![Print PDF Резюме публикатора. Когда в 1990е стали активно исследовать недавно возникшие необычно северные зимовки славки-черноголовки в Великобритании и Ирландии, выяснились, во-первых, адаптивное преимущество их использования сравнительно с обычными пиренейскими, […]](http://www.socialcompas.com/wp-content/uploads/2023/04/900-465x190.jpeg "Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки")

Самец славки-черноголовки
Резюме публикатора. Когда в 1990е стали активно исследовать недавно возникшие необычно северные зимовки славки-черноголовки в Великобритании и Ирландии, выяснились, во-первых, адаптивное преимущество их использования сравнительно с обычными пиренейскими, во-вторых, положительная ассортативность образования пар у использующих их птиц на местах гнездования в центральной Европе, вследствие сдвига прилёта на более ранние даты. В этом увидели начинающееся симпатрическое видообразование, тем более что последующие работы выявили заметную дивергенцию, морфологическую и генетическую, между среднеевропейскими птицами, прилетающими с разных зимовок, но гнездящимися на одной территории (сравнимую с дивергенцией разных географических популяций, разделённых сотнями км). Возможно, так и случилось бы, однако адаптивное преимущество британских зимовок, созданное регулярной подкормкой в садах, оказывается очень уж велико. Benjamin M. van Doren et al. (2021), исследовавшие зимнюю биологию славок в Великобритании и Ирландии, показывают, что сейчас тамошние зимовки «притягивают» птиц всей Европы, от гнездившихся в Скандинавии до гнездившихся в Средиземноморье.
Больше того, часть зимовавших там птиц вместо отлёта начала размножаться на самих островах, как в эти же десятилетия сделали ласточки-касатки Hirundo rustica и белолобые скалистые ласточки Petrochelidon pyrrhonota в Аргентине. Вместо ожидавшегося симпатрического разделения на 2 популяции славок Центральной Европы, новая зимовка на островах создала «плавильный котёл», перемешивающий птиц из географически разных районов Европы, расстояния между которыми сравнимы или больше тех, что обеспечили значимую дивергенцию аллопатрических популяций. Следовательно, особенности морфологии и экологии, созданные использованием новых зимовок, вопреки ожиданиям, не стали стимулятором формообразования и маркером отделившейся популяции.
Микроэволюционные следствия оказались обратными — интеграция популяционной системы вида на обширном пространстве Европы, а использование британских зимовок делается одной из возможных стратегий особей в разных популяциях, увеличивая их полиморфизм а, значит, устойчивость. Ещё раз подтвердилась ненадёжность и маловероятность симпатрического видообразования у птиц. Даже начавшись, оно не доходит с тем большей вероятностью, чем в большем спектр местообитаний могут дать выигрыш в приспособленности те адаптации, что исходно маркировали расщепление популяционных систем. В нашем случае это зимовка в Британии.
Резюме авторское. Человеческая деятельность глубоко влияет на мир природы.
Перелетные птицы особенно восприимчивы к её неблагоприятным последствиям, ибо глобальные сети экосистем, на которые они рассчитывают [при перелётах], очень быстро меняются. Вопреки этим трудностям, славка-черноголовка (Sylvia atricapilla) — процветающий перелётный вид. [Как мы уже писали, она даже выиграла от деятельности человека]. Уже более 40 лет данный вид пользуется высокоширотными зимовками в Великобритании и Ирландии. Их появление связано с изменением климата и массовой подкормкой птиц, выступая примером взаимодействия между человеческой деятельностью человека и биологией вида-мигранта. С целью исследовать, как антропогенные факторы формируют перемещения и экологию зимующих черноголовок, авторами помечены 623 особей в 59 пунктах Великобритании, а также собран набор данных из 9929 встреч [этого вида]. На этой выборке они изучили поведение славок при посещении кормушек, межгодовую привязанность к месту подкормки и их перемещения внутри сезона и между сезонами. На 25 птиц авторы установили геолокаторы и проанализировали миграционные треки, чтобы понять, как использование кормушек в садах влияет на миграцию и размножение в дальнейшем. Черноголовки, зимующие в Великобритании и Ирландии, высоко привязаны к конкретным местам зимовок, и редко перемещаются между ними. Славки, зимующие в традиционных местах на Пиренеях, напротив, всё время кочуют. Молодые, зимующие впервые [в Англии], меньше привязаны к конкретному месту зимовки и чаще меняют его, чем взрослые. Более частое посещение кормушек в придомовых садах даëт взрослым особям выигрыш в физическом состоянии, они менее жирные, с более длинными клювами и более закруглëнными крыльями. Однако черноголовки не питались исключительно на кормушках у домов, их посещения вызваны более суровой погодой. Особи обычно держались придомовых садиков до самого отлёта весной к местам гнездования. Результаты показывают, что подкормка [массовая и постоянная настолько, что на неë птицы «могут рассчитывать»] меняет зимнюю экологию черноголовок и изменяет их морфологию. Дополнительное кормление может иметь многогранные преимущества для выживания зимой, и эти положительные эффекты могут распространяться на миграцию и последующее размножение. Дополнительное кормление может дать разносторонние преимущества для зимнего выживания, ее положительные эффекты могут распространяться на миграцию и последующее размножение. В целом, высокая индивидуальная изменчивость передвижений и экологии кормодобывания черноголовок, а также гибкость, которую она придает [в целом биологии вида], вероятно, позволили ему процветать во время быстрых [антропогенных] изменений окружающей среды.
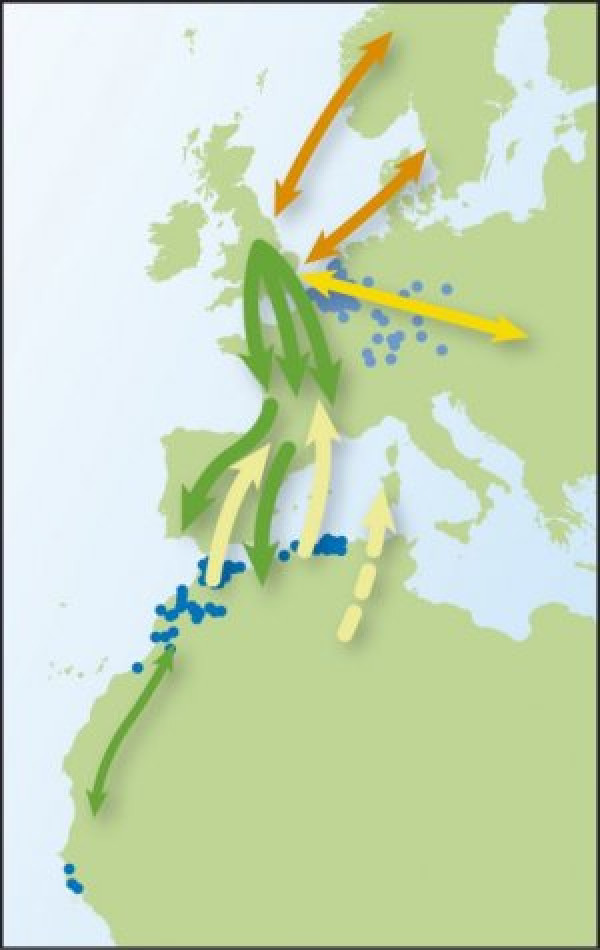
Пролётные пути славок — черноголовок Западной Европы. Зелёный — традиционный на юго-запад, желтый, оранжевый — вновь появившийся в Великобританию
Люди все больше [и/или сильнее] влияют на биосферу, [отчасти разрушая природные сообщества, отчасти преобразуя их]. Все виды биоты подвергаются целому ряду воздействий: [промышленного и городского] развития или же сельского хозяйства на исходные местообитания видов; изменений температурного режима и количества осадков, вызванных изменением климата; охоты или [других форм] промысла; а также светового и шумового загрязнения (Benítez-López et al., 2017; Gaston et al., 2013; Kunc et al., 2016; Mantyka-Pringle et al., 2012).
Организмы [разных видов] различны по своим способности приспосабливаться к средовой динамике. Неясно степень, в которой [разные виды] в сообществах смогут идти в ногу, в приспособительных ответах на изменения (Feeley & Rehm, 2012; Liang et al., 2018; Poloczanska et al., 2013; Urban et al., 2016). Перелетные птицы играют ключевую роль в здоровье экосистемы (Bauer & Hoye, 2014), но они особенно восприимчивы к изменениям окружающей среды, поскольку успешная миграция требует интеграции ряда различных этапов, каждый из которых чувствителен к текущим изменениям, а те бывают слабо коррелированы. Мигранты должны точно планировать свои маршруты; безошибочно ориентироваться [вопреки] атмосферной динамике; надёжно находить кормовые ресурсы [на остановках и зимовке], безопасно и эффективно их использовать; преуспевать в экологических условиях, меняющихся в зависимости от времени года и мест на земном шаре [куда летят птицы].
По мере того как изменение климата смещает оптимальные временные рамки, режимы ветра и характер штормов, а также по мере того, как люди изменяют как ландшафты, через которые пролетают птицы, так и районы, где они размножаются и зимуют, миграционная стратегия может становиться все более несостоятельной (Runge et al., 2015; Wilcove & Wikelski, 2008). Эти проблемы особенно остры для видов, которые мигрируют на большие расстояния (Sanderson et al., 2006). Численность перелетных птиц во всем мире сокращается, что вызывает опасения по поводу того, что многим может не хватать гибкости для быстрого реагирования и адаптации (Both et al., 2010; Rosenberg et al., 2019; Sanderson et al., 2006).
Главный вопрос современных исследований — могут ли мигранты [особенно дальние] приспособиться к быстрым изменениям [среды обитания]. У многих из них «расписание» миграций во времени и программы навигации врождëнные, что ограничивает их гибкость. Есть некоторые свидетельства микроэволюционных изменений в этих эндогенных программах, но неясно, «поспевают» ли они за темпами потепления (Charmantier & Gienapp, 2014; Helm et al., 2019; Merilä & Hendry, 2014; Van Buskirk et al., 2012).
Надёжно установлена и пластичность в ответ на изменения климата, особенно у ближних мигрантов (Gienapp et al., 2007; Usui et al., 2017), и все больше свидетельств указывает на разные проявления гибкости и у дальних мигрантов (Brown et al., 2021; Haest et al., 2020; Klinner & Schmaljohann, 2020). По мере продолжения изменений видам потребуется не только смещать «расписание во времени» для нескольких стадий годового цикла, но и претерпевать крупномасштабные изменения в распределении птиц внутри ареала, чтобы отслеживать [появление иди напротив исчезновение] подходящих условий [для зимовки или других стадий годового цикла] в связи с ними. Вызванные изменением климата сдвиги ареала были задокументированы у птиц (La Sorte & Thompson III, 2007; Lehikoinen & Virkkala, 2016; Tingley et al., 2012), но нам не хватает полного понимания способствующих им экологических и поведенческих процессов (Gill et al., 2019). Это создает сложности для прогноза будущего реагирования вида.
Евразийская славка-черноголовка (Sylvia atricapilla) — перелетная певчая птица, процветающая в условиях описанных изменений (EBCC/BirdLife/RSPB/ CSO, 2018). [Это один из видов с наиболее выросшей численностью за последние 30 лет в Европе] Миграционные стратегии этого широко распространенного вида варьируют от оседлости до полностью перелетного, при разных направлениях миграции (Roselaar, 1992). Кроме того, его европейская численность сильно выросла (на +155% с 1980 года, ЕBCC/BirdLife/RSPB/CSO, 2018) и продолжает расти.

Славка на кормушке ест сало
Наряду с этим, в последние полвека зимовочный ареал вида расширился на север, захватив Великобританию и Ирландии. Там зимний статус черноголовки изменился с редко залëтного (до 1960 года) на обычный и общепризнанный компонент зимней орнитофауны уже в 1980е (Berthold & Terrill, 1988; Leach, 1981). Его зимовочный ареал в Великобритании и Ирландии вырос на 77% в период с 1981-1984 по 2007-2011 гг., когда славок отметили этот вид в 48% квадратов со стороной 10 км, выбранных для создания атласа птиц. Эта трансформация связана с человеческой деятельностью двумя возможными способами (Berthold & Terrill, 1988; Plummer et al., 2015): во-первых, изменение климата привело к более мягким зимам, а во-вторых, ставшие многочисленными кормушки в садах у домов всю зиму предоставляют надежный источник пищи. Всего удивительней, что практически все славки, зимующие в Британии — отнюдь не из местной гнездовой популяции, но прибыли зимовать с континента, мигрировав в крайне нехарактерном (с.-з.) направлении (Berthold et al., 1992; Delmore et al., 2020; Wernham et al., 2002). Данные птицы отличны от летящих на традиционные зимовки в Средиземноморье: они используют новое направление миграции, зимуют в более высоких широтах, в местообитаниях, созданных человеческой деятельностью, и тесно связаны с дополнительным питанием на кормушках и плодами в садах. Черноголовки, зимующие в Британии и Ирландии, адаптировались ко многим характерным особенностям антропоцена, понимание детерминантов их успеха покажет, что нужно перелетным птицам, чтобы преуспеть перед лицом глобальных изменений.
В работе исследованы экология и поведение черноголовок, зимующих в Великобритании и Ирландии. Авторы используют данные о повторных обнаружениях окольцованных птиц, и подробный набор данных об отдельных поимках и перемещениях индивидуально окрашенных птиц, чтобы понять использование ими местообитаний и ресурсов в придомовых садах. Также исследовано, как на их поведение влияют местные условия и [как это влияние] опосредуется физическим состоянием и морфологией птиц.
Предположительно черноголовки зимой не полностью зависимы от дополнительного питания, но это спасательный круг в сложных условиях. Авторы также оценивали привязанность разных особей к их месту зимовки в разные зимы. На зимовках Средиземноморья и Африки она крайне низка, доля повторных отловов всего лишь 0– 5% (Cuadrado, 1992; Cuadrado et al., 1995; King & Hutchinson,).
 Рисунок 1. (а) Места мечения и число зимующих черноголовок, индивидуально помеченных цветными кольцами (размер круга, см. легенду справа). (b) Пойманный самец славки-черноголовки с цветным кольцом. (c) Другой самец в саду с геолокатором, отслеживающим уровень освещенности. Фотографии Бена Портера.
Рисунок 1. (а) Места мечения и число зимующих черноголовок, индивидуально помеченных цветными кольцами (размер круга, см. легенду справа). (b) Пойманный самец славки-черноголовки с цветным кольцом. (c) Другой самец в саду с геолокатором, отслеживающим уровень освещенности. Фотографии Бена Портера.
Поэтому изучали, развили ли славки, зимующие в Британии, большую верность конкретным местам зимовки, чтобы воспользоваться преимуществами более надежных мест подкормки в садах. Авторы использовали данные кольцевания для исследования перемещений славок между зимами и в каждую из зим, комбинируя их с прослеживанием отдельных особей, чтобы выявить места размножения славок, зимующих в Британии. Далее авторы сравниваем записи наблюдений и треки геолокаторов, чтобы проверить гипотезу о том, что высококачественная пища, доступная в садах, играет важную роль в поддержке [нового пути] миграции.
2. Методы
Содержание
- 1 2. Методы
- 2 3. Результаты
- 3 4.Обсуждение
- 3.1 4.1. Влияние человека на поведение черноголовок
- 3.2 4.2. Потенциальные будущие изменения в поведении
- 3.3 4.3. Велика ли зависимость славок от дополнительного питания?
- 3.4 4.4. Индивидуальные различия и гибкость
- 3.5 4.5. Происхождение [птиц], зимующих в Великобритании и доказательства резидентности их на кормушках
- 4 5. Заключение
2.1. Индивидуальное мечение и перемещения.
Зимующих черноголовок ловили и кольцевали на 59 участках в Великобритании и Ирландии (рисунок 1a; таблица S1) в период с ноября по апрель (рисунок 1b). Ловили их в основном в садах в зоне коттеджной застройки, изредка в местных парках. Каждая птица снабжалась уникальной комбинацией цветных колец, чтобы её можно было идентифицировать в поле. Большинство мест поимки использовались в период с зимы 2016/17 по 2019/20 годы, за исключением места, обслуживаемого GCMR и сотрудниками с 1992 года. Авторы и добровольцы регулярно наблюдали за птицами, посещающими кормушки в придомовых садах на протяжении всей зимы. Им выдали информационные листы и просили записывать все наблюдения меченых черноголовок. Анализ учитывал разницу в усилиях и обнаруживаемости [птиц] между двумя способами контроля, подробно описанными ниже: (1) путем исключения мест, где черноголовок наблюдали менее 30 дней за зиму (2) путем указания случайных эффектов местоположения. Авторы полагали, что эти черноголовки уверенно классифицируются как зимующие (в отличие от самых ранних или поздних мигрантов, проходящих через территорию), если они отмечались с 1 декабря по 1 марта, определив эти границы по данным о повторных встречах меченых особей (рисунок S1). Сохраняли все записи об особях, отмеченных с 31 октября по 22 апреля.
Собранные наблюдения состояли из поимок (паутинной сетью или лучком), визуальных наблюдений за перемещениями и данных фотоловушек. Всего получено 9929 записей о 623 окольцованных славках, 357 из них встретились один или несколько раз после первой поимки.
2.2. Морфометрия
Были проведены следующие измерения: максимальная длина крыла (выпрямленного, по хорде, рис. 11.5 в Redfern & Clark, 2001); длина хвоста (рис. 11.12 в Redfern & Clark, 2001); минимальная длина цевки (рис. 11.11а в Redfern & Clark, 2001); расстояние между верхушками первостепенных и крайних второстепенных маховых (так называемое “расстояние Киппа”; ΔS1 у Локвуда и др., 1998); расстояние между верхушкой первостепенных махов и концом 9го (дистального) первостепенного (далее “расстояние P9”, ΔQ1 у Lockwood et al., 1998); длина клюва до черепа (рисунок 11.8 у Redfern & Clark, 2001); его ширина и глубина клюва в дистальной части ноздрей (рисунок 11.10 в Redfern & Clark, 2001). Птиц также взвесили и визуально определили их уровень жирности (шкала ESF) и развитости мышц (таблицы 11.13 и 11.14 в Redfern & Clark, 2001). Птицы, как правило, накапливают запасы жира в депо на протяжении дня, авторы корректировали оценки по данным регрессии, ставящей жирность в зависимости от времени суток (использовали остатки модели). Сходно выведен индекс общего состояния тела как регрессия общей массы к длине предплюсны (показатель линейных размеров птицы), так и ко времени суток; в обоих случаях также использовали остатки модели. Время везде устанавливается непрерывным, чтобы учесть возможную нелинейную зависимость от состояния тела.
Для анализа мы использовали полевые измерения, сделанные авторами BMVD, RJP, GCE и GCMR. Исключались первые 15 птиц, измеренных каждым из них. В качестве меры предосторожности, чтобы исключить любую предвзятость, созданную индивидуальностью измерителя, данные центрировали так, чтобы между ними не было средних различий.
Если одну и ту же птицу ловили несколько раз за сезон (23% случаев), сделанные каждый раз измерения усреднялись. Для обобщения различий по морфометрии выполнен анализ главных компонент для выявленного разнообразия измерений крыла, хвоста, предплюсны и клюва, и дальше использовали первые 3 компонента (таблица S2). Первый из них дал положительные нагрузки по всем переменным, особенно по крыловым и хвостовым. Его значения сильно различны у птиц разного возраста: у сеголеток в первую зиму жизни они значимо более отрицательные, видимо, в связи с известными изменениями длины и формы крыла в онтогенезе (рисунок S2; Pérez-Tris & Tellería, 2001).
Второй компонент дает положительные нагрузки для измерений ширины и глубины клюва и отрицательные — для формы крыла; эта составляющая изменчивости форм крыла и клюва сильно варьирует в зависимости от пола. У самки обоих возрастных классов значения по этой оси выше, что соответствует более толстым клювам и несколько меньшим крыльям (рисунок S2; Rolshausen et al., 2009). Третий компонент влиял в основном на изменчивость расстояний от клюва до черепа и P9 (нагрузки положительны), что фиксирует дополнительные различия в форме клюва и крыльев, не связанные с полом и возрастом.
2.3. Ежедневная погода
Данные о погоде получены от местных метеостанций из интегрированной системы архивирования данных Met Office (MIDAS; Met Office, 2012;
http://catalogue.ceda.ac.uk/uuid/220a65615218d5c9cc9e4785a3234bd0). Станции MIDAS выбирали как можно ближе к местам наблюдений славок (от 9 до 30 км). Авторы вычисляли (1) минимум и максимум суточных температур (за 24 часа, начиная с 9 утра), (2) общее количество осадков за 12 часов, считая с 6 утра до 18 вечера и (3) среднюю скорость и направление ветра за периоды (2). Дневной температурой считали среднюю минимального и максимального значений.
2.4. Учётные работы
Для понимания факторов окружающей среды, влияющих на посещаемость черноголовками кормушек, ежедневно подсчитывали число посетивших их меченых особей. Включили лишь птиц, отнесенных к категории зимующих в Великобритании (см. выше). Данные с каждой территории сохраняли для анализа, если черноголовок наблюдали по крайней мере 30 дней в течение этой зимы. Так набрались данные по 2615 дням наблюдений на восьми участках в течение 21 года (таблица S1). Большинство наблюдений выполнено GCMR (63,5%) и GCE (16,3%).
Зависимость посещений кормушек мечеными славками моделировали регрессионным анализом с помощью обобщенной линейной модели смешанных эффектов, в зависимости от суточной температуры воздуха, осадков, скорости ветра и дня года (с 1 ноября). Поскольку мы ожидали, что влияние дат наблюдений может быть нелинейным, их указывали дату в качестве сглаженного предиктора, модель применяли, используя функцию gam в пакете R mgcv (Wood, 2017) с семейством распределений Пуассона. Рабочая гипотеза предполагала большую вероятность посещения кормушек в холодные и влажные дни с сильным ветром из-за роста потребности в энергии и трудностей с поиском и добычей природных кормов. Авторы уточнили взаимосвязь изменений температуры × осадков, температуры × скорости ветра и осадков × скорости ветра, предположив, что воздействия осадков и ветра утяжеляются при низких температурах, а ветер влияет на эффект осадков. Случайными эффектами регрессии выбраны год и местоположение, чтобы учесть различия в среднем количестве славок, отмеченных в разные годы и на разных участках. Переменные-предикторы стандартизированны, чтобы можно было напрямую сравнить размеры эффекта между предикторами и интерпретировать свободный член регрессии по средним значениям предикторов. Модель упростили, удалив несущественные взаимодействия (р > 0,05).
Использованный набор данных включал ежедневные встречи, собранные десятками наблюдателей, но не хватало данных о распределениях усилий наблюдателей, чтобы понять, что значат дни без встреч славок — птиц не было или просто не наблюдали? Следовательно, в наборе данных отсутствуют нулевые значения, а они ожидаются при использовании семейства распределений Пуассона. Чтобы убедиться, что отсутствие нулей значений не сильно повлияло на использование пуассоновского распределения, модель совместили модель с пакетом R glmmTMB (Brooks et al., 2017), который поддерживает усеченное семейство распределений Пуассона без нулевых отсчетов. Этот пакет не поддерживает сглаженные предикторы, поэтому вместо этого используется квадрат даты. Авторы сравнили его с приведенной выше моделью gam, чтобы убедиться, что выбор семейства дистрибутивов не повлиял на выводы.
2.5. Посещение кормушек
Наблюдения в садах показали значительные различия в поведении славок. Имеющийся набор наблюдений использовали для моделирования вероятности посещения кормушек разными черноголовками на протяжении зимы. Анализ ограничили 8 пунктами, описанными в предыдущем разделе про учеты, с наиболее непрерывными наблюдениями.
В моделях использовали бинарную переменную отклика: в связи с действием тех или иных факторов наблюдали данную особь в данный день или нет? Анализ ограничен периодом, в течение которого индивид наблюдался на участке конкретной зимой. Исключены птицы, встретившиеся только один раз за зиму, или с неизвестным возрастом. Использованы наблюдения за 225 особями (128 самцами и 97 самками), отмеченными в среднем 13 раз (диапазон: 2-225). Весь набор данных включал 6104 положительных наблюдения и 10 982 предполагаемых отрицательных наблюдения.
Авторы использовали обобщенную модель смешанных эффектов с биномиальным распределением и функцией логической связи для моделирования вероятности обнаружить ставок в садах в определенные дни. Модель подгоняли с помощью bam (версия gam для больших наборов данных). Он включал все фиксированные эффекты, указанные в модели “ежедневных подсчетов”, с добавлением пола и возраста. Он включал в себя сглаженный срок действия; также включена непрерывная переменная количества дней, прошедших с момента последнего физического отлова птицы, на случай, если отлов может впоследствии снизить вероятность наблюдения. Авторы упростили модель, удалив несущественные взаимодействия (р > 0,05); определили случайные эффекты года, местоположения и индивидуума, чтобы учесть различия в средней вероятности обнаружения черноголовок по годам, участкам и индивидуумам.
Затем авторы сосредоточились на проверке взаимосвязи между посещением сада и морфологией/ физическим состоянием птиц, ограничив анализ данными по славкам, отловленным и измеренным хотя бы раз за зиму. Это уменьшило выборку данных с 6104 положительных наблюдений за 225 особями до 2777 положительных наблюдений за 90 особями. Для них приведенная выше модель была расширена: добавлены жировые запасы, состояние тела и морфометрические основные компоненты 1-3 в качестве дополнительных фиксированных эффектов. Кроме того, из-за морфологических изменений по ходу онтогенеза (например, Pérez-Tris & Tellería, 2001), могущих изменить оцениваемые причинные влияния, авторы проверили взаимосвязь между фиксированными эффектами и возрастом. Как и выше, затем они упрощали модель, удалив несущественные взаимодействия (р > 0,05).
Авторы рассчитали долю изменчивости посещений [кормушек], объясняемую случайными эффектами местоположения, года и идентичности птиц. Использовали пакет rptR (Stoffel et al., 2017) для количественной оценки этих пропорций дисперсии по исходной шкале переменных и бутстреп (N = 100) для генерации 95% доверительных интервалов данных оценок. Сюда включены описанные выше фиксированные эффекты, за исключением морфологии, жировых запасов и физического состояния птиц. Таким образом, сначала рассчитывали один фиксированный эффект возраста (птицы неизвестного возраста исключены) (N = 350). Во второй модели добавили пол, дату, когда птица была впервые встречена той зимой, и взаимосвязь между датой и возрастом. Также [в регрессию] добавлены случайные свободные члены года, местоположения и идентификатора птицы, чтобы учесть различия в средней вероятности перемещений между годами, местоположениями или особями, встречавшимися несколько лет.
2.8. Перемещения
Исследовали перемещения черноголовок на протяжении зимы, между зимами и между зимовкой и гнездованием, используя данные повторных находок окольцованных птиц и индивидуальное отслеживание. Данные кольцевания получены из системы кольцевания Британского орнитологического фонда (BTO) для Великобритании и Ирландии. Для переосмысления данных как зимующих на Британских островах классифицировали славок, отмеченных с 1 декабря по 1 марта (рисунок S1).
Учитывая данные о перемещающихся особях среди черноголовок, зимующих в садах, для их изучения использовали данные кольцевания. Авторы отсортировали данные о кольцевании по особям, которые, они уверены, зимовали в Великобритании и Ирландии (см. выше), однако рассматривали все их повторные поимки между 1 ноября и 1 апреля, чтобы понять перемещения птиц в самом начале или в конце зимовочного сезона. Они также использовали данные о кольцевания для отслеживания перемещений между зимами, отфильтровав данные по встречам, между 1 декабря и 1 марта.
Авторы объединили данные кольцевания с данными по птицам с геолокаторами, отслеживающими уровень освещенности (рис.1с), чтобы определить, где проводят лето черноголовки, зимующие в Великобритании и Ирландии. Для этого отфильтровали данные по летним находкам (между 15 мая и 15 августа) птиц, окольцованных зимой в Великобритании и Ирландии. Данные геолокаторов получены из Delmore et al. (2020). Вдобавок к определению мест размножения славок, авторы сравнили сроки миграции, установленные по геолокаторам, с наблюдением зимующих птиц в садах. В том числе исследовали, как скоро после весеннего исчезновения из садов черноголовки покидают Британию. Короткий интервал предполагает важность тамошних кормушек для набора жира перед миграцией.
3. Результаты
Динамика численности
Погодные условия и месяцы наблюдений сильно влияли на число славок в садах (таблица S3). Их больше отмечали при более низких температурах, большем количестве осадков и более сильных ветрах, хотя сочетание сильных ветров и осадков, возможно, препятствовало кормлению. Численность птиц в садах росла на протяжении всей зимы и снизилась с середины марта (рис. 2а). Значимость и направленность этих эффектов были согласованы в усеченной модели Пуассона, которая включала квадратичный, а не сглаженный термин даты (не показан).
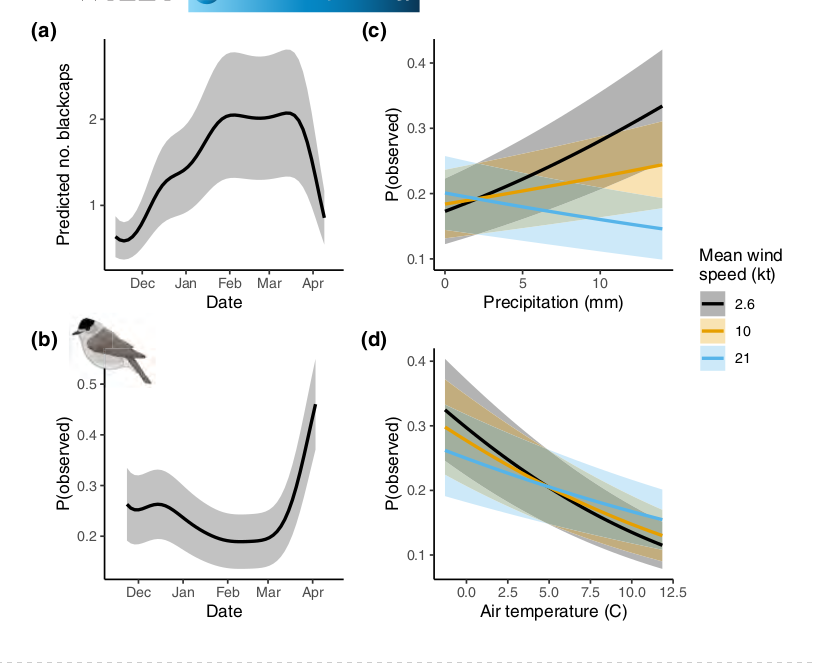 Рисунок 2. Встречаемость черноголовок в садах Великобритании и Ирландии в зависимости от времени года и погоды. Даны результаты, моделирования, предсказывающие число наблюдаемых птиц или вероятности встречи с ними в садах. (а) Прогноз динамики числа встреч славок с цветными кольцами в садах на протяжении зимы. (b–d) прогноз вероятности встречи конкретной птицы в саду между первой и последней зимними встречами, т.е. устойчивость посещений. (а) динамика численности славок в британских садах дана с учетом реакции на погоду (численность выше всего в конце зимы). (b) Динамика вероятности посещения сада славками на протяжении зимы. Общее число особей к концу зимы падает, но растёт посещаемость садов у оставшихся. (c) Зависимость посещаемости сада славками от осадков (ось x) при разной интенсивности ветра (показано цветом). (d) То же в зависимости от температуры (ось x).
Рисунок 2. Встречаемость черноголовок в садах Великобритании и Ирландии в зависимости от времени года и погоды. Даны результаты, моделирования, предсказывающие число наблюдаемых птиц или вероятности встречи с ними в садах. (а) Прогноз динамики числа встреч славок с цветными кольцами в садах на протяжении зимы. (b–d) прогноз вероятности встречи конкретной птицы в саду между первой и последней зимними встречами, т.е. устойчивость посещений. (а) динамика численности славок в британских садах дана с учетом реакции на погоду (численность выше всего в конце зимы). (b) Динамика вероятности посещения сада славками на протяжении зимы. Общее число особей к концу зимы падает, но растёт посещаемость садов у оставшихся. (c) Зависимость посещаемости сада славками от осадков (ось x) при разной интенсивности ветра (показано цветом). (d) То же в зависимости от температуры (ось x).
3.2. Индивидуальная посещаемость [садов]
В поведении зимующих славок значительны различия между индивидами. Из 140 птиц, наблюдавшихся всё вместе не менее 50 дней, 31 встречены менее чем в 15% из них, но 35% — не менее чем в 50%. Неудивительно, что номер особи объяснял большую долю дисперсии данных о посещаемости (0,13, 95% ДИ [0,08, 0,16]), чем местоположение [сада] (0,05 [0, 0,12]) или год наблюдений (0,04 [0,02, 0,07]).
Посещение [кормушек] связано с полом, физическим состоянием и морфологией [особей], как и с погодой. Первая модель авторов [для посещаемости] включала всех птиц, но не данные об их физическом состоянии и морфологии: она выявила сильную взаимосвязь с температурой воздуха, осадками и ветрами. Славки с большей вероятностью посещали кормушки в более холодную и влажную погоду; взаимодействия показали, что эти зависимости меняются при разной силе ветра (таблица S4; рисунок 2). Не найдено значимых различий ни между классами возраста (в среднем), ни влияний поимок на последующую посещаемость, самки отмечены несколько реже самцов.
Другая модель, включающая морфологию, физическое состояние птиц и возраст как дополнительный фактор воздействия, показывает сильную связь посещаемости с PC3, описывающей формы крыла и клюва. Взрослые славки с более длинными клювами и более округлым крылом встречались на кормушках значительно чаще (таблица S5; рисунок 3a). Однако у сеголеток эти зависимости не выявлены. Жировые запасы и физическое состояние тесно связаны с пребыванием на кормушках: часто посещающие сады взрослые имели меньшие жировые запасы и лучшее физическое состояние, чем бывающие там нечасто (рис.3bc). У сеголеток зависимость противоположна: часто посещающие сады имели большие запасы жира и худшее физическое состояние. Они же были чувствительней к температуре, чем взрослые.
3.3. Верность месту зимовки
Общая вероятность зимовки черноголовок в том же саду в последующие годы составила 0,4 (95% ДИ [0,16, 0,7]) для взрослых и 0,24 (95% ДИ [0,086, 0,5]) для молодых. Самцы и самки не различались по привязанности к месту зимовки, но у сеголеток она снижена вдвое. Возвращаемость была также существенно выше у птиц, впервые отмеченных в более поздние сроки и дольше находившихся у кормушек (таблица S6). Это может быть связано с перемещениями в начале зимы (см.ниже), а также с тенденцией возвращаться туда, где птицы были в конце предыдущей зимы.
Особи демонстрировали низкую повторяемость дат, когда их впервые и в последний раз наблюдали в садах каждый год. [Межгодовая] повторяемость дат первого наблюдения на кормушках составила 0,1 (95% ДИ [0, 0,35]), последнего — 0,17 (95% ДИ [0,03, 0,32]).
3.4. Резидентность [и, наоборот, перемещения] славок
Общая вероятность резидентности [встреченных особей] (т.е. наблюдения дальше более недели подряд) особей, впервые окольцованных на зимовке, составила 0,5 (95% ДИ [0,3, 0,7]). Вновь встреченные взрослые и вернувшиеся взрослые особи значимо более резидентны, чем сеголетки (новые взрослые особи: 0,67 (95% ДИ [0,44, 0,84]); вернувшиеся взрослые особи: 0,75 (95% ДИ [0,52, 0,89])). У разных возрастных групп отмечен различный паттерн резидентности в зависимости от дат появления в садах (таблица S7): вернувшиеся на зимовку взрослые, впервые отмеченные в начале зимы были высоко резидентны, но впервые отмеченные в более поздние сроки, держались очень недолго (рисунок S3). Вновь обнаруженные взрослые особи и сеголетки значительно различались по резидентности, с меньшим изменением вероятности пребывания в течение сезона.
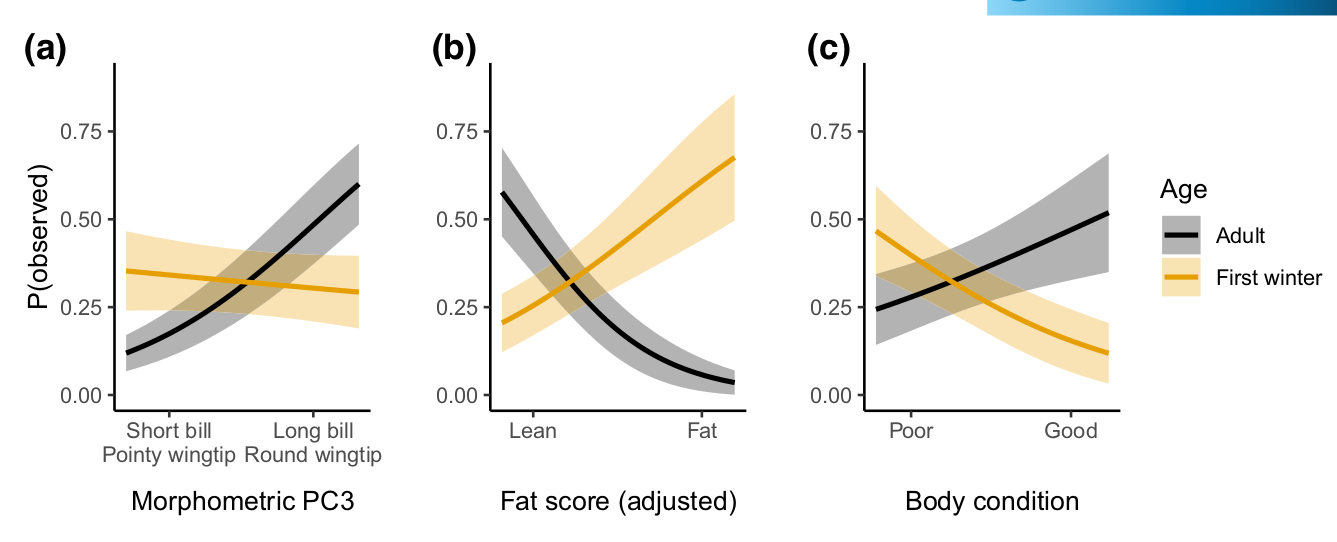 Рисунок 3. Посещаемость садов черноголовками (вероятность встречи, ордината) в зависимости от морфологии, жирности и физического состояния. Чёрные линии, серые полосы — значения и ДИ для взрослых, бежевые — для молодых славок (а) Зависимость посещаемости от PC3 (ось x). У взрослых, чаще посещающих сады, клювы, скорее всего, были длиннее, а крылья менее заостренные и более округлые, у молодых зависимости нет или она обратная. (b) То же в связи с жировыми запасами (ось x). Чаще посещающие сады взрослые, скорее всего, были менее жирными, но сеголетки наоборот. (c) То же в связи с физическим состояния птиц (ось x) при учёте жирности. У часто посещающие сады взрослых оно было наилучшим, у сеголеток — наихудшим.
Рисунок 3. Посещаемость садов черноголовками (вероятность встречи, ордината) в зависимости от морфологии, жирности и физического состояния. Чёрные линии, серые полосы — значения и ДИ для взрослых, бежевые — для молодых славок (а) Зависимость посещаемости от PC3 (ось x). У взрослых, чаще посещающих сады, клювы, скорее всего, были длиннее, а крылья менее заостренные и более округлые, у молодых зависимости нет или она обратная. (b) То же в связи с жировыми запасами (ось x). Чаще посещающие сады взрослые, скорее всего, были менее жирными, но сеголетки наоборот. (c) То же в связи с физическим состояния птиц (ось x) при учёте жирности. У часто посещающие сады взрослых оно было наилучшим, у сеголеток — наихудшим.
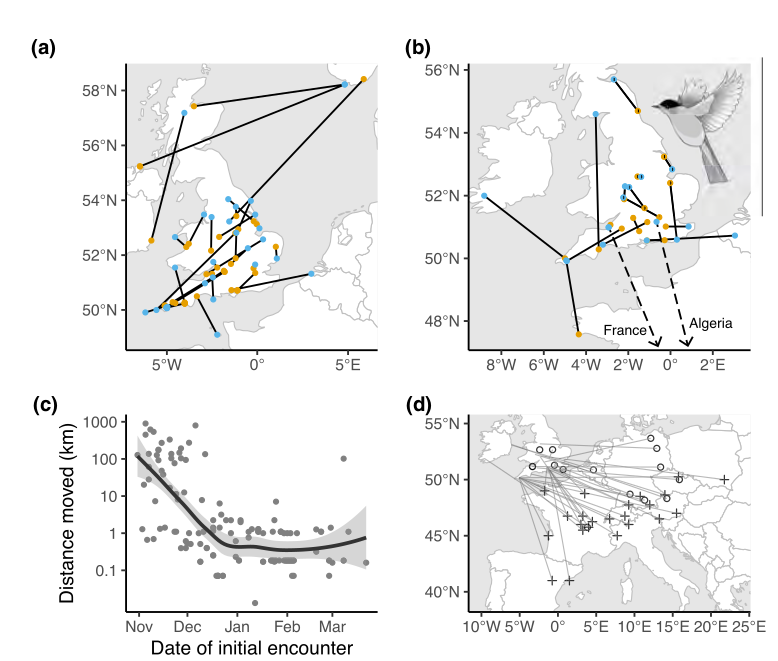 Рисунок 4. Перемещения зимующих черноголовок по данным кольцевания: синие точки — места первоначального кольцевания, жёлтые — повторных поимок окольцованных. (a) Перемещения на протяжении зимы. (b) Перемещения между зимами. Пунктир — движение птиц к местам находок в Алжире и центральной Франции. (c) Дальность перемещений на протяжении зимы в зависимости от дат кольцевания. (d) Мечта размножения (крестики и кружки) и зимовки (конечные точки линий) у славок, зимующих в Великобритании и Ирландии. Кружки — данные повторных поимок окольцованных, крестики — данные геолокаторов от Delmore et al.
Рисунок 4. Перемещения зимующих черноголовок по данным кольцевания: синие точки — места первоначального кольцевания, жёлтые — повторных поимок окольцованных. (a) Перемещения на протяжении зимы. (b) Перемещения между зимами. Пунктир — движение птиц к местам находок в Алжире и центральной Франции. (c) Дальность перемещений на протяжении зимы в зависимости от дат кольцевания. (d) Мечта размножения (крестики и кружки) и зимовки (конечные точки линий) у славок, зимующих в Великобритании и Ирландии. Кружки — данные повторных поимок окольцованных, крестики — данные геолокаторов от Delmore et al.
3.5. Перемещения
Славки, зимующие в Великобритании и Ирландии, перемещаются на протяжении зимы, но более чем на 10 км — почти исключительно в ноябре-декабре (рис. 4). Выше была показана привязанность многих славок к конкретному месту зимовки. И наоборот: особи, не демонстрирующие подобной верности, возможно, перемещаются на значительные расстояния между зимами. Дальность межзимних перемещений по данным кольцевания в среднем 148 км ± 309 SD (рис.4b). Подробные наблюдения показывают, что даже особи, проведшие в конкретном саду всю зиму, следующую могут провести вдали от прежнего места зимовки. Одна птица [N676642] отмечалась GCE в 63% дней на отрезке с 16 декабря 2017 года по 4 апреля 2018 года. Следующей зимой ее там уже не было, но ее встречали (и фотографировали) в саду в 53 км от этого места с 7 января по 12 февраля 2019 года. Ноябрьские перемещения составили в среднем 130 км ± 198 SD, декабрьские — 37,6 км ± 146 SD, январско-февральские — всего 0,735 км ± 1,04 SD.
3.6. Места гнездования и миграция
Суммируя данные кольцевания и геолокации, видим, что славки, зимующие в Великобритании и Ирландии, гнездятся обширной полосой по всей Европе, до 2000 км (рис. 4d). Данные 25 геолокаторов (Delmore et al., 2020) заставляют считать, что основной источник птиц для английских зимовок находится в Западной Европе (например, во Франции), однако почти все повторные находки окольцованных на континенте (N = 9) — с самого востока Европы. Еще шесть находок окольцованных случились в самих Великобритании и Ирландии: видимо часть ранее зимовавших птиц там загнездилась и стала обитать круглогодично (хотя см. раздел 4).
Из данных геолокаторов следует, что отлёт к местам гнездования у зимующих в Британии птиц происходит с 15 марта по 26 апреля, с медианой — 2 апреля (рис.5а). Осенью они прилетают с 4 сентября по 28 октября, с медианой 13 октября (рис.5b). Авторы сконцентрировались на птицах с геолокациоторами, в трех районах с хорошим охватом наблюдателями, чтобы определить, есть ли разрыв во времени между исчезновением славок с кормушек в садах и весенним отлётом из Великобритании. 3 птицы покинули сады вскоре после установки геолокатора, у оставшиеся 9 отлёт вообще из Англии произошëл в среднем через 4,22 ± 4,49 SD дней после последнего наблюдения на кормушке (рис.5с). Из-за небольшой дальности пролётных путей эти черноголовки прибудут к местам гнездования в среднем через пару дней после вылета, т. е. они появились в гнездовом ареале только через 5,44 ± 5,48 SD дней после последних раз наблюдений в садах на зимовках. Столь тесное соответствие между уходом с кормушек и началом миграции резко контрастирует с ситуацией осенью. Тогда славки появляются в Великобритании в среднем за 47,8 ± 18,5 SD дней до первых наблюдений в садах (N = 12; рисунок 5d).
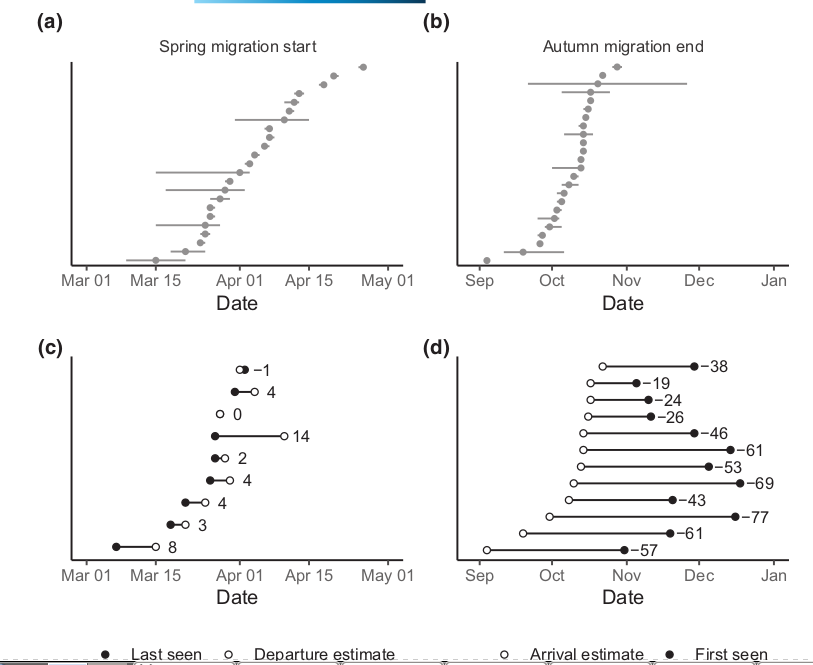 Рисунок 5. Сроки миграции черноголовок, зимующих в Великобритании. (a) и (b) распределения дат отлета в весенней и прибытия в осенней миграции; линии — неопределенность в виде межквартильного диапазона модельных оценок (Delmore et al., 2020). «Местонахождения» одних и тех же особей на горизонтали весной и осенью не совпадают. (c) и (d) сравнение данных геолокатора (полые кружки),показывающих последнее наблюдение птиц перед отлётом весной иди первое — после прилёта осенью, с датами последнего или последнего наблюдения птиц на зимовках в садах (заполненные круги). Цифры — разница между ними в днях. Положение одних и тех же особей на разных горизонталях не совпадает.
Рисунок 5. Сроки миграции черноголовок, зимующих в Великобритании. (a) и (b) распределения дат отлета в весенней и прибытия в осенней миграции; линии — неопределенность в виде межквартильного диапазона модельных оценок (Delmore et al., 2020). «Местонахождения» одних и тех же особей на горизонтали весной и осенью не совпадают. (c) и (d) сравнение данных геолокатора (полые кружки),показывающих последнее наблюдение птиц перед отлётом весной иди первое — после прилёта осенью, с датами последнего или последнего наблюдения птиц на зимовках в садах (заполненные круги). Цифры — разница между ними в днях. Положение одних и тех же особей на разных горизонталях не совпадает.
4.Обсуждение
Организмы все чаще сталкиваются с необходимостью адаптации к миру, в котором доминирует человек. Перемещения животных во время миграций могут повысить их способность к исследованию и колонизации новых подходящих районов, однако миграционный образ жизни также зависит от сложного годового цикла [связанного с последовательным посещением вовремя разных мест], где трудности, возникшие на одном этапе [или проблемы с местообитаниями на одном из этапов] могут иметь долговременные последствия [проявляющиеся на других этапах цикла]. Авторы выявили, что экология славок в недавно возникшем достаточно северном районе зимовки формируется деятельностью человека, включая зимнюю подкормку птиц, и опосредуется погодными и другими условиями зимовки.
4.1. Влияние человека на поведение черноголовок
Сады с кормушками могут быть особенно значимы при ухудшении погоды зимой, что подтверждается данными. В более холодную и сырую погоду посещение кормушек растёт, ибо в такой ситуации поедание ягод и фруктов (Snow & Snow, 2010) слишком уж расточает энергию. Кормушки дают устойчивый доступ к высококалорийной и/или жирной пище, особенно важный при повышенных требованиях к терморегуляции (когда холодно, сыро и/иди ветрено). В садах славки больше питаются жиром, фруктами и семенами, включая ядра подсолнечника (Leach, 1981; Plummer et al., 2015). Кроме того, сады — существенно изменённые местообитания с важным источником пищи для славок в виде плодов и семян неместных растений (например, Beatty, 1994; Hardy,1978; Harrup, 1998; Leach, 1981; Snow & Snow, 2010; Walton, 1983).
Хотя в ряде садов люди могут чаще кормить славок при ухудшении погоды, это вряд ли влияло на результаты исследования. По-настоящему суровые погодные явления (со снежным покровом или длительными заморозками) в использованной выборке данных редки. Кроме того, 79,8% проанализированных данных получены из садов авторов GCMR и GCE, постоянно кормивших птиц. Наконец, рост потребности в дополнительной пище при более суровых условиях обычно усиливает конкуренцию между разными особями в садах и быстро истощает эти ресурсы.
Появление постоянных и надежных источников пищи нетипично для вида, в норме зимой плодоядного [а это ресурс непостоянный и эфемерный быстро исчерпывающийся]. В традиционных средиземноморских местах зимовок черноголовки зависят от эфемерных ресурсов ягод и часто переселяются в связи с их истощением (Belda et al., 2007; Cuadrado, 1992; Rey, 1995; Tellería et al., 2008). Верность черноголовок месту зимовки в разные зимы составляет всего лишь 0-5% в Африке и Иберии (Cuadrado, 1992; Cuadrado et al., 1995; King & Hutchinson, 2001; Lövei et al., 1985). Данные по поискам–повторным отловам в Испании показали, что лишь 26% славок вероятно, будут привязаны к данному участку на протяжении зимы, считая привязанностью пребывание там более 1 недели (Belda et al., 2007). На английских зимовках существенно выше как привязанность птиц между зимами, так и резидентность на протяжении зимы: первая 0,4 (95% ДИ [0,16, 0,7]) для взрослых и 0,24 (95% ДИ [0,086, 0,5]) для сеголеток, вторая 0,5 (95% ДИ [0,3, 0,7]) у молодых, 0,67 (95% ДИ [0,44, 0,84]) у вновь встреченных взрослых славок.
Возможно, английские данные напрямую не сопоставимы со средиземноморскими, ибо получены разными способами: первые визуальными наблюдениями, вторые отловом. Будущие исследования поведения славок на зимовках только выиграют от большей стандартизации способов наблюдений и индивидуального отслеживания птиц (скажем, радиотелеметрией). Тем не менее весьма вероятно, что черноголовки, зимующие в Великобритании и Ирландии, отличаются значимо большей привязанностью к месту зимовки в разные зимы и большей резидентностью на протяжении одной зимы из-за большего обилия и надежности дополнительных источников корма в садах сравнительно с обычными ягодами. Хотя многие важные естественные корма славок широко распространены в пригородных районах, большинство их недоступны в середине зимы, когда славок в садах больше всего (например, ягоды плюща, Hedera helix, Snow & Snow, 2010).
Зимовка в Великобритании и Ирландии даëт ряд преимуществ по сравнению с традиционной миграцией в юго-западном направлении (Berthold & Terrill, 1988). Так, высокая надежность зимних кормов повысит выживаемость, выступив буфером против суровых погодных условий. Взрослые, часто посещающие сады, при меньшей жирности имели лучшее физическое состояние, чем посещавшие сады нечасто. У молодых же наоборот, часто посещающие сады были в худшем физическом состоянии но более жирными. Этот контраст может отражать знакомство взрослых с надежными местными кормами и их доминирование над молодыми. Меньшее потребление жира также снижает риск хищничества (Fransson & Weber, 1997; Kullberg et al., 1996). Данные авторов показали, что сады интенсивней всего используются как успешными взрослыми, наиболее приверженными резидентной стратегии, так и потенциально проблемными сеголетками, запасающими жир как буфер на случай проблемной погодной и кормовой ситуаций.
Обильное дополнительное питание позволяет черноголовкам перед миграцией улучшить физическое состояние и быстро накопить жир, поддерживающий их перелет. В свою очередь, подкормка может толкать к более раннему и при этом успешному размножению. Славки, отслеживаемые по геолокаторам, прибыли на места гнездования в среднем всего через 5,4 дня после последних наблюдений в садах, что заставляя считать, что тамошние условия повлияют и на состояние птиц по прибытии. Отсюда вполне вероятно, что надежный дополнительный корм на островных зимовках дает преимущество в местах размножения.
Взрослые особи с более длинными клювами и более закруглёнными крыльями сады посещали чаще. Эти различия образуют параллель с различием по морфологии клюва между ставками, зимующих в Великобритании и на Пиренеях; Rolshausen et al. (2009) предположили, что более узкие и длинные клювы британских птиц отражают их более универсальный рацион по сравнению со специализированной плодоядностью в Средиземноморье (при большей твëрдости этих плодов, вроде оливок Olea europaea) (Jordano & Herrera, 1981; Tellería et al., 2008).
Более длинные и тонкие клювы в связи с подкормкой сходно развились в Великобритании у больших синиц (Parus major) (Bosse et al., 2017). Данные авторов согласуются с этими выводами и дают дополнительные доказательства что зимняя подкормка дополнительно движет морфологическую эволюцию у птиц. Жёсткость подобной зависимости у взрослых, но не молодых, впервые зимующих, может отражать поведенческие различия между наивными и опытными особями. Другое возможное объяснение — различия давлений отбора связанные с возрастом. У молодых отбор на большую закругленность крыла, т. е. лучшую маневренность (Pérez-Tris & Tellería, 2001) может превосходить все другие селективные требования, особенно в критический период после вылета.
4.2. Потенциальные будущие изменения в поведении
Уже получены явные доказательства того, что человеческая деятельность повлияла на поведение черноголовок, зимующих в Великобритании и Ирландии, прямо и косвенно. Хотя они чаще зимуют в более теплых районах острова, но посещают сады у домов в основном в холодную и сырую погоду (Plummer et al., 2015). В ближайшие десятилетия в Великобритании ожидается потепление климата (Lowe et al., 2018). Это может повысить пригодность острова для зимовки славок, но также повлиять на их связь с подкормкой в садах. Возможно сокращение их посещаемости в будущем при более мягкой погоде, вопреки росту численности зимующей популяции.
4.3. Велика ли зависимость славок от дополнительного питания?
Возможно, дополнительная подкормка и важна птицам в тяжёлых зимних условиях, но анализ показывает, что славки далеки от того, чтобы полагаться [преимущественно] на кормушки. Высоко подвижные особи, редко бывающие в садах, составляли примерно половину всех пойманных славок. Столь высокая подвижность согласуется с возможностью большинству птиц существовать на зимовках в Великобритании и Ирландии, когда кормушки дают лишь небольшую часть рациона. Неслучайно даже самые привязанные к месту зимовки черноголовки появляются в садах много позже собственно прилёта на зимовку — иногда более чем через 2 месяца. Конечно, нельзя исключить и возможность того, что подобные особи просто переселились из одних садов в другие; в будущем прослеживание птиц с передатчиками улучшило бы понимание [пространственных связей зимующих черноголовок, с учётом возможного присутствия в популяции нескольких классов особей, различающихся в этом отношении] и/или характера использования ими разных кормов, природных и антропогенных, диффузно рассеянных и концентрированных, привязанных к садам. Однако самое правдоподобное (и простое) объяснение — кормушки для славок просто без надобности, пока не иссякнут природные источники корма (что зависит от фенологии данной осени и ранней зимы). Составляя небольшую часть рациона, подкормка может быть критической адаптацией, обеспечивая выживание в более суровую погоду. Отсюда кормушки в садах могут непропорционально сильно (и положительно) влиять на выживаемость.
4.4. Индивидуальные различия и гибкость
Высокие индивидуальные различия — определяющая черта полученных данных. При моделировании посещаемости садов наибольшая доля объясненной дисперсии пришлась на индивидуальность особей. Посещаемость изменялась в диапазоне от частой, с высоким постоянством пребывания до временного, а то и с одним посещением.
Молодые, скорее всего, были подвижной, в саду пребывали недолго, особенно в начале зимы. Возможно эти птицы неопытны и поэтому более склонны исследовать разные места и возможности, определившиеся [с жизненной стратегией] взрослые. Также взрослые могут доминировать над молодыми в садах, толкая их стать подвижней. Большая филопатрия взрослых возможно, отражает их большую выживаемость, но может говорить и о возрастных различиях в пространственных связях. Взрослые, обосновавшиеся в местообитаниях высокого качества, могут извлечь выгоду из [высоко вероятного] возвращения в данный район. И наоборот, сеголетки на своей первой зимовке могут сосредоточиться на поиске таких наилучших мест. Как бы то ни было, полученные результаты показывают, что черноголовки крайне вариативны и гибки в разных аспектах зимовочной биологии, что может быть ключом к успеху вида при быстрых антропогенных изменениях среды обитания.
4.5. Происхождение [птиц], зимующих в Великобритании и доказательства резидентности их на кормушках
Авторы объединили данные кольцевания и слежения за индивидуально маркированными особями, чтобы показать происхождение зимующих в Великобритании славок из широкой полосы континентальной Европы, около 2000 км (Delmore et al., 2020). Одни только повторные находки окольцованных славок подтверждали данные о центрально-европейском происхождении зимующих в Британии птиц, но данные геолокаторов показывают, что их места гнездования простираются далеко на юго-запад Европы, даже на Пиренейский полуостров, и уходят много восточнее, чем предполагалось по находкам колец. Отмечено 6 случаев круглогодичного нахождения птиц Великобритании и Ирландии: видимо, малая часть зимующих стала оседлой или остаётся на лето (Для масштаба: всего за несколько летних сезонов было встречено и более или менее было под наблюдением 5848 птиц). Однако вероятность повторной поиски окольцованных падает с расстоянием (пространственное искажение), что делает повторные поимки на той же территории гораздо более вероятными, чем вне её. Действительно, данные геолокаторов показали, что подавляющее большинство птиц прибыли зимовать с континента. Ни один из 25 треков геолокатора не показал признаков круглогодичного проживания в Великобритании, хотя птица, зимовавшая в Чилболтоне, Хэмпшир, Великобритания, переместилась для размножения очень недалеко, на северное побережье Франции.
5. Заключение
Жизнеспособность популяций многих перелетных птиц подорвана давлением изменений окружающей среды, они сокращаются, но славка-черноголовка процветает в этом меняющемся мире. Недавно возникшая зимующая популяция черноголовки в Великобритании и Ирландии дает возможность понять, как этому виду удается сохранять темпы роста. Полученные результаты подтверждают гипотезу о том, что изменение климата в сочетании с дополнительным кормлением птиц способствовало быстрому формированию этой зимующей популяции и продолжает формировать ее биологию. Более того, они говорят об огромной индивидуальной вариативности, которую проявляют славки-черноголовки во время зимовки в перемещениях и реакциях на изменения погоды.
Среди мигрирующих видов те, которые отличаются большой вариабельностью и гибкостью в передвижении и добывании пищи, вероятно, будут лучше всего подготовлены к реагированию на предстоящие десятилетия изменений окружающей среды.
Оригинал в Global Change Biology
Рекомендуем прочесть
 Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла»
Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла» Интеллектуальная акселерация (эффект Флинна): метаанализ
Интеллектуальная акселерация (эффект Флинна): метаанализ Восприятие шумов
Восприятие шумов- «Работа» релизеров в коммуникации животных: хороший стимул – плохой сигнал, и наоборот
 Пение самок воробьиных птиц: исключение или закономерность?
Пение самок воробьиных птиц: исключение или закономерность?- Реализация «генов образования» в зависимости от социального статуса в США
 Птицы документируют антропоцен: стратиграфия пластика из гнёзд лысухи
Птицы документируют антропоцен: стратиграфия пластика из гнёзд лысухи Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных
