![Print PDF Резюме. Фенотипическая пластичность позволяет одному генотипу производить множество фенотипов в ответ на изменения окружающей среды. Пластичность может играть решающую роль в колонизации новых сред, но ее роль в […]](http://www.socialcompas.com/wp-content/uploads/2023/02/Snimok-ekrana-ot-2023-02-12-13-11-04-465x190.png "Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла»")

Резюме. Фенотипическая пластичность позволяет одному генотипу производить множество фенотипов в ответ на изменения окружающей среды. Пластичность может играть решающую роль в колонизации новых сред, но ее роль в адаптивной эволюции противоречива. Предполагается, что быстрый и параллельный рост термостойкости гребенчатых анолисов в разных городах Пуэрто-Рико [ранее установленный авторами данной работы] вызван главным образом давлением отбора, снижающим и/или обращающим вспять ту фенотипичнскую пластичность, которая хоть и вызвана ростом температуры, но неадаптивна в городском «острове тепла».
Авторы приводят доказательства такого отбора по генам транскриптома скелетных мышц, связанным с толерантностью к жаре. «Лесные» ящерицы, выращенные в лаборатории при обычной температуре, в ответ на её рост изменяют экспрессию данных генов в сторону, коррелирующую со снижением термостойкости (т.е. их регуляторный ответ на температуру неадаптивен).
У «городских» ящериц картина противоположна. В обычных лабораторных условиях пластичность экспрессии генов в ответ на воздействие тепла у них снижена, а вот в городском «острове тепла» экспрессия генов сильно меняется в сторону, коррелирующую с большей термостойкостью (что адаптивно). Гены, в ответ на воздействие температуры проявляющие неадаптивную пластичность, неизменно сильней дивергируют при освоении ящерицами разных городов, чем гены, проявляющие адаптивную пластичность. Эти результаты подчеркивают значимость для эволюции отбора против пластичности, регулятивной по характеру, но неадаптивной.
[Введение]
Содержание
Понимание механизмов, которые порождают эволюционную новизну (новое фенотипическое разнообразие) — центральная цель эволюционной биологии. Здесь у нас есть пробел в понимании взаимодействия между изменчивостью фенотипа, вызванной воздействием изменений среды (фенотипическая пластичность, проявляющаяся как модификационная изменчивость, дающая длительные модификации в ответ на воздействие) и изменениями, детерминированными генетически (собственно эволюционными). В современной истории биологии пластичность рассматривалась в разных вариантах
а) как сдерживающая эволюцию путем буферизации генотипов от селективных давлений1-4,
б) как несущественная для эволюционных изменений источник ненаследуемых вариаций фенотипа5-7 и
в) как облегчающая колонизацию новых сред – в частности, на начальных стадиях адаптации к ним4.
До сих пор не очень ясны обстоятельства, при которых пластичность играет эти разные роли8,9, а ее роль в адаптивной дивергенции форм10-12 до сих пор дискуссионна.
Адаптивная ценность фенотипической пластичности зависит от наследуемости ответа организма на изменения среды его и направленности ее воздействия относительно некого оптимального фенотипа (рис.1). Адаптивная пластичность «приближает» популяцию к фенотипу, оптимальному для данной среды (обладающему здесь наибольшей приспособленностью), обеспечивая устойчивость популяции6 и позволяя отбору воздействовать на генетические вариации, лежащие в основе этого индуцируемого средой ответа (наследственная компонента пластичности).
Этот эффект приближает усредненный фенотип данной популяции Х к локальному пику адаптивности У4,13-15 (рис.1, координаты «фенотип — приспособленность» сверху и «изменения среды — изменения признаков» внизу).
Эволюция [за счёт] адаптивной пластичности продемонстрирована в широком диапазоне развитийных16, морфологических15, физиологических17 признаков, признаков жизненной стратегии13 и поведения18. Напротив, неадаптивная пластичность — однотипные изменения, однако неуместные в новой среде, снижающие приспособленность19–21. Отбор наследственной составляющей этой неадаптивной пластичности, может её свести к минимуму и/или полностью изменить ее направление4 (рис.1).
Например, физиологические реакции на низкий уровень кислорода исходно развились у видов равнин, чтобы справиться с острой гипоксией эндогенной природы (анемия, потеря крови или низкая оксигенация тканей)22. Однако в высокогорье недостаток кислорода — внешний фактор, создающий хроническое воздействие. Вот здесь эти же (унаследованные от предков) реакции вредны, ведут к лёгочной гипертензии и перепроизводству эритроцитов. Поэтому виды осваивающие высокогорья, эволюционировали к уменьшению или полная потере (канализация) этой наследственной пластичности23-27.
Акклиматизация и эволюционная адаптация к изменениям окружающей среды часто включают сложные скоординированные биологические реакции, такие как совместная регуляция генов, лежащих в основе физиологических функций. Усиление адаптивной пластичности и элиминация/обращение вспять неадаптивной пластичности на регуляторном и геномном уровнях, вероятно, идут сопряжëнно, подталкивая популяции к новым адаптивным пикам28. Однако относительная важность адаптивной и дезадаптивной пластичности как субстратов для отбора на начальных стадиях расхождения линий остается в значительной степени неисследованной.
Хотя выявление начальных механизмов, приводящих к адаптивной дивергенции, имеет решающее значение для понимания эволюции сложных систем в новых условиях, изучение таких явлений усложняется по мере того, как проходит время с момента расхождения линий, из-за накопления мутаций, сложных демографических историй [в разошедшихся популяциях] и последующих событий отбора. Примеры современной эволюции, такие как адаптация к городской среде, представляют собой “естественные эксперименты”, которые дают уникальное представление о механизмах, обеспечивающих эволюцию сложных признаков, меняющихся в ответ на изменение среды (или при освоении новой, вроде города). Такие примеры дают возможность количественного исследования элементарных эволюционных явлений в начале популяционной дифференциации и формирования локальных адаптаций в природе29,30.
Такое «распутывание» взаимосвязей между отбором, адаптацией и пластичностью всë более важно для понимания эволюционных последствий антропогенных изменений окружающей среды30. Быстрая трансформация местообитаний [видов] и изменения их среды обитания из-за прямого и косвенного влияния человека могут вызвать эволюцию физиологической, поведенческой и/или эволюционной пластичности31-34. Таким образом, наследственная пластичность может быть общей целью естественного отбора, если она приводит к большей устойчивости населения к изменению климата и быстрым антропогенным изменениям (HIREC)32,33,35.
Акклиматизация (экологический процесс приспособления) и адаптация (эволюционный процесс) к изменениям среды обитания часто включают сложные скоординированные биологические реакции, такие как совместная регуляция генов, лежащих в основе [нужных для этого] физиологических функций. Усиление адаптивной пластичности и снижение/обращение вспять неадаптивной на регуляторном и геномном уровнях, вероятно, происходят в тандеме, чтобы подтолкнуть [приспосабливающиеся] популяции к новым адаптивным пикам28. Однако относительная важность адаптивной и дезадаптивной пластичности как субстратов отбора на начальных стадиях расхождения линий [в связи с приспособлением, когда дочерняя линия, она же новообразующаяся популяция, осваивает новую среду] остается в значительной степени неисследованной.
Хотя выявление начальных механизмов адаптивной дивергенции критически важно для понимания эволюции сложных систем в новой среде, их исследование всё более усложняется с ростом времени с момента расхождения линий, из-за накопления мутаций, сложных демографических историй и последующих событий отбора. Случаи современной эволюции, такие как адаптация к городской среде, представляют собой “естественные эксперименты”, которые дают уникальное представление о механизмах, лежащих в основе эволюции сложных характеристик, реагирующих на окружающую среду. Их анализ позволяет количественно изучить эволюционную динамику на начальных стадиях расхождения линий и локальной адаптации в дикой природе29,30.
Кроме того, распутывание взаимодействий между отбором, адаптацией и пластичностью становится все более важным для понимания эволюционных последствий антропогенных изменений окружающей среды30. Быстрая модификация среды обитания и её трансформации из-за прямого и косвенного влияния человека могут вызвать эволюцию физиологической, поведенческой и/или развитийной пластичности31-34. Поэтому пластичность исходной популяции [авторы зовут её анцестральной] может быть «мишенью» естественного отбора, если её изменение повышает устойчивость к изменению климата и антропогенным трансформациям биотопов32,33,35.
Одним из таких антропогенных изменений среды обитания является рост температур в городах – следствие увеличения доли запечатанных поверхностей, обычно имеющих ниже альбедо, и выработки тепла, что в совокупности называется “эффект городского «острова тепла”36. В случае эктотермных видов рост городских температур сильно влияет на их жизнедеятельность37–42, воспроизводство, развитие43–45 и выживание46–49. Растет объем данных, что термоустойчивость – ключевой момент адаптации к урбосреде для наземных эктотермов37–39,41,42,50. Более того, унаследованная от предков пластичность ответа на тепловой стресс краткосрочно способствует выживанию особей, испытывающих периодические резкие скачки температуры, и может поначалу обеспечить устойчивость новообразованной популяции в урбосреде51. Например, поведенческая терморегуляция сохраняет особям оптимальную температуру тела за счёт использования температурной гетерогенности урболандшафта, чем сводит к минимуму влияние термической физиологии52 (эффект Богерта53).
Однако в экстремальной среде температура выходит за пределы предковой нормы реакции или неоднородность температур недостаточна для поведенческой терморегуляции. Эти новые и экстремальные для вида условия требуют физиологической адаптации [которая и вырабатывается отбором на термостойкость, давно и успешно исследуемой у «городских» ящериц данными авторами]. В таких случаях реализация пластичности в ответ на кратковременные температурные колебания может войти в конфликт [образовать трейд-офф] с другими моментами биологии, критически важными для выживания и размножения54,55, [и это негативно скажется на активности или жизнеспособности] включая снижение темпов роста, [ухудшение] физического состояния ящерицы, сокращение размера кладки и двигательной активности56-59. Колонизация городских «островов тепла» означает для ящериц, что тепловой стресс будет постоянным, повсеместным и экстремальным по силе, что усугубит неадаптивные последствия «ответной» физиологической пластичности.
Поэтому популяция, осваивающая урбосреду, столкнëтся с многосторонней головоломкой: как использовать индуцированную температурой пластичность для большей термостойкости, одновременно смягчив инадаптивные следствия данного изменения.
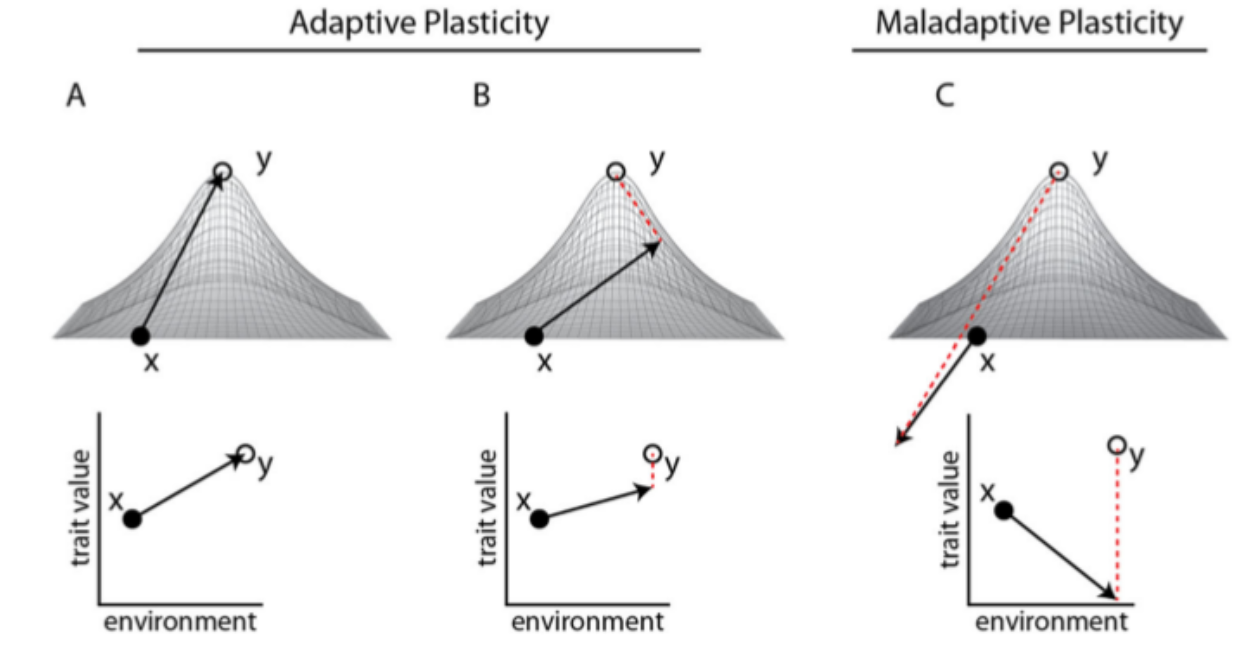 Рис.1. Эволюция пластичности в новых средах. Фенотипическая пластичность может повлиять на траекторию эволюционных изменений популяции, колонизирующей новую среду обитания. Реализация предковой пластичности может двинуть фенотип популяции (x) в разных направлениях относительно локального оптимума новой среды (адаптивный пик, y). Данные изменения адаптивны, когда они перемещают популяцию в признаковом непосредственно в У (A) или хотя бы в сторону этого пика, пусть за его пределами (В). Если здесь есть достаточный генетический компонент [как в примере с большими синицами] отбор усилит адаптивную пластичность и «переместит» усреднённый фенотип прямо к новому пику (B; пунктир). Однако отбор не «включается», если пластичность «выводит» особей прямо в У (А, опять же, как с синицами). В случае неадаптивной пластичности (С) реализованные изменения снижают приспособленность (отрицательные значения в адаптивной ландшафте). Тогда отбор сокращает эту изменчивость, сужая норму реакции или обращая её вспять. Так популяционная средняя «возвращается» сперва на исходный уровень приспособленности Х, а затем «движется» и в сторону пока У. Везде давление отбора тем сильней, чем дальше от У (пунктир в В и С). На основе рис.2 в Galambor et al. 4.
Рис.1. Эволюция пластичности в новых средах. Фенотипическая пластичность может повлиять на траекторию эволюционных изменений популяции, колонизирующей новую среду обитания. Реализация предковой пластичности может двинуть фенотип популяции (x) в разных направлениях относительно локального оптимума новой среды (адаптивный пик, y). Данные изменения адаптивны, когда они перемещают популяцию в признаковом непосредственно в У (A) или хотя бы в сторону этого пика, пусть за его пределами (В). Если здесь есть достаточный генетический компонент [как в примере с большими синицами] отбор усилит адаптивную пластичность и «переместит» усреднённый фенотип прямо к новому пику (B; пунктир). Однако отбор не «включается», если пластичность «выводит» особей прямо в У (А, опять же, как с синицами). В случае неадаптивной пластичности (С) реализованные изменения снижают приспособленность (отрицательные значения в адаптивной ландшафте). Тогда отбор сокращает эту изменчивость, сужая норму реакции или обращая её вспять. Так популяционная средняя «возвращается» сперва на исходный уровень приспособленности Х, а затем «движется» и в сторону пока У. Везде давление отбора тем сильней, чем дальше от У (пунктир в В и С). На основе рис.2 в Galambor et al. 4.
Пуэрториканский гребенчатый анолис (Anolis cristatellus) сделался отличной моделью для понимания взаимодействия между естественным отбором и пластичностью на ранних стадиях адаптации к городским островам тепла. На о. Пуэрто-Рико урбанизация территории началась с прибытия европейцев 500 лет назад, но A. cristatellus обитает в городских местообитаниях на протяжении не менее 35 поколений (Дополнительная таблица 1). Кэмпбелл-Стейтон и др.60 обнаружили, что «городские» ящерицы везде более термостойки сравнительно с ближайшей «лесной» популяцией, генетически наиболее близкой каждой из «городских» (видимо, первые произошли от вторых, т. е. разные города острова осваивались ящерицами независимо из окрестных лесов, рис.2А, Б). Однако при разведении «городских» и «лесных» ящериц в стандартных лабораторных условиях это физиологическое различие не выявляется (рис.2C), подчеркивая пластичность вызванного температурой ответа на среду обитания как главную причину данных физиологических различий в природе.
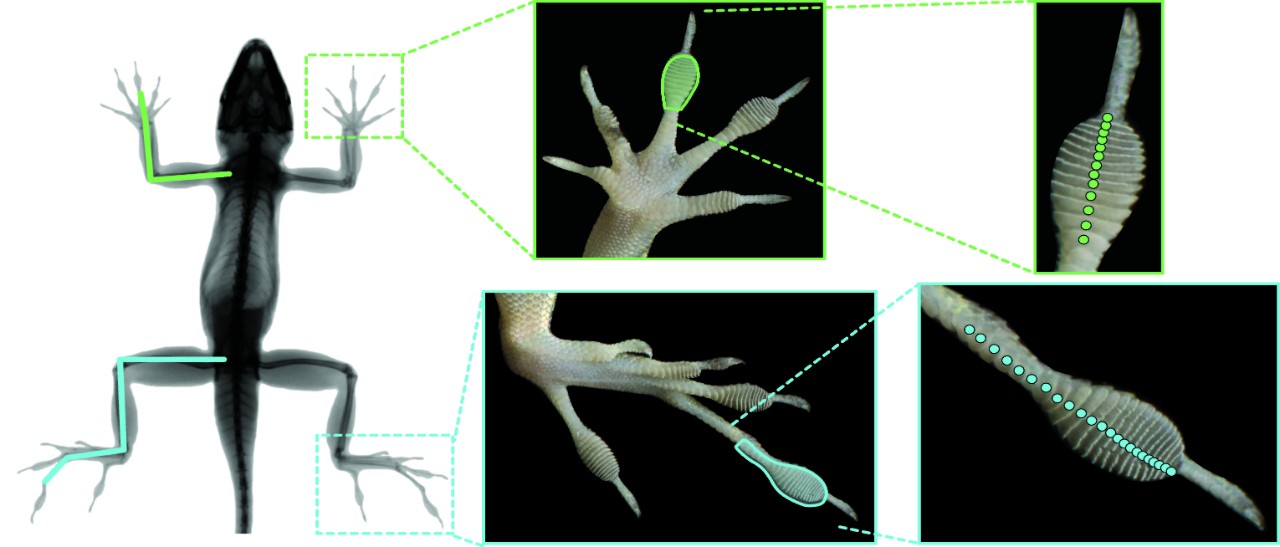
Морфологические изменения у «городских» анолисов: длинней ноги, больше подушечки лап, больше специализированных чешуй, улучшающих сцепление с вертикальной поверхностью
Тогда возможны две рабочие гипотезы о роли вызванной температурой пластичности в освоении городских «островов тепла»:
1) исходная пластичность «лесных» популяций вполне подходит для решения этой задачи, выработки термостойкости, необходимой в урбосреде, т.е. используя рис.1, температурные модификации идут в правильную сторону (гипотеза идеальной пластичности);
2) Пластичность «лесных» негодна для достижения «городской» термостойкости и должна видоизмениться отбором, чтобы «переместить» популяции, колонизирующие города, поближе к адаптивному пику новой среды (это У на рис.1, гипотеза эволюционирующей пластичности).
Основное различие между 1) и 2) в необходимости или нет естественного отбора, меняющего генетическую компоненту вызванной температурой пластичности. Кэмпбелл-Стейтон и соавт.60 сообщают о параллельной картине роста дифференциации по большому числу разных генов, чувствительных к росту температуры, наблюдаемой в популяциях разных городов острова (сравнительно с ближайшими лесными). Они же идентифицируют отдельные несинонимичные полиморфизмы, связанные с повышенной теплостойкостью, но только в пределах городских «островов тепла»60. Постоянные и повторяющиеся признаки отбора генов, связанных с термостойкостью, независимо наблюдаемые в разных городах, согласуются с гипотезой 2)60.
Однако неясно, как именно отбор изменяет существующий спектр индуцированной температурой пластичности, чтобы менять термостойкость так быстро как наблюдается.
В этом исследовании авторы опираются на выводы Кэмпбелла-Стейтона и соавт.60, чтобы изучить относительную роль адаптивной и неадаптивной пластичности в экспрессии генов как мишеней отбора в урбосреде. Во-первых, используя данные по экспрессии генов Campbell-Staton и соавт.60, они охарактеризовали регуляторную подоснову температурно-зависимых изменений активности A.cristatellus, чтобы определить возможные гены-кандидаты [эволюционных изменений в связи с термостойкостью] в транскриптоме скелетных мышц задней конечности. Их отыскивали среди генов, чья экспрессия сильно и значимо коррелирует с термостойкостью (CTmax, рис. 3А, В и дополнительный рис.S2).
Этот набор генов-кандидатов служит прокси для транскрипционных факторов формирования термостойкости, представляющих собой сплав модификационной изменчивости (пластичность развития и фенотипическая гибкость) и эволюционионно созданной дивергенции «городских» ящериц (рис.4). Дальше исследовали эволюционный разрыв по пластичности между городскими и лесными ящерицами путем оценки относительного вклада адаптивной и неадаптивной пластичности в экспрессию генов-кандидатов, используя ящериц, рожденных и выращенных в обычных лабораторных условиях.
В заключение, авторы сравнивали генетическую дивергенцию и механизмы отбора, связанные с адаптивной и неадаптивной пластичностью экспрессии генов в четырех парах городских и лесных популяций. Они делали это для измерений относительной величины генетической дивергенции между альтернативными средами обитания для генов, связанных с каждой из категорий пластичности (адаптивной или неадаптивной, рис.7). Так идентифицируются предполагаемые молекулярные мишени отбора в каждом случае (рис.8). Используя эту последовательность и дополняющих друг друга анализов, мы предполагаем, что быстрая адаптации городских анолисов к «островам тепла» результат прежде всего уменьшения или обращения вспять неадаптивной пластичности отбором (неадаптивной в условиях городских поверхностей и температур) .
Результаты и обсуждение
Идентификация регуляторной подосновы активности ящериц, зависящей от температуры
В статье представлен новый и расширенный анализ данных, собранных Кэмпбеллом-Стейтоном и соавт.60, для дальнейшего изучения регуляторной эволюции в масштабе всего транскриптома для скелетных мышц задних конечностей, важнейшего компонента локомоции61,62. Он выявил в урбосреде 4х разных городов повторяющиеся паттерны пластичности, индуцированной температурой, и её изменения отбором в городских островах тепла. Вкратце, Кэмпбелл-Стейтон и соавт.60 измерили CTmax для 4х пар популяций A.cristatellus, городских vs лесных (гг.Агуадилья, Аресибо, Маягуэс и Сан-Хуан, колонизированные ими независимо) (n = 11460).
Критический максимум температуры (CTmax) показывает порог, за которым её рост вызывает системную дисфункцию63 — неспособность координировать двигательную активность, в том числе вставать на ноги при поворачивали на спину64. CTmax также определили для городских и лесных ящериц из Маягуэза, родившихся и выросших в обычных лабораторных условиях (n = 16). Далее этих же ящериц 2 ч подвергали действию одной из трёх вариантов температур, соответствующих привычным для них местам обитания (“лесных” c дневной температурой 25,4±0,614°C; ночной 15,49±1,82°C; “городских” с дневной 32,09±3,1°C). Выращенных в лаборатории ящериц случайным образом распределили в две группы, одни испытывали на себе «лесные» условия, другие — «городские». Когда все эти ящерицы более-менее привыкли к новой жизни, от них взяли транскриптомы скелетных мышц и секвенировали РНК (RNAseq) (см. “Методы” и допматериалы рис.2).
Чтобы классифицировать регуляторную архитектуру транскриптома скелетных мышц анолисов, в данных по экспрессии генов у всех этих особей (пойманных в природе n = 114, выращенных в лаборатории = 16, Допматериалы рис.1) искали группы коэкспрессируемых генов (модули). «Работа» их сильно связана, и эту связь (силу и значимость) можно определить статистически, с использованием WGCNA65, взвешенного корреляционного сетевого анализа. Он же определяет уровень экспрессии внутри каждого модуля, выявляет в нём регуляторно важные гены (гены-хабы) и связывает паттерн экспрессии с вариацией признака, здесь — термостойкостью65.
Этим подходом авторы идентифицировали гены в транскриптоме скелетных мышц, чья экспрессия была значительно связана с CTmax, используя функцию значимости гена (GS65, определяется корреляцией уровня экспрессии гена со значением признака, в нашем случае критической температурой CTmax). Всего генов в транскриптоме было 11654; чтобы учесть модульную структуру внутри транскриптома, гены разделили на семь модулей экспрессии, которых достаточно для описания регуляторной архитектуры, установленной WGCNA60 (рис.3A,B). В модулях установили в общей сложности 632 гена, у которых экспрессия, значимо коррелирует с CTmax (с учётом поправки для множественных сравнений GS.q < 0,05) (рис.3B). Этот набор генов, ассоциированных с CTmax, считается предполагаемыми генами-кандидатами, участвующими в формировании термостойкости.
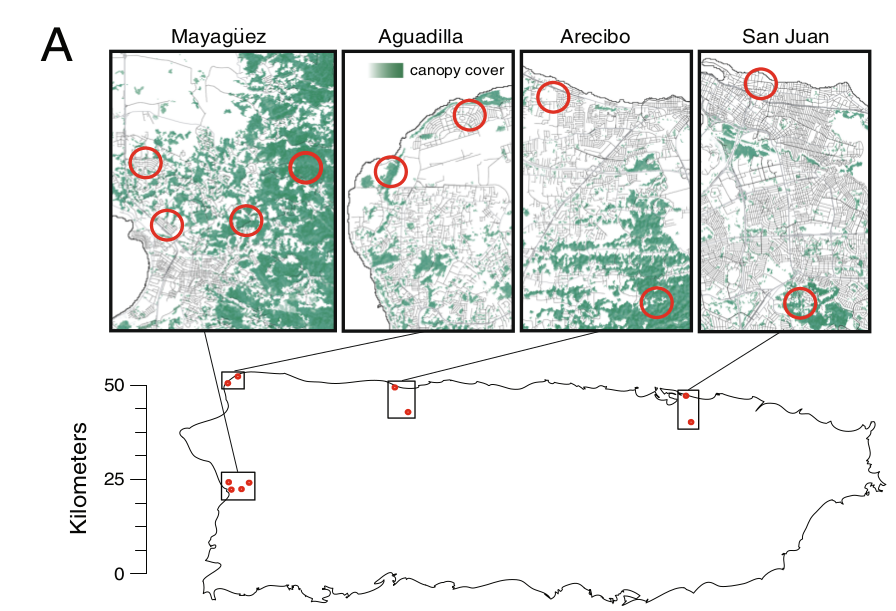
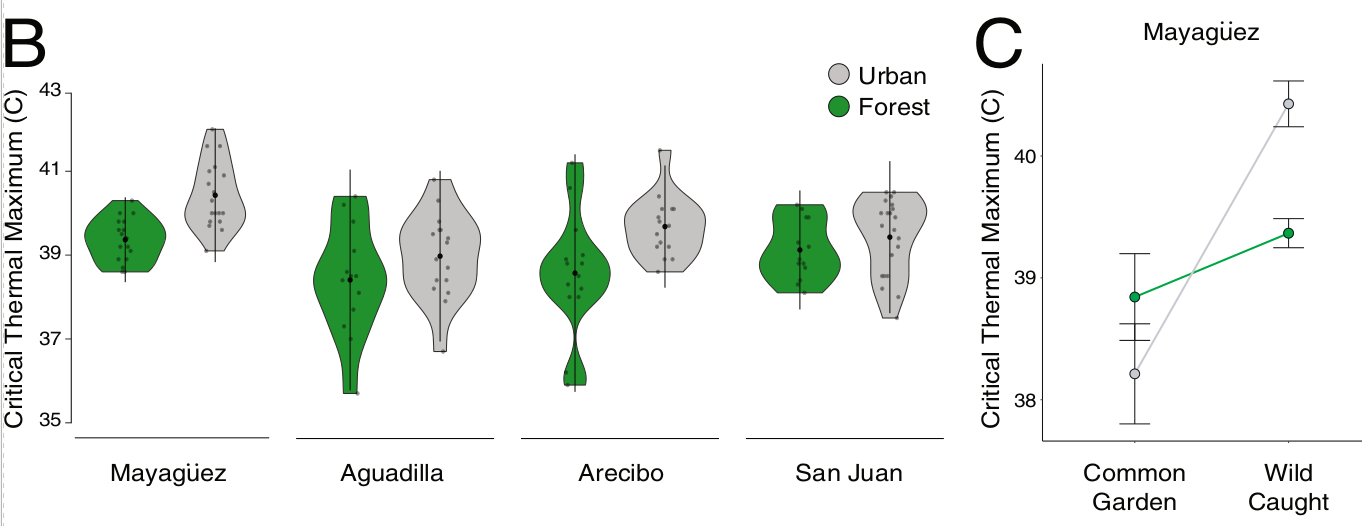 Рис.2. Дивергенция в термостойкости между городскими и лесными популяциями. А. Карта Пуэрто-Рико с местами отлова ящериц60. B. графики теплостойкости (критический тепловой максимум, CTmax), измеренные для ящериц лесных (зеленый цвет) и городских (серый) популяций из 4х городов (Агуадилья: лес n = 16, город n = 16; Аресибо: лес n = 21, город n = 20; Маягуэс: лес n = 18, город n = 18; Сан-Хуан: лес n = 16, город n = 25). Во всех 4х городские анолисы демонстрируют значимо большую термостойкость, чем их лесные собратья (линейная модель смешанных эффектов: 0,8247 ± 0,1771, χ2 = 20,093, р < 0,001, величина эффекта = 0,33760). Черные точки и вертикальные линии в центре каждой фигуры соответственно средняя и стандартное отклонение (SD). C. Сравнение CTmax “городских” (серый) и лесных (зеленый) анолисов г. Маягуэза, родившихся и выращенных в лаборатории (n = 16; “лесные”: 25 °C n = 3, 32 °C = 4; “городские”: 25°C n = 5, 32°C = 4) vs пойманных в природе (n = 36, лесной n = 18, городской n = 18). Точки с «усами» вокруг — среднее значение ± его стандартная ошибка (SE).
Рис.2. Дивергенция в термостойкости между городскими и лесными популяциями. А. Карта Пуэрто-Рико с местами отлова ящериц60. B. графики теплостойкости (критический тепловой максимум, CTmax), измеренные для ящериц лесных (зеленый цвет) и городских (серый) популяций из 4х городов (Агуадилья: лес n = 16, город n = 16; Аресибо: лес n = 21, город n = 20; Маягуэс: лес n = 18, город n = 18; Сан-Хуан: лес n = 16, город n = 25). Во всех 4х городские анолисы демонстрируют значимо большую термостойкость, чем их лесные собратья (линейная модель смешанных эффектов: 0,8247 ± 0,1771, χ2 = 20,093, р < 0,001, величина эффекта = 0,33760). Черные точки и вертикальные линии в центре каждой фигуры соответственно средняя и стандартное отклонение (SD). C. Сравнение CTmax “городских” (серый) и лесных (зеленый) анолисов г. Маягуэза, родившихся и выращенных в лаборатории (n = 16; “лесные”: 25 °C n = 3, 32 °C = 4; “городские”: 25°C n = 5, 32°C = 4) vs пойманных в природе (n = 36, лесной n = 18, городской n = 18). Точки с «усами» вокруг — среднее значение ± его стандартная ошибка (SE).
Линии, соединяющие значения критических температур в природе и в лаборатории, подчеркивают резкие изменения признака между ними, со значительным изменением пластичности отбором в городах. Поэтому не отмечено значимых различий в CTmax между ящерицами городского и лесного происхождения в условиях лаборатории. Наследственная составляющая этой пластичности может быть мишенью отбора в новых условиях городских «островов тепла».
Функциональный анализ состава генов-кандидатов выявил перепредставленность среди них участников трёх взаимосвязанных процессов, образующих клеточную реакцию на тепловой шок. Это стимуляция биогенеза клеточных компонентов, p = 0,036 (в «Генной онтологии» (англ. Gene Ontology, справочнике по функциям разных генов, его позиция GO: 0044089); регуляция сборки комплекса предварительной инициации транскрипции РНК-полимеразы II, p = 0,0004 (в Gene Ontology GO:0045898) и катаболизма клеточных макромолекул (в GO:0044265, p = 0,004). Всё три процесса разделяют основной набор генов с комплексом предварительной инициации транскрипции РНКII (Дополнительная таблица 2). Последний же играет решающую роль в регуляции клеточного ответа на тепловой шок: инициирует транскрипцию генов, продуцирующих белки теплового шока66,67. Четвертый биологический процесс [в котором также участвуют гены кандидаты] (GO:0007059) — сегрегация хромосом, p=0,025. Связанные с ним гены участвуют в репликации и репарации ДНК, процессах имеющих решающее значение для поддержания целостности ДНК во время и после клеточного теплового шока68. Сюда входят гены кодирующие белки кинезин, конденсин, белки кинетохор, РНК-хеликазу и ДНК-топоизомеразу II типа (Дополнительная таблица 2). Все вместе эти функциональные ассоциации подчеркивают потенциальные клеточные механизмы, лежащие в основе индивидуальных различий в толерантности к жаре, и образуют возможные цели отбора на термостойкость в городах.
Авторы объединили всех ящериц, из лаборатории и природы, для лучшей идентификации генов-кандидатов, связанных с термоустойчивостью. Поэтому выявленный набор генов-кандидатов отражает корреляции экспрессии генов и фенотипа (СТmax) из-за комбинированных эффектов развитийной пластичности, связанной с температурой выращивания, и фенотипической гибкости, связанной с температурой опыта, плюс взаимодействия между ними. Учет этого различия в потенциальном регуляторном вкладе был сочтен критическим для последующих анализов, поскольку отбор по любому из этих компонентов может играть важную роль в адаптивном температурно-зависимом расхождении экспрессии генов между городскими и лесными анолисами.
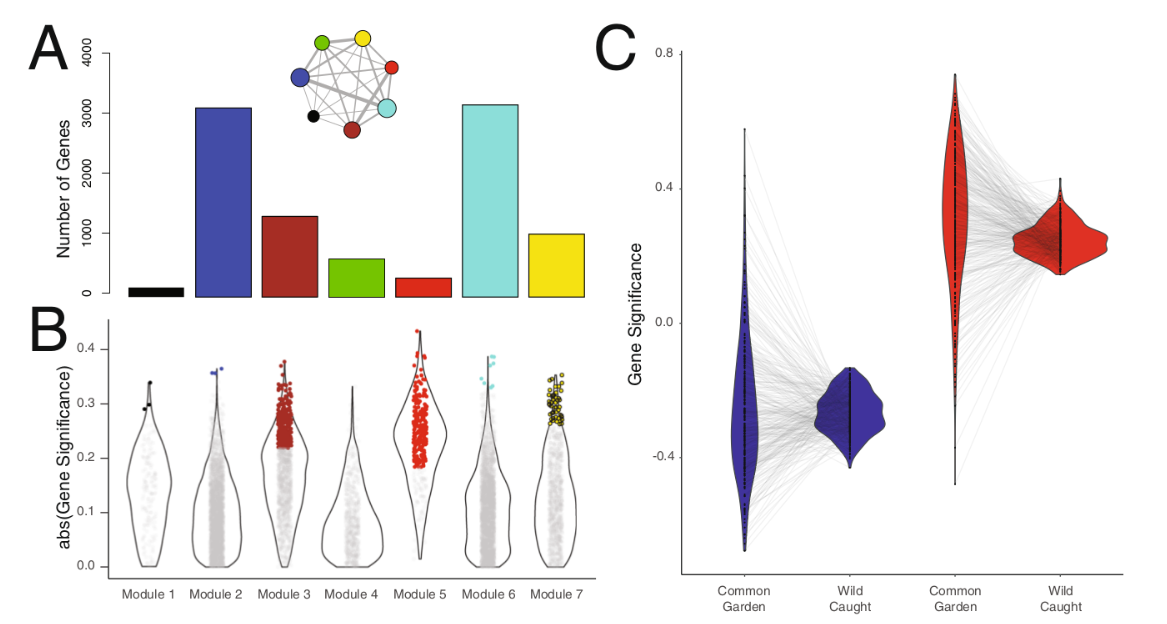 Рис.3. Идентификация генов, связанных с термостойкостью, у ящериц из лаборатории и природы. А. Для идентификации модулей коэкспрессии генов транскриптома скелетных мышц использовано 16 первых и 114 вторых, всего 130 особей. WGCNA выявил 7 модулей, гены которых, экспрессируясь согласованно в каждом из них, регулируют термостойкость: модуль 1 (черный), Модуль 2 (синий), Модуль 3 (коричневый), Модуль 4 (зеленый), Модуль 5 (красный), модуль 6 (бирюзовый), и модуль 8 (желтый)60. Столбцы — число генов в каждом модуле. Врезка над столбцами — корреляционная плеяда между активностями разных модулей, размеры кружков в ней показывают относительную величину каждого модуля (их площадь окружностей пропорциональна логарифму числа генов в каждом), а толщина линий степень ассоциированности «работы» разных модулей между собой (абсолютное значение попарной корреляции Пирсона между значениями собственных генов). В. Значимость генов для каждого из 7 модулей для термостойкости ящериц (GS, показано на абсциссе, скрипичные диаграммы слева направо — модули 1-7), определяется силой связи между экспрессией гена и CTmax. Точки внутри этих фигур — отдельные гены данного модуля. Цветные точки на каждой из фигур — гены данного модуля, чьë GS осталось значимым после множественной перепроверки (GS.q < 0,05). Это самые важные гены в смысле формирования термостойкости; их подмножество составило основной набор данных текущего исследования. C. Для подтверждения важности генов, выявленных предыдущим анализом дополнительно провели ещё 2 раздельно для ящериц из природы (n = 114) и лаборатории (n = 16), с расчётом GS уже независимо для каждой группы. Синий цвет — гены, понижающие CTmax, отрицательные регуляторы, их экспрессия коррелирует с CTmax отрицательно; красный — гены-усилители, их экспрессия положительно коррелирует с CTmax. Черные точки в центре каждой скрипичной диаграммы — оценки GS для отдельных генов, серые линии — направления изменений GS разных генов «красной» и «синей» групп между выращенными в лаборатории и пойманными в природе. Видна сильная корреляция уровня экспрессии и CTmax между данными, полученными в природе, и в лаборатории (линейная модель, R2: 0,601; p « 0,001).
Рис.3. Идентификация генов, связанных с термостойкостью, у ящериц из лаборатории и природы. А. Для идентификации модулей коэкспрессии генов транскриптома скелетных мышц использовано 16 первых и 114 вторых, всего 130 особей. WGCNA выявил 7 модулей, гены которых, экспрессируясь согласованно в каждом из них, регулируют термостойкость: модуль 1 (черный), Модуль 2 (синий), Модуль 3 (коричневый), Модуль 4 (зеленый), Модуль 5 (красный), модуль 6 (бирюзовый), и модуль 8 (желтый)60. Столбцы — число генов в каждом модуле. Врезка над столбцами — корреляционная плеяда между активностями разных модулей, размеры кружков в ней показывают относительную величину каждого модуля (их площадь окружностей пропорциональна логарифму числа генов в каждом), а толщина линий степень ассоциированности «работы» разных модулей между собой (абсолютное значение попарной корреляции Пирсона между значениями собственных генов). В. Значимость генов для каждого из 7 модулей для термостойкости ящериц (GS, показано на абсциссе, скрипичные диаграммы слева направо — модули 1-7), определяется силой связи между экспрессией гена и CTmax. Точки внутри этих фигур — отдельные гены данного модуля. Цветные точки на каждой из фигур — гены данного модуля, чьë GS осталось значимым после множественной перепроверки (GS.q < 0,05). Это самые важные гены в смысле формирования термостойкости; их подмножество составило основной набор данных текущего исследования. C. Для подтверждения важности генов, выявленных предыдущим анализом дополнительно провели ещё 2 раздельно для ящериц из природы (n = 114) и лаборатории (n = 16), с расчётом GS уже независимо для каждой группы. Синий цвет — гены, понижающие CTmax, отрицательные регуляторы, их экспрессия коррелирует с CTmax отрицательно; красный — гены-усилители, их экспрессия положительно коррелирует с CTmax. Черные точки в центре каждой скрипичной диаграммы — оценки GS для отдельных генов, серые линии — направления изменений GS разных генов «красной» и «синей» групп между выращенными в лаборатории и пойманными в природе. Видна сильная корреляция уровня экспрессии и CTmax между данными, полученными в природе, и в лаборатории (линейная модель, R2: 0,601; p « 0,001).
Относительный ранг значимости генов был высоко консервативен во всех наборах данных (ранговая корреляция Спирмена ρ = 0,75, p « 0,001). Отрицательные регуляторы не показали смещений в значимости генов между группами (парный t-тест, p = 0,26). Положительные регуляторы показали большие значения GS в данных из лаборатории, чем из природы (парный t-тест, p « 0,001). Корреляции между генами из природных vs лабораторных данных были невелики но статистически значимы для обеих групп генов (доля объяснённой дисперсии для генов-усилителей: R2 = 0,04, р < 0,0001; ослабителей R2 = 0,03, р = 0,003).
Дальше исследовали пригодность найденных генов-кандидатов для воспроизведения тех же корреляций уровня экспрессии с термостойкостью раздельно в природе и лаборатории. Для этого ещё раз использовали WGCNA отдельно для каждой из этих групп. Дальше авторы пересчитали GS для каждого гена-кандидата и сравнили их в этих двух группах (рис.3C). Оказывается, что они сильно коррелируют между собой, то есть экспрессия генов управляет термостойкостью в природе примерно также, как в лаборатории, там и там гены одинаково ранжируются по этому признаку, разница в управлении чисто количественная (линейная модель, скорректированный R2 = 0,601; p<0,001), поэтому относительный ранг GS высоко консервативен между обеими группами (ранговая корреляция Спирмена ρ = 0,631, p<0,001).
Среди генов-ослабителей термостойкости вообще не отмечено значимых различий в GS между природой и лабораторией (парный t-тест, t = 0,81, df = 295, p = 0,42), у усилителей в лаборатории был значимо больший GS, чем в природе (парный t-тест, t = 5,58, df = 335, p « 0,001). Иными словами, в лаборатории термостойкость точнее предсказывается по экспрессии найденных генов [а в природе эти корреляции слабее, ибо изменены отбором]. Большая дисперсия в лабораторных условиях снижает объяснительную силу [полученных там данных] внутри каждой категории регуляторов [при использовании опытов в лаборатории для объяснения процессов в природе].
Тем не менее, корреляция между данными из природы и лаборатории везде значима (у усилителей R2 = 0,04, p < 0,0002; ослабителей R2 = 0,03, р = 0,0003). Поэтому выявленный авторами набор генов-кандидатов годен для анализа в обоих средах обитания природной и лабораторной.
Чтобы непосредственно оценить роль эволюции в сдвигах экспрессии этих генов-кандидатов, авторы искали геномные признаки отбора в каждой из популяций с использованием всех особей (из природы = 114, лаборатории = 16). Если пластичность, образующая наблюдаемые различия в термостойкости, выступает мишенью отбора при освоении городов, должна быть выявлена геномная «подпись» селективных процессов в виде изменений «букв» генетических «текстов» именно там где находятся локусы — кандидаты.
Авторы идентифицировали 2161083 однонуклеотидных полиморфизма (SNP) в масштабах всего транскриптома с использованием считывания RNAseq у яшериц, происходящих из всех четырех муниципалитетов, изученных в Campbell-Staton et al.60 (Агуадилья: лесные n = 11, городские n = 16; Аресибо: лесные n = 18, городские n = 16; Маягуэс: лесные n = 19, городские n = 18; Сан-Хуан: лесные n = 11, городские n = 21). Используя все полиморфные локусы в генах-кандидатах в качестве фокального набора SNP-возможно изменяемых естественным отбором, авторы проверяли, есть или нет значимых отклонений от нейтральности путем прямого сравнения с нуль-гипотезой распределения SNP при отсутствии отбора. Последнее конструировали по «фоновой» вариации SNP69 в тех генах, чья экспрессия не коррелирует с CTmax (рис.4).
Авторы рассчитали неравновесие попарных связей (LD; R2 в пределах неперекрывающихся окон размером 10 кб) и протестировали отклонения от равновесия Харди-Вайнберга (HW) по всем полиморфным сайтам транскриптома. Затем они сравнили паттерны LD (привязанные с шагом 1 кб) между эмпирическими последовательностями SNP-кандидатов и «фоновыми», рассчитанными в рамках 0-гипотезы.
На равновесие LD и HW могут влиять как отбор, так и демография, поэтому на каждом участке анализ был ограничен такими сравнениями, чтобы исходные и кандидатные наборы SNP имели общую демографическую историю. Тогда если найденные кандидатные гены будут мишенями одного и того же отбора на термостойкость, можно ждать значимого роста LD в полиморфных сайтах, близких к этим генам, по сравнению с фоновым для транскриптома. Действительно, SNP в генах, экспрессия которых коррелирует с CTmax, показывает значимый рост LD (регрессионный анализ, линейная модель смешанных эффектов: p < 0,05, рис.4) во всех 4х городах. Здесь же более распространены и отклонения от равновесия HW (тест на равенство пропорций-Аресибо: χ2= 77,721, df = 1, p << 0,001; тест на равенство пропорций — Аресибо: χ2= 77,721, df = 1, p << 0,001; Агуадилья: χ2= 28,478, df = 1, p << 0,001; Майягуэз: χ2= 37,71, df = 1, p << 0,001; Сан-Хуан: χ2= 33,383, df = 1, p << 0,001; Дополнительная таблица 3).
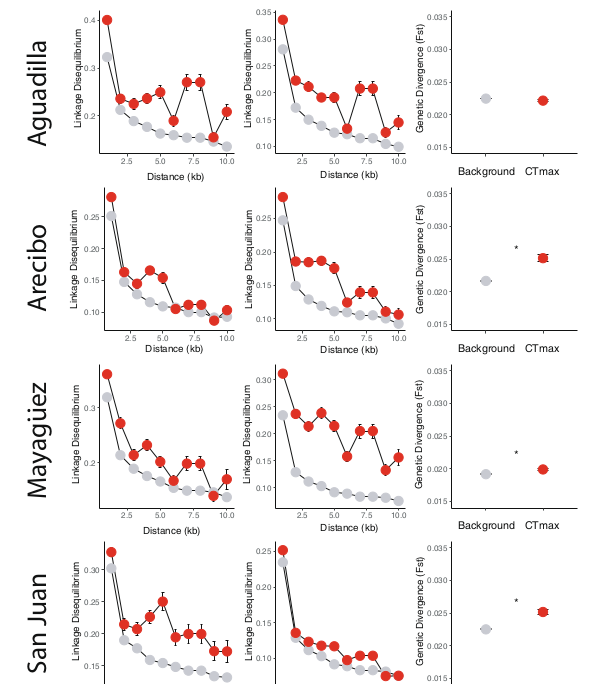
Рис.4. «Подписи» полигенного отбора среди SNP, ассоциированных с термостойкостью. Расширенное неравновесие по сцеплению (≥5 т.п.н.) отмечено в SNP, ассоциированных с CTmax (красный) (регрессионный анализ, линейная модель смешанных эффектов: p <0,05) и у «лесных» (столбец 1), и у «городских» ящериц (столбец 2), сравнение с «фоном» для всего транскриптома (серый). Это показывает продолжающийся отбор по физиологии термочувствительности на всём острове. Выявленная в 3х из 4х городов значимая дивергенция потоковых популяций от исходных (Столбец 3) по SNP, связанных с термостойкостью (красный) (Аресибо, Маягуэс и Сан-Хуан: t-критерий Уэлча; p < 0,05 (звездочки)) также показывает отбор на термоустойчивость, адаптирующий анолисов к ещё более экстремальным «островам тепла» (сравнение с «фоном» для всего транскриптома — серый). Графики LD представлены в бинах по расстоянию 1 кб. Точки и полосы ошибок - средние ± 1 SE. В анализе использованы все 130 животных, для которых были данные о последовательностях.
Эти геномные паттерны показывают параллельные направления отбора по генам, вовлечённым в регуляцию термостойкости в разных частях ареала A. cristatellus. Результат подтвердил выводы предыдущей работы, демонстрирующей неотъемлемую роль биологии терморегуляции в специализации среды обитания у анолисов, занимающих различные макро- и микроклиматические ниши70-72. Если, в дополнение к этому более широкому отбору по термической физиологии, городские популяции вида сегодня испытывают сильный отбор условиями «островов тепла», следует ожидать повышенную генетическую дивергенцию (Fst) между » городскими» и «лесными» ящерицами при сравнении SNP в локусах–кандидатах с ожидаемыми исходя из их «фонового» состава. Авторы обнаружили это в трех из наших четырех сравнений тех и других (Аресибо, Маягуэс и Сан–Хуан: t-тест Уэлча с двумя выборками, p << 0,001, рис.4). Это подтверждает предположение, что гены, ассоциированные с CTmax, являются повторяющимися объектами отбора при заселении всех городов острова, происходящем независимо из ближайших лесов.
Экспериментальная проверка дивергенции «городских» и «лесных» анолисов по пластичности генной экспрессии
Дальше авторы экспериментально исследовали в лаборатории эволюционную дивергенцию по пластичности экспрессии генов между городскими и лесными анолисами. Сперва они классифицировали индуцированную температурой пластичность экспрессии каждого гена как адаптивную или неадаптивную. Для этого они предположили, что большая теплостойкость адаптивна при более высоких температурах51, тогда как меньшая нет, ибо снижает выносливость73,74. Используя оценки GS из анализа WGCNA, описанного выше, авторы определяли знак корреляции изменения экспрессии и изменения термостойкости (фенотипа) как (+) или (-) для каждого гена-кандидата, ассоциированного с CTmax. Затем они характеризовали изменения экспрессии, наблюдаемые для каждой группы лабораторных ящериц, подвергавшихся 2 ч воздействию средних лесных (25 °C; лес n = 4, город n = 5) или городских (32 °C, лес n = 3, город n = 4) температур как конгруэнтные (адаптивные) или неконгруэнтные (неалаптивные) происходящим изменениям термостойкости.
Случаи, где направление лабораторной пластичности соответствовало ожиданиям увеличения CTmax, предполагаются адаптивными, противоположные неадаптивными. Схему процесса категоризации, разделяющей изменчивость экспрессии на данные категории см. на рис.5. Лесные популяции A.cristatellus существовали и до урбанизации, изменившей среду обитания. Поэтому анолисов содержащихся в лаборатории при «лесных» температурах (n = 7; 25°C n = 3, 32°C = 4) использовали как прокси исходного состояния генной экспрессии и сравнивали с производным (содержание при «городских» температурах, n = 9, 25°C; n = 5, 32°C = 4).
Выяснилось, что выращенные в лаборатории ящерицы «городского» и «лесного» происхождения из Маягуэза сильно и значимо различаются по пластичности экспрессии генов, вызванной высокой температурой, поддерживая [вывод] об эволюционной дивергенции между ними. На термический стресс «лесные» анолисы изменяли экспрессию преимущественно неадаптивно (82,3% генов, n = 520, биномиальный тест: p << 0,001, рис. 6A,B), при подавленном состоянии большинства (82,7%) положительных регуляторов (n = 293, p << 0,001), у большинства отрицательных (76,7%) при повышенной температуре всё наоборот (n = 227, p << 0,001).
Эти результаты показывают, что в исходных популяциях гены влияющие на термостойкость, адаптированны к меньшим критическим температурам, а при их повышении в условиях городов те же регуляторные изменения оказываются неадаптивными. Анолисы «городского» происхождения, наоборот, в лаборатории показали высокую приспособленность регуляторов к условиям городских «островов тепла». Неадаптивная пластичность генов-кандидатов на регуляцию термостойкости здесь значимо снижена (лес = 82,3%, город = 54,3%, точный биномиальный р < 0,001) при возрастании адаптивной пластичность (лес = 17,7%, город = 45,7%, точный биномиальный р < 0,001) (рис. 6B).
Отмеченная дивергенция ящериц «лесного» и «городского» происхождения по доле адаптивной vs и неадаптивной пластичности позволила предположить, что отбор связанный с освоением урбосреды, в основном направлен на сокращение второй или её обращение вспять. При таком сценарии можно ждать общего снижения пластичности экспрессии, индуцированной ростом температуры, в городских популяциях, производных от лесных своего региона75. Поэтому ожидаема отрицательная корреляция направления эволюции (усиливающего термостойкость) с направлением исходной пластичности (при урбанизации она уменьшается)75.
Проверяя эту гипотезу, авторы оценили наследственную пластичность выявленных генов-кандидатов логарифмами изменений экспрессии у ящериц «лесного» происхождения между «лесной» и «городской» температурами в лаборатории (25°C n = 3, 32°C = 4), а эволюционную дивергенцию — логарифмами изменений экспрессии между «лесными» и «городскими» анолисами, когда их содержат при температуре 32°C (по 4 шт каждых). Затем они вычислили знак и силу корреляций между исходной пластичностью и эволюционной дивергенцией по всему набору генов-кандидатов, ассоциированных с CTmax, для ящериц из Майягуэза.
Данные подтвердили отбор против неадаптивной пластичности: пластичность предковой популяции и эволюционная дивергенция коррелируют сильно и отрицательно (ρ = -0,61; 82,3% генов, n = 520, тест на равенство пропорций, р < 0,001, рис.6А). Для исключения потенциально возможного артефакта, связанного с регрессией к среднему76, авторы создали нулевое распределения коэффициентов корреляции, использовав рандомизационный тест, и сравнили с ним наблюдаемые корреляции. Последние были значительно более отрицательны чем если бы имел место лишь артефакт (эмпирический р < 0,05, врезка на рис.6А). Если отбор главным образом уменьшал неадаптивную пластичность, можно ждать значимого снижения величин индуцированных жарой изменениях в экспрессии генов у «городских» ящериц сравнительно с «лесными». Тогда они напрямую сравнили величины изменений в экспрессии при содержании первых при 25°C вторых при 32°C в условиях лаборатории (“лесные” 25°C n = 3, 32°C = 4; “городские” 25°C n = 5, 32°C = 4). Действительно, у «городских» анолисов реакция на жару притупляется (ниже логарифм количественных изменений уровня экспрессии), притом что она адаптивна.
Иными словами, отбор в новой экстремальной среде лишь «отсекает лишнее» в пластичности, вызванной напряжением сил особей предковой популяции вынужденных существовать в новых условиях и уже изменившихся в этой связи сравнительно с оставшимися в лесу. Взятые вместе, эти различия уверенно подтверждают выдвинутую рабочую гипотезу.
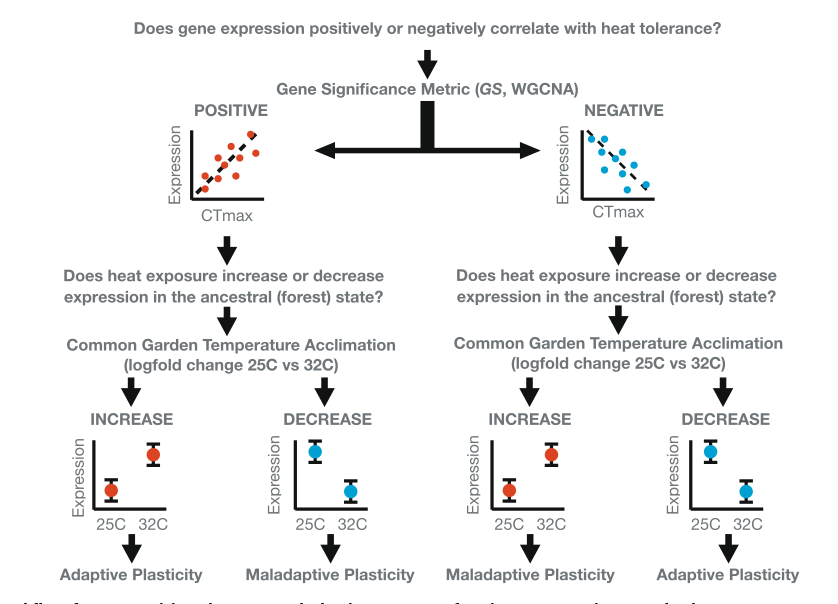 Рис.5. Теоретический алгоритм для классификации температурно-индуцированных изменений экспрессии генов предковой популяции при освоении ею городов как адаптивной или неадаптивной. На первом шаге анализа знак корреляции изменений в экспрессии генов и изменений в CTmax (измеренное в группе из 130 ящериц) использовали для выяснения, кто из найденных кандидатных генов положительный (коэффициент корреляции обоих величин >0) а кто отрицательный регулятор (он же <0). На втором шаге определяли как меняется экспрессия каждого из таких генов (усиливается или ослабевает, повышена или снижена «наработка» генных продуктов) у ящериц из «лесных» популяций (филогенетически исходное состояние), выращенных в обычных условиях (25°C) и подвергнутых температурам городских «островов тепла» (32°C). Столбцы с «усами» на схемах показывают предполагаемые изменения средних ± 1 SE. К адаптивной пластичности отнесены а) положительные регуляторы, увеличивающие экспрессию с ростом температуры, и б) отрицательные регуляторы, уменьшающие в тех же условиях (т.е. всё случаи совпадения направлений изменений пластичности генной экспрессии и эволюционирующего признака). К неадаптивной пластичности отнесли все случаи несовпадения направлений изменения признака и изменений пластичности а) пониженная экспрессия положительных регуляторов с ростом температуры иди б) повышенная экспрессия отрицательных регуляторов в тех же условиях.
Рис.5. Теоретический алгоритм для классификации температурно-индуцированных изменений экспрессии генов предковой популяции при освоении ею городов как адаптивной или неадаптивной. На первом шаге анализа знак корреляции изменений в экспрессии генов и изменений в CTmax (измеренное в группе из 130 ящериц) использовали для выяснения, кто из найденных кандидатных генов положительный (коэффициент корреляции обоих величин >0) а кто отрицательный регулятор (он же <0). На втором шаге определяли как меняется экспрессия каждого из таких генов (усиливается или ослабевает, повышена или снижена «наработка» генных продуктов) у ящериц из «лесных» популяций (филогенетически исходное состояние), выращенных в обычных условиях (25°C) и подвергнутых температурам городских «островов тепла» (32°C). Столбцы с «усами» на схемах показывают предполагаемые изменения средних ± 1 SE. К адаптивной пластичности отнесены а) положительные регуляторы, увеличивающие экспрессию с ростом температуры, и б) отрицательные регуляторы, уменьшающие в тех же условиях (т.е. всё случаи совпадения направлений изменений пластичности генной экспрессии и эволюционирующего признака). К неадаптивной пластичности отнесли все случаи несовпадения направлений изменения признака и изменений пластичности а) пониженная экспрессия положительных регуляторов с ростом температуры иди б) повышенная экспрессия отрицательных регуляторов в тех же условиях.
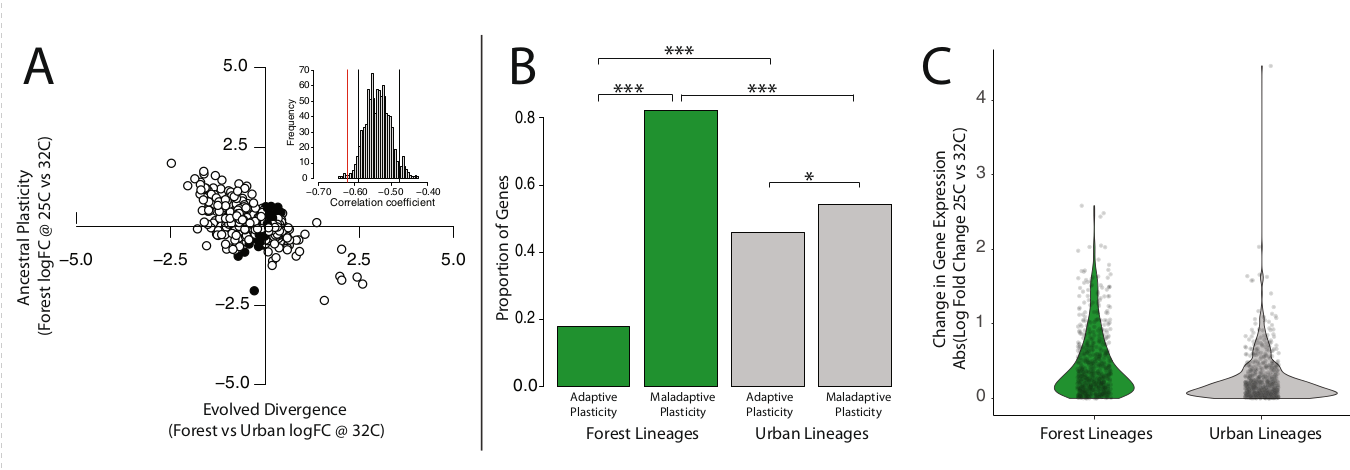 Рис.6. Экспериментальная оценка созданного эволюцией расхождения в генной экспрессии между «городскими» и «лесными» анолисами. Ящериц из той и другой популяции, отловленных у Маягуэса, растили в обычных лабораторных условиях, соответствующих «лесной» среде обитания, затем подвергали воздействию «городских» температур (32°C вместо 25°C) и смотрели изменения экспрессии генов, адаптивные и не адаптивные. А. Зависимость между изменением экспрессии генов при воздействии жары на исходную (лесную) популяцию (ордината двухкоординатного пространства) и изменением экспрессии тех же генов при образовании городской популяции, адаптированной к «острову тепла» (абсцисса). Каждая точка — один ген-кандидат,чья экспрессия коррелирует с термостойкостью (n=632), чёрный — гены, у которых изменения по обеим осям коррелируют положительно, белый — отрицательно, врезка — гистограмма распределения соответствующих коэффициентов для всех генов использованного набора в рамках 0-гипотезы (красная линия — средний коэффициент корреляции, видно, что он значимо отрицательней, чем нижний 95й процентиль рандомизированного распределения). Точки, выделенные черным цветом, представляют гены с положительной корреляцией между наследственной пластичностью и эволюционной дивергенцией. Микроэволюционные изменения генов, связанных с термостойкостью, при урбанизации вида, меняют направление фенотипической пластичности на противоположное (сравнительно с «лесной» популяцией). Вставка: наблюдаемый коэффициент корреляции (красная линия) более отрицательный, чем нижний 95-й процентиль рандомизированного нулевого распределения (черная линия). B. Столбики — число генов с адаптивной (слева) или неадаптивной (справа) пластичностью у анолисов, выращенных в лаборатории, но происходящих из лесных (зеленый цвет) и городских (серый) местообитаний. Именно у «городских» ящериц среди генов, связанных с обеспечением термостойкости, повышена доля проявивших адаптивную пластичность в ответ на рост температуры, при уменьшении доли проявивших неадаптивную. Звездочки — значимость различий между популяциями происхождения (тест на равенство пропорций в пределах типов местообитаний, точный биномиальный тест между типами местообитаний: * - p < 0,05, *** - p < 0,001). C. Скрипичные диаграммы показывают логарифм изменения экспрессии разных генов, вызванного ростом температуры с 25°C до 32°C у анолисов «лесного» (зеленый) и «городского» (серый) происхождения. Серые точки — отдельные гены. Городские линии демонстрируют в целом более низкую величину пластичности экспрессии генов (t-тест Уэлча с двумя выборками: p < 0,001), что соответствует эволюционному ослаблению неадаптивной пластичности. Для этих анализов использовали шестнадцать животных живших в обычных условиях лаборатории (25°C; происходящих из «лесной» популяции n = 4, «городской» n = 5) или «среднегородских» (32°C, «лесных» ящериц было n = 3, «городских» n = 4).
Рис.6. Экспериментальная оценка созданного эволюцией расхождения в генной экспрессии между «городскими» и «лесными» анолисами. Ящериц из той и другой популяции, отловленных у Маягуэса, растили в обычных лабораторных условиях, соответствующих «лесной» среде обитания, затем подвергали воздействию «городских» температур (32°C вместо 25°C) и смотрели изменения экспрессии генов, адаптивные и не адаптивные. А. Зависимость между изменением экспрессии генов при воздействии жары на исходную (лесную) популяцию (ордината двухкоординатного пространства) и изменением экспрессии тех же генов при образовании городской популяции, адаптированной к «острову тепла» (абсцисса). Каждая точка — один ген-кандидат,чья экспрессия коррелирует с термостойкостью (n=632), чёрный — гены, у которых изменения по обеим осям коррелируют положительно, белый — отрицательно, врезка — гистограмма распределения соответствующих коэффициентов для всех генов использованного набора в рамках 0-гипотезы (красная линия — средний коэффициент корреляции, видно, что он значимо отрицательней, чем нижний 95й процентиль рандомизированного распределения). Точки, выделенные черным цветом, представляют гены с положительной корреляцией между наследственной пластичностью и эволюционной дивергенцией. Микроэволюционные изменения генов, связанных с термостойкостью, при урбанизации вида, меняют направление фенотипической пластичности на противоположное (сравнительно с «лесной» популяцией). Вставка: наблюдаемый коэффициент корреляции (красная линия) более отрицательный, чем нижний 95-й процентиль рандомизированного нулевого распределения (черная линия). B. Столбики — число генов с адаптивной (слева) или неадаптивной (справа) пластичностью у анолисов, выращенных в лаборатории, но происходящих из лесных (зеленый цвет) и городских (серый) местообитаний. Именно у «городских» ящериц среди генов, связанных с обеспечением термостойкости, повышена доля проявивших адаптивную пластичность в ответ на рост температуры, при уменьшении доли проявивших неадаптивную. Звездочки — значимость различий между популяциями происхождения (тест на равенство пропорций в пределах типов местообитаний, точный биномиальный тест между типами местообитаний: * - p < 0,05, *** - p < 0,001). C. Скрипичные диаграммы показывают логарифм изменения экспрессии разных генов, вызванного ростом температуры с 25°C до 32°C у анолисов «лесного» (зеленый) и «городского» (серый) происхождения. Серые точки — отдельные гены. Городские линии демонстрируют в целом более низкую величину пластичности экспрессии генов (t-тест Уэлча с двумя выборками: p < 0,001), что соответствует эволюционному ослаблению неадаптивной пластичности. Для этих анализов использовали шестнадцать животных живших в обычных условиях лаборатории (25°C; происходящих из «лесной» популяции n = 4, «городской» n = 5) или «среднегородских» (32°C, «лесных» ящериц было n = 3, «городских» n = 4).
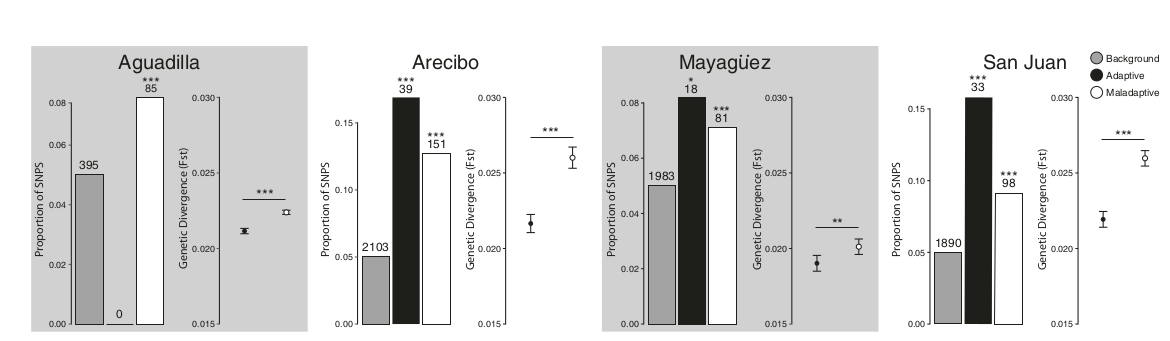 Рис.7. Параллельные сокращения неадаптивной пластичности при заселении анолисами 4х разных городов острова (колонизированы независимо друг от друга). Представлена величина дивергенции (Fst) для генов, связанных с термоустойчивостью, между «лесными» и «городскими» популяциями58. Гистограммы показывают долю однонуклеотидных полиморфизмов (SNP), чья встречаемость в генах с адаптивной (черный цвет) и неадаптивной пластичностью (белый) выходит за пределы 0-гипотезы распределения степени дивергенции, рассчитанного для всего транскриптома (серый цвет) в предположении что FST незначима, при p = 0,05. Звездочки — значимость изменений Fst (* - p < 0,05, ** - p < 0,01, *** - p < 0,001, тест на равенство пропорций в двух выборках с коррекцией на непрерывность), показывающая предпочтительную концентрацию изменившихся SNP в определённых группах генов.
Рис.7. Параллельные сокращения неадаптивной пластичности при заселении анолисами 4х разных городов острова (колонизированы независимо друг от друга). Представлена величина дивергенции (Fst) для генов, связанных с термоустойчивостью, между «лесными» и «городскими» популяциями58. Гистограммы показывают долю однонуклеотидных полиморфизмов (SNP), чья встречаемость в генах с адаптивной (черный цвет) и неадаптивной пластичностью (белый) выходит за пределы 0-гипотезы распределения степени дивергенции, рассчитанного для всего транскриптома (серый цвет) в предположении что FST незначима, при p = 0,05. Звездочки — значимость изменений Fst (* - p < 0,05, ** - p < 0,01, *** - p < 0,001, тест на равенство пропорций в двух выборках с коррекцией на непрерывность), показывающая предпочтительную концентрацию изменившихся SNP в определённых группах генов.
В популяции Агуадильи вообще не наши изменений SNP в генах, связанных с адаптивной пластичностью. Точки на графиках изменение средних значений Fst, усы — ± 1 SE. Звездочки — уровень значимости величин дивергенции «городских» и «лесных» популяций (* - p < 0,05, ** - p < 0,01, *** - p < 0,001, t-критерий Уэлча с двумя выборками). Использованы все 130 ящериц, для которых есть данные о последовательностях.
Сравнение генетической дивергенции и механизмов отбора действующего на адаптивную и неадаптивную пластичность экспрессии
Вышеописанные экспериментальные данные заставляют считать, что главную роль в эволюционной дивергенции «городских» популяций анолисов от «лесных» везде отбор, созданный необходимостью приспособления к повышенным городским температурам при урбанизации A.cristatellus. Тогда при сравнении генов тех и других можно ждать большей степени дивергенции среди генов, обуславливающих неадаптивную пластичность, чем у генов, обуславливающих пластичность адаптивную или дивергенцию по остальной части транскриптома. А разная степень дивергенции в соответствующих категориях генов, укажет различие целей отбора, вырабатывающего приспособления к урбосреде.
Поэтому авторы напрямую исследовали, действительно ли локусы, обуславливающие неадаптивную пластичность, быстрей дивергируют между вновь возникшими (городскими) и исходными (лесными) популяциями, если сравнивать с локусами, лежащими в основе адаптивной пластичности. Чтобы проверить это предсказание, авторы взяли SNP генов-кандидатов, ассоциированные с CTmax, и разделили на вовлечённые в осуществление а) неадаптивной и б) адаптивной пластичности и посмотрели значения Fst) для них по всем 4м парам исходных и производных популяций анолисов. Значимое превышение (выброс) Fst относительно «фона» доказывает полигенный отбор в данной группе генов, ассоциированных с фенотипом77, 78.
Сначала авторы численно охарактеризовали степень дивергенции генов из категорий а) и б) анализируя вклад в «выбросы» Fst генов обоих классов независимо для каждой из 4х пар сравниваемых популяций. Использовали данные по последовательностям всех особей (из природы n = 114, из лаборатории n = 16), представляющих каждый из муниципалитетов.
Выбросы Fst сравнивали с «фоном», степенью дивергенции каждой » городской» популяции от исходной «лесной», рассчитанным для всего транскриптома, учитывали различия значимые p < 0,05. Гены б) значимо обогащают «выбросы» (т.е. значимо участвуют в формировании дивергенции урбанизированных анолисов от исходных «лесных») только в 3 из 4х изученных городов — Аресибо, Маягуэс и Сан–Хуан — но гены б) — во всех (рис.7). Данные подтвердили действие отбора на оба класса пластичности, хотя оно сильней и облигатней на а).
Затем оценивали относительную силу отбора (коэффициент селекции) раздельно для а) и б), сравнив средние значения Fst для каждой пары «городских» vs «лесных» популяций (анолисов из природы n = 114, лаборатории n = 16). При освоении всех 4х городов гены а) дивергировали значимо сильнее, чем б) (t–критерий Уэлча с двумя выборками, p < 0,01) (рис.7). Хотя б) и а) равным образом проходят отбор на термостойкость в первом случае он интенсивней, особенно на начальных этапах отделения городских популяций.
Наконец, авторы исследовали потенциальный механизм, лежащий в основе а) и б), оценив долю некодирующих и кодирующих полиморфизмов, вовлечëнных в дивергенцию «городских» популяций от «лесных» (aнолисы из природы n = 114, лаборатории n = 16). Если эффект отбора проявляется главным образом в вариациях функции, связанной с несинонимичными полиморфизмами в кодирующих областях, можно ждать значимого повышения их доли среди кандидатных SNP, ассоциированных с CTmax, сравнительно с «фоном» по всему транскриптому. И наоборот, если действие отбора обусловлено в основном генетической изменчивостью цис-регуляторных элементов, среди них ожидаем значимый избыток некодирующих полиморфизмов. [Они находятся перед геном и регулируют его активность за счёт связывания транскрипционных факторов. Эти элементы обычно находятся рядом с промотором, вне кодирующей части гена, поэтому «цис-«. Регулятрные элементы, находящиеся далеко от «своего» гена — это энхансеры: обычно с них считывается РНК, но это часть механизма не синтеза белка, а обеспечения регуляции самого гена. Прим.публикатора]
Подтвердилась вторая гипотеза. В генах б) в генов выявлен значимый избыток несинонимичных полиморфизмов по сравнению с «фоном» (тест на равенство пропорций: χ2= 7,50; df = 1;p = 0,006, рис.8А). Однако такого вовсе не найдено в генах а) (χ2= 0,83; df = 1; p = 0,36, рис.8A). Иными словами, отбор формирует адаптивную пластичность за счёт изменений в структурных участках гена, а ликвидирует неадаптивную за счёт изменений регуляторных участков.
Чтобы непосредственно проверить последнюю гипотезу, авторы идентифицировали SNP в некодирующих областях транскриптома скелетных мышц, где могут быть предполагаемые цис-регуляторные элементы. Действительно, обнаружили, что некодирующие варианты перепредставлены среди генов, предположительно обуславливающих неадаптивную пластичность (χ2= 96,646, df = 1, p << 0,001, рис.8B) по сравнению с «фоном». И наоборот, среди генов б) нет избытка некодирующих SNP (χ2= 1,926, df = 1, p = 0,165, рис.8B). В совокупности эти данные говорят не только о независимости процессов дивергенции «городских» популяций по адаптивной и дезадаптивной пластичности, их течении с разной скоростью, но и том что отбор, изменяющий первую и вторую ради приспособлений к «острову тепла». В первом случае это отбор в пользу полезных кодирующих мутаций, делающих пластичность более адаптивной, во втором — против вредных цис-регуляторных генных вариантов, ликвидирующий неадаптивнойсть пластичности.
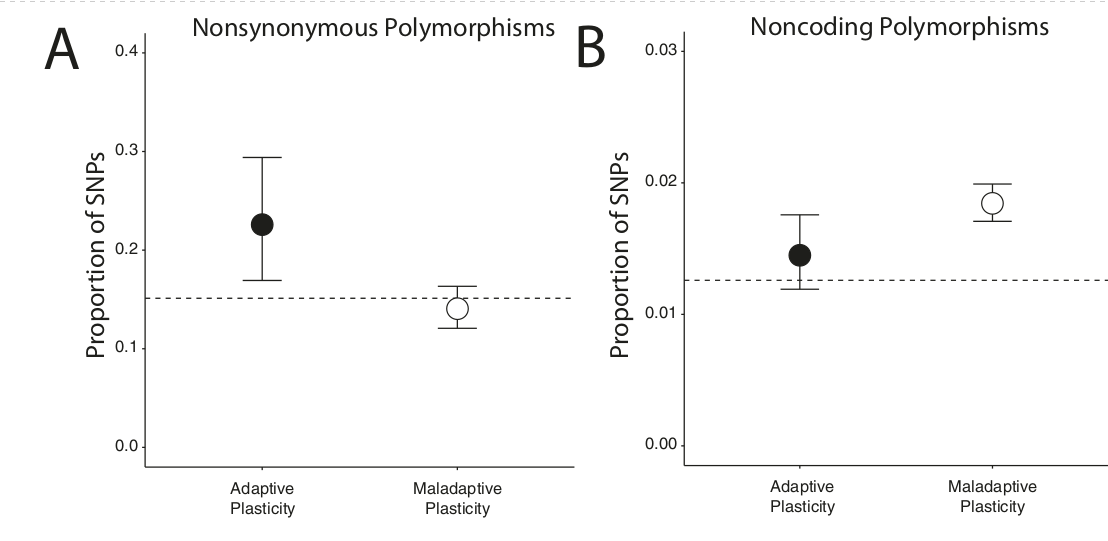 Рис.8. Доказательства различных «мишеней» по лишённого отбора по а) и б). SNP, ассоциированные с б), значимо обогащены несинонимичными полиморфизмами, связанные с а) — нет. B тот же время SNP, связанные с a) значимо обогащены некодирующими полиморфизмами, связанные же с б) нет. График показывает долю обоих типов SNP среди генов, участвующих в формировании адаптивной (черные точки) и неадаптивной (белые) пластичности, «усы» ±95%-ный интервал. Горизонтальный пунктир — доли, ожидаемые на основе «фонового» % (в основном среди генов, не связанных с теплостойкостью). Использованы все 130 особей, для которых были данные о последовательности
Рис.8. Доказательства различных «мишеней» по лишённого отбора по а) и б). SNP, ассоциированные с б), значимо обогащены несинонимичными полиморфизмами, связанные с а) — нет. B тот же время SNP, связанные с a) значимо обогащены некодирующими полиморфизмами, связанные же с б) нет. График показывает долю обоих типов SNP среди генов, участвующих в формировании адаптивной (черные точки) и неадаптивной (белые) пластичности, «усы» ±95%-ный интервал. Горизонтальный пунктир — доли, ожидаемые на основе «фонового» % (в основном среди генов, не связанных с теплостойкостью). Использованы все 130 особей, для которых были данные о последовательности
Обсуждение и ограничения использованных методов
Предшествующая работа высветила сложность интерпретировать по транскриптомным данным связи между исходной пластичностью и [пластичностью] дивергировавшей в ходе микроэволюции: отрицательные корреляции возникают из-за статистического артефакта, регрессии к среднему79. В этой работе предпринято несколько мер, чтобы учесть потенциальное влияние этого явления. Во-первых, регрессия к среднему значению, скорее всего, произойдет, когда самые экстремальные случаи одного сравнения (т.е. гены, дифференциально экспрессируемые в сравниваемых двух группах) повторно оцениваются в последующем сравнении79. Поэтому гены-кандидаты не идентифицировали по дифференциальной экспрессии между двумя состояниями (исходная пластичность vs дивергировавшая популяция), но по силе регуляторной ассоциации с экологически значимым признаком фенотипа (CTmax) с множеством независимых доказательств что именно это мишень отбора при урбанизации вида60. Рандомизация данных по экспрессии также показывает, что величина корреляции между исходной пластичностью и эволюционной созданной дивергенцией значительно выше случайного уровня75,80 (врезка рис.6A).
Важней всего, что такой статистический артефакт возникает из нейтральной вариации регуляторных данных и поэтому не ведет к одному и тому же паттерну генетической дивергенции между «лесными» и «городскими» ящерицами. Параллельно возникшие «подписи» роста генетической дивергенции у 4х независимо возникших городских популяций еще больше подтверждают роль естественного отбора в наблюдаемой эволюции пластичности генной экспрессии.
Основная цель этого исследования — понять эволюционные механизмы, приводящие к эволюционной дивергенции генной экспрессии, обуславливающей наблюдаемую разницу в термостойкости и других зависящих от температуры характеристиках между «городскими» и «лесными» анолисами.
Для этого проверяли несколько независимых, но взаимосвязанных гипотез, чтобы понять эволюцию экспрессии генов, связанную с данным различием, возникающим при урбанизации вида. В центре внимания оказываются паттерны экспрессии генов, связанные с функционированием организма при разных температурах. На их основе отыскивали паттерны изменений в экспрессии, обусловленные урбанизацией, имеющие отношение к освоению новой среды обитания, с другой температурой. Этот анализ на уровнях 1) генной экспрессии генов и 2) генных последовательностей, с отдельными SNP с разных сторон поддерживают значимость вклада изменений эволюционного характера в наблюдаемые различия по термостойкости.
Однако предложенный авторами подход к поиску кандидатных генов, вероятно, лишь часть более сложной картины адаптивной модификации приведшей к «городскому» более термостойкому фенотипу, с большим CTmax. Для более полного понимания последствий более высокого порядка от дивергенции генной экспрессии в сравниваемых парах популяций, зависящей от линий [происхождения ящериц], и механизмов, обеспечивающих данную дивергенцию, необходимо более обширное общее исследование тепловых характеристик организма в целом81. Использование параллельных изменений в нескольких парах «городских» vs «лесных» популяций, контроль условий выращивания и проведения опытов позволяют разделить и отдельно анализировать влияния среды развития (развитийной пластичности) и резких изменений температуры вокруг особи здесь и сейчас (фенотипическая гибкость).
Их приложение для модели функционирования всего организма и его регуляторных основ позволило бы дополнительно контекстуализировать результаты данного исследования и углубило бы понимание важности фенотипической пластичности как средства быстрой адаптации на начальных стадиях дивергенции. Тем не менее, представленные здесь подходы демонстрируют полезность интеграции данных полевых исследований, лабораторных опытов и данных мультиомики чтобы детально описать механизмы, обеспечившие адаптивную дивергенцию между популяциями. Здесь результаты показывают относительную роль адаптивной и дезадаптивной пластичности в стимулировании быстрой эволюции сложных регуляторных систем при урбанизации вида.
Взаимосвязь фенотипической пластичности и эволюции активно дебатируется в эволюционной биологии42,82. Это во многом обусловлено общим отсутствием данных об исходной изменчивости пластических реакций, вызванных изменениями среды, и ограниченной информацией об их адаптивной ценности или последующих влияниях на ход (микро)эволюции перед лицом новых экологических проблем. Среди исследований эволюционной роли пластичности большинство явно фокусировалось на адаптивной пластичности. Хотя эволюция через уменьшение неадаптивнрй пластичности, и предлагалось как распространенная форма адаптации в дикой природе83, ему уделялось относительно мало внимания84. Это исследование подтвердило роль фенотипической пластичности в адаптации к новым условиям, подчеркнув снижение/ обращение вспять неадаптивной пластичности как главную цель селективных давлений в городских «островах тепла». Эти данные в целом согласуются с проясняющейся (сейчас) закономерностью: хотя адаптивная пластичность способствует выживанию и сохранению популяции на ранних стадиях колонизации [новой экстремальной среды] генетические изменения, которые её не усиливают, а минимизируют и/или обращают вспять могут быть более частым предметом отбора или вести к большим изменениям28,85-87.
Лабораторные опыты в рамках этой работы использовали неродственных особей, подвергнутых обработке разной температурой. Отсюда пластичность генной экспрессии связанной с разными режимами выращивания ящериц и их обработки, с которой имели дело авторы, представляет собой популяционный феномен и не позволяет количественно оценить индивидуальные нормы реакции или учесть фоновые генетические вариации среди индивидуумов. Авторы признают это ограничением исследования, в центре которого — разница в термостойкости «городских» vs «лесных» популяций (несмотря на высокую степень обмена генами между ними, отмеченного в каждом из муниципалитетов, популяции остаются генетически дифференцированы).
Поэтому дизайн опытов данной работы действительно позволяет исследовать изменения экспрессии генов, вызванные изменением среды обитания при урбанизации, и лежащие в основе эволюционной дивергенции между популяциями (или местообитаниями). Преобладание в исходных «лесных» популяциях температурно-инлуцррованной дезадаптивной пластичности заставляет считать что на ранних стадиях урбанизации вида давление отбора обеспечивается в основном тепловым стрессом.
Параллельность генетических изменений в 4х разных городах согласуется с данными предыдущей работы60 и показывает что поведенческая терморегуляция в городе недостаточна для защиты этих тропических ящериц от жары, что «включает» отбор на термостойкость88,89. Обогащëнность [генов, оказывающихся под отбором, представителями] тех категорий генной онтологии, что связанны с транскрипцией белков теплового шока и репликацией ДНК, позволяет предположить, что индивидуальные различия в термостойкости «городских» ящериц реализуются через посредство большей или меньшей протеотоксичности, обеспеченной агрегацией неправильно свернутых белков.
В совокупности эта группа генов демонстрирует и масштабные «подписи» полигенного отбора по всему «острову (что согласуется с его термической неоднородностью и специализацией [ящериц] к разным микро- и макроклиматическим местообитаниям), так и повторяющуюся дивергенцию между «городскими» и «лесными» анолисами.
Неадаптивная пластичность, проявляемая лесными родословными, скорей всего, главная мишень естественного отбора в этом последнем контексте. Отбор сокращает долю и обращает вспять28 неадаптивные изменения, вызванные ростом температуры, и вызывает значимый сдвиг [структуры пластичности] в городских популяциях в пользу адаптивной её компоненты. Отбор адаптивной и дезадаптивной пластичности, скорей всего, также осуществляется через разные генетические механизмы — несинонимичные и цис-регуляторные полиморфизмы соответственно.
В основе исследования лежит подход к поиску генов-кандидатов, использующий корреляции между экспрессией генов и CTmax для определения групп регуляторов, важных для термостойкости на уровне всего организма. Хотя многочисленные доказательства, представленные здесь, подтверждают, что эти гены являются прямыми мишенями отбора на термостойкость, необходимы дальнейшие лабораторные исследования для прямой количественной оценки её эволюции на уровне всего организма. Действительно, результаты исследования подчеркивают необходимость одновременных и интегративных исследований адаптивной и неадаптивной пластичности на нескольких уровнях биологической иерархии, чтобы полностью понять эволюцию и адаптацию сложных признаков в новых местообитаниях.
Эта интеграция методов имеет решающее значение для прогнозирования реакции организма на предполагаемое антропогенное изменение среды обитания и изменения климата в будущем.
Рекомендуем прочесть
 «Гены урбанизации» и приспособление к средовому стрессу
«Гены урбанизации» и приспособление к средовому стрессу Экспериментальная эволюция гуппи в речках Тринидада.
Экспериментальная эволюция гуппи в речках Тринидада. Быстрая эволюция холодостойкости у анолисов в южном Техасе
Быстрая эволюция холодостойкости у анолисов в южном Техасе Быстрая эволюция в связи с урбанизацией: примеры
Быстрая эволюция в связи с урбанизацией: примеры Урбанизация и антропогенез: неожиданные параллели
Урбанизация и антропогенез: неожиданные параллели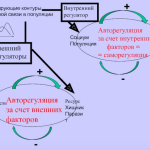 Стресс в раннем онтогенезе как адаптивное явление
Стресс в раннем онтогенезе как адаптивное явление Урбосреда как развивающая: сравнение с одомашниванием
Урбосреда как развивающая: сравнение с одомашниванием Групповые адаптации без группового отбора?
Групповые адаптации без группового отбора?
