
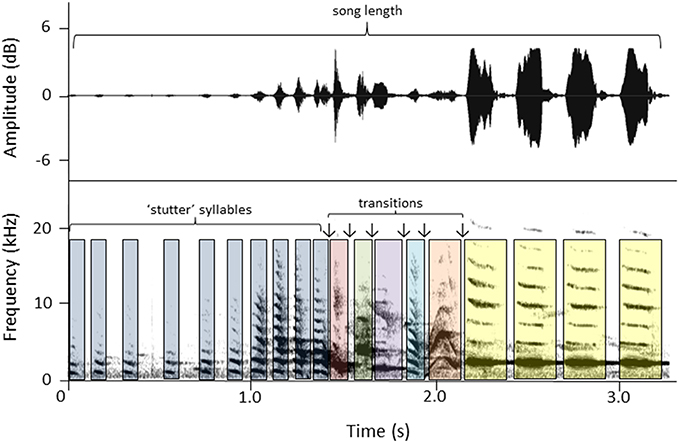
Сонаграмма самочьей песни новозеландской птицы-колокольчика Anthornis melanura. Источник
Концептуальный кризис в сравнительной этологии 1980-х гг. диагностирован многими авторами, стоящими на самых разных теоретических позициях, но указывающими одни и те же симптомы (Dawkins M.S., Halliday T.R., Dowkins R., 1991. The Tinbergen’s legacy; Панов Е.Н., 2005. Судьбы сравнительной этологии; Фридман В.С., 2013. От стимула к символу. Сигналы в коммуникации позвоночных). Так что кризис вполне достоверен и, увы, не преодолён по сей день: отсюда нынешняя неопределённость представлений об инстинкте, сигнале, сигнальной информации, демонстрациях, составляющих видовой репертуар – вместо чёткости и законченности классического период 1930-1960-х гг. Также как множественность концепций коммуникации.
Замечу, что это – один из многих примеров «некуновского» развития науки: общая парадигма («программа Лоренца – Тинбергена») не то чтобы не выдержала критики, социобиологов с одной стороны, Е.Н.Панова с другой, но «задержалась с ответом» (работы Вольфганга Шлейдта и Илана Голани конца 1970-х — начала 1980-х) на 10 лет, ставших критическими. «Дух отлетел», и интерес исследователей поведения переместился другие области. В которых новой парадигмы, сравнимой с этологической по общности, отнюдь не возникло, а пошло борение частных теорий и дробление их на всё более частные.
Почему так вышло, почему продолжается уже 40 лет, — скоро об этом напишу особо.
Здесь существенно, что один из симптомов кризиса – когда сторонники разных школ или разных теорий изучают одну и ту же проблему на одном и том же виде (-ах), он получают прямо противоположные «ответы» (естественно, в пользу своей теории). Так было в 1980-х гг., когда исследования воспроизводства разнообразия песенных диалектов у белоголовой воробьиной овсянки разными школами описывал В.В.Иваницкий. Что сохраняется и доселе; скажем, в вопросе, с чем связано пение самок воробьиных, первично оно для них или вторично. K.J.Odom et al. в 2014 г. опубликовали аж в Nature Communications статью, что первое, см. пересказ Алексея Опаева на Элементах.ру. А несколько позже И.Р.Бёме и М.Я.Горецкая на том же самом материале показывают, что второе, см. статью:
Пение самок воробьиных птиц: исключение или закономерность?
Содержание
Вокализацию воробьиных (отряд Passeriformes) подразделяют на позывы — короткие, простые звуки, издаваемые в самых разнообразных ситуациях в течение всего года; и песню, которая имеет большую длительность, сложную структуру и связана в основном с репродуктивным периодом в жизни птиц (Catchpole. Slater, 2008).
Происхождение песни исторически связывают е половым поведением самцов, а именно привлечением самки (Дарвин, 1951). Предполагали, что эволюция песни шла по направлению от вахтового позыва к сложной территориальной песне, через стадии вовлечения в структуру песни различных позывов и постепенной замены их в репертуаре на другие сигналы (Симкин, 1982).
Обладая сложной структурой, песня воробьинообразных выполняет разные, иногда противоречивые функции (Ильичев, 1972). Основными функциями песни считают:
1) обеспечение встречи половых партнеров, привлечение самки и, возможно, консолидация пары (Мальчевский, 1965; Thorpe. 1961; Catchpole, Slater, 2008; Cooney. Coekburn, 1995);
2) оповещение о занятости территории (Thorpe, 1961) и поддержание структуры поселения (Krebs, 1976; Catchpole, Slater, 2008). Роль песни (пения) состоит в том, что она сигнализирует получателям о мотивационном состоянии, статусе, индивидуальности, популяционной и видовой принадлежности поющей особи (Catchpole, Slater. 2008).
Индивидуальное и популяционное распознавание особей по голосу играет большую роль в различных социальных взаимодействиях, связанных с территориальным и брачным поведением птиц. Оно может влиять на выбор брачного партнера, осуществляемый самками (Baker, 1983), а также на уровень агрессии. проявляемой самцами при территориальных конфликтах (Catchpole. Slater, 2008). Основой для индивидуального и популяционного вокального распознавания особей является индивидуальная и географическая изменчивость песни.
Однако исторически работы по изучению песни проводили в Европе, где у большинства видов поют только самцы, и поэтому все гипотезы о роли песни высказаны применительно к ним. Хотя даже в умеренном поясе, по последним данным, пение самок не является такой уж редкостью (Garamszegi et al.. 2007). Есть виды, у которых самки ноют регулярно, и виды, у которых в тех или иных случаях отмечено их пение. В последнее время в литературе начали появляться статьи, посвященные пению самок, авторы которых указывают, что оно выполняет те же функции, что и пение самцов (Cooney, Cockburn, 1995).
Была выдвинута гипотеза, что в тропических широтах пение самок распространено гораздо шире (Kroodsma et ai., 1996; Morton, 1996; Langmore, 1998; Slater, Mann, 2004) и может быть скорее нормой, чем исключением из правил (Morton. 1996). В 2014 г. вышла статья Одом с соавторами (Odom et al., 2014). в которой авторы на основе анализа песен 1141 вида утверждают анцестральность признака пения самок в отряде воробьинообразных.
Формирование яркой красной, желтой или рыжей окраски, основанной на каротиновой пигментации, определяется содержанием каротиноидов в пище (Hill, Montgomerie, 1994; Fitze et al., 2003). Как следствие в процессах эпигамного отбора вариация подобной окраски может свидетельствовать о средовых влияниях и сигнализировать о качестве особи (Hamilton, Zuk, 1982; Horak et al., 2006). Для европейских видов была показана связь между наличием дихроматизма в окраске, обеспеченной липохромами, и пением самок (Garamszеgi et al.. 2007).
В нашей работе мы проанализировали случаи пения самок в разных семействах отряда воробьиных на разных материках, с целью понять эволюционные предпосылки этого феномена и его связь с особенностями биологии видов и путями распространения основных групп воробьинообразных.
Материалы и методы
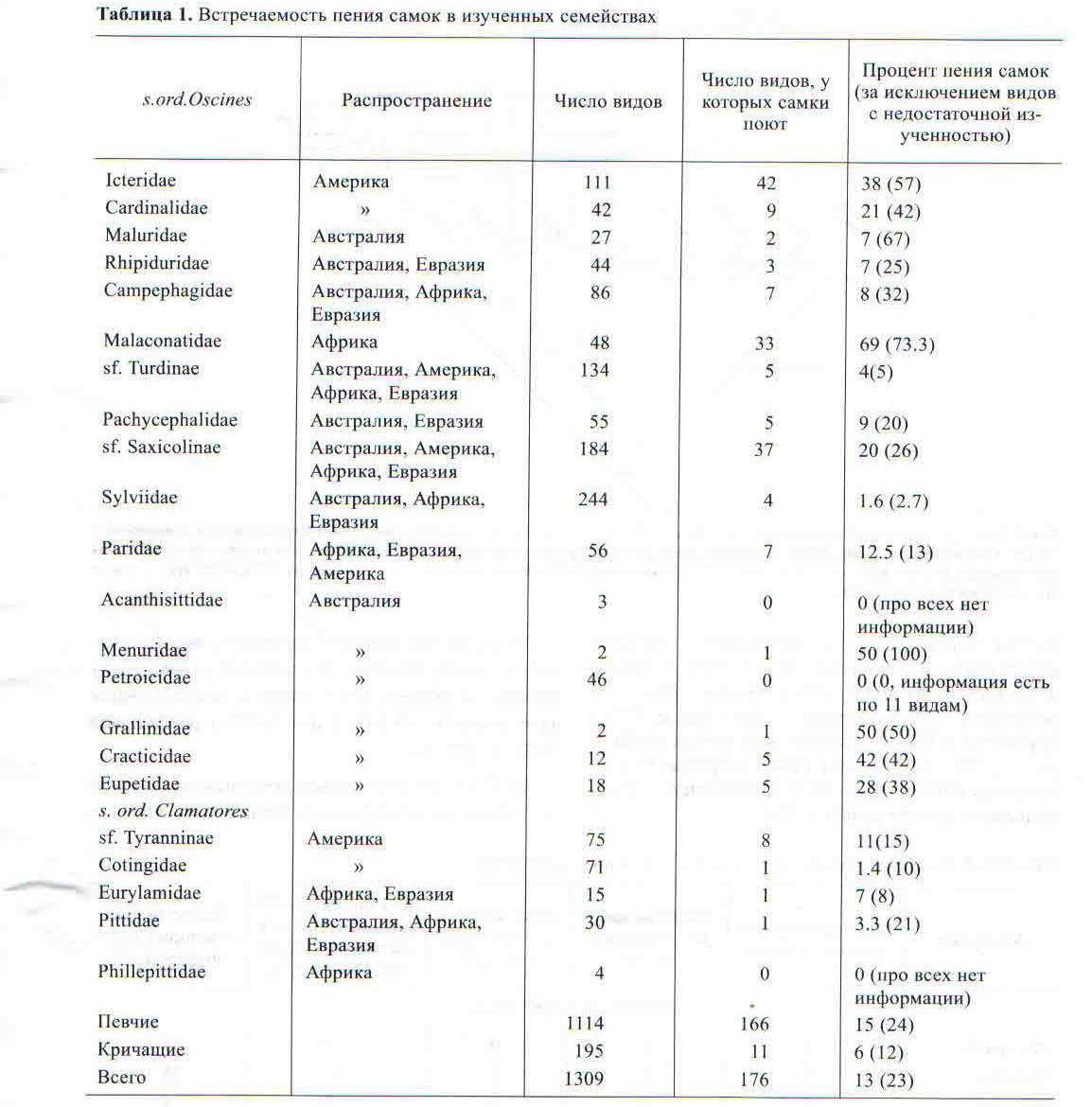
Мы проанализировали случаи пения самок воробьинообразных птиц, описанные в Handbook of the Birds of the World (Edited J. del Hoyo et al., 2004-2011), а также наши собственные данные, собранные во время экспедиции в штате Виктория (Австралия 2009 г.) и в Бразилии (2014 г.). В сводке приведена детальная информация о поведении и особенностях экологии, включая вокализацию, всех видов воробьинообразных птиц. Несмотря на то что очерки по каждому семейству и его представителям были написаны разными авторами, они выдержаны в едином стиле и основаны на современных данных. Мы проанализировали информацию по певчим воробьиным (четырнадцать семейств целиком, два подсемейства у дроздовых и три у славковых) и кричащим воробьиным (четыре семейства целиком и два подсемейства у тирановых) (табл. 1), всего проанализировано 1309 видов.
Семейства были выбраны так, чтобы в разных частях света число изученных видов было сходным. Мы придерживались систематики отряда воробьинообразных, принятой в этой сводке. Число видов в нашей работе превышает их количество в других аналитических статьях, посвященных пению самок (233 вида у Гарамзеги с соавторами (Garamszcgi et al., 2007) и 1141 в — работе Одом с соавторами (Odom et al., 2014). Наличие пения у самок считалось установленным, если в описании вокализации вида было оговорено, что самки поют, а его отсутствие — если в видовом очерке было указано, что поют только самцы. Нее остальные случаи рассматривали как отсутствие информации о пении самок. Отмечали наличие полового диморфизма, песенных дуэтов; у обоих полов по Гарамзеги с соавторами (Garamszеgi et al., 2007) отмечали тип окраски; меланиновая (коричневый, черный, красно-коричневый цвет оперения) и липохромная (желтый, оранжевый, красный); географическое распространение и наличие миграций. Для видов, у которых известно пение самок, оценивали уровень изменчивости песни самца и самки, ориентируясь на описание длительности песни и разнообразия элементов в ней (1 — простая (от одного до нескольких слогов, как у пеночки-трещотки Phylloscopus sibilatrix), 3 — очень изменчивая (много фраз, состоящих из разных слогов, как у соловья Luscinia luscinia), 2 — промежуточный вариант, как у зяблика Fringilla coelebs).
Чтобы исключить случаи недостаточной изученности вида в целом, мы проводили два варианта анализа данных. Первый вариант анализа включал оценку встречаемости пения самок у всех проанализированных видов. Второй вариант анализа включал только те виды, у которых описание вокализации в сводке занимало больше трех строк. Виды, но которым описание вокализации занимало меньше трех строк, были включены в анализ, если имелось указание о пении самок. В остальных случаях их нe учитывали, считая, что отсутствие информации о пении самок связано с малой изученностью вида в целом. Мы считаем, что второй вариант анализа может даже несколько завышать долю пения самок. Из всех изученных видов информация о пении самок в абзаце, содержащем три строки и меньше, была обнаружена 31 раз.
Для выявления связи между пением самок и наличием дуэтов, образом жизни (наличие или отсутствие миграций), половым диморфизмом и окраской использовали коэффициент корреляции Спирмeна (Лакин, 1990). Различия считали достоверными при р < 0.05.
Результаты исследования
Среди исследованных семейств в отряде воробьинообразные доля видов, у которых поют самки, составила 13%, за исключением видов с недостаточной изученностью — 23%. Среди представителей певчих воробьиных пение самок было отмечено у 15% видов, за исключением видов с недостаточной изученностью — 24%, среди кричащих оно отмечено у 6%, а за исключением видов с недостаточной изученностью — 12% (табл. 1).
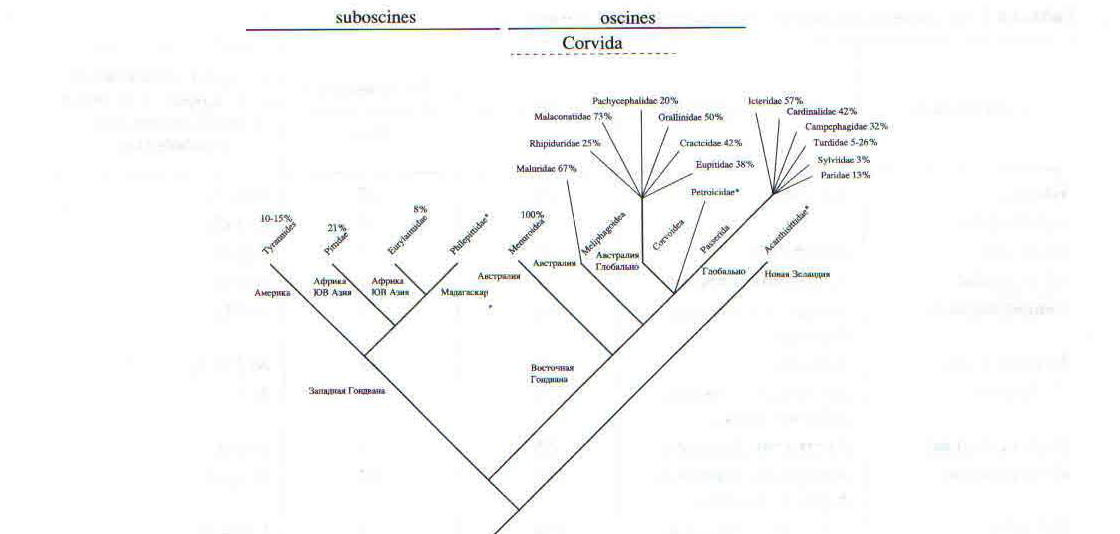
Рис. 1. Модифицированная кладограмма основных групп воробьиных на основании данных филогенетических отношений и биогеографии (Ericson et al.. 2002). Дополнительно укатаны проанализированные нами семейства и отмечен процент пения самок в них с учетом изученности видов. для всех видов семейства информация но пению не превышает трех строк, и нет указаний на пение самок.
Наши результаты показали высокий процент случаев пения самок в некоторых семействах певчих воробьиных, за исключением видов с недостаточной изученностью, до 100% у птицы-лиры (всею два вида), 73% у кустарниковых сорокопутов, 67% у австралийских славок, 57% у трупиалов и 50% у сорочьих жаворонков (табл. 1, рис. 1). Реже всего пение самок встречается в семействах синиц — 13% видов, славковых — 3%, в подсемействе дроздовых — 5%.
Среди представителей кричащих воробьиных пение самок, видимо, встречается реже, чем у певчих, за исключением видов с недостаточной изученностью, от 21% у питт до 8% у рогоклювов (табл. 1, рис. 1).
Наиболее широко, за исключением видов с недостаточной изученностью, пение самок у певчих воробьиных представлено в Америке — 37%, чуть реже в Африке — 33%, еще реже в Австралии и почти отсутствует в Евразии (табл. 2). Среди представителей кричащих случаи пения самок отмечены только в Америке и Евразии.
 В целом пение самок чаще встречается у не мигрирующих видов (83%, певчие, 100% кричащие), хотя у перелетных оно тоже встречается не так уж и редко (17% певчие) (табл. 3).
В целом пение самок чаще встречается у не мигрирующих видов (83%, певчие, 100% кричащие), хотя у перелетных оно тоже встречается не так уж и редко (17% певчие) (табл. 3).
|
Признаки |
Певчие (1114) |
Кричащие (195) |
|||
|
самки поют |
самки не поют |
самки поют |
самки не поют |
||
|
Миграции есть |
29 |
182 |
0 |
20 |
|
|
Миграций нет |
137 |
766 |
п |
164 |
|
|
Половой диморфизм есть |
79 |
339 |
3 |
83 |
|
|
Полового диморфизма нет |
87 |
609 |
8 |
100 |
|
|
Дуэты есгь |
84 |
0 |
7 |
0 |
|
|
Дуэтов нет |
82 |
947 |
4 |
184 |
|
|
Липохромы есть |
110 |
663 |
9 |
164 |
|
|
Липохромов нет |
56 |
285 |
2 |
20 |
|
|
Половой димор- |
пд нет |
Половой диморфизм |
пд нет |
||
|
физм (пд) есть |
(пд) есть |
||||
|
Липохромы есть |
309 |
464 |
78 |
95 |
|
|
Липохромов нет |
109 |
232 |
9 |
13 |
|
В целом для всех исследованных видов выявлена слабая положительная связь между наличием полового диморфизма и пением у самок (корреляция по Спирмену, R = 0.07; р = 0.01) (табл. 3). Среди видов, у которых поют самки, половой диморфизм отмечен в 48% случаев для певчих и 27% для кричащих воробьиных. Для кричащих воробьиных дуэтное пение было отмечено у 64% видов, у которых известно пенис самок, у певчих воробьиных — 51% от общего числа случаев пения самок. Дуэты в большей степени характерны для мономорфных видов (61% для певчих и 86% для кричащих). Однако дуэтное пенис отмечено и для довольно большого числа видов, имеющих половой диморфизм (39%). Дуэтное пение чаще встречается у оседлых видов (отрицательная корреляция с наличием миграций, по Спирмену, R = -0.3, р< 0.001) (табл. 3)).
Нами показана положительная связь между степенью изменчивости песни у самцов и у самок (положительная корреляция но Спирмену, R = 0.6, р < 0.001).
Меланин присутствует в окраске всех изученных видов. Среди видов, у которых отмечено пение самок, липохромовые пигменты в окраске встречаются у 72%. На основе анализа 1309 видов нами не выявлено связи между наличием ли- нохромов в окраске и пением самок (табл. 3).
Обсуждение результатов
В целом, по нашей оценке, доля пения самок в отряде воробьинообразных не превышает 30%. Процент пения самок широко варьирует между разными семействами, даже в тропических широтах есть семейства, где пение самок встречается очень часто до 100% и достаточно редко 5-25%. Однако несомненно этого вполне достаточно, чтобы разрушить устоявшиеся представления о пении, как о прерогативе самцов.
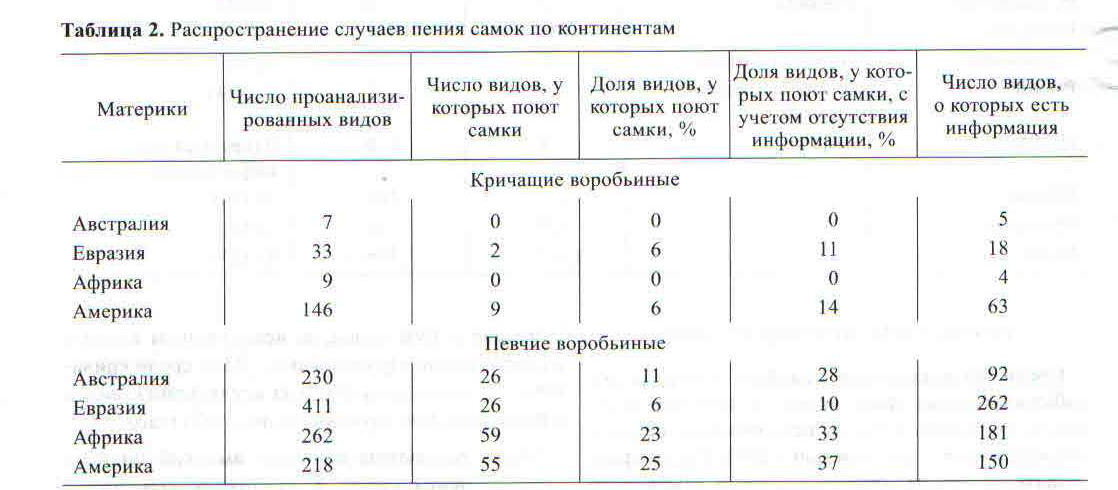
Географическое распространение случаев пения самок позволяет выдвинуть гипотезу о путях становления этого явления у воробьинообразных птиц. Согласно данным Эриксона с соавторами (Ericson еt al., 2002), становление и распространение кричащих и певчих воробьиных шло после распада Гондваны в разное время и разными путями (рис. 1, 2). Наш анализ показал, что в Австралии доля пения самок у певчих воробьиных составляет 28%, менее всего пение самок распространено в Евразии (10%) и максимума оно достигает в Африке (33%) и Америке (37%). Разные ветви певчих воробьиных вначале заселили Австралию, потом Евразию, затем Африку и наконец Северную и Южную Америку (рис. 1, 2). Разные ветви кричащих воробьиных вначале заселили Африку и Азию и лишь затем Южную и Северную Америку.
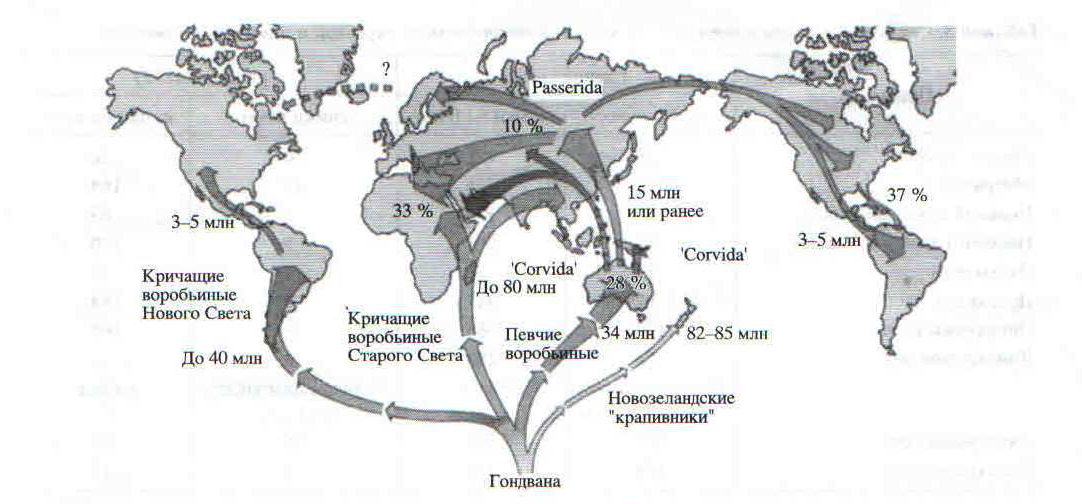
Рис. 2. Предполагаемые пути дисперсии основных групп воробьиных из Гондваны на основании данных филогенетических отношений и биогеографии (Erieson et al., 2002). «млн.» означает “миллионов лет назад». Проценты показывают встречаемость песни самок у певчих воробьинообразных на разных материках.
Нами в Евразии отмечено 11 % пения самок, в то время как в более молодых семействах в Америке пенис самок встречается в 14% случаев. Таким образом, пенис самок чаще встречается в более молодых группах как кричащих, так и певчих воробьиных.
Полученные нами результаты практически полностью расходятся с данными Одом с соавторами (Odom ct al., 2014), в которых для шести семейств, исследованных как нами, так и ими, процент поющих самок различается больше, чем в 50 случаях. И это тем более странно, что авторы ссылаются на тот же источник информации (HandBook of the Birds of the World (Edited J del Hoyo et al., 2004-2011)), и по их описанию применяли те же критерии наличия пения у самок. Так, для австралийских зарянок мы не обнаружили ни одного случая пения самок, в то время как Одом с соавторами отмечают около 50% таких случаев, сходная картина и с другими семействами. Возможно авторы использовали и другие источники, не оговоренные в статье. Такое явное несоответствие в данных позволяет критично отнестись и к выводам авторов статьи.
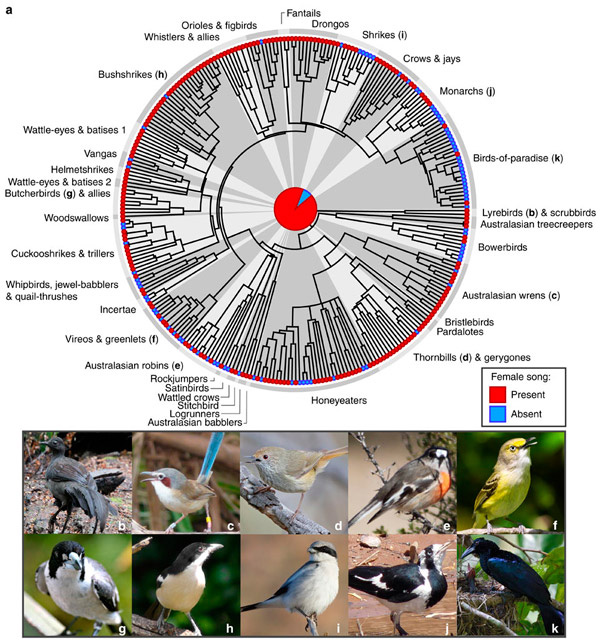
Противоположные выводы (об анцестральности пения самок) Odom et al., 2014, полученные анализом тех же данных и по тем же критериям. Источник.
По данным литературы сложно оценить степень выраженности пения самок, а именно, являются ли описанные случаи единичными или самки поют регулярно. Так, по данным Гарамзеги с соавторами (Garamszegi et al., 2007), из 223 европейских видов у 101 (45%) отмечено пение самок, хотя очевидно, что для одних видов такое пение является нормой, а у других это единичные случаи, подтверждающие принципиальную способность самок петь. При рассмотрении континента Евразии наш анализ показал, что самки поют только у 11% видов, несмотря на то, что это один из наиболее хорошо изученных материков.
В свою очередь причины пения самок, видимо, действительно связаны, как предполагалось ранее, с приуроченностью к теплому климату и оседлому образу жизни (Price, 2009; Price et al., 2009), это подтверждает и наш анализ. Дуэтное пение нами было выявлено только в 51% от случаев пения самок в отряде воробьинообразные. Видимо, как дуэтное пение, гак и соло самок возникают независимо.
Поданным Гарамзеги с соавторами (Garamszegi ct al., 2007), анализировавшими европейские виды, пение самок связано с наличием полового диморфизма в окраске, причем только в тех случаях, когда окраска обусловлена липохромовыми пигментами. Выраженность последних более подвержена воздействию внешних факторов и связана с физиологическим состоянием особи, наличие липохромовой окраски, по мнению Гарамзеги с соавторами, может являться честным показателем качеств ее носителя. Нами не выявлена связь между пением самок и наличием липохромов в окраске, однако обнаружена положительная связь пения самок с наличием полового диморфизма, возможно полученные Гарамзеги с соавторами данные также связаны не с окраской как таковой, а с выраженностью полового диморфизма. К сожалению, в обсуждаемой статье авторы не разделяли на отдельные переменные: наличие полового диморфизма и пигменты, формирующие окраску, поэтому они не могли проанализировать эти признаки раздельно.
Пение самок появляется, как правило, у оседлых популяций и предпочтительно в тропическом климате. Причем причины его возникновения у конкретных видов не всегда очевидны. Так, среди австралийских славок у двух видов самки поют не хуже самцов и песня несет территориальную функцию (Cooney, Cockburn, 1995), в то время как у 25 других представителей этого семейства, обладающих сходным образом жизни, пение самок не отмечено. Сходная картина обнаружена у австралийских свистунов и трупиалов, у которых пенис самок распространено довольно широко, а близкие виды отличаются по этому признаку.
Результаты нашего анализа показывают, что, в отличие от самцов, пение самок чаще встречается в более молодых группах воробьинообразных, и не поддерживают гипотезу о том, что пение является анцестральным признаком для самок. Видимо, пение у самок независимо возникало в разных семействах, причем для каких-то семейств (например, трупиаловые согласно Прайсу (Price, 2009; Price et al., 2009)) оно действительно является анцестральным.
Вокальные сигналы изначально характерны для обоих полов в классе птицы, во многих отрядах распространены вокальные дуэты (Брагина. Беме, 2010). Поскольку центральные вокальные моторные и сенсорные механизмы взаимосвязаны, отбор на пение ведет к усилению способности воспринимать вокальные сигналы и петь. В отряде воробьинообразные пение получило наибольшее распространение и очевидно, что отбор на пение и восприятие песни шел не только среди самцов, но и среди самок. Известно, что у самок в процессе онтогенеза также развиваются вокальные ядра головного мозга, а при стимуляции тестостероном в норме непоюшие самки начинают петь (Catchpole, Slater, 2008). Таким образом, способности самок к пению должны развиваться параллельно со способностями самцов, а проявление этого признака зависит от конкретной ситуации. Самки кричащих воробьиных после проникновения в Старый Свет практически утрачивают способность к пению (надо заметить, что песни и самцов этих видов крайне примитивны и просты), в то время как в Америке доля поющих самок существенно выше. Для певчих воробьиных наблюдается сходная картина. Таким образом, пение самок независимо проявляется в различных семействах воробьинообразных птиц как кричащих, так и певчих.
Авторы благодарны сотрудникам университета Монаш (Австралия) Александре Павловой и Полю Санок и университета Дикон (Австралия) Кейт Бучанон за предоставленную возможность работать в составе их экспедиции в штате Виктория в Австралии. Мы благодарим сотрудников кафедры зоологии позвоночных Биофака МГУ В.В. Иваницкого и И.М. Марову за предоставление литературных источников.
Работа выполнена при поддержке гранта РФФИ № 14-04-00108а, 15-29-02771 офи м и гранта РИФ 14-50-00029.
Список литературы
Брагина Е.В.. Беме И.Р., 2010. Дуэты у животных почему птицы поют хором? // Природа. № 6. С. 13— 18.
Дарвин Ч.. 1951. Происхождение человека и половой отбор. Соч. в 8 томах. Т. 5. М.; Л. Изд-во АН СССР. С. 199-658.
Ильичев В.Д.. 1972. Биоакустика птиц. М.: Изд-во МГУ. 468 с.
Лакин Г.Ф., 1990. Биометрия. М.: Высш. шк. 350с.
Мальчевский А.С., 1965. К вопросу о голосовой имитации у птиц // Сложные формы поведения. М.; Л.: Наука. С. 139-143.
Симкин Г.Н., 1982. Актуальные проблемы изучения звукового общения птиц// Орнитол. Т. 17. М.: Изд-во МГУ. С. 111 129.
Baker М.С., 1983. The behavioral response of female Nuttall’s White-crowned Sparrows to male song of natal and alien dialects // Beh. Ecol. Sociobiol. V. 12. Is. 4. P. 309 315.
Catchpole C.K.. Slater P.J.B.. 2008. Bird Song: Biological Themes and Variations. Cambridge: Cambridge Univ. Press. 335 p.
Cooney R., Cockburn A., 1995. Territorial defense is the major function of female song in the superb fairy- wren (Malurus cyaneus) H Anim. Behav. V. 49. № 6. P. 1635-1647.
Ericson P.G.P., Christidis L., Cooper A., /rested! M.. Jack- son J., Johansson U.S.. Norman J.A.. 2002. A Gondwanan origin of passerine birds supported by DNA sequences of the endemic New Zealand wrens // Proc. R. Soc. Lond. Scr. В. V. 269. P. 235-241.
Fitze P.S., KoUiker M, Richter II., 2003. Effects of common origin and common environment on nestling plumage coloration in the great tit (Parus major) // Evolution V. 57. P. 144-150.
Журнал общей биологии. 2016. Т.77. №3. С.239-246.
Рекомендуем прочесть
 Быстрая перестройка фенологии в новых местах гнездования ласточек
Быстрая перестройка фенологии в новых местах гнездования ласточек Что не так с критикой Ильенкова
Что не так с критикой Ильенкова Птицы документируют антропоцен: стратиграфия пластика из гнёзд лысухи
Птицы документируют антропоцен: стратиграфия пластика из гнёзд лысухи Восприятие шумов
Восприятие шумов Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла»
Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла» Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки «Жесты», вόроны и шалашники
«Жесты», вόроны и шалашники Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных
