

Популяция как [хорошо выделенная] система
Содержание
Помню, был у меня с ivanov-petrov’ым в ЖЖ диалог относительно популяции, которую он считает плохо выделенной системой, а я – наоборот. Я говорил: но как же, у неё есть целостность и собственная устойчивость, она регулирует перемещения и взаимодействия индивидов так, как нужно для наиболее точного воспроизводства характерной структуры системы в данной мозаике местообитаний, год от году меняющейся, вопреки образующим эти изменения средовым и демографическим флюктуациями, а не вслед за ними? Нежели ей можно отказать в самости (или сущности), ведь это — почти видовой морфотип, только другим механизмом «делается» и поддерживается – не онтогенезом, а внутрисистемной коммуникацией… между которым не зря же есть интересные изоморфизмы…
А он возражал: есть и целостность и устойчивость – но только их источники не в самой популяции, а, например, в генах и прочей «биологии» особей, телодвижения которых «складываются» в должные демонстрации в должных точках коммуникативных процессов, которыми популяционная структура сперва «делается», а потом воспроизводится на данной территории, в данной мозаике местообитаний. «Грубо говоря, сигнальная поза хранится в индивиде, а не в популяции».
«Поза» — да, но даже совокупность «поз» — видовая этограмма не исчёрпывает сигналов, существенных для воспроизводства характеристических отношений системы. Важная их часть, образующая сигнальное поле группировок и, шире, популяции в целом, представляет собой элементы среды, специфическим образом преобразованные деятельностью индивидов и, тем самым, вынесена вовне.
Благодаря сигнальному полю популяция обладает своего рода наследственностью, но «внешней» и как бы ламарковской, где преемственность воспроизводства структуры населения вида на конкретной территории обеспечивается элементами среды, соответствующе преобразованными активностью особей и в меру преобразованности «покрытыми сигналами». Поле ориентирует и «направляет по цели» во время перемещений нерезидентных особей, пополняющих группировки и связывающих их в системное целое – ту самую популяцию.
Благодаря ему даже в однородной среде особи перемещаются между поселениями и группируются в них неслучайными образом, а воспроизводят «кружева» и «фигуры», отвечающие видовой «конструкции» пространственно-этологической структуры. Больше того, точное воспроизведение этих последних для индивидов важней собственной приспособленности. Сплошь и рядом ради первого они жертвуют вторым, если в неблагоприятных условиях невозможно достичь этих двух целей одновременно; если возможно, сперва особи во взаимодействиях между собой воспроизводят типическую структуру социальных отношений (образующих ту структуру отношений тока, колонии, территориальной группировки, что характерна для вида), и лишь потом направляют усилия на эксплуатацию кормовых запасов участка, или привлечение самок, т.е., на рост собственной приспособленности. См. примеры; С.П.Харитонов, анализируя поведение колониальных видов птиц, называл это «приматом системности над адаптивностью«. Он проявляется сполна во всех иных формах социальной организации позвоночных (кроме сугубого меньшинства «несоциальных» видов).
В реальной среде, со сложными формами рельефа и мозаикой местообитаний разного качества направленность переселений между группировками – как резидентов, так и птиц из «подвижного резерва» популяции – ещё заметней. Она резко контрастирует со случайным перемещением, предполагаемым в нынешних представлениях о популяции (в крайнем случае ограниченном направляющими линиями ландшафта).
Вообще, крайне важно, что при местных переселениях особи направленно движутся к цели, а не «рыщут» в её поисках (тем более нет случайного блуждания). Это верно как для резидентных особей, меняющих поселение в связи с вытеснением с прежней территорией или, наоборот, эффективной её охраной и прочим успехом (первые движутся в «худшие» — наименее плотные и постоянные во времени/пространстве поселения данной местности, вторые — в «лучшие»), так и для нетерриториальных птиц из резерва, занимающих участок везде, где возможно «воткнуться».
В том числе во временно возникающих местообитаниях, где возможно интенсивное размножение — но недолго. Появление таких мест — вполне случайный процесс, поэтому кажется, что в подобные поселения попадают исключительно в ходе «блужданий». Но нет; практически все вселенцы, занявшие здесь территории, двигались в нужное место направленно. Подчеркну, что «дистанционное наведение» переселенцев всех категорий осуществляется на дистанциях, много больших обычного расстояния исследовательских выходов за пределы собственного участка. Иными словами, оно направляется сигналами «привлекающих» группировок, а не прошлым опытом пребывания в районе возможного поселения раньше.
Действительно, птицы и млекопитающие движутся по «ландшафтам привлекательности» и «против» «ландшафтов страха», о которых писал ранее. Эти два типа градиентов образуют сигнальное поле, они «протягиваются» во всех устойчиво заселённых видом местностях и, помимо направления перемещающихся птиц, воспроизводят характерную плотность, число резидентов и пр. характеристики социальной среды группировок.
Их притягательность прямо пропорциональна плотности (или интенсивности) элементов сигнального поля (вторичных сигналов, в противоположность первичным – параметрам местообитания как такового). Она, в свою очередь, пропорциональна преобразованности активностью животного тех элементов среды, на которые направляется эта последняя – вроде плотности кузниц больших пёстрых дятлов, «забитых» раздолбанными шишками, или изрытости грунта норами сурков, песчанок и пр. Эта трансформированность среды, а не сами вторичные сигналы, обеспечивает устойчивость привлечения особей «со стороны» на временных интервалах, сравнимых с жизнью одного или нескольких поколений.

Старые шишки и кузницы больших пёстрых дятлов; их обилие и сменяемость показывает соседям/вселенцам интенсивность использования данной части территории

См. обзор современного состояния концепции сигнального поля, Е.А.Ванисовой и А.А.Никольского1 (2012).
Концепция сигнального поля: современное состояние
«Перемены в среде, производимые популяцией, приобретают информационное значение и служат своеобразной памятью группы, организующей поведение особей (Наумов, 1973б). Следы жизнедеятельности, которые передаются из поколения в поколение, Н.П. Наумов называл “долгоживущими сигналами”. Он видел в них механизм, с помощью которого новым поколениям животных передаётся опыт предшествующих поколений, связывая между собой разные генерации (Наумов, 1973а). К “долгоживущим сигналам” относятся норы, тропы, скопления помёта и другие запаховые и визуальные метки. Отражая деятельность (опыт) предшествующих поколений, эти следы жизнедеятельности представляют собой своеобразный “аппарат памяти” в биогеоценозе (Наумов, 1973а, в).
Этот механизм, позволяя каждому последующему поколению использовать опыт предыдущих поколений, сокращает время и энергию на освоение территории со всеми находящимися на ней ресурсами. Считывая информацию “долгоживущих сигналов”, новые поколения животных воспроизводят пространственную структуру популяций своих предшественников, тем самым повторяя траектории предыдущих поколений. Вот что, например, пишет Н.П. Наумов (1973а, с. 813) об убежищах, как “долгоживущих сигналах”:
“Распределение и строение длительно существующих убежищ отражают опыт поисков оптимальных мест обитания и типов убежищ всеми предшествующими поколениями. Использование убежищ как пространственных ориентиров молодыми расселяющимися особями повышает их выживание, упрощая задачу поисков и сокращая расход их времени и сил.”
В свою очередь последующие поколения животных так же вносят вклад в поддержание сигнального поля. Они обновляют “долгоживущие сигналы” – наиболее стабильные элементы, оставляя в пространстве биогеоценоза следы своей жизнедеятельности. Благодаря перманентному процессу обновления стабильных элементов, сохраняется образ использования территории предыдущими поколениями (образ пространственной структуры популяций).
Существенно, что неограниченно долгое существование стабильных элементов сигнального поля сопоставимо по длительности с существованием самой экологической системы. Каждая популяция млекопитающих наследует не только видовой генотип, но и запахововизуальный образ территории. Точно так же биоценоз наследует не только биомассу и видовое разнообразие, но и информационное поле, оставляемое рядом поколений образующих его популяций».
[Получается как бы ламарковская наследственность. Долговременно значимые элементы среды, не меняющие характер значимости, трансформируются деятельностью новых особей в одном направлении, почему обновляются и усиливаются как сигналы, при неизменном характере действия. А меняющие значение или утрачивающие его начинают подновляться иначе, или перестают вовсе, что меняет и характер вторичных сигналов. Поэтому популяционная структура, воспроизводимая деятельностью разных поколений особей, сопрягаемых и координируемых через сигнальное поле, сочетает консерватизм главных черт, относящихся к видоспецифической «конструкции» системы, с изменчивостью побочных, приспасобливающих «конструкцию» к конкретным условиям данного места и демографии2.]
Элементы сигнального поля делятся на стабильные и нестабильные. Первые наиболее константны, они долговременно меняют среду, образуя сеть устойчивых ориентиров на территории особи, важных как для неё, так и для соседей с вселенцами. Нестабильные «привязываются» к стабильным, их распределение зависимо от них и меняется с изменением состояния матрицы стабильных элементов.
«В поселении песца функцию стабильных элементов выполняют норы, тропы, сигнальные кочки, наблюдательные лёжки, временные убежища. Нестабильными элементами могут быть, например, поеди или фекалии песцов, сосредоточенные у стабильных элементов. Позже наличие системы стабильных и нестабильных элементов было показано на уникальной структуре сигнального поля бобра (Castor fiber) в заповеднике “Рдейский” (Завьялов, 2009). К стабильным элементам относятся, в частности, легко выделяющиеся участки в поселениях бобров с интенсивной многолетней маркировкой.»
«Даже при сильных депрессиях численности вида на соответствующей территории элементы сигнального поля сохраняются и направляют повторное расселение. Благодаря этому восстановленная сеть группировок в значительной степени повторяет прежнюю, хотя новое население вида «составлено» уже иными особями [в т.ч. генетически иными, вследствие «бутылочного горлышка» при депрессии]».
Первые поселенцы, расширяющие ареал вида в процессе расселения, начинают формировать сигнальное поле, маркируя базовые аттракторы и облегчая тем самым каждому новому расселяющемуся поколению экспансию территории. Так, Г.А. Савченко (2002), наблюдая за меченными степными сурками, обратила внимание, что новый хозяин начинает осваивать территорию с ближайших окрестностей участка, где следы жизнедеятельности предыдущего хозяина наиболее устойчивы.
Неравномерная плотность распределения следов жизнедеятельности, стабильных и нестабильных элементов создает неравномерную интенсивность сигнального поля. Их скопления образуют “сгущения”, а изменения в пространстве – градиенты сигнального поля. Следы жизнедеятельности, образующие сигнальное поле как аттрактор второго порядка, зависимы от аттракторов первого порядка, имеющих ключевое значение как экологические факторы, в организации ритма активности животных во времени и в пространстве. При этом имеет значение не только положение аттракторов первого порядка в пространстве, но и их свойства. В условиях эксперимента на волках было показано, что наиболее эффективными аттракторами становятся наиболее заметные, выделяющиеся из окружающей среды объекты, концентрируя вокруг себя активность как отдельных животных, так и группы в целом (Ванисова и др., 2012).
Для многих видов выявлены определенные предпочтения в выборе мест маркировочной активности. Запаховые метки, образующие сигнальное поле многих видов млекопитающих, приурочены к различным конкретным объектам среды (Рожнов, 1988). Песцы (Наумов и др., 1981), тигры (Panthera tigris) (Кучеренко, 1985) и околоводные куньи (Рожнов, 2011) выбирают для нанесения запаховых меток наиболее заметные элементы окружающего ландшафта, с наибольшей вероятностью попадающие в поле зрения других особей. Согласно нашим наблюдениям в Туве, аналогичные объекты выступают в качестве эффективных аттракторов для ирбиса (Uncia uncia). Возле них наблюдается повышенное скопление следов жизнедеятельности – элементов маркировочной активности. А.Д. Поярков с соавт. (2011) отмечают, что значительная часть поскребов ирбиса располагается возле крупных камней и скальных стенок. Соотношение запаховых и визуальных меток, их взаимная приуроченность могут зависеть от возраста, пола, физиологического состояния животного и времени обитания на участке, что было отмечено, например, для дальневосточного леопарда (Panthera pardus orientalis) (Пикунов, Коркишко, 1992).
К определенным элементам биотопа приурочены постоянные переходы-лазы рыси. Большинство из этих образований в течение многих лет остаются практически неизменными, и хищники перемещаются по ним даже по прошествии 30 лет, несмотря на смену поколений (А.С. Желтухин, С.А. Желтухин, 2009).
Некоторые виды млекопитающих сами формируют выделяющиеся объекты, к которым привязывают запаховые сигналы. Песчанки, например, сооружают из песка и снабжают запаховой меткой “сигнальные холмики” (“сторожки”), которые становятся для них хорошо заметным визуально-ольфакторным ориентиром и эффективным элементом маркировки территории (Рожнов, 1988; Громов, 2000). Похожие сигнальные холмики с секретом мускусной железы формируют бобры (Завьялов, 2009). Выразительным запахово-визуальным аттрактором являются “утолоки” волков.
Они уже издали различимы по желтеющей траве и белеющим костям, а вблизи определяются по характерному запаху (Федосенко и др., 1978). Весной 2012 г. мы наблюдали в заповеднике “Тигровая балка” (Таджикистан) множество уборных джейрана (Gazella subgutturosa) – сложно устроенные, далеко заметные аттракторы, которые, располагаясь на тропах, регулярно привлекают антилоп. Уборная представляет собой участок земли без растительности, овальной формы размером примерно 100 × 50 см, который заполнен помётом джейранов, отложенным в разное время и разными особями. По периферии уборной разрастается травянистая растительность, представленная 5–7 фоновыми видами. Всё вместе – расчищенный копытами джейранов участок почвы, кучки помёта, бордюр из яркой густой зелени и, конечно, запах помёта, оставленного многими особями, делают уборные джейрана стабильными элементами сигнального поля (рис. 1)».

Для характеристики элементов сигнального поля Гольцманом и Крученковой было предложено понятие аттрактора. Само поле формируется аттракторами разной степени устойчивости (и разной степени долговременности сигнала, связанной с преобразованностью среды обитания) и разной силы привлечения или отталкивания особей своего вида или биоценотических партнёров.
Перемещаясь по градиентам сигнального поля, млекопитающие оптимизируют усилия на использование ресурсов в пределах занимаемой ими территории. Зонирование территории по экологическим свойствам и функциям (кормовой участок, гнездовой участок, защитный участок и т.п.) у большинства видов млекопитающих “размечены” соответствующими аттракторами, содержащими конкретную информацию. В частности, норы, логова, гнезда сигнализируют не только о занятости территории конкретными животными, но также о её «качестве», интенсивности эксплуатации, успешности владельца в сравнении с соседями и много чём ещё.
«Информационная актуальность зонирования территории обеспечивается за счет регулярного подновления элементов сигнального поля. Для сурка Мензбира, например, характерно маршрутное мечение, когда зверек регулярно движется определенным маршрутом и целенаправленно ставит метки (Машкин, Батурин, 1993). В результате перемещения по одним и тем же маршрутам и маркировочной активности дальневосточного леопарда в заповеднике “Кедровая падь” сформировалась сеть постоянных троп, вдоль которых располагаются визуальные и ольфакторные метки (Пикунов, Коркишко, 1992), что наглядно отражает структуру использования животными пространства.
В Таджикистане мы неоднократно встречали маркировочные деревья бухарских оленей (Cervus elaphus bactrianus) с метками разных лет давности. Самцы оленей рогами сдирают кору с деревьев и заламывают ветки, что придает этим аттракторам характерный выразительный облик (рис. 2). Много лет подряд одни и те же сигнальные деревья метит и бурый медведь (Ursus arctos).

В Удмуртии были обнаружены метки возрастом до 9 лет. Более того, одни и те же деревья могут маркироваться разными самцами бурого медведя и кабаном (Sus scrofa) (Пучковский, 1988). По нашему мнению, такие деревья, приобретая значение стабильных элементов в структуре сигнального поля, выступают в качестве сигнальных пунктов. Они играют роль универсальных аттракторов, привлекая внимание разных видов животных, в данном случае медведя и кабана».
Интересно, что сигнальное поле зоологов повлияло на концепцию политического дискурса А.К.Михальской. Согласно ей, он тоже имеет полевую структуру, и включает репелленты и аттрактанты. Это слова и образы, соответственно с позитивным или негативным эмоциональным воздействием, которое проявляется вне и помимо собственно семантического содержания этих слов и выражений (своего рода изоморфизм запаховых и прочих меток в сигнальном поле животных, привлекающих и отталкивающих).
«Аттрактанты и репелленты – обладают свойством прямо, вне и помимо текста влиять на поведение человека, выполняя роль сильного элементарного положительного или отрицательного стимула» (Михальская, 2013, с. 282). И аттрактанты, и репелленты относятся к классу т.н. фасцинаторов, то есть значений, связывающих и удерживающих внимание». В том числе они действуют «автоматически» и тогда, когда способность людей извлекать содержательную информацию из текста или непосредственно наблюдаемой ситуации (а тем более использовать её по назначению) «отключена» страхом, агрессией, жаждой наживы и т.д. сильными эмоциями. Скажем, во время массовой паники толпа тем не менее воспринимает людей в погонах, в форме и т.д., и склонна следовать именно их решениям, чем можно воспользоваться для спасения ситуации.
Всякая политическая сила для сплочения сторонников и противостояния противникам использует не только убеждение со всеми багами и фичами пропаганды, но также взращивает «удобное для себя» поле. В его системе репеллентов и аттактантов, если она достаточно развита и устойчива, оппонент просто не может внятно выразить свои идеи. Соответственно, задача его публичной риторики — «разложить и обезвредить» чужое сигнальное поле.
«Возвращаясь к работе Н.П. Наумова с соавт. (1981), посвящённой сигнальному полю медновского песца, мы обращаем внимание, что особое место в структуре матрицы стабильных элементов занимают сигнальные кочки вдоль песцовых троп (рис. 3). Для формирования такого сложного образования, каким являются сигнальные кочки, требуется длительное время и смена многих поколений песцов. В образовании кочек принимают участие не только песцы, но и сложный почвенно-растительный комплекс, свойственный островному биоценозу. Эти невысокие холмики образуются в результате разрастания дерновины злаков, вызванного составом почвы, изменённой экскрементами песцов. Кочки хорошо заметны на местности и, являясь объектом преимущественно запахового маркирования, становятся, таким образом, самостоятельным зрительно-запаховым ориентиром.
На о. Врангеля стабильными элементами сигнального поля песцов являются норы. В период гона они выполняют роль сигнальных центров, обеспечивая встречу брачных партнеров. Совокупность нор в поселении можно рассматривать как матрицу, на [лучше — «по»] которой уже в начальной фазе периода размножения формируется или восстанавливается территориальная структура поселения (Овсяников, 1993).
В качестве другого примера матрицы стабильных элементов в зрелых экосистемах можно привести систему троп и нор в колонии степных сурков (Marmota bobak). Зональные степи представляют собой зрелое сообщество, а их облик в значительной степени создают поселения этих крупных грызунов, обычно имеющих высокую плотность популяции. Холмики земли, возвышающиеся над постоянными норами, и проложенные между ними тропы (рис. 4), которыми сурки пользуются регулярно, образуют матрицу стабильных элементов, ограничивая использование сурками территории с имеющимися на ней ресурсами. Причём ресурсами не только кормовыми, но и защитными, что для зимоспящих млекопитающих имеет исключительное значение, так как норы, в которых сурки проводят спячку, отличаются особым температурным режимом и специфической сезонной динамикой (Nikol’skii, 2009). Дополнительным стабильным элементом, усиливающим информационный эффект, является растительность на холмиках земли (бутанах) около нор. Как и на сигнальных кочках песцов, она сильно отличается от окружающего фона и хорошо заметна на большом расстоянии. Кроме того, около входа в каждую из постоянных нор сурки устраивают туалеты, дополняя зрительный образ колонии запаховым.

Норы сурков, образуя базовый элемент матрицы стабильных элементов в степных биоценозах Евразии и Северной Америки, имеют большое сигнальное значение не только на популяционном уровне, но и на биоценотическом. Они привлекают десятки видов как позвоночных, так и беспозвоночных животных, а на их поверхности формируются специфические растительные ассоциации (Бибиков, 1967). Похожую функцию на биоценотическом уровне выполняют поселения барсука (Meles meles) в таежной зоне. На протяжении многих лет в Дарвинском заповеднике одни и те же поселения используются не только барсуком, но также лисицей (Vulpes vulpes) и енотовидной собакой (Nyctereutes procyonoides), регулярно посещаются копытными и птицами, а в нежилых поселениях иногда устраивает логово волк (Canis lupus) (Сидорчук, Рожнов, 2010).
В различных экологических системах, особенно в лесных, на зрелой стадии сукцессии универсальным стабильным элементом сигнального поля являются магистральные тропы. Ими пользуются многие виды млекопитающих, причем как хищники, так и их жертвы. По магистральным путям зайцев, например соединяющим места жировок и лежек, перемещаются лисица, енотовидная собака, куница (Martes martes), рысь (Lynx lynx), волк, косуля (Capreolus capreolus). При этом магистральные тропы оказывают влияние на пространственное распределение не только тех видов, которые эти тропы создавали, но и многих других (Гайдук, 1988).
Перемещаясь из поколения в поколение по одним и тем же маршрутам, натаптывая магистральные тропы, звери сильно изменяют поверхность почвы, придавая ландшафту своеобразный, легко узнаваемый облик (рис. 5). Опытнейший следопыт, Е.Н. Матюшкин (1977, с. 148), пишет, ссылаясь на Н.П. Наумова (1973): “Одни и те же тропы нередко используются разными видами зверей, приобретают значение длительно сохраняющихся ориентиров и вплетаются в общее сигнальное биологическое поле биоценоза”.
Закономерности формирования сигнального поля биоценотического уровня недавно были показаны на примере тропы в Тверской области (Блидченко, 2011). Млекопитающие консервативны в использовании магистральных троп. Оказавшиеся на тропе препятствия (например, бревно) они не только не обходят, но используют их в качестве стабильных элементов, регулярно оставляя на них следы своего присутствия (Никольский, Фроммольт, 1989; Никольский, 2003) (рис. 6).

Структура сигнального поля, реализуемая через матрицу стабильных элементов, неоднородна. Следы жизнедеятельности млекопитающих располагаются и накапливаются в пространстве биогеоценоза неравномерно. Например, сигнальные кочки и фекалии песцов на некоторых отрезках троп образуют заметные скопления, связанные с определенными точками участка (Наумов и др., 1981). Неравномерное распределение следов жизнедеятельности, сочетание стабильных элементов с менее стабильными создаёт градиенты сигнального поля, различные уровни его интенсивности, облегчая млекопитающим ориентацию в пространстве биогеоценоза. Так, нарастающий градиент биологического сигнального поля, образуемый смываемыми с лежбища зоогенными элементами, может служить надежным ориентиром ластоногим на подходах к лежбищу (Лисицына, 2002). В значительной степени по градиентам биологического сигнального поля происходит расселение зубров в Кавказском заповеднике. Матрицей стабильных элементов в данном случае является сеть троп, соединяющих такие жизненно важные объекты, как сезонные пастбища, солонцы, броды (Трепет, 2008).
Зрелость стадии сигнального поля косвенно подтверждается тем фактом, что, наследуя от предыдущих поколений его структуру, последующие поколения животных удивительно точно повторяют использование территории своими предшественниками. Об этом наглядно свидетельствует структура индивидуальных участков в ряду поколений. У северной (Ochotona hyperborea) и алтайской (O. alpina) пищух в случае смены хозяина участка его новый владелец воспроизводит использование территории прежним хозяином (Nikol’skii et al., 1990; Nikol’skii, Mukhamediev, 1997). Сходная картина при смене хозяина участка наблюдается и у степного сурка (Nikol’skii, Savchenko, 1999). В.А. Зайцев (1991), описывая аналогичные территориальные замещения у кабарги (Moschus moschiferus), назвал эти постоянные участки территории – “ячейки-вакансии”. После длительного отсутствия волки также занимают участки обитания своих предшественников (Назаренко, Гурский, 1969; Буневич, 1989; Лихацкий и др., 1995).
Сигнальное поле, насыщенное следами жизнедеятельности ушастых тюленей, обеспечивает функционирование лежбищ на протяжении многих лет. Участки старых лежбищ, долго сохраняющие запах помёта и секрета желез, расположенных на ластах зверей, являются устойчивыми элементами сигнального поля, стимулируют выход первых секачей на сушу. Мощный запаховый фон лежбищного субстрата не исчезает даже в течение зимнего отсутствия ластоногих (Лисицына, 1988, 2002). Лишенные следов жизнедеятельности участки побережья заселяются тюленями крайне медленно. Для формирования полноценного лежбища необходимы многие годы, пока за счет накопления следов жизнедеятельности сигнальное поле не достигнет определенной интенсивности.
Таким образом, зрелые экологические системы, вероятно, характеризуются наиболее устойчивым сигнальным полем, образуемым множеством стабильных элементов. Млекопитающим в этом процессе принадлежит ведущая роль, так как они оказывают наибольшее физическое воздействие на ландшафт и наиболее активно используют информацию, сохранённую в оставленных ими следах жизнедеятельности».
Ванисова, Никольский, 2012, op.cit.
Сигнальное поле — блок памяти, регулирующий точность и устойчивость воспроизводства популяционной структуры через управление перемещениями и взаимодействиями особей. Чем выше плотность поселения и интенсивней взаимодействия, тем выше продукция вторичных сигналов, разность устойчивых концентраций которых в среде образует аттракторы, градиенты и т. д. элементы поля. Если собственно сигнальное поле наиболее важно в плане дистанционного наведения активных вселенцев (направленность переселений которых связывает устойчивые группировки в системное целое, популяцию, позволяет «перебрасывать» особей из резерва в места недавнего истребления для восстановления численности), то для резидентов внутри группировок аналогичную функцию выполняет коммуникативная сеть с постоянным «подглядыванием» компаньонов друг за другом.
Так в каждой из местностей, заселённых видом, возникает система аттракторов, подправляемых и воспроизводимых из года в год, соотношение «силы» которых даёт представление о градиентах изменений плотности и устойчивости группировок в данной местности, т.е. градиентов их привлекательности для мигрантов и особей из «подвижного резерва популяции», выбирающих место для подселения. Так в каждой из местностей сигнальное поле прочерчивает градиенты социальной плотности: «вверх» по ним движутся успешные особи, переселением в более плотные/устойчивые группировки старающиеся повысить свой статус, «вниз» — лузеры, вытесненные с территорий, потерявшие партнёра и пр.
Между разными местностями то же сигнальное поле обеспечивает сортировку особей по потенциям развития поведения, перенаправляя предпочитающих социальный стресс энвиронментальному всё ближе к центральной части, а особей с противоположными предпочтениями — сбагривающее на периферию. В результате перемещения особей между поселениями не случайны, а организованы соответствующей системой градиентов (см. рис.6Б ниже).
Популяция как система с определённой структурой (информационным компонентом которой выступает сигнальное поле) «умеет управлять» перемещениями и взаимодействиями особей — своих элементов, так что с высокой вероятностью особи с определёнными потенциями развития поведения оказываются именно в той части ареала, в тех группировках, в той экологической среде, которая наиболее благоприятна с точки зрения развития именно их поведения (относящегося обычно к одной из N возможных жизненных стратегий). Лучше всего это выходит в урбанизированных популяциях, почему их структура устойчиво воспроизводится в ландшафтах с наиболее нестабильной и быстро меняющейся (и случайным образом и направленно) мозаичностью среды — в городах.
Поэтому популяции позвоночных в значительной степени автономизированы от среды: мало того, что социальная плотность уже существующих группировок создаёт как бы кокон, опосредующий все внешние воздействия на особей (так что действия, скажем, экологических факторов или паразитов воспринимаются через производимые ими изменения социальной среды), но также благодаря направляющей функции сигнального поля система так регулирует разнокачественность особей, растасовывая их между группировками в разных частях ареала, с разной социальной плотностью, разной устойчивостью и пр., что каждая особь с высокой вероятностью оказывается именно в той среде которая оптимальная для развития именно её поведенческих потенций. Если конечно, не погибнет в процессах сортировки и переброски, или в период пребывания в «подвижном резерве». Об этом же см. статьи Н.А.Щипанова с соавт. про мелких млекопитающих; иллюстрация механизма сортировки см. рис. 1А-Б здесь.
От саморегуляции к номогенезу?
Некогда Б.Б.Жуков при объяснении, почему достижения эпигенетики — не прорыв в эволюционной теории и не доказательство, что «Ламарк был прав», замечательно выразился об ошибочности «исходить не из веры в то, что организм всегда знает, как ему меняться под действием того или иного фактора». «На первом шаге» анализа эволюционных процессов это действительно так — пока мы абстрагируемся от того, что особи, существующие в мозаике местообитаний и подвергающиеся отбору, не независимы, а связаны вполне конкретными отношениями — социальными (к компаньонам), территориальными (к пространству), биотопическими (к растительности и пр. параметрам местообитания).
«Конструкция» этих отношений (видо)специфична не менее видовой морфологии, разные типы их действуют на разном уровне — сигнальное поле ориентирует нерезидентов и активных вселенцев между группировками, коммуникативные сети направляют выбор следующих взаимодействий (их места, времени и форм поведения внутри группировки) и пр. Так или иначе индивиды большую часть годового цикла остаются не независимыми, а связанными — со всеми последствиями, что описаны выше.
Всё перечисленное можно рассматривать как инфраструктуру, создающую «дальний порядок» внутри популяционной (или социальной) системы вида. Она даёт всем «подключённым» к ней особям информацию о «качестве» местообитаний и группировок далеко за пределами их непосредственного окружения как в пространстве, так и во времени. Причём речь идёт о возможности оценить ситуации и опережающе среагировать на расстояниях, много больших размеров не только индивидуального участка особей, но и «исследовательских выходов», из него, и на временах, сравнимых с несколькими поколениями.
Точно также как сейчас ясно, что все экологические взаимодействия — с хищниками, конкурентами и пр. — происходят не напрямую, а опосредуются и предваряются информационными. Что даёт их участникам возможность опережающе реагировать на «слишком опасные» места или «слишком напряжённые» моменты жизни, уходя от элиминации и всё больше оставляя отбору роль «закрепителя» приспособительных изменений, созданных сортировкой особей по потенциям в «теле» популяционной системы.
Сходным образом действует «сигнальная наследственность», «оттискивающая» стиль родительского, агрессивного и пр. видов поведения потомства «по образу и подобию» родителей. Этот эффект осуществляется самыми разными способами — через копирование поведения «учителей», сигнальную индукцию, воздействием изменений гормонов и нейромедиаторов в желтке яйца (возникших в период его формирования в связи с обстоятельствами брачных и пр. отношений данной самки) на поведение потомства и пр.
В самом деле, главная проблема, встающая перед индивидом в борьбе за существование — как эффективно находить те «пятна» местообитаний и те группировки, которые по параметрам экологической и социальной среды наиболее соответствуют его физиологическим и поведенческим потенциям. Проблема важна, поскольку вне их он размножиться практически не может, попасть туда надо за конечное время, а тут ещё на пути хищники, непогода, стресс и все прочие риски…
Поэтому надо уметь обнаруживать ближайшие «острова комфорта и безопасности» и при перемещениях направленно двигаться в их сторону, не тратя время на «слепой рыск». При этом расположение благоприятных и неблагоприятных «мест поселения» меняется в силу разных причин, непрогнозируемых и неподконтрольных для особи, почему может рассматриваться как случайное и с т.з. факторов направляющих её поведение, и с т.з. факторов определяющих её место рождения.
За счёт этого снимается существующее в Природе забавное противоречие. С одной стороны – исключительно сильное «напряжение жизни». Судя по всему, борьба за существование у животных намного напряжённей и жёстче той нашей собственной, социальной конкуренции, — на работе ли, дома, на митинге или при посадке в автобус. Так, С.А.Полозов восхищается «напряжению», с которым самец зелёной жабы Bufo viridis в копетдагской полупустыне должен отыскать нечастые здесь подходящие лужицы, а затем интенсивно петь, привлекая самку.
«Посмотришь на такое – одна лужица в округе, а он нашёл её и сидит, и поёт в ней, и ждёт самку, надеясь, что она услышит, и придёт, и сразу становится понятно, что жизнь во многих местах натянута как струна. К счастью, струна весьма крепкая. Воды ведь может и не быть в нужный сезон; или она может испариться быстрей, чем разовьются головастики» (Фасциатус. Ястребиный орёл и другие. Документальная орнитологическая сказка. М.: Армада-пресс. С.111).
Вообще говоря, «напряжение жизни» на порядок выше у самки, откликнувшейся на его призыв, ведь самец выбирал водоём и безопасный путь к нему по собственному усмотрению, тогда как самка движется, уже «доверяя» его сигналу и его выбору. В случае экстремальной среды и одном и том же «напряжении жизни» самец (и вообще активный партнёр в коммуникации) может изменить/скорректировать выбор, если вдруг окажется ошибочным, тогда как самка (и любой пассивный партнёр) потеряет всё из-за ошибки самого выбора.
Впрочем, это дело личной позиции – одних восхищает жёсткость конкурентной среды или эффективность преуспевших в ней, других – доверие к сигналам системы при неущербной способности распознавания. Наверно, поэтому Сергею Александровичу удобней жить в США, а мне – скорей здесь.
Например,
«хронометрировали самца серой вороны (Corvus cornix), самка которого насиживала кладку. Стояла тёплая погода конца мая; за 4 дня птица израсходовала на самообеспечение и брачное поведение 9,6 ч, а 6,4 ч были заняты весьма разнообразной деятельностью «активного безделья». По своей форме эти действия «могли быть отнесены к кормёжке, запасанию, патрулированию территории, внутри- и межвидовым контактам, если бы объекты, на которые они направлялись, были более естественными.Под постоянным наблюдением вороны находились три собаки (одна молодая, «своя», жившая на том же дачном участке, где было и гнездо, и две соседских), два кота, старик и старушка на «своём» участке, люди на двух смежных участках, пара зябликов, пытавшихся гнездиться на дубах, и две пары соседских ворон. «Своя» собака использовалась для охотничьих игр – ворона подходила или подлетала к ней вплотную, провоцируя погоню. Кроме того, собака должна была разыскивать предметы (как съедобные, так и несъедобные), которые ворона прятала у неё на глазах, в местах то доступных, то недоступных.Первая соседская собака, сидевшая на цепи, использовалась для игр в нарушение чужих владений: ворона разгуливала по двору вне досягаемости цепи, трогала и перекладывала разные предметы до тех пор, пока собака не пыталась на неё броситься. Вторая соседская собака, имевшая привычку зарывать куски пищи в землю и охранять их, использовалась для игр в похищение этих кусков, причём в наиболее сложных акциях участвовали то один, то оба соседних самца.
Оба кота побаивались вороны и использовались как модель вторгшихся врагов: все их перемещения по двору ворона сопровождала, разрешая ходить только по периметру дома, сарая, забора и не позволяя залезать на высокие предметы. В противном случае они атаковались и на них обращалось внимание собак.
Ворона наблюдала за зябликами, пытаясь разгадать, где они очередной раз пытаются вить гнездо, а выследив, разрушала постройку.
Старушка сажала огород, и стоило ей удалиться, как ворона находила в грядке семена и рассаду, выдёргивала, а иногда и перепрятывала. Мужчины в эти дни занимались ремонтом крыши сарая, ворона воровала гвозди, носила их на сарай на соседнем участке и засовывала в дыры в кровле.
Помимо этих главных занятий за 4 дня ещё 25 мин ушло на безуспешную попытку отвязать от забора верёвку с бельём, успешное сбрасывание с ограждения крыльца банок с комнатными растениями и на попытки вылить воду из лежащего шланга. По 5 мин в день обследовалась мусорная куча, и ещё 5 мин уходило на запрятывание найденных в ней предметов» (Дольник В.Р., 1995. Ресурсы времени и энергии у птиц в природе. СПб: Наука (Труды ЗИН. т.179). С.173.). Вопрос – как сочетается одно с другим, исключительное «напряжение жизни» и «бездельничанье», единственная цель которого — этот избыток «спустить», и «напряжение жизни», которое карает гибелью за малейшую неточность в выборе биотопа, партнёра и пр.?
Но чтобы реакция особей на управляющие воздействия со стороны социума или популяции как целого была эффективной и точной, необходим некий общий «язык», система сигналов. Она «притягивала» бы особей в места заведомого избытка ресурсов, где можно в полной мере развернуть видоспецифические социальные отношения, и «отвлекала» бы от попыток осесть в местах, где «напряжение жизни» уж чересчур.
Обычно всё занятое населением вида пространство (и ареал в целом, и отдельные местности, занятые соответствующими популяциями) анозотропны. В силу как истории расселения, так и сдвигов качества местообитаний в них выделяются градиенты противоположных изменений экологической и социальной среды. Скажем, в видовом ареале в целом прослеживается градиент «центр — периферия»; от первого ко второй растёт экстремальность среды обитания и падает напряжённость конкуренции за территорию, партнёра и пр. социальных взаимодействий. Точно такой же градиент прослеживается в большинстве заселённых видом местностей, только идёт не в направлении расселения, а от излюбленных («лучших») местообитаний к пессимальным. См. рис. 6А.
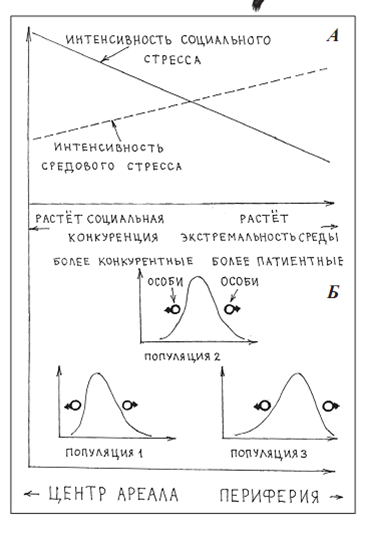
Популяционная система вида согласно «морфологическому подходу»: градиенты «центр – периферия» ареала (А) с сортировкой особей по потенциям (степень развития конкурентной и патиентной стратегий) вдоль них (Б).
Соответственно, особям «более конкурентным», более устойчивым к социальному стрессу, «любящим и умеющим» побеждать в соответствующей конкуренции, лучше концентрироваться на одном полюсе таких градиентов («в центре», -ах). И наоборот, особям, бегущим конкуренции с себе подобными в плотных поселениях, проигрывающим её но устойчивым к экстремальности внешней среды (как периферийные самцы тетерева в сравнении с центральными, куропёрые — мухоловок — пеструшек в сравнении с чёрно-белыми), лучше жить «на периферии».
Родиться же каждый из них может где угодно (рис.6Б); поэтому исключительно важная задача — направленное попадание особи в ту среду (экологическую и социальную), которая наиболее соответствует её потенциям, поведенческим и прочим, минимизация неопределённости и риска на этом пути.Что есть операциональная формулировка проблемы приспособления полиморфной популяции к мозаичной и флюктуирующей среде (особенно в плане устойчивости воспроизводства, а не «использования средовых изменений»).
Ортодоксальный СТЭшный взгляд на эту проблему именно таков, как сформулировал Б.Б.Жуков: особи разных потенций перемещаются по ареалу популяции более-менее случайно или в лучшем случае — по направляющим линиям ландшафта. Им «не дано знать, куда лучше меняться», поэтому пробуют во все стороны, что если и ограничивается, то только внешними барьерами. Кто-то добирается до нужного «пятна» в приемлемый срок, кто-то нет, «невидимая рука» отбора поддерживает первых и вычёркивает вторых, одновременно распространяя в популяции признаки, влияющие на «добирание вовремя» (и на успешные взаимодействия в найденной группировке).
Эта идеализация хороша лишь как нулевая гипотеза. Ведь реальные популяции (как минимум птиц и млекопитающих) включают в себя сигнальное поле, благодаря которому особи разных потенций направленно добираются в «пятна» экологической и социальной среды, благоприятной именно им. Всё происходит значительно быстрей и с меньшим грузом элиминации, чем в «классическом варианте» (что лучше всего видно опять-таки на примере урбанизации «диких» видов).
Сигнальное поле производит дистанционное наведение, благодаря которому чем социальнее вид, тем важней для него «сортировка вместо отбора» (для «несоциальных» видов сказанное bbzhukov верно без поправок или изъятия). Последний конечно, никуда не исчезает, но теряет творческую роль, остаётся только стабилизирующим. Переставая быть «делателем признаков, влияющих на добирание» он поддерживает видовую норму — формы демонстраций и формы отношений, совершенствует коммуникативные реакции на них. Т. е. отбор лишь «очищает», а содержательные изменения популяционной структуры чем дальше, тем больше производятся «сортировкой» (также как изменения морфологии/физиологии индивидов — эффектом Болдуина).
Всё сказанное про перемещения особей в «нужную» группировку верно для социальных (и иных) взаимодействий внутри неё с компаньонами и конкурентами. Роль сигнального поля здесь выполняет коммуникативная сеть, о которой писал уже многажды; она также создаёт «дальний порядок», предоставляя особям информацию о значимых сигналах и событиях за пределами непосредственного окружения, и теми же точно последствиями для выбора моделей поведения в будущем — его формы, места и времени, только в более узком хронотопе. Благодаря этим системам особь-таки «знает» куда ей стоит двигаться и, по прибытии, как себя вести — не всегда и во всём, но в некоторых существенных (с т.з. прошлой истории) аспектах и на некоторый период вперёд — достаточно точно.
Думаю, в начале эволюционного процесса у живого действительно было «незнание, в какую сторону лучше меняться», «часовщик» был совершенно «слепым», и процесс изменений «тыкался во все стороны». Но чем дальше, тем больше, у него «прорезаются глазки»: сперва развиваются инерционность и направленность филогенеза, затем появляются «предпочтительные» и «запрещённые» направления изменений, и, наконец, с появлением положительной обратной связи между сложностью социальной организации и развитием индивидуальности звери-птицы и фактически «берут эволюцию в свои руки».
Эти процессы идёт одновременно с двух сторон. Во-первых, чем более развиты, многоуровневы и сильны внутрипопуляционные регуляторы (сигнальные поля и коммуникативные сети), тем более значима в формировании стратегий поведения сортировка особей по потенциям, отбор выступает лишь «закрепителем». А дифференциация поведения между группировками рис.6 (или внутри них) «тянет за собой» физиологические и дальше морфологические изменения — с понятными поведенческими последствиями (п.19). Во-вторых, сейчас подробно изучены факторы, реализующие «ламарковские эффекты» без всякого ламаркизма.
Отсюда понятно, что возрождение номогенетических моделей эволюции связано с изучением предпочтительных направлений развития, созданных системной природой эволюционирующих объектов и «надстраивающихся» над стохастикой селектогенеза. Она бы действовала без видоизменения или изъятия, если бы каждый вид был представлен «россыпью» особей (ещё лучше — «эгоистичных репликаторов» без тел), друг с другом никак связанных и взаимодействующих= исключительно эксплуатационной конкуренцией через среду. Увы, они связаны, причём двояко — через тела, общевидовая морфология которых «строится» в ходе онтогенеза; и социальными и т.д. отношениями в популяцию, пространственно-этологическая структура которых «строится» в ходе коммуникации. А дальше «построенное» устойчиво воспроизводится, для чего управляет перемещениями, взаимодействиями и формированием поведения особей. И чем лучше это делает, тем больше включает сортировку и исключает отбор как творческий фактор в появлении нового (что и есть эволюция).
Иными словами, для возрождения номогенеза не стоит бесплодно растрачивать силы на отрицание эволюционной роли отбора (понятно, что он действует). Надо, приняв «селектогенез» за нулевую гипотезу, исследовать случаи, когда приспособительные изменения совершаются сильно быстрей, с существенно меньшим грузом элиминации или иным способом (прямым приспособлением, через стресс-реакцию, но не из-менением признаков, «связанных» с действующим давлением среды). Т.е. когда проявляется системность эволюционирующего объекта, по типу «Ударь кобылу в нос, она махнёт хвостом». Что самое замечательное, в этом случае можно исследовать номогенез в «заповедных угодьях» СТЭ — в области микроэволюции.
Примечания
1Откуда взяты все цитаты. См. также материалы конференции по сигнальному полю (2013 г.), где описывается его структура и функционирование у разных видов млекопитающих.
2Растительность той же территории сходным образом «наследует» ландшафтное разнообразие, бывшее в прошлом. «Органические сообщества действительно прямо включают и видоизменяют («ассимилируют») элементы неживой среды — почву, воздух, водоёмы, и эти элементы прослеживаются («наследуются») в живом покрове даже тогда, когда уже перестают существовать как таковые (например, заросшее сплавиной озеро долго ещё выделяется среди окружающего ландшафта совершенно иной растительностью)». Жерихин В.В. Искажение мира// Нева. 1991.
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Используются ли «понятия» в коммуникации животных?
Используются ли «понятия» в коммуникации животных? Об истории, культуре и современных проблемах коми. Часть 3.
Об истории, культуре и современных проблемах коми. Часть 3. Год — с винтовкой и плугом
Год — с винтовкой и плугом Виртуальные миры: анализ содержания психологических эффектов аватар-опосредованной деятельности
Виртуальные миры: анализ содержания психологических эффектов аватар-опосредованной деятельности О борьбе в науке
О борьбе в науке Об истории, культуре и современных проблемах коми. Часть 2.
Об истории, культуре и современных проблемах коми. Часть 2. Влияние озеленения и благоустройства на микроклимат
Влияние озеленения и благоустройства на микроклимат
