
Сигналы-символы, изоморфные «понятиям» в коммуникации животных.
Сигналы-символы: «сухой остаток» после скепсиса и перепроверки
Содержание
Читаю обзоры о функционировании сигналов-символов (referential signals) в коммуникации разных видов позвоночных: при привлечении к корму у птиц и млекопитающих; при предупреждении об опасности у птиц; в разных контекстах общения у приматов и млекопитающих вообще. Напомню: это эволюционно наиболее продвинутая (и самая редкая) категория сигналов. В противоположность более частным сигналам-стимулам (классическим релизерам, motivational signals) они передают информацию в символической форме (отсюда название). Поэтому соответствующий сигнал «действует только своим значением», и независим от каких-либо контекстуальных «подсказок» от поведения продуцирующей его особи или изменения ситуации в «общем поле зрения» обоих участников взаимодействия.
См. спектрограммы сигналов домашней курицы: 1 – «опасность с воздуха», 2 – «опасность с земли», 3 – «пищевой крик». Обратите внимание на альтернативность форм разных типов сигналов и отсутствие переходов между ними.
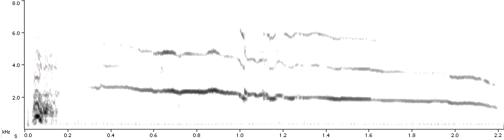
1.
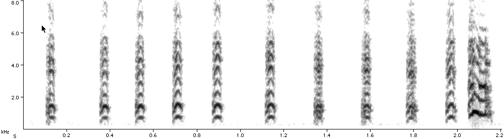
2.

3.
А вот как показывается его действие.
«Ставили две серии опытов, в контрольной серии (половина всех повторностей, выбранная случайным образом) проигрывание пищевого крика и крика «опасность с земли» предваряли выдачей зерна («food trials»), птице давали возможность кормиться. Соответственно, в первом варианте опыта кормлению предшествует проигрыванию пищевого крика (заполненные кружки), во втором — кормлению предшествует проигрыванию крика «опасность с земли» (пустые кружки), в обоих случаях сигнал мотивирован прошлым состоянием животного.
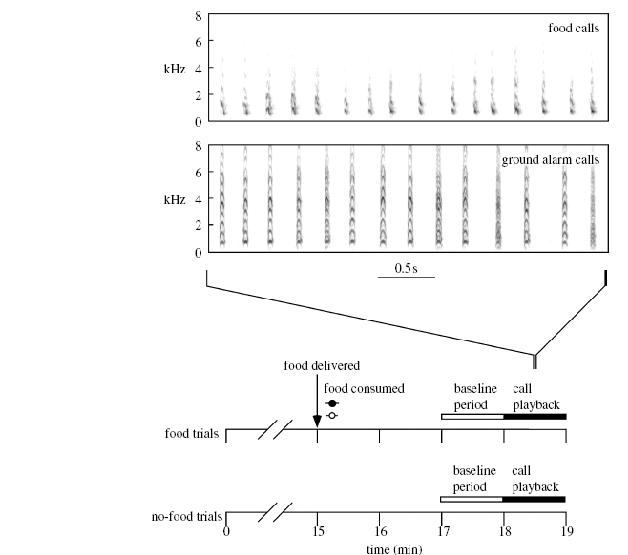
Исследование коммуникативного эффекта пищевых сигналов домашних кур – информирование или манипуляция?
В экспериментальной серии те же крики проигрывали в том же порядке, но им не предшествовала выдача зерна («non-food trials»), то есть птицы ничем не мотивированы «правильно» реагировать на первый и второй крик. Реакцию на сигнал оценивали по интенсивности поискового поведения, при котором птица характерным образом меняет позу, опуская голову к земле, так что оказывается неспособной сканировать окружающее пространство на предмет потенциальной опасности (см.рисунок). Поисковое поведение после сигналов сравнивали с таким же поведением в предшествующий период (baseline period) и оценивали приращение либо уменьшение.
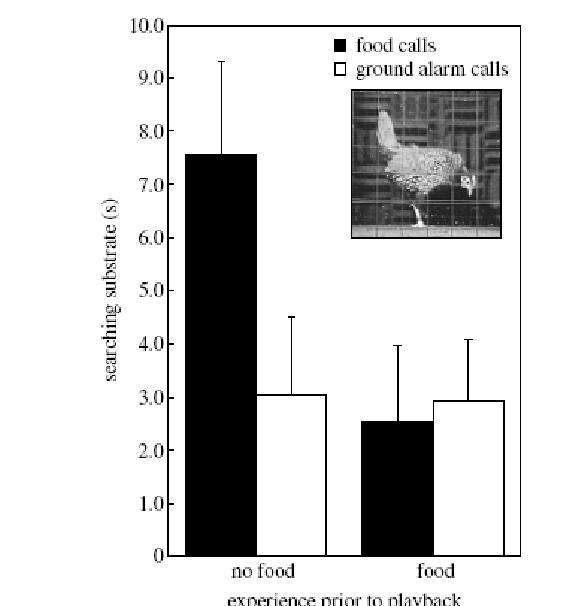
Чёрные столбики – изменения интенсивности поискового поведения после проигрывания пищевого крика, белые – после проигрывания крика «опасность с земли». Фото – поза поиска на субстрате, принимаемая в ответ на пищевой крик именно в случае отсутствия подкрепления (то есть, когда ничего в реальности не мотивирует искать корм, кроме данного крика).
Адекватность восприятия выражается выдачей специфической реакции на сигнал даже у немотивированной особи, которая из наблюдения ситуации вокруг не может получить независимое подтверждение о необходимости действовать определённым образом – так, как «сообщают» сигналы, и приходится «полагаться» именно и только на последние. Вышеописанный эксперимент подтверждает подобное «полагание» в отношении пищевого крика. Предшествующие исследования тех же авторов показали аналогичное полагание и в отношении обоих «тревожных криков» — «опасность с земли» и «опасность с воздуха» (Evans et al., 1993a- b; Evans, Marler, 1995).
«От стимула к символу. Сигналы в коммуникации позвоночных животных«
Система отличий тех и других типов сигналов по форме и функции описана здесь; я её свёл в таблицу:
|
Сигналы-стимулы (Motivational signals) |
Сигналы-символы (Referential signals) |
|
|
Форма |
Формы разных демонстраций в последовательности связаны переходными элементами |
Формы взаимно альтернативны, между последовательными элементами чёткая грань |
|
«Значение» |
Сигналы состояний — показывают уровень стресса, вызванного проблемной ситуацией взаимодействия, и уровень мотивации (агрессиии, страха, половой), отражающий готовность «проблему» решать |
- Сигналы ситуаций; «именуют» логически альтернативные категории объектов, значимых в данном контексте общения; |
|
Действуют ли как стимулы |
Да, разной степени специфичности |
Нет |
|
Функционирование как сигналов |
- «cмазана» переходными элементами — не выражена, так как изменения контекста влияют на означаемое сигнала — уровень мотивации — не выражена, так как обстоятельства предъявления этих стимулов влияют на уровень мотивации — сигнал выражает разные состояния внутреннего мира животного (уровни мотивации). |
- специфичность формы при продуцировании сигнала (англ. production specifity) |
После первоначального ажиотажа, когда эту категорию сигналов хотели найти везде и всюду, естественным образом пошла противоположная волна скепсиса. С одной стороны, она подстёгивалась крахом очень сильных надежд найти их у антропоидов и другими последствиями третьего, самого лукавого вида антропоморфизма — антропоморфизма противопоставления.
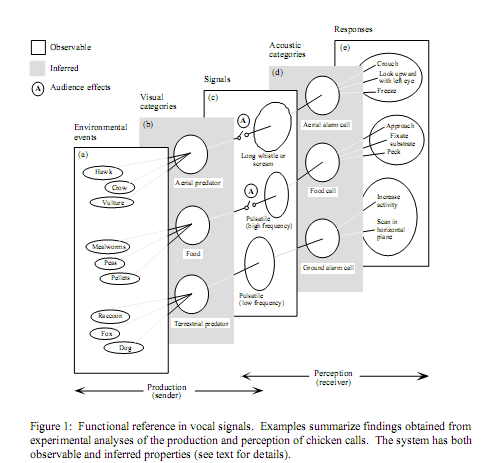
Надежды подогревались стремлением найти в коммуникативных системах «типа верветок» хоть плохонький, но «такой же» знак, как в человеческой коммуникации — с намеренным информированием, ментальными репрезентациями. Где хотя бы в одном, пусть узком аспекте, мышление и «речь» (в смысле, их коммуникативные сигналы) были бы «сцеплены по — человечески», когда речь интеллектуальна, а мышление речевое. Эти надежды воплощает правая часть схемы механизмов восприятия коммуникативных сигналов животных из обзора Ch.Evans’а (1997), показывающая тогдашнее понимание действия (или отреагирования) сигналов-символов — так, как будто определение референциальные к ним применимо на том же основании, что к речевым знакам.
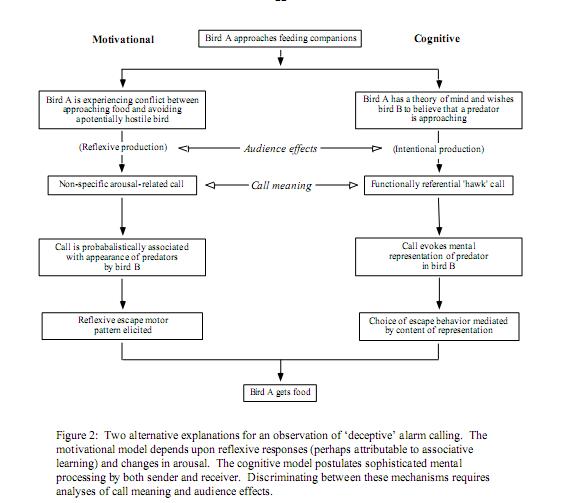 Сегодня мы точно знаем, что это неверно: в эффекте сигналов-символов «нет ничего «когнитивного», только инстинкт». Правда, некоторые надежды остались; так, авторы одного из обзоров пишут «действительно ли эти сигналы действуют так же символически, как в человеческом языке, где условные звуки отражают ментальные репрезентации внешних объектов, созданные мыслью у нас в голове, остаётся неясным, поэтому использование термина «функциональная референтность» несколько противоречиво».
Сегодня мы точно знаем, что это неверно: в эффекте сигналов-символов «нет ничего «когнитивного», только инстинкт». Правда, некоторые надежды остались; так, авторы одного из обзоров пишут «действительно ли эти сигналы действуют так же символически, как в человеческом языке, где условные звуки отражают ментальные репрезентации внешних объектов, созданные мыслью у нас в голове, остаётся неясным, поэтому использование термина «функциональная референтность» несколько противоречиво».
На деле его данные, как и все прочие, показывают вполне ясно, что
-
эти сигналы действуют вполне символически, «именуя» дискретные типы важных объектов или значимых ситуаций во внешнем виде животного направленно, точно и независимо от контекста (таблица);
-
«именование» происходит так хорошо и точно, что животные «доверяют» этим сигналам больше чем непосредственному виду хищников или корма, чем дают интересный изоморфизм к управляемости словом людей;
-
однако «намеренность издавания» сигнала, «осознанность его значения» и, соответственно действия, и связывающие их ментальные репрезентации здесь отсутствуют — хотя обезьяны или, например, вороны, прекрасно осознают важные социальные ситуации, и действуют намеренно и направленно, чтобы менять их в свою пользу.
То есть, как и предполагал Л.С.Выготский, у животных мышление и «речь» разобщены; соединение их (в виде некого «дополнения» или «изменения» значений видовых сигналов собственным опытом использующего их животного) делается всегда «на живую нитку» и крайне ограничено даже у столь умных существ, как мартышки Дианы. Поэтому, кстати, название «сигналы-символы» лучше кальки с англ. referential signal — не наталкивает читателя на ложную мысль о присутствии «человеческого» обеспечения референтности, здесь оно иное, инстинктивное.

Мартышки Диана Cercopithecus diana
Наряду с чёткими отрицательными результатами (совокупность которых более менее чётко очерчивает «область использования» таких сигналов в отношении таксономии и/или контекстов общения) было выдано на-гора много сомнительных работ, требующих перепроверки.
Пример проверки: какой тип сигналов используется?
Один из примеров проверки — связанная с хищничеством вокализация у североамериканской рыжей белки: кому адресованы тревожные крики и как они работают?
Этот вид издаёт громкие, хорошо слышимые тревожные крики, но использует их эпизодически, и мнения исследователей о функции криков противоречивы (поэтому статья, где они специально исследуются, названа осторожно — «вокализации, ассоциируемые с появлением хищников», и это правильно). Авторы изучали примерно 47 меченых белок в течение 3-х сезонов в предгорьях канадских Скалистых гор. Фиксировали издавание соответствующих криков в ответ на появление реальных хищников, наземных, воздушных и/или змей, и в опытах с макетами, имитирующими появление и приближение хищника. тестировали 3 рабочие гипотезы, соответствующие наиболее распространённым концепциям о функциях тревожных криков — что это специальные предупреждения об опасности либо родственникам, либо брачному партнёру, либо владельцам соседних территорий, с которыми у него прочные связи.

Рыжая белка Tamiasciurus gudsonicus
Все три гипотезы подтверждаются плохо (только частично и подтверждение неустойчиво). Так получается потому, что на деле это сигнал совершенно иного рода — это сигнал хищнику, что он обнаружен, животное настороже (что, как показано многажды, его «обескураживает», останавливает, заставляет отказаться от атаки и пр.). Соответственно, изменчивость параметров крика отражает уровень стресса и страха, возникающий у белки при обнаружении опасности. Т.е. это сигналы — стимулы, а не сигналы-символы, как было бы, если бы они действительно информировали о характере опасности. Что вполне понятно — белки в целом примитивнее сусликов, у анцестральных видов сусликов вокализации, ассоциированные с появлением хищников, тоже представляют собой сигналы-стимулы.
Как было показано Шели и Блюмштейном (2005), сигналы-символы развиваются независимо в разных филогененетических ветвях наземных беличьих (и, шире, дневных социальных грызунов), лишь когда социальность разовьётся настолько, что станет необходимым информирование многих соседей о характере опасности. Когда крик слышат лишь некоторые (поселения недостаточно плотные, недостаточно развито внимание зверьков к вокализациям друг друга), выгодней только воздействовать на хищника «энергией стресса», ретранслируемой через крик. И действительно, эти белки — одиночный вид, охраняющий территории, на которых они создают запасы корма, с активным противодействием попыткам соседей их растащить. Хорошо документированный пример такой эволюции «от стимула к символу» — рост референциальности значений и одновременно дискретности/противопоставленности друг другу форм тревожных сигналов в одной из ветвей американских сурков рода Marmota.
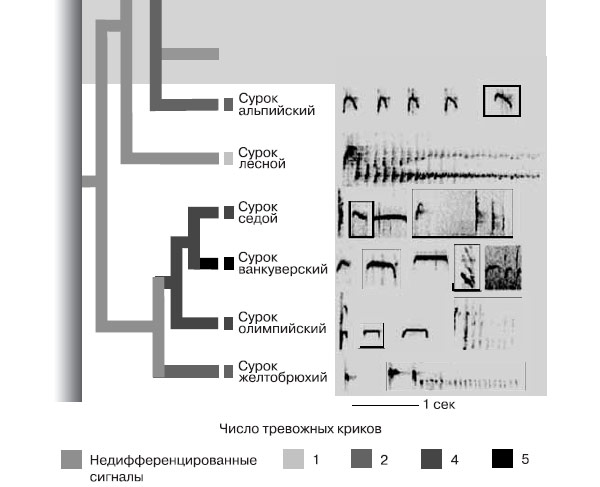
Формирование дифференцированной системы сигналов предупреждения об опасности в филогенетическом ряду разных видов сурков. По: Blumstein, 2007. Прямоугольники — различные типы сигналов: видно увеличение их числа при переходе от более древних видов к молодым. Это позволяет сделать вывод о переходе от недифференцированной системы сигнализации у более анцестральных видов к дифференцированной у более продвинутых. Источник
Вполне понятно, что одна и та же вокализация (именно та, что исследуется в статье) выражает у них «стресс и возмущение» от воровства (ретрансляция которого в крике останавливает вора, побуждает отказываться от таскания чужих шишек), и «стресс и страх» при появлении хищника. То есть не всякий крик, структура которого закономерно меняется в случаях появления разных хищников, является сигналом — символом, как часто предполагают этологи вследствие «моды» на сигналы-символы (referential signals). Они могут отражать разную степень страха, обычно связанную с разными видами хищников, когда те придерживаются присущей им скорости крейсирования и способа появления в поле зрения жертвы. Но если характер появления и скорость сближения с жертвой пернатого или наземного хищника изменяются, отклоняясь от видового стереотипа (зверь резко и неожиданно выскакивает из укрытия, пернатый хищник сидит на земле или спокойно парит в небе), то крик изменяется сообразно изменившейся степени стресса и страха. У видов с действительными сигналами-символами, такого не бывает никогда — разные виды хищников «обозначаются» разными сигналами (ещё и альтернативными друг другу по форме, без связывающих их переходов) независимо от того, как движутся и что делают.
Исходно эта вокализация выражала стресс и агрессию резидента при попытках растаскивания его запасов, а затем тот же самый сигнал мотивации по понятным причинам смог работать и в прочих контекстах.
Перечисленные обзоры дают нам «сухой остаток» достоверного использования сигналов-символов, подтверждённого разными способами и/или авторами. Скажем, см. какие сигналы и когдак используются в разных контекстах изученными видами млекопитающих.
Сигналы-символы в коммуникации: использование «понятий вслепую»?
Помимо прочего, вышеописанный материал важен тем, что отвечает на вопрос заголовка. Классические инстинкты «вслепую» выполняют логическую операцию «если, то… иначе…», которую разумное существо производит сознательно. В обоих случаях критически важно распознавание образов, благодаря которому «то, что нужно» отличается от «не того» или «всего прочего» (фигуры друг от друга и от фона).
В «образе мира» животного (иначе — умвельт) первое и второе представляет «фигуры», третье — «фон». Обычно это специфические формы демонстраций, которыми обмениваются особи в данном взаимодействии и континуальный поток поведения, куда они «вкраплены», из которого «выделяются», как льдины из шуги в ледостав. Реже — значимые объекты внешнего мира, вроде различных хищников и видов корма, однако распознаваемых тем же способом
— детекция по специфическим характеристикам формы, выделение «на фоне» изменчивой и континуальной «прочей» реальности, категоризация (воздушные vs наземные хищники vs корм как у курицы) и
- означивание сигналами, формы которых взаимно альтернативны, так что не спутаешь. См. «Ещё про расшифровку «языка животных», Вступление и Ч.2.
Подчеркну, что инстинкт всегда и везде срабатывает в ответ на характерную форму «ключевого раздражителя» (или образ, устойчиво воспроизводимый в телодвижениях демонстратора и предъявляемый партнёру), но не на эффекты воздействия в виде меняющегося внутреннего состояния после «укола» им, «толчка» или «давления».
Контроль переменных состояния при помянутом распознавании образов показывает, что и в социальном взаимодействии, и в реакции на внешнего мира животные сперва выделяют значимый объект по параметрам, «кодирующим» его форму и отделяющим от других объектов того же ряда (форм демонстраций данного контекста, других категорий хищников и пр.) и от «фона», и отреагирует его должным образом — это как раз инстинктивный ответ на сигнал. Что видно лучше всего на недавно расшифрованной системе коммуникации коростелей Crex crex и давно — предупреждения об опасности у американских черноголовых гаичек Parus atricapillus.
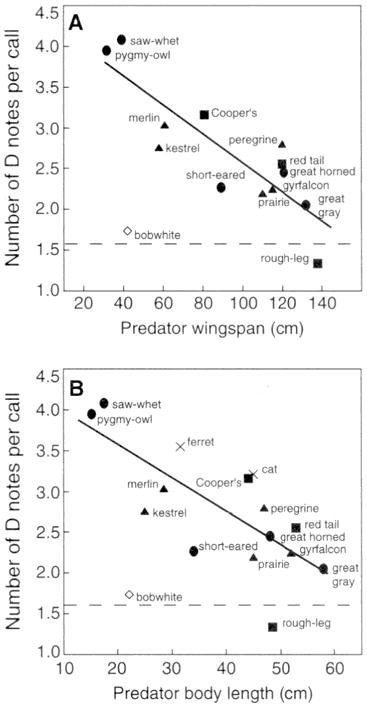
Кодирование информации о степени опасности в тревожных криках американских черноголовых гаичек. По: Templeton et al., 2005. Ордината – среднее количество слогов «Dee» тревожного крика как функция размаха крыльев возможных хищников в см. (А; регрессия y = 4.4 — 0.02x; r2 = 0.512, P < 0.0001) и как функция размера тела возможныхпернатых и наземныххищников (В, регрессия y = 4.4 — 0.4x; r2 = 0.361, P < 0.0001). Разные группы хищников показаны разными символами: • — совы, х – млекопитающие, маленький чёрный квадрат – ястреба, маленький чёрный треугольник – сокола. Пунктирная линия – интенсивность вокализации в ответ на контроль (виргинский перепел Colinus virginianus). Примечание. Параметр А отражает «размер» и «ловкость» хищника одновременно.
Лишь потом, при точности первого и успешности второго внутреннее состояние исполнителя изменяется (в т.ч. концентрации тестостерона, кортикостерона и других существенных в данной ситуации молекул-посредников). Фактически, если воспринимаемые формы демонстраций воспроизведены в телодвижениях исполнителя достаточно стереотипно и дифференцированно от других форм (демонстраций, предшествующих и последующих в данном взаимодействии), мотивационные изменения «внутри» реципиента происходят «сами собой», см. про действие сигналов-стимулов.
Благодаря сходному механизму мы различаем фонемы и лексемы независимо от интонации сказанного, от того, говорят их или поют, или даже когда говорят с акцентом. А если не различаем, не понимаем сказанного, заменяем восприятие фантазией. Соответственно, каждое коммуникативное взаимодействие суть кооперативная реализация инстинкта (одного из N видовых), невозможная в одиночку. Она воплощается в «цепочках» последовательных демонстраций, которыми особи обмениваются во взаимодействии (по определённым «правилам», стремясь его выиграть); каждая демонстрация — сериальный элемент соответствующего инстинкта, вроде сегмента у нереиса, видовая этограмма — репертуар демонстраций это как бы «конструктор» из разнотипных «сегментов». См. «Про выигрышные стратегии в коммуникации животных«. Верно и обратное — инстинкт всегда коммуникативен.
«Так, у цыплят домашней курицы специфическую реакцию затаивания и бегства вызывает не только крик «опасности с воздуха», но и стилизованное изображение «ястреба», – силуэт с широкими крыльями, короткой шеей и длинным хвостом, если его двигать над цыплятами. Реакция на соответствующий стимул впервые была исследована еще в классических экспериментах Лоренца-Тинбергена и повторена в опытах Ch.Evans et al. (1993a), когда использовали компьютерную анимацию движения модели «хищника» с разной скоростью и на разной высоте. В обоих случаях оборонительная реакция следовала, когда модель напоминала «ястреба», и отсутствовала, когда модель двигали в противоположную сторону (тогда она напоминает «утку» – длинный «хвост» интерпретируется как «шея»). Наряду с бегством при предъявлении «ястреба» цыплята также издавали крик «опасность с воздуха» (именно и только этот), но лишь в том случае, когда находились в группе, и не вокализировали в одиночку (Evans et al., 1993b)…
В других экспериментах у цыплят вырабатывали условную связь между нейтральным стимулом (красный свет) и появлением корма. После выработки условного рефлекса красный свет вызывает пищевой сигнал, свет иной длины волны – нет. Другой пример: у цыплят существует значительная степень дифференциации функций между «системой левого глаза» и «системой правого глаза» при обработке зрительных стимулов. «Система левого глаза» решает в основном задачи пространственной локализации удаленных объектов, «система правого глаза» настроена на категоризацию объектов, например распознавание корма. Когда цыплятам проигрывали крик «опасности с воздуха» они пытались фиксировать соответствующий объект именно в верхней части поля зрения и именно при помощи левого глаза (Evans, 1997)».
«От стимула к символу», op.cit
Инстинктивный акт же — функциональная единица процесса: единичный ответ реципиента на демонстрацию данной особи, действующую как релизер и/или передающую информацию. А организация, выявляемая в «цепочках» — временнАя и по порядку следования разных демонстраций репертуара — выступает как «синтаксис» взаимодействия с информационной точки зрения и одновременно как форма соответствующего видового инстинкта с т.з. «морфологии поведения», как её представляли классические этологи.
Иными словами, ритуализированные демонстрации отличает примат формы над действием, и распознавания образов (детектирующих первое) над изменением состояния (после второго) даже когда (и особенно когда) они выступают как классические релизеры.
Сегодня мы знаем, что это особенно верно для так называемых сигналов-символов (referential signals), открытых в конце 1980-х и активно исследуемых в 1990-е и 2000-е годы. Они информируют реципиента о дифференцированных категориях значимых объектов его внешнего мира, т. е. это сигналы ситуаций, а не состояния. Среди них — разные типы хищников, виды корма, разные шансы завершить конфликт победой или поражением большего (меньшего) «качества» и пр.
Они столь же инстинктивны, как сигналы-стимулы, т.е. продуцируются без намерения информировать реципиента и считываются им без «понимания« их значений. И, вообще, без ментальных репрезентаций, опосредующих использование знаков и символов у нас или language-trained apes — т.е. существ, у которых инстинкты отсутствуют.
Достаточно лишь неспецифичной готовности продуцировать «должные» системные сигналы в «должных» ситуациях взаимодействия с другими, социальными компаньонами, а при восприятии сигналов других — столь же «автоматически« (и вероятностно, а не детерминистически) корректировать собственное поведение на основании сигнальной информации, и тем самым ретранслировать этот общий сигнал по коммуникативной сети всего сообщества. Почему выгодны «подглядывания« и прочие формы умеренного информационного «жульничества».
В отличие от сигналов-стимулов-символы не «понуждают» реципиента к определённым ответам, оставляя ему свободу выбора. Он сам активно ищет данную информацию, корректирует по ней собственное поведение и одновременно ретраслирует по коммуникативной сети. Всё происходит примерно как модем опрашивает сеть, и получив (в «должном» месте взаимодействия) «должный» сигнал, животное сообразно ему автоматически же меняет характер ретранслируемого сигнала – или оставляет прежде передаваемый без изменений. См. про живые машины Тьюринга.
Животное лишь, в отличие от модема, продуцирует сигнал, воспроизводя его «материальный носитель» — одну из демонстраций видового репертуара в собственных телодвижениях, исполняемых со стереотипностью не ниже некоторой, чтобы получатель мог устойчиво/однозначно воспринять инвариант формы сигнала. Поэтому он исполняется вопреки «шуму» и «возмущениям», созданным всеми прочими (тело)движениями участников и событиями вокруг, и никогда не использует такого рода «переливы фона» для сигнализации[1]. Также для восприятия сигналов, продуцируемых оппонентом, лучше, чтобы получатель уже предъявлял некоторую демонстрацию (одну из N поз репертуара), чем находился в обыденной позе. Та индивидуальность животного, которую изучают зоопсихологи[2], тут совершенно не при делах.
Это значит, что обмен демонстрациями инстинктивной природы «вслепую» воспроизводит «разумное» использование понятий в анализе окружающего мира, как инстинкт «слепо» воспроизводит логику разума (в отдельных операциях). Ибо сигналы-символы «именуют» логические альтернативные категории внешних событий, важных для вида. Почему формы разных сигналов этого класса также взаимно-альтернативны, в то время как формы чаще встречающейся и «более примитивной» категории — сигналов-стимулов связаны взаимными переходами.
Следовательно, наиболее продвинутые (и самые редкие) формы инстинктивного поведения — обмен демонстрациями, функционирующими как сигналы-символы при агрессии, ухаживании, предупреждении об опасности, подзыве к корму и пр. — «вслепую» повторяют не просто логическое мышление, но и использование понятий. Важно подчеркнуть, что они и самые редкие.
Коммуникация человека и животных: главнейшие различия
Что связано с двумя важными различиями коммуникации животных и словесного общения (в кибернетическом отношении):
1. Люди при речевой коммуникации сотрудничают, как показал Томазелло в диалоге люди побуждают друг друга к высказываниям, а уже высказанное поддерживает говорение. То же верно для невербальной коммуникации у нас и антропоидов — обмен жестами и взглядами между матерью и ребёнком, потенциальными половыми партнёрами, вообще членами группы помогает понять «в чём проблема» и поддерживает общение.
У животных наоборот, обмен демонстрациями инстинктивной природы направлен на то, чтобы демонстрирование партнёра «преодолеть» и, если не «заткнуть», то «обернуть в противоположность», переключив его поведение из конкурентного режима в управляемый и ведомый. Скажем, чтобы он в территориальном конфликте вместо предъявления всё более эффективных демонстраций угрозы «сдулся» и перешёл к позам подчинения, зеркальным по отношению к демонстрациям угрозы владельца. В ухаживании это демонстрации активного и пассивного партнёра при сближении и пр.
[«Животные» здесь — все имеющие репертуаре видовые демонстрации инстинктивной природы как отдельности, специфические по форме и специализированные по функции, т.е. «до мартышек» включительно. У макак-павианов сие разрушается, видовые демонстрации превращаются в индивидуальные реакции, мотивированные агрессией, сексуально и пр., т.е. «расплываются» в форме и теряют «значение», и употребляются редко в сравнении с собственно действиями. У антропоидов этот процесс завершён: остались лишь индивидуальные реакции, неспецифические и выражающие лишь общее возбуждение. См. «Человек» и «животное»: где провести грань?«].
2. У животных обмен демонстрациями — предельно рискованная и затратная штука, по возможным потерям сравнимая с передачей радиста разведгруппы в зоне действия функцабвера. Сигналы, которыми обмениваются участники процесса, передают информацию не только себе любимому, но и хищнику, и оппоненту. Первый ею воспользуется, чтобы словить, второй — чтобы бить и травмировать, а поскольку сигнальное действие демонстраций прямо пропорционально стереотипности исполнения образующих их телодвижений, точности «начертания» их специфических форм, то оно обратно связано с ловкостью и возможностью ухода от когтей хищника и удара под дых.
Плюс демонстрирование сильно энергозатратно, и если бюджет энергии особи здесь и сейчас напряжён (длительная плохая погода), приходится устанавливать отношения «сокращённым способом», без коммуникации. Иными словами, развёртывание обмена сигналами подчинено императиву «Короче и реже!», «только при крайней необходимости». У людей же обоих ограничений нет, болтать можно сколько угодно. Интересно, что коммуникация участников рынка, опосредованная деньгами, или танцоров в танце, опосредованная их символическими фигурами, по обоим параметрам соответствует таковой у животных.
И так как сигналы животных инстинктивны, их распознавание и отреагирование – процесс автоматический, отнюдь не задействующий их индивидуальность, а проходящий на более низком уровне центров среднего и промежуточного мозга. Чему есть красивые нейрофизиологические данные (1-2), противопоставляющие животных и человека.
В отличие от Homo sapiens, где всякая передача сигналов и вообще восприятие мира, его форм и структур, с соответствующей реакцией на них в деятельности совершенно немыслимы без ментальных репрезентаций, индивидуального понимания, личной эмоциональной окраски происходящего, и иных форм включения индивидуальности в коммуникацию с миром. У животных – наоборот, из коммуникации с миром индивидуальность участника исключена (она работает в сфере витальных потребностей, обеспечиваемых вне сферы коммуникации – где-то это корм, где-то безопасность, где-то – и спаривание; от вида зависит), это область кооперативной реализации видовых инстинктах в соответствующих сообществах, в соответствующем кругу социальных компаньонов животного (лоренцевский Kumpan) и пр.
Непонимание этого есть подспудный антропоморфизм, когда участников коммуникации животных считают действующими по той же схеме, что и мы грешные – руководствуясь собственным умом, чувствами и пр., только ум сильно хуже, а чувства порезче. То есть по умолчанию «мерят на свой аршин» (или наш общий), не понимая, что филогенетический путь от человека к животным предполагает качественный скачок, связанный с разрушением инстинктов и деградацией «общеобязательных» систем видовых сигналов «типа верветок».
Сегодня мы знаем: они есть у низших приматов, до мартышек включительно, затем деспециализируются («расплывается» форма, исчезает специфичность значения) у павианов с макаками и точно отсутствуют у горилл с шимпанзе. У них нет инстинктов так же, как у нас, и во всех поведенческих актах их индивидуальность участвует в той же степени, что и у нас, их жестикуляция, крики, прочие действия – индивидуальные реакции, а не видовые сигналы, как у верветок и более низших позвоночных.
Собственно, можно или нет «мерять на наш аршин», видеть в коммуникации животных деятеля с его мотивацией, кое-каким умишком и пусть примитивными, но «расчётами»[3], или это чисто автоматический процесс кооперативного запуска инстинктов, как в аналогии с модемом, определяется местом этой самой границы на филогенетическом древе позвоночных. Увы, она проходит достаточно близко к нам, так что у животных искать «деятелей» не приходится.
Следовательно, в коммуникации животных знак образуется и отреагируется индивидами инстинктивно, без корректировки их собственным опытом и собственным специфическим «пониманием» ситуации. В таком «автоматизме» всегда присутствуют промедления и «сбои», достаточно частые у молодых. Но отсутствие ответа, неточный или неадекватный ответ на сигналы партнёра – всегда «ошибка» коммуникации, а не намеренный «обман». Таковы неточные ответы на крики бедствия у верветок, Chiney, Seyfarth, 1990[4], или замедленный ответ – у синиц-пухляков Parus montanus, Rajala et al., 2003, или прямое преследование самки вместо ритуализованной демонстрации дупла у мухоловки-пеструшки Ficedula hypoleuca (Благосклонов, 1988). «Неправильные» индивиды быстро выбраковываются отбором, что и показано в соответствующих работах. Состояние особи – актора в процессе обмена демонстрациями меняется одним образом после эффективного разрешения проблемной ситуации взаимодействия, и другим – после неэффективного, что сходно с поведением конечных автоматов. Животное выступает здесь как конечный автомат, изменяющий состояние в процессе считывания информации, передаваемой при изменении состояния других автоматов, взаимодействующих с данным (Цетлин, 1969; Варшавский, 1973; Варшавский, Поспелов, 1984).
Отсюда приём-передача информации в актах демонстрирования и обмена демонстрациями делается неотъемлемой частью реализации инстинкта, так сказать, «программным управлением» инстинктивного механизма, составными элементами которого являются видовые демонстрации. Информационный обмен происходит так же «автоматически» и непроизвольно, как и «складывание» демонстраций из специфических телодвижений в ответ на предшествующие демонстрации партнёра, или их смена на основании информации о вероятных следующих действиях партнёра. Поэтому в математическом отношении коммуникация животных может быть описана как решение задач на синхронизацию в популяции K автоматов с L дискретными состояниями, каждый из которых меняет состояние на некоторую величину в зависимости от взаимодействий друг с другом (Цетлин, 1968; Варшавский, Поспелов, 1984).
Заключение
Вернёмся к сигналам-символам. Попытавшись поставить под сомнение само существование этой категории сигналов, критики добились противоположного — проверка «отшлифовала» предложенные ранее чисто операциональные техники ответа на вопрос «используются сигналы-символы данным видом в данном контексте коммуникации или нет?», и отделения их от «-стимулов» как сигналов ситуаций вовне vs сигналов мотивационных состояний внутри (см.таблицу).
И таки да, применение тех и других показывает, что ряд видов птиц и млекопитающих Х использует именно сигналы-символы, тогда как близкие формы — лишь стимулы, и это сказывается на представлении демонстраций вышеописанным образом (в случае гомологии их форм). И это именно наиболее напряжённые и важные контексты общения с т.з. биологии вида (и у видов первой категории это «более узкие« места, чем у видов второй).
Важней всего здесь два типа реакций. Во-первых, возможность специфического ответа на тревожные сигналы другого вида, причём не всегда близкого (т.е. его особи реагируют на значения адекватно, не испытывая воздействия их «материальных носителей« демонстраций как стимулов). Птицы направленно выделяют «значащие формы» из звукового фона вокруг и реагируют вполне адекватно, чем окончательно подтверждают их символическую природу. Во-вторых, возможность «обмана», во внутри- или межвидовой коммуникации равным образом.
Таким образом, реализация инстинкта в коммуникации всегда изоморфна разумному выбору. В отдельных же случаях (использование сигналов-символов) она также изоморфно использованию понятий — но, конечно, не гомологична ему.
Рекомендуем прочесть
 Первобытные деньги и становление работорговли
Первобытные деньги и становление работорговли «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Год — с винтовкой и плугом
Год — с винтовкой и плугом Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции
Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции Об истории, культуре и современных проблемах коми. Часть 3.
Об истории, культуре и современных проблемах коми. Часть 3. Опыт работы по поддержанию местообитаний редких видов ржанкообразных в Виноградовской пойме
Опыт работы по поддержанию местообитаний редких видов ржанкообразных в Виноградовской пойме Виртуальные миры: анализ содержания психологических эффектов аватар-опосредованной деятельности
Виртуальные миры: анализ содержания психологических эффектов аватар-опосредованной деятельности Социальный ранг и психика. Синантропные грызуны в «жилой комнате»
Социальный ранг и психика. Синантропные грызуны в «жилой комнате»
