

В продолжение темы парфорсной теории эволюции как механизма быстрой адаптации
В.В.Суслов
Аннотация. Рассмотрены противоречия эволюции при освоении особями новизны. Показана неадекватность оптимизационных сценариев. Предложен неоптимизационный сценарий, базирующийся на свойствах стресса.
Мозг решает проблему адаптации при недостатке актуальной информации и времени путем целенаправленного поведения (ЦП). Появление мотивационного возбуждения вызывает отрицательную эмоцию. Появление препятствий усиливает ее, а не мотивацию. Сравнивая лабильно зависимый от ситуации неспецифический[1] эмоциональный фон с устойчивой специфической мотивацией, особь оценивает вероятность достижения цели еще до действий, in situ, и очерчивает круг и иерархию мобилизации средств (компенсаторная и подкрепляющая роль эмоций), либо меняет цель коррекцией акцептора результата под обстановочную афферентацию (замещающая роль эмоций), либо терпит цель как невыполнимую (переключающая роль эмоций). Т.е. эмоция не только оценивает, но и повышает вероятность успеха, канализуя перебор средств памятью (рис.1), причем рост вероятности достижения результата вызывает положительную эмоцию [тез.I-68].
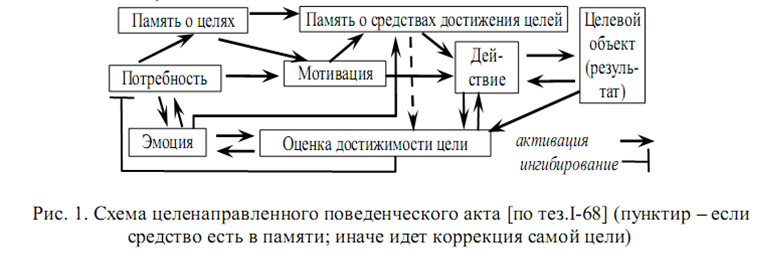
Считается: в эволюции нет целеполагания из-за постериорности отбора и стохастики мутабильности μ. Но несоответствие стадий освоения новизны (инвазия в новые экониши/биотопы, адаптация к ним) и потенциальной скорости эволюции заставляет в этом усомниться.
Столкнувшись с засухой, небольшая популяция травяных лягушек не только освоила сухие биотопы, но и конкурентно оттеснила аборигенов – морфофизиологически более адаптированных жаб. Начав с элиминации поведения “сушебоязни” отбор за несколько поколений сформировал ряд параллельных жабам адаптаций. В свою очередь, у жаб комплекс адаптаций к суше не возникал один раз у общего предка, а формировался у разных таксонов параллельно, одновременно с расселением по континентам [тез.I-III].
Популяции городских сизарей, годами хранившие наследованную от диких предков неспособность сидеть на ветвях, быстро (~5-30 лет) и независимо освоили ветви. Если в Иркутске ясна кормовая адаптация (после высадки яблонь Палласа в 60 гг. голуби подбирали опавшие плоды, часто – за стаями воробьиных, в зимы 1970 гг. отмечены голуби, питающиеся на ветвях, в 1980-1990 гг. поведение рутинно и утеряло связь с кормом [3, Фефелов, личн. сообщ.]), то на Кавказе инвазию на ветви запустили между 1980–1985гг. снова проникшие в регион кольчатые горлицы[2].
После кормежки на земле рядом с голубями редкие горлицы летели на деревья. Часть голубей подражала (стадный инстинкт развит у голубей), большинство после двух-трех попыток летели на привычные присады, но некоторые голуби с трудом продолжали попытки. К 1990 гг. поведение уже не редкость. В предковом ареале оба вида хорошо различают друг друга.
Резкий переход от замедления к ускорению инвазии (i) и несоответствие успеха инвазии и адаптации к новой эконише[3] (ii) дополнит несоответствие скоростей эволюции и адаптации (iii). Палеоданные подтверждают современные: при инвазии эволюция ускоряется, охватывая малые популяции (МП). Чем больше генов под отбором, тем больше особей он бракует за поколение (Дилемма Холдейна, ДХ). Но особи гибнут и случайно. Репродуктивный резерв при прочих равных условиях тем меньший, чем меньше популяция, должен компенсировать смертность от всех причин. Геномные проекты [тез.I-III] и проект ENCODE [7] показали: популяции насыщены полиморфизмами, псевдогены и некодирующая ДНК имеют ненулевую вероятность синтеза. Большая часть этой изменчивости нейтральна, но оставшаяся грозит размыть уже достигнутые адаптации, что ставит на первое место их охрану, а не поиск редких полезных аллелей. Отсюда, МП опасны рост и μ, и нормы реакции (НР), и случайной гибели (γ). Отсюда, опасна инвазия в новизну и даже приближение к пессимальному краю предковой экониши до формирования эвадаптивных преадаптаций. МП лучше специализироваться, что ведет к катастрофе эндемизма (КЭ): от ухода в область предсказуемости/комфорта экониши до закрепления там стабилизацией онтогенеза (путь I) и/или через ПЗА, превентивнозащитные адаптации (путь II), направленные не на противодействие, а на недопущение контакта с фактором пессимальности. ПЗА, работающие все более на опережение, в итоге более устойчивы, чем защищаемая функция и в конечном итоге смогут ввергнуть в стазис даже мономорфную популяцию, для которой сценарий стазиса-контрбаланса отбора не работает.
Путь I отражен в правилах Гаузе (межвидовые отношения), контрбаланса отбора Cеверцова (внутривидовые отношения), раздела адаптивных субниш Креславского (борьба с абиогенными факторами): все три правила – о разделе предковой ниши, а не инвазии в новую [тез.I-III].
Путь II, запрещая накопление новизны, эффективно обеспечивает рост знаний, увеличить который можно как экстенсивно (широта кругозора, дилетантизм[4], агон [cм. тез V] и др. – IIa), так и интенсивно (педантизм, погружение в проблему за счет игнорирования всего остального – IIb). Критерий роста для IIa и IIb – рост предсказуемости, но лишь в IIa этот рост сопровождаем работой как можно большего числа и типов рецепторов, в пределе – рост перцептивной схемы за пределы умвельта. Отсюда, вопреки традиции [тез.I-68; 1], опережающее отражение и рост предсказуемости не помогают адаптации и выживаемости, а амбивалентны к ним.
ПЗА делятся на циклические (а), циклические по исчерпанию (б), наведенные (в), индуцибельные (г), неспецифические (д), конститутивные (е) и комбинированные. Пример типа а – генная сеть (ГС) циркадного ритма [8]. Тип б – ГС напоминания, чей принцип регуляции – синтез пула побочного расходуемого/самораспадающегося вещества в ходе основной работы ГС. Исчерпание этого пула вновь активирует ГС (обратная связь по исчерпанию).
Многие нейромедиаторы, месенджеры и др. сигнальные молекулы происходят от побочных продуктов метаболизма клетки [8]. ГС напоминания широко распространены: от поддержки аппетита [8] до репарации ДНК [9]. Тот факт, что ПЗА определяют базовые функции организма говорит об их глубокой эволюционной укорененности. Тип в – инстинкты, г – условные рефлексы и производная от них когнитивика [тез.I-68; 2], д – неспецифические адаптивные синдромы, нетерминализованные (боль) и терминализованные[5] (стресс). Тип е отбор может создать из любого признака[6]. Итак, из 9 типов ПЗА рост IIa дают г и д. Самый легкоформируемый и частый тип е не вызывает роста ни IIa, ни IIb.
Противоречия i-iii традиционно пытаются решить в рамках оптимизационно-балансовых сценариев (ОБС) и эффекта Болдуина. В ОБС отбор поддерживает поиск новизны, несмотря на рост γ, так как кумулятивный сбор информации о среде в итоге должен повысить ее предсказуемость [1, 13]. Эффект Болдуина [14] ускоряет эволюцию, если приобретенное ЦП и врожденная адаптация сходны. Не отрицая достигнутых успехов [1, 13, 14], отметим – в работах игнорируется КЭ, ДХ и разница IIa и IIb (копится новизна или забывается[7]?). Кроме того, ОБС требует много времени на повторные исследовательские мотивы (ИМ), когда отрецептированная картина мира сравнивается с афферентной – запомненной в ходе предыдущего ИМ. Поиск новизны в ОБС отключается при опасности [1, 13, 14], которая часто сама по себе признак новизны. Т.e. ОБС хороши при инвазии не в новую, а в филогенетически знакомую и комфортную, т.е. эфферентнодостаточную эконишу – миграция наземной птицы через океан просто не могла возникнуть в ходе серии повторных ИМ. Ее удаление от берега, равно как и удаление рыбы от кромки воды отбор быстро заблокирует, создав ПЗА.
Наконец, при инвазии в эфферентнодостаточную нишу сама эфферентная достаточность заранее детерминирует эффект Болдуина как адаптивный[8]. Итак, ИМ опасны или бесполезны при отсутствии, недостаточной разрешающей способности или медленном срабатывании рецептора или акцептора результата[9]. Отсюда, отбору проще запретить ИМ и эффект Болдуина это лишь ускорит (негативный эффект Болдуина). Действительно, в незнакомой обстановке любой неожиданный, но яркий раздражитель, успешно блокирует ИМ: в психологии стресса [тез.I-III; 18, 19] много примеров, когда бурное срабатывание ПЗА типов в (неофобия, рвота, дефекация и т.д.), г (страх) и комбинированного вг (тревожность[10] в ответ на неопасный, но незнакомый (часто – незнакомый лишь в новом контексте) фактор блокировало желание изучать не только данную, но и любую новизну[11], даже при успешном решении задачи[12].
Из биотопов, недавно бывших островными, инвазия растений – пассивных расселителей – идет эффективнее, чем животных с их неофобией и страхом [тез.I-III].
ЦП не подменяет (экранирует [14]) эволюцию, а блокирует ее, ничего не давая взамен. Решение проблемы косвенным отбором – одна адаптация повышает вероятность другой, формируя цепи или циклы (рис.2, цикл 1-5) – предложено в сценарии таких инвазий в новизну, как выход на сушу хордовых [тез.I-99] и артропод [тез.II-4]. Но шунты (рис.2, др. циклы) обесценивают сценарий: он хорошо объясняет смену органов в новой и/или филогенетически знакомой средах (пре- и постадаптации), а не смену сред.
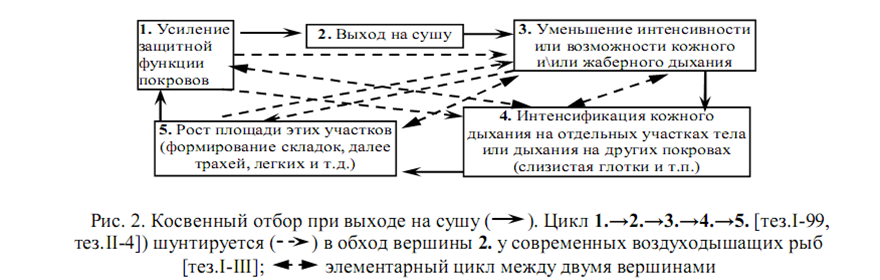
Действительно, современные рыбы с эвадаптациями к дыханию воздухом тяготеют к “стратегии кита”: питание в О2-бедном гидробиотопе↔отдых в О2-богатом гидробиотопе[13]). Зато Periophthalmus с инадаптивным кожным дыханием покидает воду систематически (противоречие ii) [тез.I-III]. Еще труднее объяснить по типу рис.2 смену умвельтов: если морфофизиологический признак используют в одной-двух ситуациях, то рецептируемая картина мира нужна во многих взаимоисключающих ситуациях, поэтому консервативна. Прозревшие слепорожденные признают важность зрения для общей оценки картины мира, но тонкую детализацию ведут привычно — тактильно [27]. Рост НР сигнализации у гибридов непевчих птиц затрудняет формирование пар [4, подробнее – «Красота и стресс…»]. Эволюцию рецепторов часто выявляют геномные проекты, а рост умвельта при этом редок[14][тез.I-III; 37].
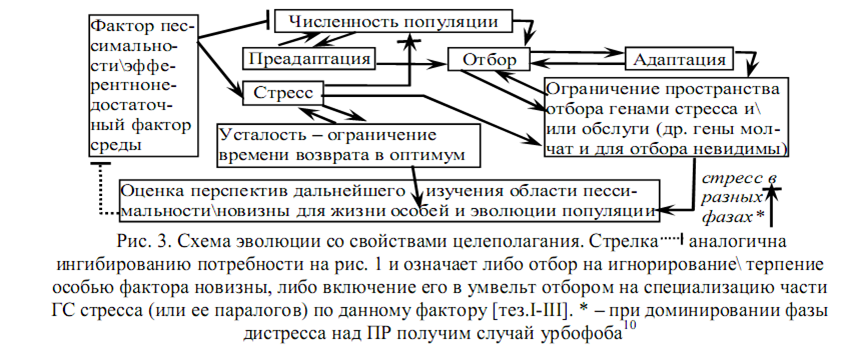
Стресс, как и эмоция, неспецифичен – кратковременно дает особи устойчивость к многим факторам (фаза перекрестной резистентности, ПР), запускаясь, угашает множество ГС (включая ряд ПЗА[15]), кроме собственной ГС и ГС базовых обслуживающих инфраструктур организма (часто это одно и то же). Долгий стресс ведет к истощению и/или гибели (фаза дистресса), но задолго до истощения провоцирует усталость. Т.о. особь in situ может оценить опасность новизны по усталости даже без рецепторов к фактору новизны. Стресс канализует отбор – он проверяет лишь неугашенные гены. Их мало, но их адаптивность многократно тестирована в филогенезе в силу неспецифичности (отсюда быстрая эволюция в МП и параллелизмы). Т.е. возможна оценка перспективности новизны для жизни особей и для адаптивной эволюции МП (рис. 3) [тез.I-III].
Наконец, именно усталость придает стрессу свойство гиперкомпенсации при восстановлении после отдыха. Прочие молекулярно-биологические, эндокринные, популяционные и экосистемные механизмы регуляции, использующие отрицательную обратную связь, способны лишь к гомеостазу, в лучшем случае параллельному (отсюда к гомеостазу принято сводить остальные механизмы как эволюции, так и ЦП). Поскольку ПЗА лишь способствуют гомеостазу, борьба с ними с его помощью бесперспективна. Зато гиперкомпенсация, порождая возможность ненаследственной тренировки, позволяет временно угашать ПЗА типов а[16]-д (причем сама временность адаптивна – без нее нельзя оценить вышеотмеченную перспективность новизны), компенсировать ПЗА типа е и оценивать их вредность (ср. тез. V, гандикап).
Стресс в эволюции (рис.3), как и эмоция в поведении (рис.1) придает обоим процессам свойство целеполагания в поиске новизны. Поэтому в новизну широты кругозора IIa стремятся особи, прошедшие отбор не на оптимизацию уровня стресса [тез.I-68; 2, 13][17] (они стремятся в новизну педантизма IIb [31]), а на максимальную пролонгацию стресса [тез.I-III].
Литература
Ссылки на предыдущие тезисы: тез. I, II, III[18] – cм., cоответственно, тезисы Cуслов В.В. Оптимизация стресса и ароморфная эволюция. I, II, III // XXV Любищевские чтения. – Ульяновск, 2013. – С. 173-197; тез. V – тезисы Cуслов В.В. Оптимизация стресса и ароморфная эволюция V. Красота и Стресс: возможный подход к дарвиновскому объяснению эпифеномена красоты // в текущем выпуске Любищевских чтений, стр.127-144; cсылки на литературу, приведенные в предыдущих тезисах даются по схеме тез.I-1 = тезис I, пункт 1 в его списке литературы.
1.Жданов А.А. Автономный искусственный интеллект. – М., 2009. – 359с.
2. Найсер У. Познание и реальность. – М., 1981. – 232с.
3. Фефелов И.В. Такие непростые простые голуби // http://www.irk.ru/news/blogs/Fefelov/300/
4. Фридман В.С., Еремкин Г.С. Урбанизация «диких» видов птиц в контексте эволюции урболандшафта. – М., – 240 с.
5. Шипанов Н.А. Некоторые аспекты популяционной устойчивости мелких млекопитающих // Усп. Совр. Биол. 2000. Т. 120, № 1 С. 73-87.
6. Северцов А.С. Эволюционный стазис и микроэволюция. – М., 2008. – 176с. 7. Mudge J.M., Frankish A., Harrow J. Functional transcriptomics in the post-ENCODEera // Genome Res. 2013. V. 23, N 12. P. 1961-1973.
8. Колчанов Н.А. и др. Интеграция генных сетей, контролирующих физиологические функции организма // Вестник ВОГиС. 2005. Т. 9, № 2. С. 179-198.
9. Dianov G.L., Hübscher U. Mammalian base excision repair: the forgotten archangel //NAR. 2013. V. 41, N 6. P. 3483-3490.
10. Brown C.R., Bomberger M. Where has all the road kill gone? // Curr. Biol. 2013. V23, N 6. P. R233-R234.
11. Гаврилов В.М. Затраты энергии на полет, аэродинамическое качество и освоение птицами лесных биоценозов // Зоол. Журн. 2011. Т. 90, № 7. С. 773-783.
12. Brown P.M.J. et al. Harmonia axyridis in Europe: spread and distribution of a non-native coccinellid // BioControl. 2008. V. 53, N 1. P. 5-21.
13. Непомнящих В.А. Адаптация и автономия в поведении животных //Нейроинформатика-2013. Лекции по нейроинформатике. – М., 2013. – С. 106-123.
14. Редько В.Г. Механизмы взаимодействия между обучением и эволюцией //Нейроинформатика-2013. Сборник трудов. Ч.2. – М., 2013 – С. 257-266.
15. Малеев Е.А., Даревский И.С. Драконы острова Комодо // Природа. 1963. №3. C.24-35.
16. Hoelzel A.R. Killer whale predation on marine mammals at Punta Norte, Argentina; food sharing, provisioning and foraging strategy // Behav. Ecol. Sociobiol. 1991. V. 29. N3. P. 197-204.
17. Алфераки С. Тигры в Кульджинском районе // Охота и рыбалка, век XXI. 2013. № 11. C. 158-160.
18. Обухова Л.Ф. Детская психология. – М., 2006. – 203 с.
19. Сигеле С. Преступная толпа. – М., 2011. – 128 с.
20. Кирпичёв С.П. О возможности разведения выводковых птиц в естественной среде // http://www.youtube.com/watch?v=BrCGbrppCME .
21. Амелькина О.А. и др. Эффекты отбора по поведению и неонатального хэндлинга у серых крыс // XIV междунар. cовещ. и VII школа по эвол.физиологиии. – СПб., 2011. – С. 14.
22. Saltz J.B. Genetic composition of social groups influences male aggressive behaviour and fitness in natural genotypes of Drosophila melanogaster // Proc. Biol. Sci. 2013. V.280, N 1771. doi: 10.1098/rspb.2013.1926.
23. Трапезов О.В. Селекционное преобразование оборонительной реакции на человека у американской норки (Mustela vison Schreb.) // Генетика. 1987. Т. 23, № 6. С. 1120-1127.
24. Ims R.A., Yoccoz N.G., Killengreen S.T. Determinants of lemming outbreaks // PNAS USA. 2011. V. 108, N 5. P. 1970-1974.
25. Wilson P.J., et al. Discordance between genetic structure and morphological, ecological, and physiological adaptation in Lake Magadi tilapia // Physiol. Biochem. Zool. 2004. V. 77, N 4. P. 537-555.
26. Cабанеев Л.П. Рыбы России. – М., 2008. – 592 c.
27. Кроль В.М. Психология и педагогика. – М., 2001. – 319 с.
28. Goldsmith T.H. Evolutionary tinkering with visual photoreception // Vis Neurosci 2013. V. 30, N 1-2. P. 21-37.
29. Cheney K.L., et al. Colour vision and response bias in a coral reef fish // Exp Biol. 2013. V. 216 (Pt. 15). P. 2967-2973.
30. Ogura A. et al. Loss of the six3/6 controlling pathways might have resulted in pinhole-eye evolution in Nautilus // Sci. Rep. 2013. N 3. 1432 doi: 10.1038/srep01432.
31. Тимофеев М.А. и др. Некоторые аспекты эволюции терморезистентных способностей байкальских эндемичных амфипод по вертикальному градиенту условий среды обитания // Развитие жизни в процессе абиотических изменений на Земле. – Иркутск, 2011. – С. 233-239.
Источник: XXVIII Любищевские чтения, Ульяновск, 2014. С.119-127.
Комментарии:
[1] Т.е. разные стимулы (в данном случае мотивации) вызывают одинаковые реакции (в данном случае эмоции). Напротив, мотивации специфичны: вызваны данной потребностью и гасятся комплементарным ей результатом, чье невыполнение генерализует возбуждение, вытормаживая соседние участки мозга (доминанта). Cигналы, связанные с потребностью и\или результатом, тоже влияют на эмоцию, а не на мотивацию [тез.I-68]. Отсюда, доминанта интегративно оценивает организм по потребностям [1], а эмоция – по сигналам [тез.I-68; 1] и обе они требуют рецепции.
Совокупность рецепторов и циклических процессов организма формируют по Я. Икскюлю умвельт – эфферентную картину мира организма. Близкий термин – перцептивная схема [2], но умвельт более связан с наследуемой, а перцептивная схема – с ненаследуемой, приобретенной составляющей картины мира.
[2] Нерегулярные, но систематические попытки роста ареала этих горлиц на север – тоже пример быстрой инвазии, чья мотивация до конца не изучена. Типовой цикл птичьих инвазий начинает фаза широкого расселения (ФШР) отдельных особей и\или негнездующих стай – ценопопуляций. Птицы беспокойны, делят территорию, но легко бросают ее, могут формировать пары, но и отказ от гнездовья может длиться несколько лет. Только затем формируется популяция (ФП – фаза популяции), но, несмотря на успешное, даже многолетнее гнездование, инвайдеры вновь откочевывают к прежнему ареалу (ФО – фаза отступления) [4, 5, Фридман, личн. сообщ.]. Цикл, труднообъяснимый ОБС (см. ниже), может повторяться.
[3] Показателен Байкал, где изолирующие факторы – низкая температура и содержание солей в оксидированной воде [тез.I-77]. Хотя многие эндемики нерестятся в реках бассейна Байкала, т.е. преадаптированы к инвазии и преадаптации тестируются в каждом поколении, но инвазии из Байкала редки. Инвайдеры же из рек – осетр, щука, амурский сом – дали жилые морфы, хотя вода Байкала им некомфортна. Сравним: голуби, освоившие деревья, даже морфы не образовали (контбаланс светлых и темных морф голубей важен при освоении урбобиотопов [6]), т.е. если у них и отбирались комбинации аллелей, они уже рассеяны панмиксией.
[4] Как в общепринятом, так и в итал. значении – радующийся, познающий что-либо не из выгоды.
[5] Подробнее см. «Красота и Стресс: возможный подход к дарвиновскому объяснению эпифеномена красоты».
[6] Белолобые ласточки укоротили крылья, охотясь у автострад ≈ 30 лет: столкновение с машинами отбирало маневренных особей, но от короткого крыла страдали прочие летные качества [10] (у воробьиных птиц зарослей для компенсации вреда адаптации-аналога потребовался особый энергозатратный стиль полета [11]). Натиск асфальта редуцировал плодовитость и расселительные адаптации урбопопуляций растений [тез.I-III]. Инвазию азиатской божьей коровки сдерживала инбредная депрессия, элиминированная при искусственном разведении для борьбы с тлями, т.е. долгим противоестественным отбором на прожорливость при высоких численностях популяций [12]. Итак, ввиду прямого снижения γ, отбор легко образует, но плохо элиминирует ПЗА типа е, хотя они не дают роста IIa и IIb, и вообще могут быть не связаны с рецепцией.
[7] Иначе выражено в парадоксе Найсера [2]: верно достигнутый результат несет недостаточно информации о своем достижении.
[8] При моделировании эффекта Болдуина априорно задаются сходство приобретенной и врожденной адаптации [14], способность к инвазии [тез.I-III].
[9] Неспособность отличить Луну от открытого огня (лесной пожар) ночью губит массу насекомых, хотя оба фактора сопровождали весь филогенез насекомых. Гипертрофированная демонстрация лишь подобия раздражителя, провоцирующая ИМ (подманивающее поведение рыб-удильщиков, каймановых черепах [тез.I-34]), или, наоборот, назойливая демонстрация – типичное поведение хищников [тез.II-49; 15,16]. В последнем случае хищник может либо эфферентно четко – не особо прячась – позиционировать себя, угашая таким образом и ИМ, и стресс у жертв (если жертва легко выдерживает безопасное расстояние: охота рептилий на млекопитающих [15], косаток на молодых морских львов у лежбищ [16]), либо постоянно провоцировать ИМ и/или стресс (вплоть до паники) жертв, ярко, но нечетко позиционируя себя. Так вокализация, мечение, реже патрулирование в виду жертв – элемент поведения пантерин [тез.II-49]. Копытные же уходят на расстояние, недостаточное, чтоб снизить интенсивность их атак, но достаточное, чтобы не чувствовать постоянного присутствия хищника (или его запаха [17]) – налицо рост IIb, но не IIa.
[10] Глухарь – типичный урбофоб, незнакомые сигналы повышают его тревожность, побуждая отход вглубь биотопа. ♀ импринтирует уровень тревожности птенцов, вокализируя перед вылуплением. Проигрывание вокализации спокойной ♀ снижало тревожность детей, которые, в свою очередь, снижали тревожность внуков [20]. Аналогичен эффект при хендлинге (ласка руками) беременных крыс [21].
[11] Универсализм бихевиоризма не устраняет, а маскирует эффект: в целях контроля животному дают филогенетически знакомую обстановку [тез.I-34]. Иначе – отбор на редкое поведение (1, агрессия… у дрозофил [22]), на сосуществование с незнакомым стрессором (2, доместикация [23]), на незнакомое сочетание знакомых стрессоров (3, дрессура [тез.II-38]) – нужна настройка отбора на рецепцию, т.е. выработка метода селекции, тем более особого, чем стереотипнее поведение вида. Так, в 1,2 нужно дозирование действия стрессора во времени и пространстве [22, 23], в 3 – подбор видов стрессора и стресс-лимитатора [тез.II-38]. Бихевиоризм здесь ведет к невозможности ответа на отбор на популяционном (даже при высокой дисперсии) и на индивидуальном (даже при наличии преадаптаций) уровнях [тез.II-38; 22].
[12] “Никто не помнил как добрался домой” – А.Конан-Дойль, “Cобака Баскервилей”; забвение череды даже успешных действий при аффекте известно в психологии [18, 19], но блокада аффектом влияет и на эволюцию. Темп вспышек численности леммингов предетерминирован экологией [24], а заплывы через узкие фьорды – постледниковым ландшафтом Норвегии [тез.I-59]. Но адаптаций к плаванью, несмотря на вековые повторности и некоторую изоляцию норвежской популяции нет: вне стаи особь, переплывшая фьорд (или ее потомок), воды боится.
[13] “Стратегия кита” отмечена и в аэрированной воде, если в ней повреждающие жабры агенты (содовые термоозера Африки [25], “темное половодье” наших рек [26]), или если рыбы резко ускоряются или дельфинируют (череда прыжков резко снижает сопротивление воды) [тез.I-II]. Отсюда само по себе формирование элементарного дивергентного цикла Гилярова-Симпсона (ЭДЦ) порождает собственный вектор эволюции даже в совершенно стабильной среде [тез.I-99]. ЭДЦ объясняет смену траекторий онтогенеза при полной их эквифинальности. ЭДЦ двух полов объясняет “cценарий турухтана” (формирование фригидных ♀ для возбуждения которых нужно несколько ♂), который А.А.Любищев считал дарвинистски невозможным [тез.II-27]. На молекулярном уровне ЭДЦ гомологически соответствует режиму эволюции коадаптивных замен.
Т.е. циклы Гилярова-Симпсона широко распространены и смена сред скорее мешает, чем помогает им. Утрата легочного дыхания описана у хвостатых и бесхвостых амфибий [тез.I-99, тез.I-26; 6]. Сценарий ЭДЦ-шунтирования онтогенеза в стабильной среде объясняет оба случая, а адаптивная смена дыхания при пищевой специализации подъязычного аппарата – только случай хвостатых амфибий [тез.I-26] (не исключена у них работа обеих механизмов). Утрата жаберного дыхания при ЭДЦ-шунтировании онтогенеза рыб не описана, но не исключена.
[14]Известно восстановление путем обратных или супрессорных мутаций полихромного зрения у приматов и рыб коралловых рифов [28]. Но рифовые хищные рыбы олигохромны, хотя полихромное зрение облегчает на фоне кораллов поиск жертв, использующих аналогичное зрение для внутри- и межвидовых коммуникаций [29], а структура родопсина облегчает реверсии [28]. Устойчивость умвельта позволяет объяснить сохранение глаз-обскур у наутилуса. Если утрата хрусталика у его предков и была по какой-то причине адаптивной, то отсутствие реверсий у современных видов загадочно: кристаллины – лучший пример конвергенции негомологичных белков [28]. Но у наутилуса даже дисперсии по признаку нет) [30].
[15]Ср.в тез.V феномен ускорения течения времени (угашение ПЗА типа а) при опасности при одновременном сокращении перцептивного поля – отключения слуха, тактильности и т.п. Эксперименты [тез.V-86] показали субъективность феномена ускорения, однако были поставлены так, что отсчет времени никак не повышал адаптивность. Ю.Росциус привел подборку наблюдений, где именно отсчет времени повышал адаптивность, что возможно только при объективном переводе работы ГС типа а в новый режим.
[16] А.А.Любищев отмечал: практикуемый им учет времени привел к улучшению способности оценки временных затрат при любой деятельности.
[17]Такой отбор ведет к КЭ, именно его прошли эндемики Байкала [31]. Ввиду сложной динамики стресса (три фазы – тревоги, ПР и дистресса без учета субфаз), ОБС эволюции стресса часто вырождается в отбор на оптимизацию одной фазы в ущерб другим, или снижает неспецифичность, переводя стресс в другой тип ПЗА. Даже просто снижение стресса из-за роста комфортности абортирует успешную инвазию. У птиц в оценке комфортности биотопа важны территориальные сигналы: кормный биотоп без соседей кажется менее комфортным, что объясняет противоречивое поведение птиц в ФШР [тез.I-59; 4,5]. Лишь когда территория более-менее сигнально покрыта, ее стрессогенность падает – наступает ФП: птицы гнездуют, растет численность [тез.I-59] и сигнальное покрытие [4], что провоцирует отбор на оптимизацию стресса (a), новые миграции стресс-неустойчивых особей из предкового ареала (b), но снижает отбор на пролонгацию стресса (c). В итоге, успешная инвазия замирает: субпопуляции b и a вместе запускают КЭ, и ФП сменяется ФО.
[18] Их краткий пересказ см. «Ламарк и стресс: дарвинистское объяснение основных эпифеноменов ламаркизма»
Рекомендуем прочесть
 Как Венгрия стала коричневой
Как Венгрия стала коричневой Преподавание эволюции в Саудовской Аравии и Иране
Преподавание эволюции в Саудовской Аравии и Иране Про акмэ идеи «естественности неравенства полов»
Про акмэ идеи «естественности неравенства полов» Почему лгать лучше только за деньги?
Почему лгать лучше только за деньги? За пределами роста-1
За пределами роста-1 Динамика психологического состояния российского общества: экспертная оценка
Динамика психологического состояния российского общества: экспертная оценка О происходящем на Украине
О происходящем на Украине Кошка, пахнущая попкорном
Кошка, пахнущая попкорном
