

(о книге Д.А.Жукова по «биологии поведения человека и других зверей»).
«В поле бес нас водит, видно,
Да кружит по сторонам».
А.С.Пушкин
Исследования поведения: «морфология» vs «побуждений» и «нравов»
Содержание
- 1 Исследования поведения: «морфология» vs «побуждений» и «нравов»
- 2 Инстинкт и механизмы поведения
- 3 Что первично, поведение или состояние животного?
- 4 Запасание и поиск запасов
- 5 Родительское поведение грызунов
- 6 Тактильная стимуляция и другие социальные влияния на заботу о потомстве
- 7 Рецепторная концепция действия гормонов
- 8 Методологические изъяны
- 9 Заключение
Активность животных, развёртывающуюся перед наблюдателем, можно анализировать с двух разных позиций. Первая следует наивному антропоморфизму: наблюдаемые паттерны активности одиночных особей или участников взаимодействия автоматически полагаются внешними проявлениями «намерений», «побуждений», «стремлений», «нравов», «привычек», «эмоций» и пр., как если бы это был действующий человек. Получается «платонова пещера наоборот»: всё непосредственно наблюдаемое во внешнем мире есть пляска теней внутренней жизни, «выражение эмоций», если вспомнить известную работу Дарвина, или «намерений», «привычек» и «нравов», если Эспинаса. Задача исследователя – интерпретация «внутренней жизни» животного, её напряжений и конфликтов по деятельности во внешнем мире.
В XIX веке, когда антропоморфизм господствовал безраздельно, упор делался на «психическую» составляющую внутреннего мира, «намерения», «привычки» и «нравы». В ХХ, по мере того, как он всё больше дискредитировался и изживался, перевешивало «физиологическое», от мотиваций до концентраций конкретных гормонов и медиаторов в крови и других жидкостях тела. С этой методологической позиции началась зоопсихология; исходный упор на объяснение наблюдаемой деятельности через действие внутренних детерминантов (всегда остающееся предполагаемым, с возможностью найти лучший детерминант или лучшую их комбинацию) сохраняется по сей день.
Замечу, что «ультрасовременная» позиция биологизаторов, объясняющих действия «человека и других зверей» эффектом гормонов и медиаторов (или «генами», или иной «биологией»), вполне изоморфна объяснениям своих предшественников XIX, оперирующих «привычками» и «нравами». Просто сегодня мы больше знаем о физиологии, молекулярной биологии и генетике процессов внутри организма, почему соответствующие объяснения выглядят «научнее» (а, значит, и убедительней, Аронсон и др., 2008). Однако они страдают теми же дефектами в сравнении с альтернативой, обсуждаемой ниже.
Книга Дм.Жукова, как популярный жанр, написанный красочно и доходчиво, демонстрирует эти изъяны исключительно выпукло, даже карикатурно. Собственно, главный изъянов два. Первый — это концепция автора, с позиций которой он объясняет читателю поведение животных и человека; она не соответствует фактам даже для группы, с которой он сам работает как физиолог (грызуны). Второй — методологическая позиция: поскольку объяснения автора не соответствуют фактам не только для человека, но и животных, он компенсирует это подачей собственных взглядов как «единственно научных», соответствующих «бритве Оккама». Но пользуется ею неадекватно (о чём отзывавшиеся о книге уже писали), и забывает про не менее важный методологический принцип — необходимого разнообразия Эшби. Что сильно вредит и «научности» излагаемого автором, и состоятельности способов доведения этих идей или фактов до нас, читателей.
Вернёмся к интерпретациям активности животных, особенно социальной. Подход, альтернативный зоопсихологии позавчера (фактически и биологизаторству сегодня) – это сравнительная этология. Она появилась позднее, как только в начале ХХ века возникло понятие о поведении животного (Гороховская, 2001). Е.А.Гороховской в исследовании истории этологии специально показано, что это не то же самое, что «поведение» в обыденном языке или у зоопсихологов.
Инстинкт и механизмы поведения
Также как «инстинкт» в обычном словоупотреблении или у неспециалистов – это феномен внутреннего мира, специфическое (наследуемое) побуждение. У этологов же это не гибкое слово, а строгий термин. Это формы и структуры поведения в мире внешнем, закономерно развёртываемые животным в определённой ситуации взаимодействия, «высвобождаемые» там специфическим ключевым стимулом (также репрезентируемым одной из N инстинктивных форм поведения, распознаваемых по характерному – стереотипно воспроизводимому – облику и составляющих видовой репертуар, этограмму). Или при длительном отсутствии специфических ключевых раздражителей, высвобождающиеся спонтанно в т. н. «реакциях вхолостую» (обычно повторяющимся образом).
Поэтому соответствующие формы и структуры (видо)специфического поведения исследуются с использованием «морфологического подхода» к поведению (неслучайно и Лоренц — ученик сравнительного анатома Фердинанда Хохштеттера). Что хорошо понимали не-этологи, также работавшие именно с поведением как развёртывающейся и самоорганизующейся структурой деятельности, а не «внутренней жизнью», скажем психологи культурно-исторической школы. «Инстинкт, эта генетически первичная форма поведения, рассматривается как сложная структура [курсив мой — В.К.], отдельные части которой слагаются наподобие элементов, образующих ритм, фигуру или мелодию» (Словарь Л.С.Выготского, 2004: 44).
Иными словами, инстинкт — это «должные» формы (видо)специфического поведения в «должной» ситуации взаимодействия во «внешнем мире» животного (сходство с онтогенезом здесь совсем неслучайно), а не «врождённые потребности» в мире внутреннем, лишь проявляющиеся во «врождённой программе действий». См. пример такого (неверного) понимания: неслучайно без ссылок, да и стереотипность воспроизведения «должных» форм телодвижений в «должной» ситуации взаимодействия для инстинкта куда важней их «врождённости», см.ниже.
Увы, 80 лет развития этологии прошли мимо Дм. Жукова: «инстинкт» и «инстинктивный» он везде использует в обыденном, не в этологическом смысле — как специфическое врождённое побуждение (или потребность) из «внутреннего мира» животного, а не специфические формы демонстраций во внешнем. [При этом, увы, рекомендует «книги по этологии» на Постнауке: из 5 книг одна — «мифология в этологии» В.Р.Дольника, другая же — замечательная книга Р.Левонтина — убей бог, не имеет к этологии отношения.]
И игнорирует (хуже, если не знает) огромную массу данных, показывающих неадекватность такого подхода. Скажем, у трёх наших видов поморников, крайне похожих друг на друга, территориальная агрессия и агрессия в отношении хищников (песцов, сов, зимняков, 2-х других видов поморников) выражена радикально различными демонстрациями, позными и акустическими, при общей физиологии того и другого. И, естественно, те и другие демонстрации видоспецифичны, т. е. одинаковые внутренние состояния состояния разных видов, существенно сходных внешне, выражены различными демонстрациями (Рябицев, 1995).
Другая ошибка — определяя «инстинкт»/«инстинктивное», автор концентрируется отнюдь не на специфике форм демонстраций и закономерной организации последовательностей обмена ими во взаимодействиях, в ходе которого особи кооперативно реализуют соответствующий видовой инстинкт — территориальный, брачный и пр. (притом что прогресс в этологии 1940-1970-х гг. связан с исследованием именно этого, см. основные вехи истории дисциплины). Нет, он определяет его через «врождённость побуждений», выраженных в инстинктивных формах поведения (а на деле его лишь «питающих» его своего рода энергией действия).
На деле у высших позвоночных — птиц и млекопитающих — врождённа лишь «болванка» инстинктивного действия, в которой при первом появлении дефинитивную форму никак не узнать. Затем оно «доводится до кондиции» в ходе взаимодействий между соответствующими компаньонами; скажем, взрослыми чайками и птенцами, всё точнее клюющими цветное пятно на конце. См. классическое (пере)исследование J.P.Hailman (1967, 1968, 1971) «реакции клевания» у чаек («Как птицы обучаются инстинктивному поведению?»).
Значительное участие обучения и других форм «матричного копирования» поведения других особей в становлении видоспецифических форм поведения ничуть не мешает последующему функционированию их как инстинктов, т.е. «автоматическому» высвобождению в ответ на специальный ключевой раздражитель (релизер) и некорректируемость «высвобожденного» инстинктивного действия средой или индивидуальным опытом животного, даже если без этой коррекции действие неуспешно и контрпродуктивно. Это т. н. «слепота» и «автоматичность» инстинктивного действия, которые и позволяют его распознать, отделив, с одной стороны от рефлексов, с другой — от стереотипов, созданных обучением. Больше того, исследование поведения близких видов показывает, что однотипные реакции, вроде поведения накалывания у сорокопутов, избегания жертв с предупреждающей окраской у синиц или ныряния при накрывании сверху у одного вида могут быть инстинктивными, у другого же — созданными обучением. «По первому впечатлению» не догадаешься, в каждом случае надо специально проверять… а авторский рассказ «о поведении» всё смешивает в кучу.
Фактически автор игнорирует весь массив знания об инстинктах, накопленный этологией в ХХ веке, и вместо него предлагает читателю нечто своё, фактически обыденное представление, в котором инстинкт путается с рефлексом, и оба они вместе взятые — со стереотипами, созданными культурой (последние столь же «автоматичны» и «слепы», но, в отличие от инстинктов, допускают «размягчение» и «переделку», см. про танец и преодоление различий «мужских» и «женских» профессий в СССР 1930-х гг.). В итоге читатель оказывается дезинформированным и даже обведённым вокруг пальца — ему обещали «биологическое» объяснение человеческого поведения как «наиболее научное», на деле же скармливают старый добрый антропоморфизм.
Животное, как и человек, в интерпретации автора, действует на основе побуждений и представлений «изнутри», только, в отличие от нас, оно «будет попримитивней», его «внутреннее» включает максимум физиологии, минимум психики с интеллектом и практически ноль социальных влияний. Что неверно: формообразующая роль социальных влияний у позвоночных животных не меньшая, чем у человека, просто влияния другие (а частично и механизмы их передачи «внутри» организма), о чём далее.
Понимая опасность антропоморфизма, будущая этология строго отделила наблюдение от интерпретаций. Первое – это структура действий животного, развёртываемых во времени их пространстве (1; единственное эмпирическое, фиксируемое наблюдателем), формы и фигуры, описываемые телодвижениями актора при подобном развёртывании (2) и (3) численные закономерности такого развёртывания, скажем, порядок следования (или правила смены) форм отдельных элементов в последовательностях. (1) – это и есть поведение, (2) или (3) – его описание, выполненное на основе «морфологического подхода» к нему, в центре которого – выделение специфических форм и структур, установление эмпирических закономерностей их развёртывания и пр. Поведение понимается как временный орган животного, устойчиво воспроизводимый «должным образом» и в «должной ситуации» взаимодействия. Первое означает стереотипность воспроизведения каждого элемента (иначе не «выпишешь» его специфической формы) и отделённостью от форм других элементов (которая может быть большей или меньшей, редко – почти никакой, рядом континуума). Второе означает ситуационную специфичность употребления элементов с разной формой, которая тоже бывает большей или меньшей, границы и правила которой надо ещё установить (и часто это нелегко).
Что первично, поведение или состояние животного?
Для этолога «поведение» 1) характеризуется специфической формой каждого элемента или паттерна активности (своими телодвижениями «описывающего» эту форму как огоньком сигареты чертят буквы в темноте) и 2) объясняется «из себя самого», причиной употребления «должных элементов» в «должной ситуации» взаимодействия является одновременное поведение другого участника, или предшествующее поведение самой особи. Параметры состояния же, «психические» или «физиологические» оказываются не причиной, а одним из условий, влияющим на качество реализации данного поведения в данных обстоятельствах контекста – наряду с риском хищничества, погодой, и другими событиями внешнего мира, не относящимися к обмену поведением между участниками взаимодействия.
Второе – это параметры внутреннего состояния актора, его «мотивация», «побуждение» и пр. Будучи лишь интерпретациями, они существуют практически в статусе полезных фикций: для них не предполагается онтология, они сохраняются лишь там и тогда где полезны как промежуточная переменная, связывающая сигналы (или релизеры) на входе системы организм с закономерной структурой поведенческого ответа на выходе. Соответственно, они рассматриваются не как «причины» происходящей активности и не «объяснения» выбора (или смены) действий в проблемной ситуации.
Всё это этологи ищут «во внешнем мире», в поведении же – либо в одновременно удерживаемых демонстрациях партнёра, либо в паттернах активности, ранее реализованных самой особью). Это всего лишь «источник энергии», поддерживающий формы поведения, реализуемые особью, причём неспецифический; специфика, видовая и ситуационная, задана поведением же, результатами прошлой активности; она интериоризируется во «внутреннем мире» участников в виде специфической мотивации, сдвига гормонального состояния и пр. лишь вследствие достижения специфических результатов поведения.
Биологический смысл самоорганизации инстинктивного поведения под действием ключевых раздражителей извне (а не побуждений «изнутри») актора виден на примере осенне-зимнего долбления большим пёстрым дятлом шишек сосны и ели на «кузницах». Под влиянием чего птица, раздолбив шишку, летит за другой; срывает отточенным движением её с кормового дерева; несёт в клюве (реже в лапах) на кузницу, там прижимает грудью к стволу, после чего вынимает старую шишку из «станка« и выбрасывает, а потом ставит на её место новую и начинает долбить? Под влиянием голода? Отнюдь нет: исследования бюджета времени и энергии показывают, что питание семенами хвойных обеспечивает дятлу в среднем девятикратный перевес «прихода« энергии над «расходом« (Бардин, 1982).
Остаётся даже на «активное бездельничанье», как и у других видов птиц и млекопитающих при устойчивой жизни, когда нет затяжной непогоды и других бедствий (см. его уморительное описание у самца серой вороны). Можно было бы и отдохнуть: но нет, владелец участка таскает шишки на «кузницу» и меняет старые на новые как часы. Что управляет такой регулярностью? Ключевой раздражитель в виде старой шишки в «станке» из которой, как знает владелец, выбраны все семена. Когда это произошло, дятел не может не удержаться, чтоб не слететь за новой, а, принеся её, обязательно прижмёт новую грудью и дёрнет клювом за старую шишку, чтобы вынуть.
Это инстинкт, автоматичный настолько, что используется для отлова дятлов на «кузницах«. Орнитологи настораживают лучок за старую шишку а дятел, прилетев, дергает за неё и спускает сторожок (Приклонский, 1962). Он не может не дёрнуть, хотя видит это странное сооружение; дёргает даже в случае, когда уже попадался и пр. Хотя вообще дятлы — умные птицы, с высокоразвитой рассудочной деятельностью, на инстинктивную часть поведения это не распространяется не только у них но и, скажем, у мартышек Диана; инстинкт неизменно «слеп». Именно отсутствие такого рода реакций у людей и антропоидов (или, например, «инстинктивного смещения» Брелендов) заставляет считать, что у тех и других нет инстинктов.
Так или иначе, видоспецифическое поведение здесь организуется прошлыми действиями, их успехом или неудачей, но отнюдь не голодом или иным побуждением (если дятла спугнуть при долблении, он летит на кормовое дерево, «автоматом« срывает шишку, но летит не на прежнюю кузницу, а в другую часть участка, где также есть кузница со старой, невытащенной шишкой, стимулирующей её заменить и «работать« над новой). До «толчка изнутри», полагаемого Жуковым обязательным, дело попросту не доходит, поведение опережает его. Сходным образом богач вносит деньги на счёт никак не для того, чтобы сберечь их от повседневных трат, а для последовательного увеличения свободы в будущих операциях, каковы бы они ни были.
Другой пример: хищническое поведение, особенно в отношении крупных жертв, способных эффективно сопротивляться, «спускается» отнюдь не голодом охотника, а беспомощностью (или, шире, уязвимостью большей чем обычно) добычи. Соответственно, оно не имеет других тормозов, кроме их исчерпания, когда все доступные жертвы мертвы или сумели отбиться. Если беспомощных жертв слишком много, хищник не останавливается, пока не зарежет всех, независимо от собственной сытости или усталости. «Волки убивают всех находящихся в пределах досягаемости оленей или косуль, вязнущих в глубоком снегу, т.е. больше, чем могут съесть. Нет торможения реакции хищника при нападении на стадо домашних овец, в телятниках, либо на средних и малых копытных в вольерах и др.»
Вообще, для хищных млекопитающих показано, что одним из сигналов доступности жертвы, запускающих упорное преследование, являются изменения микрофлоры на поверхности мочки носа, свидетельствующие о долговременном стрессе, приведшем уже к какого- либо рода патологии. Практическим все животные, добытые хищником в природе, или же пойманные борзыми в специальных напусках на сайгаков и зайцев для изучения соответствующего феномена, несли разные серьёзные патологии. Напротив, охотники, стремившиеся отстрелять именно больных сайгаков или зайцев, примерно в трети случаев убивают вполне здоровых (Северцов, Шубкина, 2015).
Поэтому мнением автора, что «действия осуществляются на основе побуждений« неверно даже для животных, не то что для человека. В нормальной ситуации до этого дело просто не доходит, почему организмы могут устойчиво существовать и направлено двигаться к определённой цели, преодолевать неблагоприятные обстоятельства и т.д. «сопротивление среды». Тем более это верно для человека: никакое из его побуждений «не работает» без опосредования «должными« символами соответствующей культуры и не удовлетворяется иначе, как в соответствующих культурных формах.
Скажем, автор почему-то считает, что неудача «сухого закона» говорит о необоримости «потребности в удовольствии», и именно алкогольном. Однако французы, некогда больше всех потребляющие этанола в Европе, примерно за 30 лет радикально снизили потребление, и курение в цивилизованных странах успешно теснят — хотя удовольствие от него не меньшее. Фиаско «сухого закона» много разумней объяснять особенностями американского неограниченного капитализма и следующем отсюда торжестве организованной преступности, а не «природой человека».
В рамках этологического подхода ошибочно полагать, что «гормоны», «гены» или иная физиология «определяют» поведение, как причина — следствие. Верно как раз обратное: пламя поддерживает и регулирует само себя, а дрова лишь «питают» костёр, но не «управляют» пламенем. Также поведение регулируется и определяется поведением же (что бы ни вкладывай в эти два слова). Специфическое состояние «внутреннего мира» животного, гормональное ли, мотивационное ли, лишь поддерживает будущее поведение и в свою очередь, формируется результатами прошлой активности. Почему следует принимать именно эти рамки, а не милые сердцу Дм. Жукова? Потому, что сейчас уже точно известно, что вопрос «что первично — поведение или состояние животного?» разрешается в пользу первого.
Формообразование поведения, его спецификация идут в первую очередь под воздействием социальных влияний, одновременно «интериоризующихся» в виде специфики мотивационного состояния (о чём см.рассказ). И наоборот: ошибочна противоположная точка зрения, что формообразование и спецификация поведения обеспечиваются эффектом гормонов или иным влиянием «внутреннего мира». Действительно, когда социальное влияние входит в конфликт с эндогенной детерминацией, именно оно пересиливает, а не наоборот. Поэтому эффект тестостерона и других гормонов, на которые делается упор в книге Жукова, неизменно социально-релятивен.
Данные об эффекте социальных влияний в сочетании с опытами Дарио Маэстрипиери и др. исследованиями «сигнальной наследственности» млекопитающих и птиц показывают, что всякое формирование агрессивного, полового, родительского и т. д. поведения объяснимо этими двумя причинами «без остатка». Причём экзогенные влияния на формирование поведения наиболее значимы, а эндогенные особенно слабы, с одной стороны — у грызунов (группа, которой занимается автор, правда, не в поле, а в лаборатории), с другой — у приматов (Крученкова, 2002).
Квинтэссенции эти тенденции достигают, естественно, у человека разумного, т. е. используемый Жуковым объяснительный принцип именно к нам всего непригодней. Впрочем, он не годится и для «социальных» видов грызунов и других позвоночных (а их большинство в сравнении с «несоциальными» родичами). Однако же он проводится автором «железной рукой», при полном игнорировании несовместных с ним данных исследований. В чём можно видеть уже не научную, а идеологическую позицию: все привержены собственным теориям и подходам сверх меры, но полное исключение из обсуждение конкурирующих объяснений и особенно, поддерживающих их данных, говорит об идеологии. Научный подход, пусть в популяризации, требует рассмотреть альтернативные объяснения (т. е. изложить их читателям) и отвергнуть на неком достаточном основании.
Несколько искажая действительность (и тем самым следуя подходу Дм.Жукова, пусть в отличие от него — в направлении, подкрепляемом фактами), можно сказать, что какие-либо гипотезы об эндогенных детерминантах наблюдаемого поведения просто излишни и должны быть «отрезаны» бритвой Оккама. На деле, конечно, они крайне важны, когда речь идёт о поддержании (видо)специфического поведения «должное время и в должном месте», чтобы преодолеть сопротивление оппонента, неблагоприятные обстоятельства демонстрирования и пр. Без «подпитки дровами костра» ничего здесь не сделаешь, но в формировании специфичности поведения их вклад минимален.
Действительно, сейчас показано отсутствие «собственной онтологии» у гидравлической модели мотивации, врождённых разрешающих механизмов и других популярных моделей эндогенных влияний на поведение; у разных видов и в разных ситуациях это «делается» разными физиологическими эффектами. Вот два ярких примера, когда реальные звери и птицы (также как рыбы и гады, а тем более все мы, грешные) ведут себя совершенно не так, как объясняет автор.
Запасание и поиск запасов
Он пишет: «Потеряв нужную вещь, конечно, можно строить различные гипотезы: похищение врагами, провал в четвертое измерение, одушевление предмета и т. п. – и действовать в соответствии с ними, планируя поиски. Но наше поведение будет более эффективным, если мы будем исходить из того, что сами куда-то положили эту вещь. Следовательно, надо лишь вспомнить, когда и где мы видели ее в последний раз».
Неправильно! Поскольку память и воображение — это одно и то же чувство, усилия по «припоминанию» лишь плодят фантомы, см. исследования ложных воспоминаний Э.Лофтус и др. Чем пользуются нечистоплотные следователи: давя на подозреваемых и свидетелей, они фабрикуют ложные самообвинения и «доказательства». Поэтому так организованный поиск только плодит разочарования после каждой надежды «вот, точно! Сюда положил». Как только мы исходим не из «внутреннего» (побуждение вспомнить), а из реальных закономерностей организации поискового поведения у людей и животных, то действуем совершенно иначе. Бездумно, но методично и тщательно перебираем все «уголки», куда можно засунуть искомое, и так как их число в любой комнате, кабинете и т. п. «участке обитания» ограничено, оно находится в одной из ниш.
Именно так ищут спрятанные запасы корма птицы, славящиеся исключительно сильной пространственной памятью — синицы-гаички, кедровки и сойки, осенью запасающие корм в количествах, достаточных для питания в течение холодного времени года. Осенне-зимние участки обитания этих видов покрыты ранее сделанными запасами, и владелец устойчиво эксплуатирует территорию, последовательно возвращаясь к ним, причём помнит исключительно точно.
На протяжении осени и зимы они ловко и сразу отыскивают созданные запасы, практически без проб и ошибок вокруг спрятанного. Но! Они почти полностью неспособны к отысканию таких же запасов (созданных в этой же микростации, их товарками или экспериментатором), но на незначительном отдалении. Это различие исключительной точности поиска в первом случае и ненахождением во втором не зависит от индивидуальных топомнезических способностей птиц и сохраняется даже если последние велики (разные виды врановых; они все запасают корм, даже виды, не столь специализированные в этом, как сойки с кедровками) См. Vander Wall, Balda, 1981; Sherry, 1989; Sherry, Vaccarino, 1989.
Это означает, что механизм поиска запасов «работает» не через «припоминание» их размещения, а следствие инстинктивной реакции движущейся по участку птицы на релизеры, заданные особенностями «просматриваемых» микростаций (для синиц или врановых это архитектурно различные части кроны или участки земли). Примерно также развёртывается поведение ухаживания и угрозы каждой особи в ответ на демонстрации второго участника взаимодействия (или собственные прошлые демонстрации), но достаточно независимо от уровней специфической мотивации или общего возбуждения акторов.
Действительно, когнитивные карты осваиваемого птицами пространства скорее перечень правил принятия решений в ответ на сигналы местообитания, нежели карты в собственном смысле, поскольку птицы не запоминают напрямую, где и что им надлежит искать (Krebs, Biebach, 1989; Sherry, Vaccarino, 1989; Vallotigara, Zanforlin, 1989). Этот взгляд позволяет одновременно объяснить и быстрое расхищение запасов птиц, и неспособность их найти запас, заложенный экспериментатором совсем рядом с запасом самой птицы (Бардин, Марковец, 1990). Иными словами, птицы и животные вообще не «ищут» спрятанное, а «наталкиваются» на него, эта находка — «автоматический» продукт развёртывания их активности по определённым правилам. По той же причине неспособность птиц найти гнездо с кладкой при малейшем его перемещении резко контрастирует со способностью неограниченно следовать за гнездом после появления птенцов, с которыми родители связаны общей социальной активностью (Дерим-оглу, 1964; Благосклонов, 1977).
Поэтому физиологам, исходно предрасположенным к «зоопсихологическому» способу интерпретации, надо быть особенно осторожным, чтобы не «наступать на грабли», связанные с умалением достижений главного («этологического») подхода. Один из способов «предохранения» от такого фиаско — стать «компетентным читателем» этологических исследований (выполненных в рамках второго подхода), чтобы корректировать свои мнения, по их результатам.
Увы, Дм.Жуков не то что не делает этого, но полностью игнорирует соответствующие данные (упоминая работы Лоренца и других этологов там, где они сходятся с ним во мнении, но не в их основных достижениях — что ещё раз показывает не научный, но идеологический подход автора к изложению проблемы). И рецензенты — также физиологи — нигде не поправили автора, почему получается нечто вроде средневековых попыток взлететь на искусственных крыльях: вроде похоже на «настоящий полёт» птиц, но не работает. Может быть, книгу, посвящённую закономерностям поведения (особей в популяции), а не физиологии организма «внутри себя», стоило дать на рецензию специалистам — этологам и социальным психологам.
Тогда, наверное, можно было бы избежать большей части ляпов, связанных с экскурсом автора в чужую предметную область (аналогичный контроль специалистов-гуманитариев позволил бы избежать ещё больших ляпов при экскурсах автора в вовсе чуждую ему область, истории, экономики, социологии и пр. культуры).
Родительское поведение грызунов
Другое важное правило, следующее из разделения «поведения» и «состояния» животного как наблюдаемого и интерпретаций: «животных» нельзя объединять в противопоставлении «человеку». Каждый вид обладает собственной социальной системы, репрезентируемой в пространственно-этологической структуре популяции: она непрерывно воспроизводится в контактах и взаимодействиях особей, регулируя их течение и результат (а не наоборот, как может казаться), обеспечивая определённые частоту и интенсивность взаимодействий индивидов и пр.
Обусловленные ею различия в социальных связях и образе жизни у разных видов и/или у особей с разным социальным статусом влияют на их поведение много сильней и (главное) специфичней, чем гормональные влияния «изнутри». Хороший пример здесь: родительское поведение грызунов, подробно исследованное В.С.Громовым (2013); первый раздел книги (с.12-39) как раз посвящён интересующему нас вопросу.
Прежде всего, гормональные влияния на родительское поведение грызунов противоречивы, часто неустойчивы, их характер меняется от вида к виду (таблица).

Напротив, паттерн соответствующего поведения у исследуемых видов, резко различных по экологии и социальным системам, «устроен» достаточно универсально и сходным образом «самособирается» в деятельности животного, см. рис. 2.7.6. книги.
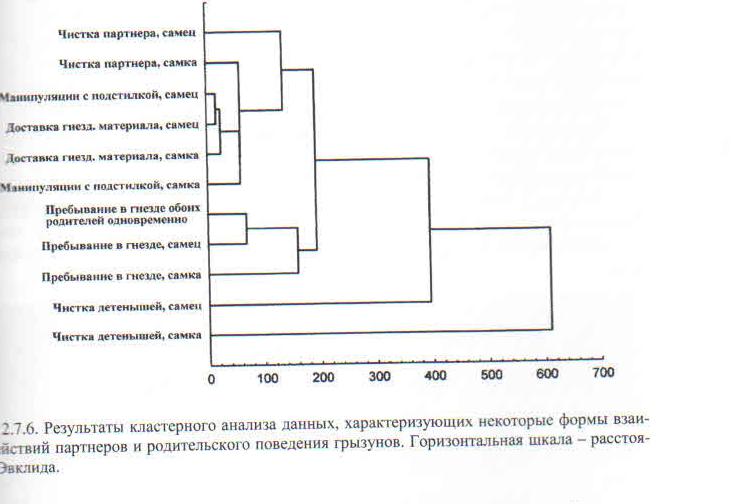
Обратите внимание, что тесная взаимосвязь между разными формами родительского поведения сохраняется независимо от видовой принадлежности (и гормонального состояния, очень разного у этих видов см.ниже). Наконец, эффект социальных влияний постоянен, определёнен, единообразен у всех видов.
Так, у самцов пролактин активирует те же центры головного мозга, ответственные за стимуляцию родительского поведения, что у самок. В том числе введение пролактина в мозг стимулирует заботу о потомстве у взрослых самцов крыс. И наоборот: у молодых самцов уровень пролактина повышается, когда демонстрируют заботу о потомстве. У видов с семейно-групповым образом жизни и отцовской заботой о потомстве (калифорнийский хомячок, монгольская песчанка, хомячок Кэмпбелла) концентрация пролактина в сыворотке крови самцов возрастает после рождения детёнышей, совпадая с первыми признаками проявления отцовской заботы. С другой стороны, самцам прерийной полёвки длительно вводили антагонист пролактина — бромкриптин, но снижение концентрации гормона не влияло на уровень родительской заботы.
У крыс и мышей родительское поведение стимулируется эстрадиолом и блокируется прогестероном (что действует и на крыс, не имеющих сексуального опыта). Функция прогестерона двойственна: его длительная секреция адаптирует организм матери к высокому уровню секреции эстрадиола в конце беременности и при родах, снижение же его концентраций незадолго до родов своевременно активирует материнское поведение. У самок домовых мышей, наоборот, поведение подготовки гнезда стимулируется высоким уровнем секреции прогестерона и пониженным — эстрадиола. В отличие от них обеих, у сирийского хомяка стероидные гормоны не участвуют в регуляции прямой материнской заботы, и уровень секреции эстрадиола и прогестерона в последние дни беременности падает до минимума.
Таким образом, роль стероидных гормонов в регуляции материнского поведения разная у разных видов грызунов. Кроме того, секреции одних лишь стероидных гормонов недостаточно для полноценного проявления материнской заботы.
После родов материнское поведение практически не зависит от гормонального статуса (во всяком случае от уровня секреции половых стероидов). Больше того, после выкармливания первого выводка самки могут заботиться о втором без предварительной гормональной стимуляции. Полноценная материнская реакция на новорожденных здесь формируется в течение 48 ч.: главную роль играет тактильная стимуляция поверхности живота новорожденными, но не запах их различных экскретов. У мышей также действенны ультразвуковые сигналы детёнышей.
Инъекции вазопрессина непосредственно в мозг самцам прерийной полёвки стимулируют у них обнюхивание, груминг и чистку самок, что характерно также и для других видов с семейно-групповым образом жизни. Этот эффект отсутствует у одиночного вида — пенсильванской полёвки. Также экспрессия генов, ответственных за секрецию вазопрессина, у видов первой категории увеличивается после спаривания с самками, а также после рождения детёнышей, под влиянием контактов с ними.
Социальная релятивность тестостерона наиболее известна и подробней всего изучена. В целом у грызунов с семейно-групповым образом жизни и заботой самцов о потомстве концентрация тестостерона позитивно связана с проявлением родительского поведения. Однако то же самое обнаруживается у рыжей полёвки — вида с системой агрегаций индивидуальных участков обитания и неустойчивостью связей партнёров. У условно-одиночных видов и серой полёвки (вида со слабо консолидированными семейными группами), напротив, экспрессия родительского поведения самцов негативно связана с секрецией тестостерона в семенниках. У самцов же степной пеструшки (эта же категория видов) эти два показателя не коррелируют вовсе.
Как и у других гормонов, влияние тестостерона на родительское поведение самцов часто неустойчиво и/или противоречиво (не приобретает определённости без учёта социальных факторов и другого контекста). У домовых мышей уровень секреции тестостерона повышен, если при внутриутробном развитии их эмбрионы располагались в матке между эмбрионами-самцами. И, наоборот, если они были между эмбрионами-самками, уровень тестостерона снижался, и эти самцы отличались более феминизированным поведением, в т.ч. в отношении детёнышей. Однако реакция самцов домовой мыши в стандартном тесте, когда они или убивали предъявленных детёнышей, или затаскивали их в укрытие, не зависела от уровня содержания тестостерона. При любой концентрации тестостерона негативное отношение к детёнышам и детоубийство у самцов домовой мыши подавляются после спаривания с самками и т. д.
Отцовское и материнское поведение грызунов имеет общую нейрофизиологическую основу (в отличие, например, от агрессии). Одни и те же гормоны, активирующие определённые центры головного мозга самок и самцов, должны стимулировать одинаковые реакции, связанные с заботой о потомстве. Экспериментально доказано, что нейроэндокринные механизмы активации материнского поведения имеются и у самцов. В том числе кастрированные самцы крыс, в норме не проявляющие какой-л. заботы о детёнышах, делаются столь же заботливыми, как самки, после подкожной имплантации эстрадиола, эстрадиол-бензоата и прогестерона. Вообще, половой диморфизм родительского поведения грызунов определяется не различиями в церебральных структурах самцов и самок, а дифференциальной экспрессией генов (управляемой в первую очередь социальной стимуляцией «извне»).
Противоречивы данные о влиянии церебрального пролактина (синтезируется передней долей гипофиза и действует как медиатор) на материнское поведение крыс и мышей.
Отдельная составляющая родительского поведения — материнская агрессия. Она подчиняется тем же закономерностям. У мышей этот феномен выражен иначе чем у крыс: уровень агрессивности невысок на всем протяжении беременности и во время послеродового эструса, и достигает максимума на 2е сутки после родов. Иные и гормональные влияния: эстрадиол здесь материнскую агрессию подавляет, а прогестерон стимулирует демонстрацию угрожающих (у крыс — наоборот).
Тактильная стимуляция и другие социальные влияния на заботу о потомстве
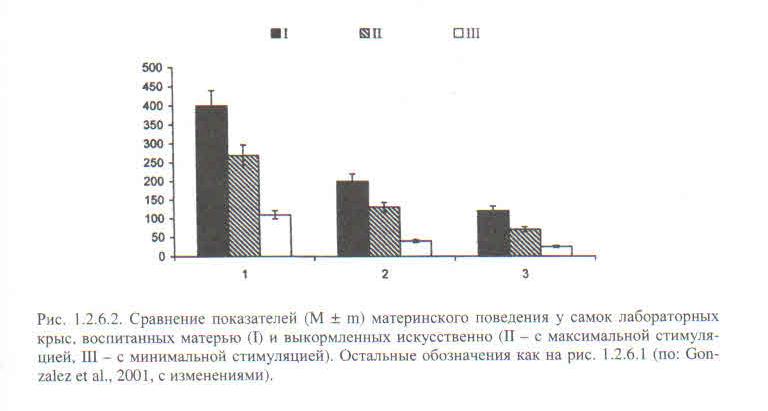
Среди прочего, на грызунах были поставлены опыты, аналогичные опытам Харлоу на обезьянах. Часть детёнышей лабораторных крыс выкармливали искусственно, лишая любой дополнительной стимуляции, часть (контроль) была с матерью, как обычно. Третья часть выращивалась как первая, но им компенсировали отсутствие материнского груминга поглаживанием мягкой кисточкой. К тому же их разделили на 2 подгруппы, с минимумом стимуляции (поглаживали только аногенитальную область дважды в сутки по 45 с.), в другой — максимум стимуляции (всё туловище 5 раз в сутки по 2 минуты, что, правда, сильно меньше, чем материнское грумингование, Громов, 2013). В зрелом возрасте самки всех 3-х групп принесли потомство, и с 6 по 10 день выкармливания последнего у них анализировали развитость материнского поведения по 4-м параметрам (рис.1.2.6.1 книги).
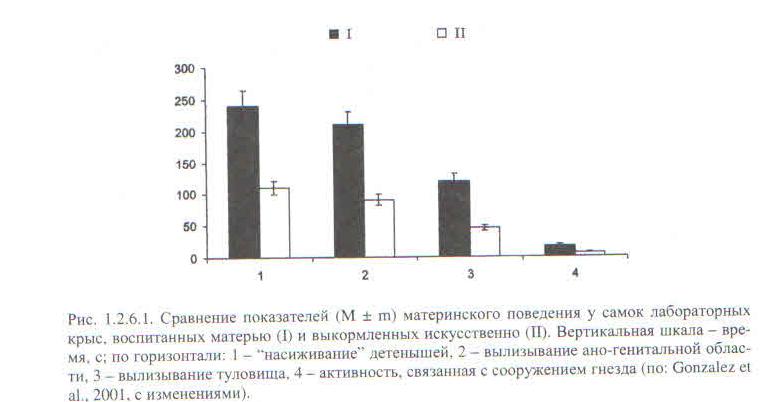
Вполне предсказуемо искусственно выкормленные самки демонстрируют существенный дефицит материнской заботы по всем направлениям (рис.1.2.6.2). Искусственная стимуляция до некоторой степени восстанавливает положение, причём степень компенсации пропорциональна интенсивности стимуляции кисточкой: максимальная примерно наполовину — две трети, минимальная — на треть. У видов с семейно-групповым образом жизни и облигатной заботой отцов о потомстве роль родительского груминга в формировании одноимённого поведения у детёнышей ещё выше. См. Gonzalez et al., 2001.
Сейчас интенсивно исследуется «передаточный механизм» от тактильной стимуляции родителей к развитости заботы о потомстве у детёнышей, в том числе между поколениями (Громов, 2013, разд.1.2.6). Показано, что сигналы тактильный рецепторов поверхности тела идут в соответствующие отделы гипоталамуса, модулируют гипоталамо-гипофизарную систему, влияя на формирование эмоциональных реакций и устойчивость к стрессу, что в конечном итоге влияет и на родительское поведение (не препятствует его полному проявлению, специфические формы которого заданы социальным влиянием). Другой «передаточный механизм» связан с изменениями функциональных связей в коре головного мозга, гиппокампе и других структурах лимбической системы.
Недостаток тактильной стимуляции сказывается на развитии средней преоптической области гипоталамуса и связанных с ней центров — миндалины, ложа терминальной пластинки и вентральной области покрышки среднего мозга. Либо (что более вероятно) на уровне активации рецепторов в процессе синтеза и высвобождения медиаторов, связанных со стимуляцией или подавлением материнского поведения: окситоцина, дофамина и ряда других. У «недолюбленных» детёнышей меняется метаболизм, увеличивается экспрессия с-fos протеинов в некоторых ядрах гипоталамуса, при её уменьшении в средней преоптической области. Выше, чем в норме, и уровень глюкортикоидов в плазме крови. Правда, эти исследования проведены на ограниченном круге видов (крысы, мыши, сирийский хомяк), среди которых нет наиболее интересных для нашей проблемы — с устойчивыми брачными связями и отцовской заботой о потомстве.
Другой вид социального влияния, интенсифицирующий экспрессию родительского поведения — это опыт помощничества у подрастающих детёнышей из старших выводков, особенно самок. Находясь вместе с младшими собратьями, они волей-неволей проявляют родительскую заботу: скучиваются с ними, вылизывают и затаскивают в гнездо, если те оказываются снаружи. Этот эффект сензитизации положительно влияет на ухаживание за собственными детёнышами: он отмечен как у крыс, так и прерийной полёвки, и максимален у самок первого вида в возрасте 22-24 дня.
Далее, в подавляющем большинстве случаев для запуска «должного» родительского поведения самцов и самок недостаточно одних гормональных влияний, необходима тактильная стимуляция (скучивание и груминг). Лишь благодаря ей они становятся заботливыми родителями и партнёрами. Эксперименты на лабораторных крысах показывают, что самки, выращенные собственными или приёмными матерями, уделявшими вылизыванию детёнышей и скучиванию с ними большое количество времени, став половозрелыми, также отличались высокими показателями материнской заботы в сравнении с самками, испытывавшими в детстве недостаток внимания матерей. Ещё резче этот эффект выражен у самцов.
Наконец, когда эффект социальных влияний и действие гормонов антагонистично, пересиливает первое. Что хорошо видно в экспериментах по пересадке детёнышей между парами близких видов грызунов, у одного из которых — семейно-групповой образ жизни с заботой самцов о потомстве, в другого — промискуитет, отсутствие постоянных пар и лишь материнская забота. Воспитание детёнышей видов второй группы в семьях первой даёт им в норме несвойственную тактильную стимуляцию от отцов (через скучивание с согреванием и вылизывание). Благодаря этому, повзрослев, «пересаженные» самцы сами заботятся о детёнышах, и ведут себя «несвойственным им» образом (пересадки от пенсильванских полевок к прерийным, McGuire, 1988) и от белоногих хомячков к калифорнийским (Bester-Meredith, Marler, 2003). То же самое зафиксировано в опытах с перекрёстным выращиванием между разными популяциями прерийных полёвок (штатов Канзас и Иллинойс). Родительское поведение у вторых было выражено больше, чем в первых; зверьки, выросшие в чужих семьях, особенно самцы, следовали стереотипу поведения приёмных родителей, особенно отцов, больше чем биологических (Roberts et al., 1998a).
И наоборот: самцы видов с семейно-групповым образом жизни, выращенные с недостатком тактильной стимуляции (монгольские песчанки, в течение 3-х поколений выращиваемые без самцов), оказываются худшими и родителями, и брачными партнёрами.
То же верно для внутривидового разнообразия стилей заботы о потомстве у самок, из млекопитающих наибольшего у грызунов и особенно у приматов. Оно воспроизводится в первую очередь «сигнальной наследственностью» и при вышеописанных пересадках стиль поведения соответствует таковому приёмных родителей, а не родных (Champagne, Meany, 2001; Крученкова, 2009).
Рецепторная концепция действия гормонов
Отдельное замечание: если формообразующий эффект социальных влияний автор блистательно игнорирует, даже когда это касается его группы (грызунов), рецепторную концепцию гормональных влияний он упоминает. Лишь для того, чтобы отвергнуть как «полную чушь» без каких-либо оснований. Ведь данные по сравнению родительского поведения близких видов грызунов, различающихся социальной организацией, прочностью социальных связей в группе (и/или брачных связей родителей в парах, трио или «коммунах»), и степенью участия самца в заботе о потомстве, показывают различия в первую очередь в плотности рецепторов в соответствующих отделах мозга. Концентрации гормонов и нейромедиаторов, включённых в запуск родительского поведения, у таких видов различаются существенно меньше.
Так, у видов со слабыми парными связями (лабораторные крысы, домовые мыши) родительское поведение самцов «включается» лишь в ходе сензитизации, т. е. постоянных или, как минимум, регулярных контактов с детёнышами. Важно отметить, что для всех вдов показана одинаковость влияния гормонов и/или нейромедиаторов на родительское поведение у обоих полов. Сензитизация может происходить в очень раннем возрасте, скажем, уже по окончании периода молочного вскармливания, когда самка воспитывает 2 разновозрастных выводка. Если контакт с детёнышами недостаточно регулярен, у этих видов высок риск инфантицида со стороны самцов.
Аналогичная сензитизация описана и у самок. Так, самки лабораторных крыс, не имеющие родительского или сексуального опыта, в тестовой ситуации чаще всего отвергают чужих новорожденных детёнышей или убивают их. Но если они регулярно контактируют с ними, то через 4-7 дней избегание сменяется устойчивым проявлением родительской заботы. Поведение самок меняется именно воздействием комплдекса запаховых, акустических и тактильных стимулов от детёнышей. В организме девственных самок сензитизация производит физиологические изменения, аналогичные идущим при беременности, в том числе интенсифицирует два важнейших гормональных влияния — эстрадиола и пролактина.
У видов с семейно-групповым образом жизни родительское поведение самцов «включается» автоматически. Инфантицид здесь подавлен, отцовская забота формируется при участии нейроэндокринных изменений, связанных с формированием устойчивых социальных связей (о них см.Громов, 2013). После приобретения отцовского опыта самцы этих видов демонстрируют родительскую заботу в любой ситуации и независимо от репродуктивного состояния самки. При этом у них растёт концентрация пролактина в крови и снижается содержание андрогенов (Громов, 2013, раздел 1.3.3).
Как и у самок, за стимуляцию родительского поведения самцов ответственна средняя преоптическая область гипоталамуса (СПО), дополнительные обонятельные луковицы и средняя область миндалины. Повреждение СПО отражается на реализации этого поведения негативно, имплантация эстрадиола непосредственно туда — позитивно.
Межвидовые различия в реализации родительского поведения самцов, у грызунов очень резкие, опосредованы столь же резким различием в концентрации специфических рецепторов в разных частях СПО. Так, исследовали распределение вазопрессиновых рецепторов V1a СПО у горной и прерийной полёвки. Первый вид — условно одиночный, о потомстве заботятся только самки; второй с постоянными брачными связями в парах или трио, облигатной отцовской заботой. V1a рецепторы в ядрах латеральной перегородки и диагональной связки ЛПО у второго вида распределены гуще и плотней, чем у первого, что понятным образом связано с развитостью родительского поведения самцов (Young et al., 1999). Однотипные различия в концентрации V1a выявлены у гомологичной пары видов: калифорнийский хомячок — белоногий хомячок (Bester-Meredith et al., 1999).
Распределение рецепторов вазопрессиина также варьирует у самцов одного вида в зависимости от их репродуктивного статуса. Самцы прерийной полёвки, живущие в паре с самками и вместе с ними воспитывающие потомство, демонстрируют большую плотность распределения V1a в латеральной перегородке, чем не имеющие сексуального опыта и/или живущие изолированно.
В соответствующих парах видов различается и распределение рецепторов окситоцина в структурах головного мозга. У видов с семейно-групповым образом жизни их концентрация повышена в прилежащем ядре и предлимбической коре, у их условно одиночных родичей — в латеральной перегородке. Инъекции антагонистов окситоцина в данные области блокируют поведение, связанное с формированием семейных пар у прерийной полёвки (но не родительское, влияние окситоцина на которое скорей косвенное, Young et al., 2001).
Вывод отсюда может быть только один: если не специфичность эффекта, то возможность его проявления (говоря генетическим языком, если не экспрессивность, то пенетрантность) обеспечивается именно плотностью специфических рецепторов в «нужных» отделах мозга. Она также меняется под действием социальных влияний.
Соответственно, прежде чем отрицать, стоило бы поинтересоваться.
Методологические изъяны
И последнее замечание — методологическое. Пользоваться «бритвой Оккама» надо с умом, чтобы «не отрезать лишнее», в смысле неудобные факты, не укладывающиеся в твою концепцию (что автор с успехом и проделывает). Сущности не следует умножать лишь «сверх необходимости», и главное здесь — не «экономия мышления», а определение соответствующей «необходимости», в смысле достаточной сложности объяснения. Тут надо пользоваться принципом минимально необходимого разнообразия Эшби: сложность конструкций теории, применяемых для объяснения определённой реальности, не может быть ниже сложности содержания этой реальности, иначе нечто существенное обязательно будет упущено, в том числе нельзя будет решить проблему, ради которой создаваласьэта теория.
Иными словами, объясняющее (explanans) в концептуальном отношении не может быть проще объясняемого (explanandum), но сложнее быть тоже не должно. Иначе производимые объяснения будут недопустимо упрощены и/или упущены некоторые критически важные факторы, и объясняющий будет склонен уклоняться от рассмотрения всех относящихся к делу фактов (ибо они «высвечиваются» лишь подходящей теорией). Примитивизм объяснений автора и ошибки интерпретаций во многом — результат игнорирования этого правила.
Заключение
Из вышесказанного следует, что организатором видоспецифического поведения «должного, и в должные моменты активности» являются социальные влияния. Гормоны и другие эндогенные факторы лишь «поддерживают» (или ослабляют) уже выбранное и реализующееся поведение, а их уровень, сила эффектов и пр. характеристики прямо зависят от успеха предшествующей активности, уже реализованного поведения. Это верно для всех «социальных» видов позвоночных, но максимального развития эта тенденция достигает у приматов и человека. Именно в этой филогенетической линии социальные влияния на поведение (агрессивное, родительское, брачное) максимально усиливаются, гормональные — последовательно ослабляются (Крученкова, 2009).
Научный дефект всех биологизаторских концепций (в «Стой, кто ведёт!» представленных как единственные — без рассмотрения альтернатив) заключается как раз в «непримечании слона», уклонении от рассмотрения вышеописанных данных. Другой важный момент — популяризуемые книгой представления об организации поведения не работают на той самой группе — грызунах, которой он занимается. Если бы не идеологические шоры, он бы это увидел, сопоставив собственные данные (более чем интересные в соответствующей частной проблеме) с другими исследованиями, показывающими как разные стратегии преодоления стресса вплетаются в общую организацию поведения и участвуют в формировании индивидуальности особи.
Поэтому биологизаторство (или, мягче, примат эндогенных влияний «изнутри» над социальными извне в организации поведения) не только плохая теория, но и идеологическая позиция. Реакционная, потому что не позволяет рассматривать все относящиеся к делу факты, препятствует прогрессу исследований… — не человека, нет, высших позвоночных как социальных существ, хотя «анатомия человека — ключ к анатомии обезьяны», и к социальности это тоже относится.
В случае с человеком же эта как бы научная позиция мешает понять действительную роль «биологии» в общественной жизни нашего вида и, воспринятая публикой, оказывается отличным оправданием существующих форм угнетения, представляя различия, «сделанные» социальной стратификацией и дискриминацией «низших» как «естественные».
Увы, «Стой, кто идёт», как и прочие произведения Дм.Жукова, написан в том числе и для этого, помимо собственно популяризации его теорий с присущими им огрехами. Отсюда присутствующие в произведении натяжки, передёргивания, умолчания, прямые искажения фактов, и другие приёмы дезориентации читателя (подробно рассмотренные в известной книге Сергея Поварнина «Спор. Об искусстве спора»). Часть их уже отмечена рецензентами, другие обсуждаются ниже.
Наконец, исключительно негативно при чтении книги воспринимается безапелляционность утверждений автора. В настоящей научно-популярной литературе (столь разной тематики, как «Язык тела: природа и культура» М.Л.Бутовской и «Обман в науке» и «Вся правда о лекарствах» Б.Голдакра) на каждое утверждение есть ссылка на оригинальное исследование, позволяющая вдумчивому читателю не принимать на веру.
Утверждения же Д.А.Жукова тем резче и бездоказательней, чем хуже подкреплены фактами: это общечеловеческий грех, встречающийся и у других, но здесь больно уж ярко выраженный. Вполне закономерно в таких утверждениях он попадает «в молоко»; в первую очередь касательно истории, культуры, психологии и других гуманитарных дисциплин, но и в собственной специальности тоже.
***
Честно скажу: книгу не дочёл до конца, осилил лишь треть. Но ведь чтобы понять, что суп прокис, не надо есть всю тарелку, понюхать достаточно?
Список литературы
Аронсон Э., Уилсон Т., Эйкерт Р., 2008. Большая психологическая энциклопедия. Психологические законы человеческого поведения. Спб.: Прайм-Еврознак. 560 с.
Бардин А.В., 1982. Бюджеты времени и энергии большого пёстрого дятла Dendrocopos major major (L.) в зимний период // Тр. Зоол. ин-та АН СССР. Т.113. С.45-57.
Бардин А.В., Марковец М.Ю. Скорость расхищения запасов синиц//Экология. 1990. №6. С.48-53.
Благосклонов К.Н. Экспериментальный анализ территориального поведения птиц// Адаптивные особенности и эволюция птиц. М.: Наука, 1977. С.128-134.
Гороховская Е.А., 2001. Этология: Рождение научной дисциплины. Спб.: Алетейя. 224 с.
Крученкова Е.П., 2002. Принципы отношений мать-детёныш у млекопитающих. Автореф. и рукопись дисс. доктора биол.наук. М. 409 с.
Крученкова Е.П., 2009. Материнское поведение млекопитающих. М.: URSS. 208 с
Приклонский С.Г., 1962. Отлов больших пёстрых дятлов на кузницах// Тр. Окского зап-ка. Вологда. Вып.4. С.427-430.
Рябицев В.К., 1995. Механизмы и результаты межвидовых территориальных отношений у тундровых поморников [на англ.яз. Ryabitsev V.K. Patterns and results of interspecific territorial relations in tundra skuas]// Русский орнитол. журнал. Т.4. №1/2. С.3-12.
Северцов А.С., Шубкина А.В., 2015. Эволюционная роль хищных млекопитающих и механизмы её реализации// Журн. общей биологии. Т.76. №5. С.355-368.
Hailman J. P. 1967. The ontogeny of an instinct: the pecking response in chicks of the laughing gull (Larus atricilla L.) and related species // Behav. Suppl. Vol. XV. E.J. Brill, Leiden. 196 pp.
Hailman J. P., 1968. Visual-cliff responses of newly-hatched chicks of the laughing gull Larus atricilla// Ibis. Vol.101. P.187-200.
Hailman J. P. 1971. The role of stimulus-orientation in eliciting the begging response from newly-hatched chicks of the laughing gull (Larus atricilla)// Anim. Behav. Vol.19. P.328-335.
Krebs J., Biebach H. Time-place learning by garden warbler (Sylvia borin)// Ethology. 1989. Vol.83. №3. Р.248-256.
Sherry D. Food storing in the Paridae// Wilson Bull. 1989. Vol.101. №2. Р.289-304.
Sherry D., Vaccarino A. The hippocampus and memory for food caches in black-capped chickadees// Behav. Neurosci. 1989. Vol.103. №3. Р.308-318.
Vander Wall S., Balda R. Ecology and evolution of food storage behavior in conifer-seed-catching corvids.// Z. Tierpsychol. 1981. Bd.56. S.217-242.
Vallotigara G., Zanforlin M. Place and object learning in chicks (Gallus gallus domesticus)// J. Comp. Psychol. 1989. Vol.103. №2. Р.201-209.
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Красивое доказательство «семантичности» демонстраций у птиц
Красивое доказательство «семантичности» демонстраций у птиц Используются ли «понятия» в коммуникации животных?
Используются ли «понятия» в коммуникации животных? Социальный ранг и психика. Синантропные грызуны в «жилой комнате»
Социальный ранг и психика. Синантропные грызуны в «жилой комнате» Об истории, культуре и современных проблемах коми. Часть 2.
Об истории, культуре и современных проблемах коми. Часть 2. Социальные функции нетерпимости к атеизму в США
Социальные функции нетерпимости к атеизму в США Два механизма «копирования поведения»
Два механизма «копирования поведения» Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции
Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции
