруист мог при случае оказывать помощь родственникам, последние должны быть всегда «под рукой» — рядом, в пределах досягаемости, и часто действовать вместе, например, при поиске корма, перемещении в составе группы или спасении от хищника. То есть, чем более выражен альтруизм в пользу родственников, тем выше вероятность того, что сообщества – локальные популяционные группировки, в которых и происходит воспроизводство – будут образованы особями, сильно более родственными между собой, чем в среднем по популяции. А это, в свою очередь, противоречит другим построениям социобиологов, не менее важным, чем идея родственного отбора – о том, что животные «избегают инбридинга». Точней, что вредные – в эволюционном масштабе времени – последствия близкородственного скрещивания настолько велики и сильны, что животные при прочих равных будут избегать инбридинга, даже если это потребует дополнительных перемещений и большей «разборчивости выбора», всегда увеличивающих риск.
Так социобиологи загоняют себя в ловушку – с одной стороны, животные должны предпочитать близких родственников ради развития альтруизма и социальности, с другой стороны – избегать их ради снижения вредных последствий инбридинга, которые сводят на нет всю выгоду от альтруизма в пользу родичей.

Вроде бы получается патовая ситуация, но противоречие это мнимое. Оно основано на ложной посылке, что субъектом выбора при предпочтении или, наоборот, избегании родственников является изолированный индивид, «эгоистически просчитывающий» свои «платы» и «выигрыши» и «принимающий решение» о лучшей стратегии поведения. Или точней, уже отшлифованный для автоматических выборов нужных стратегий отбором. Я уже писал о механизмах саморегуляции в популяционных системах, сортирующих особей с разными потенциями между группировками разной плотности, так что потенциальные носители «нужных» стратегий попадают в «нужную» социальную среду. Эти же механизмы саморегуляции системы, перераспределяя индивидов между группировками, определяют уровень инбридинга или аутбридинга (скрещивания предпочтительно неродственных особей) в конкретных локальных сообществах.
Это хорошо видно на примере сравнения степени инбридинга в популяционных группировках, находящихся внутри сплошного ареала, и включённых в общую «сеть» обмена нерезидентными особями, и в изолированных поселениях, находящихся вне пределов ареала, за 500-1000 км от какой-либо из его границ. Степень инбридинга в изолированных группировках всегда исключительно высока (обычно максимум из того что можно обеспечить данным составом резидентных особей и их потомков), что не снижает ни успех размножения особей, ни жизнеспособности группировки.

Более того, что большинстве случаев в таких группировках в период миграций в массе появляются нерезидентные особи, остаются, и пытаются закрепиться, образовать пару и начать размножение (поскольку здесь территория видом недонаселена, есть избыток пространства, корма и др. ресурсов). Резиденты ж упорно предпочитают образовывать пары между собой, игнорируя всякий инбридинг, и также упорно отвергают вселенцев.
Например, в популяции желны Dryocopus martius на п-ове Ютландия в 50-70 км севернее границы основного ареала, в течение нескольких поколений зафиксировано постоянное предпочтение собственных потомков в качестве половых партнёров (варианты инбридинга «мать х сын», «отец х дочь», и никогда «брат» х «сестра»). Ежегодно в период позднелетней дисперсии птицы из основной части ареала наполняют поселение, многие из них держатся здесь всю зиму, но местные особи отвергают их как брачных партнёров (Christensen, 1995[13]).
И наоборот: если какие-то поселения включены в систему направленного обмена особями внутри популяции, то в них поддерживается неродственное скрещивание даже при условиях, которые в соответствии с социобиологическими представлениями о кин-отборе должны бы превращать все популяционные группировки в объединения заведомых родственников. Однако наблюдается прямо противоположное. Например, у манакина Chiroxiphia linearis и Ch.caudata зафиксировано репродуктивное разделение труда между самцами на току – токуют все, сплочённо и синхронизовано, а вот спаривается только один альфа-самец, все прочие ему ассистируют, препятствуя чужим или молодым особям своими вторжениями сорвать сближение главного самца с самкой. По социобиологическим представлениям такой странный отказ от собственного размножения в пользу других особей, как и в случае общественных насекомых, возможен лишь под действием родственного отбора, а тока, совпадающие у этого вида с элементарными популяционными группировками, должны представлять собой группы родственников.
Естественный вопрос – в чём выигрыш «побеждённых» самцов-участников процесса коммуникации? Во-первых, в устойчивом сохранении их резидентного статуса на токовом дереве. В силу его наибольшей привлекательности для самок демонстрирование здесь сопряжено с минимальным риском, в отличие от обычного образа жизни самцов без брачного наряда, перемещающихся между локальными аренами. Во-вторых, коммуникация предполагает использование некой системы сигналов-посредников и процесс обмена сигналами по определённым «правилам». Наличие «общих правил» в процессе коммуникации оставляет шанс побеждённому победить в следующем взаимодействии, более эффективно использовав эти «правила». У него этот «шанс» в принципе тот же, что у победителя, если только поражение не снизило устойчивость пребывания особи в данном сообществе (ток, группировка и пр.), структурированном и поддерживаемом именно процессом коммуникации.
В этом случае система «гарантирует» долговременный выигрыш, «обеспеченный» адекватным предъявлением нужных демонстраций в нужные моменты взаимодействия, вопреки широкому ассортименту «помех», как оплата наличными подтверждает надёжность сомнительного клиента. При надёжных «гарантиях» долговременного выигрыша, связанных с устойчивым течением специфических процессов коммуникации, индивиды во взаимодействиях соблюдают специфические «правила игры», даже ведущие к их отказу от немедленного размножения. Например, экзотическая форма «репродуктивного разделения труда» между доминантом в центре и субдоминантными особями на периферии обнаружена у манакинов р.Chiroxiphia, также у Pipra fascicauda. Демонстрации всех самцов на току высоко синхронны и довольно чётко согласованы во времени и в пространстве. Но у альфа-особи они ведут к спариванию, у бета-особей вокруг – только привлекают самок на ток как целое (Robbins, 1986).
В этой системе пребывание на току и синхронизация брачной активности доминантами ничего не даёт b-особям, кроме ожидания долговременного выигрыша в виде постепенного перемещения «на позиции» альфа-особей по мере их естественной убыли. На таком току исчезает разделение на индивидуальные территории самцов: во всех трёх названных и неродственных группах подчинённые демонстрируют на одной территории с доминантом.
Например. у Chiroxiphia linearis самцы демонстрируют на диффузных токах: на участке наблюдений в 80 га в 1983-1986 гг. держалось около 50-60 самцов, из которых лишь 6-8 занимали социальный ранг a. Лишь они участвовали в копуляциях, прочие самцы – «ассистирующие» b-особи. У самцов отсутствует территориальное поведение, агрессия доминантов по отношению к подчинённым зафиксирована лишь в моменты взаимодействий с самкой. У видов с развитой территориальностью на токах происходит нечто обратное – массовые вторжения соседей, в том числе и низкоранговых особей, в момент сближения самого доминанта с самкой. Между a- и b-самцом возникает стабильное объединение: птицы вместе поют, вместе привлекают рецептивных самок в центр активности, вместе демонстрируют перед самкой, хотя спаривается только a-самец. Доминант придерживается одного центра активности, низкоранговые самцы могут иметь до 6, с расстоянием между ними до 800 м. Здесь они «ассистируют» разным a-самцам, рассчитывая занять «вакансию» после естественного выбытия (McDonald, 1989).
Подчеркну: долговременный выигрыш – единственное приемлемое объяснение пользы подчинённым самцам от кооперирования с доминантом. Спекулятивная идея «отбора родичей» не проходит вовсе. Анализ полиморфных микросателлитных локусов в 33 парах «ассистирующих» b-самцов с a на току Chiroxiphia linearis показал, что средний коэффициент родства отрицательный с 95% доверительным интервалом. Следовательно, все участники «репродуктивного разделения труда» менее родственны друг другу, чем особи случайной выборки из той же популяции. Реципрокный альтруизм также отсутствовал. Тщательный анализ выгод бета-особей показа, что все они были отсроченными, в том числе восхождение к альфа-статусу, редкие копуляции, жёсткая связь самок с определённым токовищем (McDonalds, Potts, 1994).
Поскольку птицы собирающиеся на току, менее родственны друг другу, чем случайная выборка из популяции, то система «следит» за пресечением непотизма и оказывает разнообразящее воздействие на генетический состав локальных группировок, не допускает группирования особей сходного происхождения в одном месте. То же обнаружено для токов тетерева: их тоже социобиологически пытались истолковать как скопления более или менее близких родственников – но получили прямо противоположный результат (Foster, 1981[14]; McDonald, 1989; McDonald, Potts, 1994[15]).
То же самое видим и в человеческих популяциях, в отношении которых социобиологии много пролили чернил, описывая универсальность «избегания инбридинга» и всеобщую эволюционную обусловленность запрета на инцест. Но нет, то и другое возникает исторически, и связано с чисто экономическими причинами. И как только те же экономические причины делают близкородственное скрещивание выгодным, антропологи фиксируют его широкое распространение, не встречающее ни внутренних преград, ни культурных запретов. В первую очередь речь идёт о горных изолированных поселениях на Памире, в Гиндукуше и пр., с их нехваткой земли, так что делиться семьям невыгодно, в сочетании с экстремальными природными условиями и редкостью сети поселений, так что брачная экспедиция сложна и опасна.
При крайне ограниченном количестве земли, используемой для земледелия, близкородственные браки в традиционном обществе делаются предпочтительными, так препятствуют дальнейшему дроблению земельных участков. В этом случае коэффициент инбридинга F поднимается выше 5% (уровень, превышение которого заставляет считать близкородственные скрещивания не редкой аномалией, а распространённым явлением). В частности, F очень высок у памирцев сёл Пастхуф и Хуф (0,0594 и 0,0545), история которых прямо подтверждает влияние попыток воспрепятствовать парцеллярному дроблению участков как причины распространения близкородственных скрещиваний. Тот же фактор действует и в другом изоляте, у киргизов с.Мургаб, где коэффициент инбридинга пока низок но, судя по наличию той же побудительной причины и повышенной жизнеспособности потомства от близкородственных скрещиваний, развитие идёт в ту же сторону, просто Мургаб находится пока в начале этого пути.
Традиции нерегулируемой рождаемости у памирских горцев и других общин, практикующих близкородственные скрещивания в разных районах земного шара (в табл. 57 Н.Х.Спицына перечисляет 13 сообществ с F>0.05 не считая тех, которые изучала она сама) сами по себе способствуют поддержанию высокого уровня инбридинга, поскольку чем выше рождаемость внутри изолята, тем выше вероятность кровнородственного скрещивания, вносящего свой вклад в коэффициент инбридинга.
Как это неудивительно, в изолированных им полуизолированных популяциях высокий коэффициент инбридинга далеко не всегда только отрицательно влияет на репродукцию – есть и мощные положительные моменты. Точно также как высокий коэффициент аутбридинга в крупных городах – скрещивания более неродственных особей, чем в случайной выборке из популяции – имеет не только положительные следствия в виде гетерозиса и акселерации, но и возможный отрицательный эффект, связанный с разрушением локально-коадаптированных генных комплексов при скрещивании в крупных городах представителей территориальных групп одного этноса. Например, русских из степной зоны и поморов в Москве[16].
Для корректности во всех памирских изолятах сравнивалась репродукция в близкородственных и неблизкородственных, которые уже вышли из репродуктивного возраста. В Пастхуфе в родственных брачных парах выявилось большее число рождений и живых детей, приходящихся в среднем на одну женщину, меньшее число мертворождений и выкидышей, а также отсутствие случаев бездетности в сравнении с теми же параметрами в неродственных браках. Однако достоверно значимы лишь различия по мертворождениям, выкидышам, и уровню бесплодия. В Хуфе по всем исследуемым признакам «более благополучно» выглядят неродственные брачные пары, хотя различия статистически незначимы. Напротив, в Мургабе, только начинающем эволюцию в сторону распространения близкородственных скрещиваний, отмечено достоверно большее число рождений и живых детей на 1 женщину в близкородственном браке, чем в неродственном[17].
Н.Х.Спицына связывает эти положительные эффекты инбридинга с повышенной биологической совместимостью гамет у родственных партнёров, связанной с действием системы генов протеиназ, влияющих на легочную и печёночную деятельность.
Другим примером популяций с исторически сложившейся системой браков, включающей предпочтение браков между родственниками, являются азербайджанцы. В 1986 г. было изучено население с.Абдал Агдамского района Азербайджана. Выявлено 109 близкородственных браков, что составляет 25% от общего числа. Преобладают браки между двоюродными сибсами – 68,8%, доля троюродных и четвероюродных 17,4% и 13,7%. Полностью отсутствуют одноперекрёстные браки типа дядя-племянница, тётя-племянник, F=0.0642, то есть выше чем в памирских изолятах. При этом реальный уровень инбридинга может быть ещё выше, из-за неточностей применённого генеалогического метода оценки родства и многочисленных отказов респондентов сообщить характер брака.
По всем сравниваемым критериям: среднее число рождений живых детей, частоте мертворождений и выкидышей – кровно-родственные браки в Абдале имеют несколько большее значение, чем неродственные. Однако в родственных браках повышен темп репродукции: женщины в родственных браках, завершившие период репродукции, показывают более высокую плодовитость (7,86±0,74 детей на женщину и 6,75±0,48, живых детей к моменту выхода родителей из репродукции в родственных браках 6,27±0,001 в неродственных 5,46±0,43, при том что во вторых мертворождений выкидышей и прочих «сбоев» существенно больше.
Эти данные Н.Х.Спицыной показывают, конечно, не то, что близкородственные скрещивания полезны или, наоборот, вредны, а нечто более существенное и важное. Биология каждого отдельного индивида настолько пластична и богата возможностями для изменений, совместимых с жизнеспособностью и даже увеличивающих её, что практически любое необходимое изменение в системе не встретит препятствий в виде биологии особей, скорей наоборот, в биологических свойствах индивидов – малых и заменимых элементов популяционного и социального целого – всегда найдутся такие генретические, физиологические, поведенческие характеристики, которые смогут это изменение на уровне целого поддержать и даже выиграть от подобной поддержки.
Это в равной мере касается изменений на уровне популяционного целого у животных – постоянные преобразования «кружева ареала» и пространственно-этологической структуры группировок под влиянием изменений среды, колебаний численности и т.п., и изменений в человеческом обществе, связанных с экономическими интересами, классовой борьбой, общественным прогрессом и пр.
При наличии выраженного желания людей изменять нормы общежития в эту сторону, преодолевая консерватизм нежелающих биология никогда не является ни непреодолимым препятствием изменений, ни обязательным побудительным стимулом. Хотя может быть попутным ветром, который надо уметь ловить в паруса. Поскольку, как писал по сходному поводу А.А.Ухтомский:
«Мы – не наблюдатели, а участники бытия. Наше поведение – труд… Природа наша делаема[18]».
Примечания:
[1]Clutton-Brock T.H., 1989. Female transfer and inbreeding avoidance in social mammals// Nature. Vol.337. №6202. Р.70-72.
[2]Hoogland J.L., 1985. Infanticide in the prairie dogs: lactating females kill offspring of close kin// Science. Vol.230. №4729. Р.1037-1040.
[3] P.H.Harvey и Lyles A.M., 1985. The limits to infanticide// Nature. Vol.318. №6043. Р.235-236.
[4]Gomendio M., Colmenares F., 1989. Infant killing and infant adoption following the introduction of new males to an all-female colony of baboons// Ethology. Vol.80. №1-4. Р.223-244.
[5]Nishida T., Kawanaka K., 1985. Within groop cannibalism by adult male chimpanzees// Primates. Vol.26. №3. Р.274-284.
[6] Agoramoorthy G., Mohnot S. M., 1988. Mohnot Infanticide and juvenilicide in Hanuman langurs (Presbytis entellus) around Jodhpur, India// Human Evolution. Vol.3. P.279-286.
[7]Чалян В.Г., Мейшвили Н.В., 1990. Инфантицид у павианов гамадрилов// Биол. н. №3. С.99-106.
[8]Aureli F., Cozzolino R., Cordishi C., Scuccht S., 1988. Interfamili revenge in a group of Japanese monkeys// Boll. Zool. Suppl. 55 pp.
[9]Berger J., Cunningham C., 1987. Influence of familiarity on frequency on inbreeding in wild horses// Evolution (USA). Vol.41. №1. Р.229-231.
[10] Noё R., 1989. The veto game played by baboons: a challenge to the use of prisoner’s as a paradigm for the reciprocity and cooperation// Anim. Behav. Vol.39. №1. Р.78-90.
[11] Smuts, 1987. Why are friends for// Natur. Hist. Vol.96. №2. Р.38-44.
[12] Berkovitch F., 1987. Reproductive success in male savanna baboons// Behav. Ecol. Sociobiol. Vol.21. №3. Р.163-172.
[13] Christensen H., 1995. Bestandsentwicklung und Verwandschaftsbeziehungen in einer kleinen Population von Schwarzspechten (Dryocopus martius) in deutch-dänischen Grezraum // Corax.16: 196-197.
[14] Foster M., 1981. Cooperative behaviour and social organization of the shallow-tailed manakin (Chiroxiphia caudata)// Behav. Ecol. and Sociobiol. Vol.9. №3. Р.167-177.
[15] McDonald D., 1989. Cooperation under sexual selection: age-graded changes in a lekking bird// Amer. Natur. Vol.134. №5. Р.709-730.
McDonald D., Potts W., 1994. Cooperative display and relatedness among males in a laked-mating bird// Science. Vol.266. №5187. Р.1030-1032.
[16]Ю.П.Алтухов. Генетика природных популяций. М.: Академкнига, 2003. С.231-238. См. подробнее «Естественный отбор и фенотипическая норма в популяциях человека»
[17] Н.Х.Спицына. Демографический переход в России. М.: Наука, 2006. С.147-153.
[18] См.также интересное обсуждение этой темы в комментах
Рекомендуем прочесть
 Как Европа открыла для себя Московскую Русь, а потом закрыла
Как Европа открыла для себя Московскую Русь, а потом закрыла COVID-19 вскрывает правду
COVID-19 вскрывает правду «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве «Жесты», вόроны и шалашники
«Жесты», вόроны и шалашники Синдром темпа жизни (pace-of-life syndrome, POLS): эволюция концепции
Синдром темпа жизни (pace-of-life syndrome, POLS): эволюция концепции Малоизвестная победа короля Фейсала над коммунизмом и СССР
Малоизвестная победа короля Фейсала над коммунизмом и СССР Социальная сегрегация в городах
Социальная сегрегация в городах От экокризиса к экоустойчивости: три принципиальных момента
От экокризиса к экоустойчивости: три принципиальных момента

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}