

Резюме. Быстрые антропогенные преобразования местообитаний, глобальное потепление и урбанизация, рождают приспособления в виде новых маршрутов миграций и мест зимовок. Для славок-черноголовок Центральной Европы показано, что это ведёт к быстрой эволюции, формирующей не только приспособления (использование кормушек на зимовке, более короткий пролётный путь и более ранний прилёт, большая кладка), но и ведёт к расщеплению исходной популяции. Симпатрически обитающие черноголовки с разных зимовок образуют пары преимущественно между собой, т.е. имет место положительная ассортативность. В дальнейшем показаны генетическая дифференциация и морфологическая дивергенция между разделившимися частями единой популяции, сравнимая с таковой между аллопатрическими популяциями вида и даже превосходящая её. Иными словами, новый режим отбора, созданный климатическими изменениями/урбанизацией, не только вырабатывает приспособления, но и дифференцирует их носителей от основной части популяции, т.е. создаёт новые единицы популяционной системы вида и потенциальную возможность их обособления как отдельных подвидов и далее видов.
Для степного конька показаны начальные стадии этого разделения, до начала генетической и морфологической дифференциации популяций с новых зимовок. На примере пеночек-зарничек, встречающихся в Европе всё чаще, исследован первый шаг этого процесса — как бродячие особи могут стать регулярными мигрантами, как идёт переход от бродяжничества к использованию новых зимовок.
Дихотомическая эволюция славок-черноголовок Центральной Европы
Содержание
- 1 Дихотомическая эволюция славок-черноголовок Центральной Европы
- 2 Генетическая/морфологическая дивергенция между частями разделившейся популяции черноголовок
- 3 Степные коньки сменили места зимовок в тропической Азии на Средиземноморье
- 4 Зарничка «переходит» от бродяжничества к регулярному использованию нового пути миграций
Быстрая эволюция последние 50 лет происходит в германской популяции славки-черноголовки (Sylvia atricapilla). В первой половине ХХ в. вид зимовал в основном на Пиренейском полуострове. C 1959-1961 гг. отметили его зимовку на Британских островах и в Ирландии, где ранее черноголовки только гнездились. Доля зимующих там птиц постоянно росла; по опросу 2004 г., ~31% всех встреч черноголовок в Великобритании приходится на октябрь-март. Также в 1950-1990-х гг. увеличивалась доля птиц в популяциях Центральной Европы, выбравших новый район зимовки на северо-западе, достигнув 7-10% (Berthold et al., 1992; Bearhop et al., 2005).
Благодаря глобальному потеплению буквально на наших глазах возник новый район зимовок и новое направление миграции (северо-западное) в дополнение к прежнему. Работы начала 1990-х гг. показали значительную генетическую подоснову такой новизны (Berthold et al., 1992; Helbig et al., 1994). Черноголовки в неволе в период миграций испытывают так называемое миграционное беспокойство. Они мечутся в определённом секторе круглой клетки (так называемой воронки Эмлена, придуманной для исследования врождённой составляюющей миграционного поведения), как бы пытаясь начать движение, а избранный ими азимут совпадает с направлением от мест гнездования на места зимовки. У зимующих в Португалии это юго-западное направление, у британских – северо-западное. Наследуемость вектора миграционного беспокойства значительна у черноголовок, как и у других видов. Скажем, оно сохраняется у птенцов «британских» или «португальских» птиц, выкормленных вручную в лаборатории, потомство от скрещивания между ними имеет промежуточный азимут (Berthold, Pulido, 1994; Helbig, 1996). Следовательно, новая адаптация — продукт направленных генетических изменений, а не следствие фенотипической пластичности.
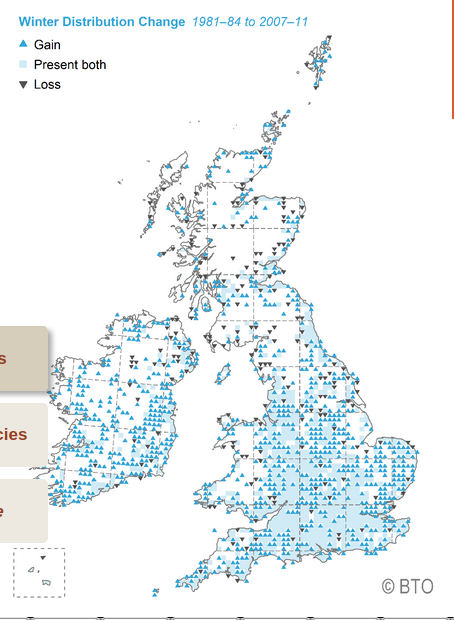
Изменение зимнего распространения черноголовок, 2007-2011 гг. в сравнении с 1981-84 гг. Синий треугольник — появились, где ранее не встречались, голубой квадратик — оба присутствуют, чёрный треугольник вершиной вниз — перестала отмечаться. Источник British Trust of Ornithology
Дальше по содержанию 2Н и 13С в роговых кончиках коготков птиц было показано (Bearchop et al., 2005, fig.1-2), что в центральноевропейской популяции черноголовок произошла дизъюнкция. С северо-западных зимовок птицы в среднем прилетают на 10 дней раньше (вообще, зимующие северней прилетают раньше), делают большие кладки и выводят больше птенцов, чем с юго-западных (Bearhop et al., 2005, fig.4). Выявлена и положительная ассортативность скрещиваний: «британские» и «португальские» птицы образуют пару с себе подобными значимо чаще, чем случайным образом (Bearhop et al., 2005, fig.3). Освоение новой зимовки, вызванное изменением климата, не просто повысило приспособленность, но привело к дизъюнкции, обособляющей этих особей в качестве отдельной популяции (по образцу возникающих при освоении урболандшафта).

Как это было сделано, рассказывает статья Stuart Bearhop et al. (2005) в Science: её перевод следует ниже. Рис.1 статьи показывает область гнездования исследуемой «расщепившейся» популяции, новый и старый район зимовки (эллипс штрих-пунктиром и эллипс сплошными линиями соответственно). Главной проблемой было: когда мы наблюдаем славок на местах гнездования в Германии или Австрии, как определить, где они зимовали, в новом или в старом районе зимовок.
Кольцевание здесь не очень помогает. Поскольку возвращаемость взрослых, а тем более молодых птиц, у мелких воробьиных мала, вследствие значительного % гибели и ещё более значительной дисперсии, уводящей птиц от мест рождения/размножения, особенно молодых.
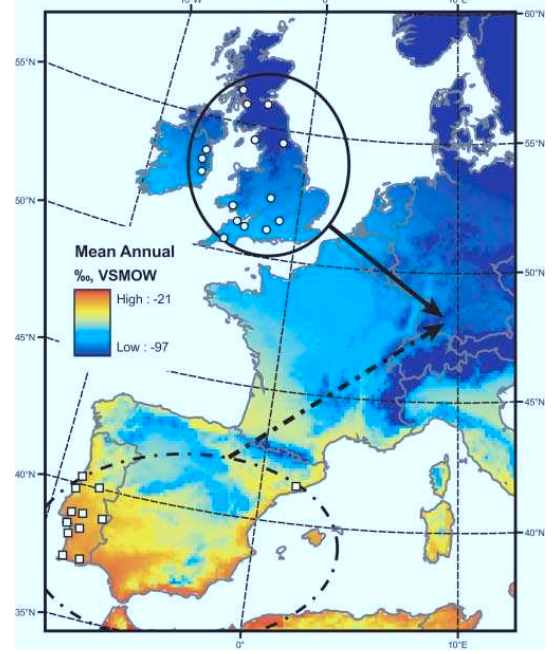
Рис.1. Среднее содержание дейтерия в осадках (синее — меньше, жёлтое и оранжевое — больше) в разных местах Европы, в том числе в старых (обведены шрих-пунктиром) и новых (сплошной линией) местах зимовок славки-черноголовки. Стрелки — направления весенней миграции птиц из первых или вторых в места размножения, где они обитают в симпатрии.
Поэтому для определения «происхождения» птиц из конкретного района зимовки воспользовались изотопным анализом – именно, относительным содержанием стабильных изотопов водорода {измеренного в промилле: δ2Н = (2H/1Hsample ÷ ([2H/1Hreference]-1)x1000} и углерода (δ13C) в метаболически инертных тканях птиц, вроде рогового чехла когтей или клюва. Предполагается, что содержание изотопов в тканях зимующих птиц в целом соответствует их содержанию в осадках, выпадающих в районе зимовки (или в пище), и если содержание этих изотопов в природных ландшафтах имеет хорошо выраженный широтный градиент, то «разницу» в содержании 2Н между Португалией и Ирландией можно использовать для определения места зимовки птиц, занявших территории и размножающихся в Германии или Австрии.
На счастье, это именно так: широтный градиент относительной концентрации дейтерия в осадках в Западной Европе хорошо выражен, и птицы из разных районов зимовок должны разделяться друг от друга с максимальной чёткостью (рис.1 статьи). Если, конечно, концентрация изотопа в тканях точно отражает его концентрацию в природе, без всяких фокусов с разной интенсивностью включения его в ткани у разных видов или в разных популяциях. Чтобы проверить, так ли это, исследовали содержание дейтерия у местных английских/португальских видов птиц, обитающих здесь круглогодично, и сравнивали с зимующими черноголовками. В Англии «из местных» использовали лазоревок Parus caeruleus и больших синиц Parus major, на Пиренеях – средиземноморскую славку Sylvia melanocephala. Соответственно, δ2Н в использованном роговом веществе острия когтя должен а) резко различаться между Англией и Пиренеями и б) в каждый из сезонов быть сходным у зимующих славок и местных видов, даже если он изменяется по сезонам (как и оказалось).
Именно это и было обнаружено при измерениях δ2Н зимой 2001-2002 и 2002-2003 гг. (рис.2 статьи). Относительное содержание дейтерия в промилле в тканях черноголовок обоих районов зимовки: а) значимо не отличается от таковой местных птиц, хотя сильно варьирует от года к году, б) разница в относительном содержании дейтерия между черноголовками из северного и южного района зимовок точно соответствует градиенту, который наблюдается на карте рис.1. У черноголовок, зимующих в Великобритании и Ирландии, δ2Н существенно ниже, чем у зимующих на Пиренеях.
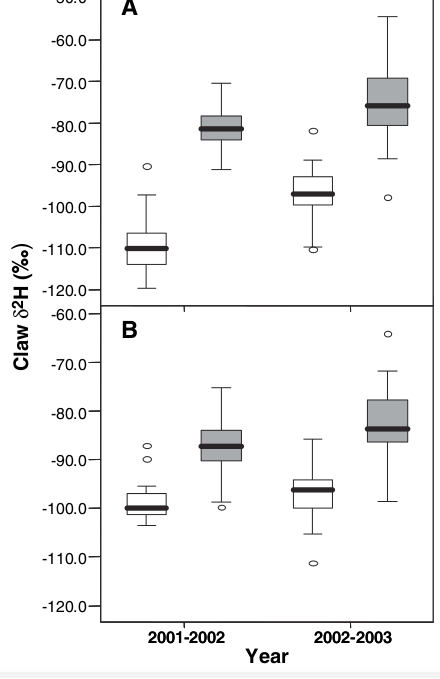
Рис.2. Содержание дейтерия в коготках в разные годы, А — местные птицы, В — зимующие черноголовки, заполненные столбики — Португалия, пустые — Британские острова
Это значит, что, отлавливая птиц в сезон гнездования, мы по относительной концентрации дейтерия можем определить, из какого района зимовок они прибыли, что даёт возможность оценить наличие/отсутствие ассортативности скрещиваний птиц внутри каждой из групп, связанной каждая со «своим» районом зимовки. Это было проделано в Южной Германии и Австрии летом 2002 и 2003 года. Птиц отлавливали так быстро, как возможно, после установления их гнездовых территорий. Старались отловить обоих членов пары одновременно и далее смотрели их последующее размножение. Обнаружена сильная корреляция между δ2Н в коготках самца и самки, образующей с ним пару (рис.3 статьи, обратите внимание на практическое отсутствие особей, промежуточных по δ2Н).
Иными словами, ассортативность есть и она положительная; особи, зимовавшие в Великобритании, и зимовавшие в Ирландии, в Центральной Европе скрещиваются преимущественно с себе подобными. Иными словами, «расщепление» исходной популяции на две произошло и, если новая жизненная стратегия даёт птицам достаточные адаптивные преимущества, разделение будет скорей усиливаться (в чём можно видеть «зародыш» симпатрического видообразования – темы, сейчас более чем популярной).
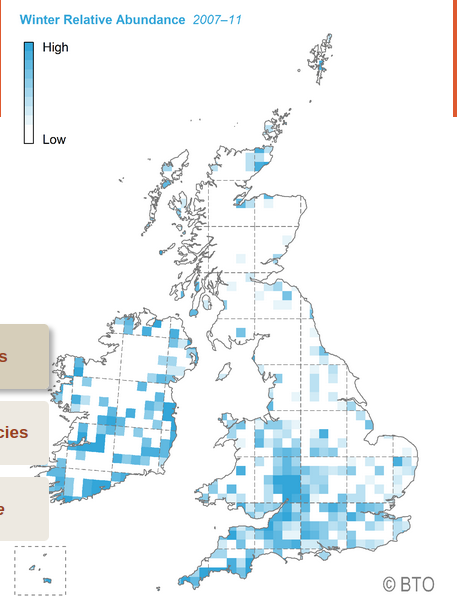
Относительное обилие зимующих черноголовк в 2007-2011 гг., чем синее тем больше
Для более точной оценки степени ассортативности наблюдаемое скрещивание сравнивали с нуль-гипотезой случайного скрещивания, созданной на основе разницы в численности черноголовок в новом и старом районе зимовки, и, соответственно, разной вероятности происхождения немецких птиц из первой или второй. Образование пар из птиц, «происходящих» с общей зимовки, оказывается высоко неслучайным (χ2=40,1, df=1; Р<<0,001): в 2,63 раза более вероятно, чем если бы эти различия были для него существенны. Важно исключительно быстрое появление ассортативности (<30 поколений).
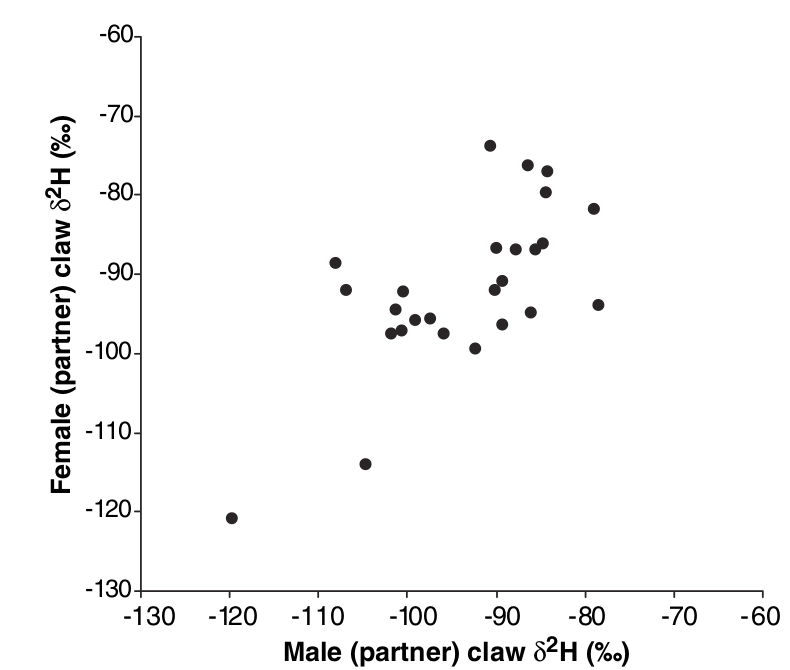
Рис.3. Положительная ассортативность скрещиваний по месту зимовки. Показано содержание дейтерия в коготках самцов и самок, образующих брачные пары в южной Германии
Это вполне совместимо с гипотезой, что при зимовке птиц в более северных широтах критический фотопериод запускает миграционное беспокойство и вызывает ускоренное развитие гонад раньше, птицы раньше прибывают в гнездовой район, что само по себе ведёт к ассортативности скрещиваний. Дальше оно усиливается отбором против потомков «гибридных пар», углубляющим «расщепление» надвое S.atricapilla Центральной Европы. Их генетически детерминированное направление и дальность миграции должны быть какими-то промежуточными, вызывая зимовку где-то в районе Бискайского залива. Такое направление миграционного беспокойства отнюдь не наблюдается в ориентационных экспериментах со взрослыми славками, да и в парах, чьё размножение изучали летом 2002 и 2003 гг., не было птиц, «промежуточных» по месту зимовки (Bearhop et al., 2005).
Далее выяснилась большая адаптивность черноголовок с новых зимовок. У самок, образующих пару с самцами с низким содержанием дейтерия в коготках (=улетающих на северо-запад, Bearhop et al., 2005, fig.1-2), значимо выше размер кладок (использовали множественную логистическую регрессию, Wald = 7.02; P=0.008; влияние даты откладки яиц не значимо, n=48 самцов). Видимо, это следствие качества территории: раньше прибывающие самцы могут занять «лучшие», а самки на это отвечают увеличением кладки. И «лучшие» самки также точнее оценивают территории, быстрей выбирают лучшие, образуя пары с владельцами. Правда, часто не разделить влияние на размер кладки двух «качеств» — территории и самой самки. Тут, видимо, действует правило, хорошо известное орнитологам: репродуктивный успех выше у пар, начинающих кладку раньше сравнительно со средним по популяции (Bearhop et al., 1992).
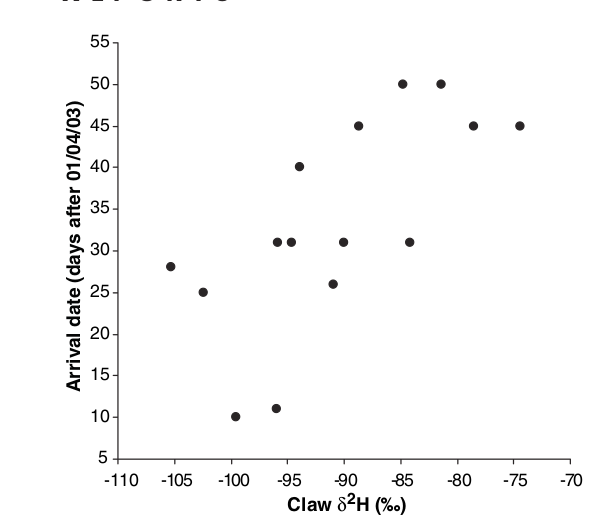
Рис.4. Черноголовки, зимующие северо-восточнее, прилетают раньше. Даты прилёта даны по оси У, в днях относительно «нуля», 1.04.2003, место зимовки определили через содержание дейтерия (абсцисса).
Вслед за большей успешностью размножения самцов с минимумом дейтерия в коготках (т.е. в Великобритании и Ирландии) это было показано и для самок (использовали множественную логистическую регрессию, Wald = 5.08; P=0.015; n=48). Так подтвердилось первоначальное мнение Bertold et al. (1992) о большем «выигрыше» использования новых зимовок. Возможно, выше и «плата», вследствие необходимости полёта над морем и большей суровости зимы сравнительно с Пиренеями (хотя и благоприятность в кормовом отношении значительно выше).
 «Выигрыш» новых зимовок вызван тремя причинами.
«Выигрыш» новых зимовок вызван тремя причинами.
1. Прилетающие с них птицы могут выделить больше ресурсов для размножения, что вместе с занятием лучших территорий усилие обеспечивает лучшую выживаемость птенцов.
2. Современный паттерн урбанизации Великобритании и Ирландии обеспечивает большую кормность зимовки в виде подкормки в садах и выращивания экзотических растений, дающих круглый год ягоды, потребляемые славками. Лучшее питание зимой позитивно влияет и на миграционное состояние.
3. Приспособление к более суровым погодным условиям новой зимовки увеличит выживаемость при более раннем прилёте черноголовок в Центральную Европу (Bearhop et al., 2005).
Эти работы первыми связали дизъюнкцию исходно единой популяции с адаптивными сдвигами в поведении, жёстко контролируемом генетически с положительной ассортативностью скрещивания внутри возникающих частей, показав возможность симпатрического обособления форм внутри неё. Позднейшие — выявили генетическую дифференциацию и морфологические различия между обоими частями разделившейся популяции S.atricapilla, несмотря на симпатрию птиц с «британских» и «пиренейских» зимовок в местах гнездования (Rolshausen et al., 2009).
Источник Science
Генетическая/морфологическая дивергенция между частями разделившейся популяции черноголовок
Позднейшие исследования выявили генетическую дифференциацию и морфологические различия между обоими частями разделившейся популяцию, несмотря на симпатрию птиц с «британских» и «пиренейских» зимовок в местах гнездования (Rolshausen et al., 2009; она пересказана ниже) . Первая выше, чем генетические различия между аллопатрическими популяциями S.atricapilla, разделёнными дистанцией в 800 км, и обуславливает значительность вторых, относящихся к приспособительным признакам. Улетающие в северо-западном направлении отличаются более округлыми крыльями и более узким клювом. Возможно, последнее — следствие использования кормушек в городах Великобритании. Удлинение и утончение клюва (за столь же быстрые сроки, с 1970-х гг.) отмечено также у «городских» больших синиц в Великобритании. Оно отсутствует у нидерландских больших синиц, где конструкция кормушек иная, клюв не надо просовывать через сетку). Они отличаются также цветом клюва и опережения от улетающих в юго-западном направлении.
Ограничение потока генов, созданное разницей фенодат и лучшим состоянием птиц, прилетающих с «северо-западных» зимовок, ускоряет дивергенцию между субпопуляциями в Центральной Европе, по механизму, показанному С.П.Харитоновым — одновременное предъявление альтернативных вариантов отбираемого признака существам, выступающим агентом отбора (или «подставление» их под природный процесс, делающий то же самое), существенно увеличивает коэфффциент селекции, ускоряет выработку приспособительных изменений, т.е. потенцирует его действие. (В наблюдениях Сергея Павловича это были средние поморники Stercorarius parasiticus, разоряющие гнёзда зимняков Buteo lagopus в условиях ослабления охраны гнезда, созданной нехваткой леммингов, и появившийся маскировочный вариант окраски яиц). Похожее расщепление может происходить у более чем 50 видов птиц, недавно изменивших миграционное поведение (чаще всего под влиянием изменений климата. Это работа 2010 г., сейчас таких случаев уже много больше).
Сегодня всё чаще чаще признаётся, что экологические и эволюционные изменения происходят в одном временном масштабе и влияют друг на друга [а не независимы, как раньше считалось]12, 13, 14. Хороший пример — современная эволюция славки-черноголовки Sylvia atricapilla: у популяций Центральной Европы в 1960е гг. возник новый миграционный путь (северо-западный, NW) и новое место зимовки (Великобритания и Ирландия), что «расщепило» их надвое. Успешному освоению новых зимовок благоприятствовало потепления климата и лучшие кормовые условия зимой, созданные использованием кормушек, массовых в Соединенном Королевстве 8, 9. Новые районы зимовки находятся в 1200-1800 км северней западносредиземноморских зимовок, традиционных для другой части популяции S.atricapilla южной Германии и Австрии (летящих на юго-запад, SW, и симпатричных с теми, кто летит в Великобританию) 8, 9. Менее чем за 30 поколений доля NW среди черноголовок, гнездящихся в южной Германии, достигло примерно 10% [15]. Скрещивание тех и других выявило промежуточное направление миграции у гибридов NW и SW; схема наследования согласуется с контролем миграционного направления (непосредственно видимого по азимуту миграционного беспокойства при помещений птиц в круглые клетки) только одним или несколькими основными генами [15].
Дальнейшие эксперименты показали, что миграционное поведение черноголовок может эволюционировать за нескольких поколений благодаря существенной аддитивной генетической вариабельности составляющих его характеристик [16] [включая отбор на оседлость и большую перелётность среди частично мигрирующих средиземноморских черноголовок — за 6 поколений. Прим.публикатора].
По прибытии на места гнездования славки образуют пары на один сезон, NW прилетают раньше SW. Сегрегация по срокам прилёта и занятия территории обеспечивает положительную ассортативность скрещиваний между птицами с одних и тех же мест зимовок. Черноголовки образуют сезонные парные связи по прибытии на места размножения. Различия в миграционных путях ведут к сильному различию фенодат прилёта и, следовательно, к ассортативному спариванию между птицами, прибывшими с одних и тех же зимовок. 8, 17. Таким образом, дивергенция по направлению миграций может заблаговременно изолировать возникающие экотипы NW и SW и, возможно, представляет собой зародыш экологического видообразования у черноголовки.
Доказательств того, что различия в миграционном поведении способствуют видообразованию, у птиц очень немного. Предыдущие [до 2009 г.] исследования популяций птиц, чётко различных по направлению миграций, показали незначительную генетическую дифференциацию, причём по нейтральным маркерам 18, 19. Хотя есть некоторые возможные исключения 20, 21, симпатрическая дифференциация [популяций, тем более подвидов и видов] у птиц считается редкостью 22, 23. Учитывая, что перелетные птицы — яркий пример высокомобильных организмов, отсутствие генетической диверсификации может быть вызвано скрещиванием между различными популяциями. Кроме того, это может отражать ограниченность генотипических изменений в популяциях, испытывающих отбор по одному и тому же экологическому измерению [23]. В этой авторы, однако, исследуют адаптивную дивергенцию в симпатрической популяции славки-черноголовки южной Германии. Они использовали анализ стабильных изотопов, чтобы связать разных особей этой популяции с разными местами зимовки, и тем самым показать ещё «расщепление» надвое, а также связать это диверегенцией по признакам фенотипа и нейтральной генетической дифференциацией. Так они попытались определить, идёт ли в такой популяции дизруптивный отбор, и по каким экологическим измерениям он идёт, одному или нескольким.
В соответствии с низким уровнем генетической структурированности у перелетных птиц, предшествующие анализы гаплотипов митохондриальной ДНК (мтДНК) не выявили генетической структированности у черноголовок Западной Европы, т.е. что любые различия в миграционном поведении очень недавние [24].
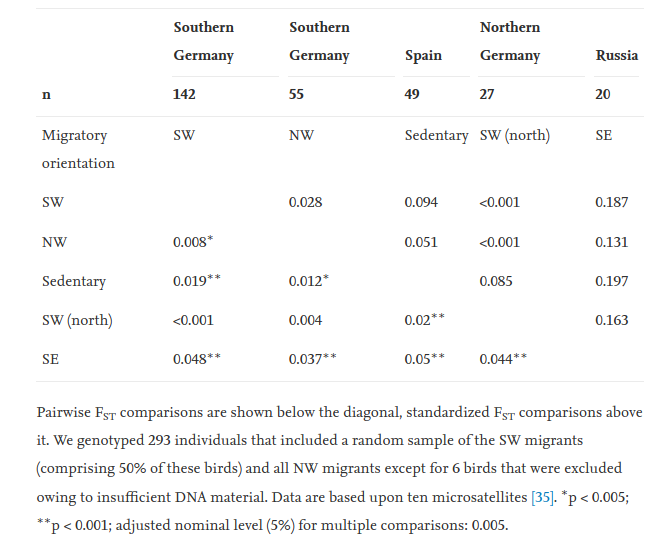
Табл.1 статьи
Появление NW птиц вызвало разделение популяций Центральной Европы по нейтральным маркерам. В табл.1 приведены данные Fst между разными популяциями и между птицами одной популяции, использующими альтернативные пути миграций, NW и SW, также как оседлыми из Испании. Обратите внимание на их сравнимость (данные основаны на 10 микросателлитах), причём дифференциация возникла за <30 поколений. Генотип особей предсказателен: с вероятностью >85% он верно относит птицу к конкретной популяции (рис.1) и [25].
Важно отметить, что низкая, но значительная генетическая дивергенция между NW и SW в условиях симпатрии (Fst = 0,008; p < 0,005) выше дивергенции между аллопатрическими популяциями черноголовок из Северной и Южной Германии, разделенных 800 км (она к тому же незначима Fst = 0,0001–0,004; все p > 0,5; табл.1). Генетическое расхождение между NW и SW в условиях симпатрии последовательно обнаруживалось в разные годы и в разных местах сбора проб (таблица S1). Интересно, что генетическая дистанция между NW и SW выше в данных каждого отдельного года, нежели в объединённой выборке разных лет. Этот результат показывает рост генетической дивергенции их внутри симпатрической популяции.
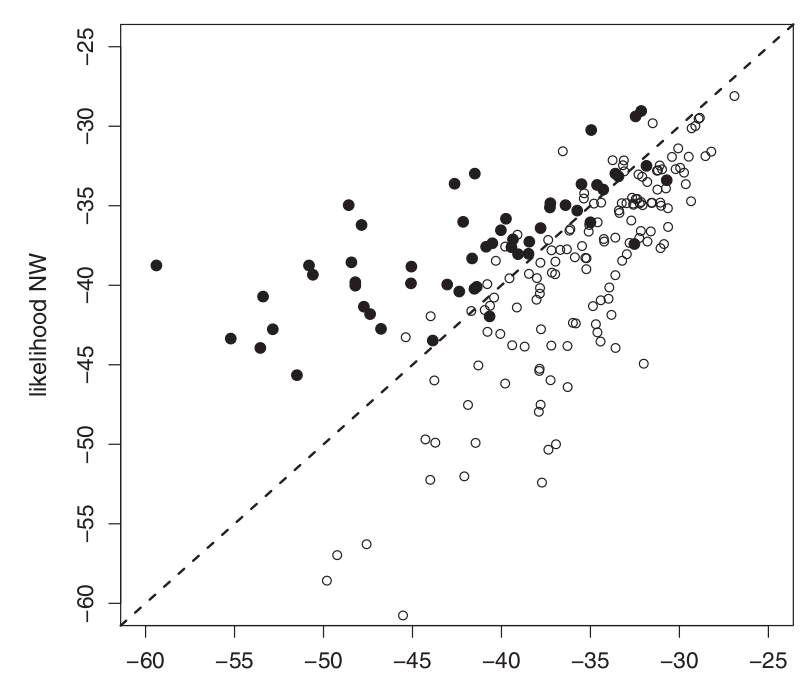
Рис.1. Логарифмы вероятности отнесения черноголовок из симпатрической популяции к одному из альтернативных мест зимовок на основании генотипа.
Авторы использовали многолокусную идентификацию происхождения отдельных особей без какой-либо априорной информации, кроме использующей изотопную «подпись». Черные круги (•) — прилетевшие с северо-западных зимовок (NW); белые круги (○) с юго-западных (SW). Пунктирная линия показывает соответствие 1:1. Обратите внимание, что две популяции генетически схожи, поскольку большая доля особей группируется близко к линии соответствия 1:1.
Эти данные заставляют считать, что благодаря позитивной ассортативности скрещиваний появление нового места зимовки/пути миграций инициировало репродуктивную изоляцию в симпатрических популяциях черноголовки. Имеющаяся генетическая дивергенция вряд ли может быть создана дрейфом генов, он не успеет за <30 поколений (или даже <60 поколений, если NW появился 100 лет назад, но остался незамеченным, из-за того что первоначально в нём участвовало мало особей). Более вероятна связь генетической дивергенции с эффектом основателя, поскольку доля гомозигот среди NW выше, чем у SW (G.R., неопубликованные данные). [Это аргумент в пользу недавнего появления — у этого и других видов певчих птиц новые пути миграций возникают благодаря относительно небольшому числу бродяг, способных успешно перезимовывать в новых местах и возвращаться в места гнездования. На наших глазах такой путь сейчас формируется у пеночки-зарнички Phylloscopus inornatus, о чем ниже. Прим.публикатора]
Дисперсионный анализ генетической изменчивости (AMOVA) пяти популяций черноголовок Европы, включающих разные миграционные стратегии (рис.2), направленный на определение их генетической структурированности, выявил низкие, но значимые уровни последней (общий Fst = 0,016; 95% доверительный интервал = 0,005–0,028; p = 0,00228). Попарные сравнения Fst выявили самую сильную генетическую дифференциацию между мигрантами на юго-восток Европы (SE) из Рыбачьего, Россия, и другими популяциями (Fst = 0,037–0,050; все p < 0,00001).
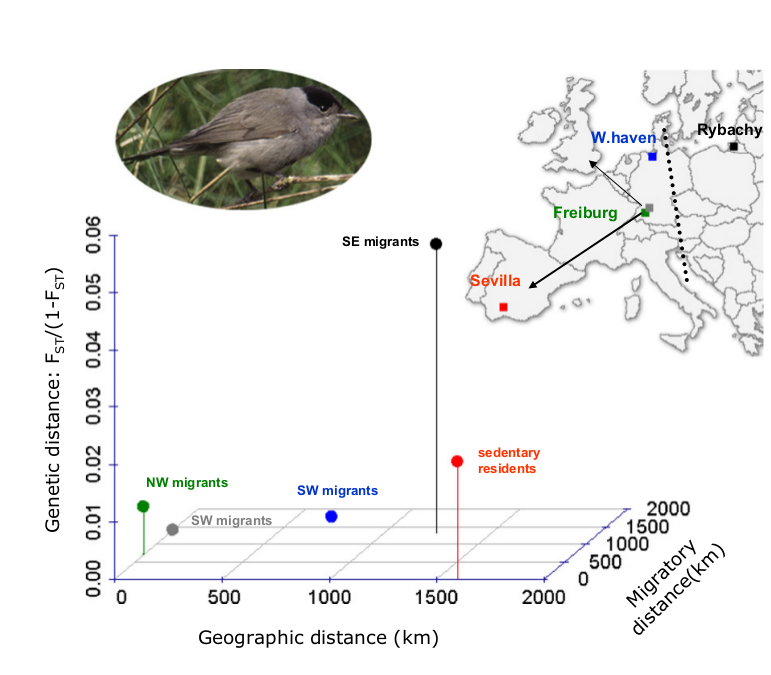
Рис.2. Генетические дистанции между популяциями черноголовок больше зависят от направлений миграционных путей [их членов], а не от географического расстояния между ними.
Генетические и географические дистанции показаны относительно SW-черноголовок из южной Германии (серая точка), дальность миграций показана на оси z, разные цвета — разные географические популяции. На карте Европы показаны места их размножения и два ярко выраженных миграционных водораздела в черной шапке. Пунктир — водораздел между направлениями миграции SW и SE, проходящий по 12°E–13°E [24]. Стрелки — недавно возникшие альтернативные пути миграций, SE vs NW. “W. Hafen” -Вильгельмсхафен.
Эта дифференциация отражает зону вторичного контакта особей, происходивших из разных плейстоценовых рефугиумов, и использующих разные миграционные пути, SW и SE. Граница между ними проходит по 12°–13° восточной широты в Центральной Европе [24]. Аналогично оседлые черноголовки из южной Испании сильно дифференцированы от трех мигрирующих центральноевропейских популяций (Fst = 0,012–0,020; все р ≤ 0,005). Однако в этих последних альтернативность миграционных путей, NW или SW, связана с большей генетической дивергенцией, чем «изоляция расстоянием» между аллопатрическими популяциями (тест Мантеля на генетическую и географическую дистанцию: r = 0,504, p = 0,255).
Помимо хорошо известных примеров интродуцированных видов и популяций, это одно из немногих исследований, доказывающее, что экологические изменения, связанные с деятельностью человека, достаточно сильны, чтобы инициировать становление репродуктивной изоляции внутри симпатрических популяций. Альтернативная гипотеза об аллопатрическом происхождении черноголовок NW не подтверждается ни данными авторов, ни предыдущими данными по генетике других европейских популяций черноголовки [24]. Эта гипотеза едва состоятельна, ибо мигрирующие на северо-запад черноголовки до 1960-х годов не отмечались в Европе или где-либо еще.
Фенотипическая дивергенция
Авторы обнаружили морфологическую дифференциацию между черноголовками NW и SW. Новый миграционный путь NW короче (≈1090 км), почему крылья более округлые, чем у летящих по более длинному SW (≈1640 км). Наблюдаемое различие в форме крыла внутри симпатрической популяции хорошо согласуется с закономерностью, установленной для разных популяций черноголовок Европы: удлинение миграционных маршрутов увеличивает остроконечность крыльев (индекс Холински, рис. 3). Эти различия вряд ли могли быть вызваны эффектом основателя, ибо тогда мигранты с северо-запада происходили бы из более южной популяции с её более коротким миграционным путём (что значительно удлинило бы NW), а это не подтверждается данными кольцевания. Птицы NW и SW равно линяют в местах гнездования в своих обычных местообитаниях перед миграцией [27]. Хотя линька не обязательно случается в одно и то же время, маловероятно, что различия в форме крыла между NW и SW связаны с разными средовыми условиями во время линьки. Куда более вероятен эффект начинающейся репродуктивной изоляции между этими группами птиц и отбора на путях миграций, связанного с их разной длиной. я мигрантов СЗ и ЮЗ способствовала адаптивному фенотипическому расхождению в направлении нового миграционного маршрута.
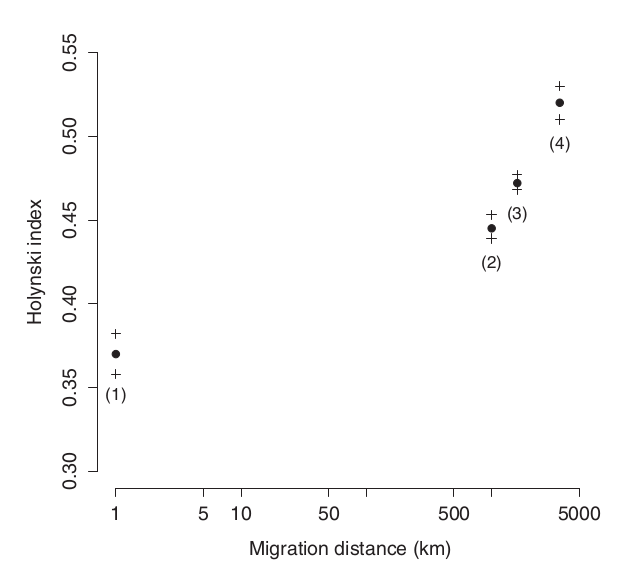
Рис.3. Индекс Холински меняется в соответствии с дальностью миграционного пути
Крыло мигрантов на короткие расстояния (<500 км) или малоподвижных средиземноморских черноголовок ([1]; n = 99) (дополнительные данные из [38]) более закруглённое, чем у более дальних мигрантов из других популяций (на 1800-5000 км, [4]; n = 315, среднеквадратичная ошибка по Холинскому = 0,52 ± 0,009). Черноголовки NW ([2]; n = 61) имеют более округлые крылья, чем симпатричные с ними SW у которых миграционный путь длиннее ([3]; n = 264; F = 6,23, p < 0,05, дисперсионный анализ типа II; см. рисунок S2 в дополнительных материалах для распределения индекса Холински среди двух симпатрических популяций). У южногерманских черноголовок наследуемость признаков крыла >0,6 [38]. Обратите внимание на логарифмический масштаб оси x. Показаны средние значения и стандартные ошибки.
Птицы NW и SW также различны по морфологии клюва, но не по длине цевки (р > 0,4). Форма клюва этого вида варьирует между популяциями и полами (анализ главных компонент, PC2: F = 12,32, p < 0,001). Пол влияет на размер клюва (PC1: F = 10,05, p < 0,01; мигранты nNW = 61, мигранты nSW = 264, дисперсионный анализ типа II): в обеих популяциях клювы самок крупнее, чем у самцов. Различия в форме клюва могут быть вызваны дифференцированным использованием пищевых ресурсов [28], сценарий, вероятно справедливый и для черноголовки. На зимовках в средиземноморском регионе 95% их рациона составляют ягоды [29], тогда как на зимовках в Великобритании они питаются в основном семенами и жиром в садовых кормушках [30]. Учитывая, что ширина зева является основным ограничением, определяющим размер плодов, которые может использовать птица [31], мы предполагаем, что относительно более широкие клювы SW адаптированы к более специализированной, плодоядной диете. Напротив, относительно более узкие и длинные клювы NW, вероятно, отражают более генерализованные способы кормления у кормушек для птиц [и необходимость просовывать клюв в сетку, как это показано для удлинения клюва у английских больших синиц].
Цвет клюва и оперения также существенно различается между NW и SW (в пространстве цветов оперения, вообще присущем данному виду, т.е. речь идёт о большем и меньшем выражении окрасок, уже имеющихся, не о возникновении новых цветовых паттернов). Клюв и перья спины NW относительно более коричневые (с оливковым оттенком у самцов), у SW они относительно более серые независимо от пола или возраста (таблица S2). Это говорит о разном соотношении эу- и феомеланина в обоих частях разделившейся популяции, что может объясняться различными стратегиями линьки. Разница в цвете может быть связана с разным износом перьев: черноголовки, зимующие северней, меньше меняют перьев в предбрачной линьке, чем зимующие южнее [27]. В отличие от пера, клюв представляет собой быстро реагирующую ткань, чувствительную к изменениям уровня средового стресса, состояния здоровья и рациона питания [32]. Поэтому разница цвета клюва отражает разную напряжённость существования и разные виды корма на разных зимовках, NW и SW.
В общей сложности пять признаков, не коррелирующих между собой (морфология крыла, форма клюва, цвет оперения на спине, цвет оперения на голове и цвет клюва; все р > 0,2; см. таблицу S2) оказались различны у разных частей симпатрической популяции. Эти отличия достаточно очевидным образом связаны с разными проблемами приспособления, решаемыми NW и SW: форма крыльев с дальностью миграционных путей, форма клюва связана с кормовыми объектами и/или методами, окраска оперения со стратегиями линьки или синтезом меланина.
Поскольку отбор и наследуемость пяти признаков, вероятно, различаются, различные эволюционные механизмы, вероятно, способствуют фенотипическому расхождению среди симпатрических черноголовок. Во-первых, репродуктивная изоляция может способствовать адаптивной фенотипической дивергенции в ответ на контрастирующие режимы отбора, созданные на каждом из миграционных маршрутов. В этом случае дивергенция является адаптивной и эволюционировала вследствие расхождения миграционных путей. Мы предполагаем, что этот сценарий особенно вероятен для различий в форме крыла и клюва. Во-вторых, фенотипические различия могли быть вызваны дрейфом после зарождающейся репродуктивной изоляции. В-третьих, эффекты основателя могут объяснить фенотипические различия, которые присутствовали бы до — и, возможно, даже способствовали — эволюции расхождения миграционных путей. Поскольку адаптивная роль окраски клюва и спины неизвестна, сочетание эффектов дрейфа и основателя может объяснить различия в этих признаках.
Даже если предположить появление дивергенции [по этим 5 признакам] после установления дихотомии миграционных путей, темпы микроэволюции (измеряемые в холдейнах) будут высоки (хотя и не исключительны). Степень дивергенции NW и SW по форме крыла (hp (1,43) = 0,014) и клюва (hp (1,43) = 0,020) оказываются на верхнем пределе 3го квартиля выборки наблюдаемых скоростей эволюции (выборка соответствующих исследований обобщена Hendry et al. ([33]). По базе данных оттуда квартили скоростей эволюции составляют (в холдейнах) первый: 0,002; второй: 0,006; третий: 0,014. Если новый миграционный путь возник не 30 (как следует из первых встреч черноголовок в Великобритании), а 60 лет назад, то темпы микроэволюции будут ещё ниже (таблица S3), но все же выше медианы, представленной в [33].
Неизвестно, приведут ли контрастирующие режимы отбора к более глубокому расхождению между NW и SW. Авторы отмечают, однако, что возникшие различия в окраске могут запустить механизм усиления, связанный с лучшим распознаванием «своих» и более активным их предпочтением, при ещё большем отвержении «чужих». Если ещё у гибридов снижена приспособленность, т.к. их миграционный маршрут промежуточен, и они оказываются в неподходящих местах для зимовки, положительная ассортативность будет усиливаться, а расхождение NW и SW углубляться, т.е. обособление экотипов оказывается побочным следствием появления новых зимовок. Авторы считают, что последнее инициирует репродуктивную изоляцию в условиях симпатрии, что ведёт к появлению генетически различных экотипов (NW и SW) и их последующей эволюции, всё более независимой. Мы предполагаем, что экотипы испытывают различные режимы отбора, которые пока что действенны лишь на зимовках (т.е. в условиях аллопатрии), но их последствия сильно влияют на поведение особей в гнездовой период, когда оба экотипа симпатричны в Центральной Европе. В этом эволюционном сценарии одно изменение — новые места зимовок и направления миграций — изначально ограничивает поток генов между частями ранее единой популяции, и «подставляет» каждую из них под свой собственный режим отбора. Генетическая и фенотипическая дивергенция, вызванная различием этих давлений, вторично приводит к дальнейшему сокращению потока генов и — благодаря петлям положительной обратной связи между сокращением потока генов и адаптивной дивергенцией — делает правдоподобным сценарий зарождающегося экологического видообразования. [Подобное расхождение уже было показано, правда не на птицах, Прим.публикатора]
Теоретические соображения предсказывают, что если контрастирующие режимы отбора ведут к дивергенции внутри ранее единой популяции, вызванные им изменения [элементарные эволюционные явления] могут быть более эффективны в стимулировании видообразования, чем ответ на отбор по одному признаку. Поскольку начало использования черноголовками нового миграционного пути «подставляет» их под отбор другими факторами среды, ведущими к изменению других признаков, некоррелированных с данным, различие между отбором по одному признаку и нескольким может быть менее выраженным, чем недавно предполагалось [7].
Наше исследование иллюстрирует глубокое влияние быстрых антропогенных изменений среды обитания на эволюционную траекторию популяций. Зимняя подкормка птиц в Соединенном Королевстве породила северо-западный миграционный маршрут, который ранее, вероятно, был бы неадаптивным. Поскольку антропогенные изменения, как правило, происходят одновременно во многих экологических измерениях, они ведут к разнообразному отбору по множеству признаков и тем самым стимулируют современную эволюцию.
Мы предполагаем, что эволюция мигрирующих экотипов — это тонкий, но потенциально широко распространенный процесс, совершенствующий приспособления к разнообразным экологическим условиям, с которыми сталкиваются мигранты. Мы прогнозируем, что она происходит у видов с низкой связанностью между популяциями, когда особи расходятся по разным местам зимовки или местообитаниям.
Экспериментальные процедуры
Птиц отлавливали в 4х регионах. Весной 2006-2008 гг. мы ловили черноголовок по прибытии в места их размножения на юге Германии в Радольфцелле (47°45′ с.ш., 08°59′ в.д.) и Фрайбурге (48°00′ с.ш., 07°51′ в.д.). Выборка отловленных особей включала мигрантов с юга (в основном с юга Испании) и с северо-запада (зимовавших в Великобритании), которые были пойманы на каждом участке площадью около 50 гектаров в одни и те же паутинные сети. Также были взяты образцы от: 1) оседлых черноголовок, пойманных зимой 2006-2008 гг. недалеко от Севильи, Испания (37°39′ с.ш., 5°35′ з.д.); 2) северогерманских черноголовок, пойманных летом 2007 г. в местах их размножения в Вильгельмсхафене (53°31’с.ш., 08°07’в.д.); 3) черноголовок, пойманных весной/летом 2007 года в Рыбачьем, Россия (55°09’с.ш., 20°51’в.д.). Птицы из Вильгельмсхафена мигрировали над морем, направляясь в Средиземноморье и дальше в Африку к югу от Сахары по юго-западному пролётному пути, из Рыбачьего — мигрируют в Африку по юго-восточному маршруту в Африку [24]. Мы подсчитали количество поколений, разделяющих симпатрические популяции, разделив время с момента появления нового пути миграций (отмеченного в 1959 году) на среднеожидаемую продолжительность жизни южногерманских черноголовок [27].
Изучены следующие фенотипические признаки птиц южной Германии и Севильи: длины предплюсны и крыла; длины всех длинных первичных маховых и первых вторичных маховых (P1–P9, S1); длина, ширина и высота клюва; окраска оперения перьев спины, груди и головы; окраска клюва. Вариации в морфологии клюва анализировали с помощью анализа главных компонент (PCA) по измерениям клюва. Первую компоненту PC1 интерпретировали как размер клюва (объясняет 50% изменчивости; нагрузки отдельных измерений по этой оси: длины клюва: -0,31, ширины: -0,64, высоты: -0,69). Вторую PC2 — как форму клюва (33% изменчивости; нагрузки: длина: -0,91, ширина: 0.39, высота: 0,04). Кроме того, мы получили 2-4 дистальных среза когтей от каждого индивидуума для изотопного анализа, позволяющего определить место происхождения (см. рисунок S3).
В оценке окраски оперения и клюва использовали спектрометр AvaSpec 2048 и дейтериевую галогенную лампу AvaLight-DH-S (Avantes). выполнены 5 измерений для каждой части тела птицы, спектры усреднены в интервалах 5 нм от 300 до 700 нм. Используя модель птичьего глаза [34], мы смоделировали вероятность улавливания фотонов четырьмя типами птичьих колбочек, используемых для цветового зрения.
Генетический анализ
ДНК извлекали из образцов крови с помощью набора для анализа крови и тканей DNeasy (QIAGEN). Характеристики локусов, амплификация локусов и данные о полиморфизмах таковы, как описано в [35] и таблице S4. Мы провели расчеты попарного Fst, разделения молекулярной дисперсии (AMOVA) и точности характеристики популяции по индивидуальному генотипу популяции. Чтобы учесть вариации гетерозиготности в популяциях, различающихся по степени изолированности [36], мы использовали стандартизированную меру дифференциации, масштабирующую Fst в один и тот же диапазон (0-1) для всех уровней вариации между локусами и популяциями [37]. Мы сообщаем о стандартизированном показателе вместе с исходными значениями Fst. Более того, мы проверили надежность нашего набора из десяти локусов, поскольку наши оценки могут быть предвзятыми из-за низких размеров выборки относительно количества аллелей (таблица S5).
Источник Current Biology
Степные коньки сменили места зимовок в тропической Азии на Средиземноморье
Степные коньки, мелкие певчие птицы с равнин Сибири, Казахстана, Монголии и Китая, нашли новое место для зимовки. Обычно эти пернатые проводят холодное время года в тропической Азии, однако в последние десятилетия их все чаще встречают зимой в Юго-Западной Европе и Северной Африке. Как отмечается в статье для журнала Current Biology, скорее всего, сдвиг в миграционном поведении коньков произошел из-за антропогенных климатических изменений: благодаря росту температур запад Средиземноморья стал идеально подходить для зимовки этих птиц.
Перелетные птицы мигрируют по одним и тем же маршрутам в течение тысяч лет. Тем не менее, в случае необходимости они способны довольно быстро менять их. Например, при расширении ареала пернатые иногда выбирают более удобный путь к местам зимовки и обратно. А в некоторых случаях они и вовсе отказываются от перелетов или начинают проводить холодное время года в совершенно новых регионах. Так, славки-черноголовки (Sylvia atricapilla) из континентальной Европы, которые раньше мигрировали в Средиземноморье и Африку, в последние десятилетия все чаще зимуют в Великобритании и Ирландии, где благодаря климатическим изменениям и обилию кормушек для них сформировались идеальные условия.

Степной конёк на зимовке в Таиланде
Степной конек (Anthus richardi) — еще один вид, миграционное поведение которого, возможно, меняется прямо у нас на глазах. Эти певчие птицы, родственные трясогузкам, гнездятся на травянистых равнинах и горных склонах от Южной Сибири и Казахстана до Восточного Китая. На зиму они улетают в Индию и Юго-Восточную Азию. При этом отдельные птицы движутся в противоположном направлении и достигают Европы, где их каждую осень в небольшом количестве замечают орнитологи и бердвотчеры.
Традиционно считалось, что в Европу прилетают молодые степные коньки, которые сбились с маршрута из-за неопытности и никогда не смогут вернуться к местам размножения. Однако в последние десятилетия таких птиц становится все больше. Например, во Франции в 1981-1993 годах каждый год регистрировали от трех до шести степных коньков, а к 2018 году эта цифра выросла до 144. Подобный рост можно было бы объяснить тем, что европейцы стали активнее искать и лучше распознавать редких пернатых гостей. Но частота встреч с другими заблудившимися птицами сибирского происхождения увеличилась далеко не так значительно, как в случае степных коньков. Более того, степных коньков начали замечать в Европе в зимний период. Первый такой случай был зафиксирован в 1991 году на юге Франции. С тех пор этих птиц регулярно встречают зимой во Франции, Италии, Португалии и Испании.
Часть орнитологов считают, что у степных коньков появилось новое место зимовки в Западном Средиземноморье — и новый миграционный маршрут. Согласно альтернативной гипотезе, в последние годы молодые особи этого вида просто чаще сбиваются с пути и вместо тропической Азии по ошибке оказываются в Европе. Разобраться в данном вопросе решила команда исследователей во главе с Полем Дюфуром (Paul Dufour) из Университета Савойя Монблан. Они предположили, что если степные коньки действительно начали зимовать в Западной Европе, то среди птиц, которые сюда прилетают, должны быть особи разных возрастов. Напротив, если коньков заносит сюда случайно, подавляющее большинство из них будут молодыми особями.
Чтобы проверить эти идеи, Дюфур и его коллеги в течение трех зим отлавливали на средиземноморском побережье Франции и Испании степных коньков и оценивали их возраст. В общей сложности в их руки попала 81 птица. Взрослыми оказались 28 из них, что соответствует 34,6%. Аналогичные доли взрослых особей характерны для воробьинообразных, которые регулярно зимуют в Средиземноморье, например, для зарянок (Erithacus rubecula). Таким образом, результаты отловов подтверждают гипотезу о новом месте зимовок. Кроме того, в течение двух первых зим орнитологи пометили индивидуальными кольцами и выпустили 68 коньков. Одиннадцать из них вернулись в Средиземноморье в следующие зимы.
Исследователи допускали, что птицы, которых они ловили на средиземноморском побережье, принадлежат к ранее неизвестной популяции, обитающей к западу от традиционных мест размножения вида. Чтобы исключить эту возможность, Дюфур с соавторами пометили геолокаторами семь особей, пойманных зимой 2019-2020 годов. Трех из них удалось повторно поймать следующей зимой и вернуть устройства. Проанализировав собранные данные, ученые пришли к выводу, что все три конька провели лето в западной части известного гнездового ареала вида, а именно в Новосибирской области. По крайней мере один из них участвовал в размножении. Это дополнительный аргумент в пользу идеи о новом миграционном маршруте.
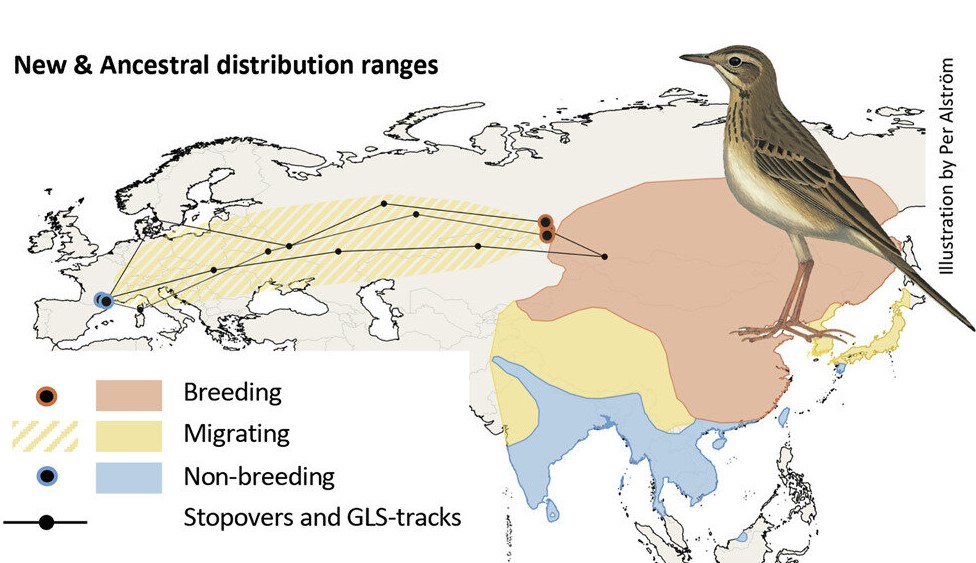
Миграции степных коньков (Anthus richardi). Красным отмечены места размножения вида, синим — традиционные места зимовок в тропической Азии, желтым — регионы, где коньки встречаются во время миграций. Точки с синей обводкой отмечают места отлова птиц на побережье Средиземного моря, а точки с оранжевой обводкой — места размножения трех птиц, помеченных геолокаторами. Также на карте изображены их миграционные пути.
На последнем этапе авторы решили удостовериться, что их выводы верны и для других регионов Европы и Северной Африки, где степных коньков замечают вне сезона размножения. Для этого они оценили возраст 331 особи с фотографий, размещенных орнитологами и бердвотчерами в онлайн-базах данных. Оказалось, что в Северной Европе взрослые особи среди степных коньков встречаются редко: их доля здесь достигает всего 10% осенью и 15,2% зимой. Таким образом, в этот регион в основном прилетают бродячие молодые особи, которые сбились с пути. Напротив, в Юго-Западной Европе и Северной Африке взрослых коньков отмечают намного чаще. Их доля на изученных снимках составила 48,5% осенью и 59% зимой. Таким образом, взрослые особи регулярно зимуют на Пиренейском полуострове, во Франции, Италии и Марокко.
Судя по всему, степные коньки начали зимовать в Юго-Западной Европе и Северной Африке совсем недавно (либо отдельные особи зимовали здесь регулярно, но в последнее время их доля многократно увеличилась). Толчком к этому, вероятно, послужили антропогенные изменения климата. Сравнив температурный режим в Западном Средиземноморье в 1961-1990 и 1990-2018 годах, Дюфур с соавторами обнаружили, что за последние десятилетия зимы в данном регионе стали куда мягче, в результате чего заметно расширилась площадь территорий, подходящих для зимовок степных коньков. В первую очередь это касается севера Испании, юга и запада Франции, а также Италии.
Если раньше молодые коньки, которые случайно прилетали сюда осенью, погибали (что обычно происходит со сбившимися с пути перелетными птицами), то несколько десятилетий назад они начали успешно перезимовывать на новом месте и возвращаться к местам гнездования. Затем они передавали свои гены потомству, в результате чего численность особей, зимующих в Средиземноморье, постепенно росла. Впрочем, следует отметить, что подавляющее большинство степных коньков, как и прежде, проводят зиму в тропической Азии. Возможно, европейские зимовки используют только птицы с крайнего запада гнездового ареала.
Сергей Коленов для N+1
Зарничка «переходит» от бродяжничества к регулярному использованию нового пути миграций

Зарничка

Рис.1. Гнездовой ареал зарнички (розовый), пунктир — наиболее западная часть ареала по Уралу, жёлтое — миграционные пути, в Южную Азию даны по литературе, в Европу — гипотетические. Голубой — исторически сложившиеся зимовки вида, зелёные точки — регулярные встречи птиц в октябре и ноябре 2018. Большинство встреч — в Западной Европе и Юго-Восточной Азии, за некоторыми исключениями на Ближнем Востоке и в Центральной Азии. Обратите внимание, что частота зеленых точек не отражает частоту встречаемости вида в правильной пропорции из-за смещения наблюдателя/регистратора. На фото — зарничка, встреченная на острове Ouessant, Франция, осенью 2020 года
Быстрый рост численности зарничек в Западной Европе объясним двумя причинами. Во-первых, большинство птиц, отмеченных вдали от обычного пути миграций, это бродяги, т.е. дезориентированные молодые особи, которые движутся в “неправильном” направлении во время своей первой миграции [27, 28] [и, в конечном счёте, обречены]. Во-вторых, вид, возможно, «прокладывает» новый миграционный путь в совершенно ином направлении (рис. 1). Второе было предложено Gilroy & Lees [29] в контексте общего увеличения численности сибирских видов птиц в Европе (их гипотеза о “псевдо-бродяжничестве”). Оба варианта не взаимоисключающие: их разграничение сложно, поскольку “бродяжничеству” не хватает концептуализированного определения, и поэтому его трудно отделить от “миграции” (см. [30]) [плюс одно может перерастать в другое]. Хуже того, для каждой отдельной особи не определишь, мигрант она или бродяга [если не проследить её перемещения в будущем — сможет ли успешно перезимовать, полетит ли весной в места размножения и пр., скажем при помощи геолокаторов. Прим.публикатора].
Независимо от относительного вклада миграции и бродяжничества в статус зарнички Европе, данный вид — идеальная модель для лучшего понимания эволюционных основ изменений миграционных путей и мест зимовок у видов, где время первых и место вторых контролируется генетически ([20, 21, 31]). Даже если зарнички в Европе — всё ещё просто бродяги (т.е. среди них нет особей, регулярно зимующих там и возвращающихся обратно в гнездовой ареал — скажем, в самые западные его части), значительное число птиц, участвующих в этих перемещениях, и их ежегодное повторение дают уникальную возможность изучить механизмы подобного. На самом деле было бы удивительно видеть, как каждую осень тысячи птиц увлекаются в эволюционный тупик. И наоборот, если на наших глазах возникает новый — западный — миграционный путь, зарничку можно будет включить в очень небольшое число изученных орнитологами случаев быстрого изменения миграционных путей у видов, где они детерминированы генетически. В их числе евразийская славка-черноголовка Sylvia atricapilla: [20]; североамериканская ласточка-касатка Hirundo rustica: [32]; белолобая скалистая ласточка Petrochelidon pyrrhonota: [33]; возможно, степной конёк: [8]; см. также [34,35,36] для других примеров изменения миграционных маршрутов).
Во-вторых, бродяжничество может быть полностью обусловлено «иной», «ошибочной» обработкой сигналов ориентации в ходе самой миграции или изменением развития систем, обеспечивающих их обработку в ходе онтогенеза. В обоих случаях это необратимо влияет на миграционный фенотип без каких-либо изменений на уровне ДНК (т.е. это фенотипическая пластичность [46, 79]). Зеркальное отображение обычного пути миграций или обратный ему маршрут обычно объясняются неправильным использованием птицей компасов врождённой природы: птицы либо не смогли определить точку отсчета на компасе (т.е. север; путь, обратный обычному: [28]), либо не смогли «взять» правильный угол относительно контрольной линии север–юг (зеркально отображённый путь: [65]). Thorup [27] предположил, что YBW, встречающиеся в Европе (и, вероятно, происходящие из наиболее западных гнездовых популяций), возможно, движимы сбоем, «отразившим» естественное направление миграционного пути на 180°, и следуют по маршруту, противоположному исходному «большому кругу» маршрутов миграции предков [80] (обратите внимание, что зеркальное отображение данной траектории также может объяснить появление зарнички в Южной Европе [10]). Глубинные причины различного выражения ориентации по врожденному компасу могут быть связаны с онтогенетическими проблемами [81] и интеграцией разных компасов [82] без генетических модификаций.
Эти два объяснения не являются взаимоисключающими. Бродяги могут быть смесью генетически изменённых птиц и птиц, чья система ориентации изменена (точнее искажена определённым образом) негенетическими механизмами, ведущими к увеличению вариабельности [направлений миграции] и [главное] новым миграционным путям [46, 83].
Временные и пространственные закономерности появления зарничек в Европе
За последние несколько десятилетий сильно выросло число зарничек, отмеченных в разных странах Европы (рис. 2); большинство встреч были после 1985 года [84], причём в ряде публикаций дополнительно подчеркнута исключительно высокая численность птиц в так называемые годы «притока» [85,86,87]. Даты первых наблюдений варьируются от 1845 (Германия [88]); 1937 (Норвегия [89]) до 1987 (Болгария [90]); 1988 (Польша [91]) и 2013 (Румыния [84]). В нескольких странах (особенно в Великобритании) число встреч настолько возросло, зарничек стали считать обычными мигрантами, а не редко залётными или бродячими [84, 88]. В 1963 г. её исключили из списка Британского комитета по редким птицам, а после 2018 года — и из «Отчета о [встречах] редких видов», поскольку за предыдущие три года было минимум 1950 ежегодных встреч. Важно отметить, что в ряде районов рост числа встреч был независим от увеличения числа наблюдателей, и именно там (о. Фэйр, Шотландия, о.Гельголанд, ФРГ), где давно были созданых орнитологические станции, ведшие долговременный мониторинг перелётных птиц. Это гарантировало, что рост встреч зарничек реален, это не артефакт интенсификации наблюдений (рис. 2).
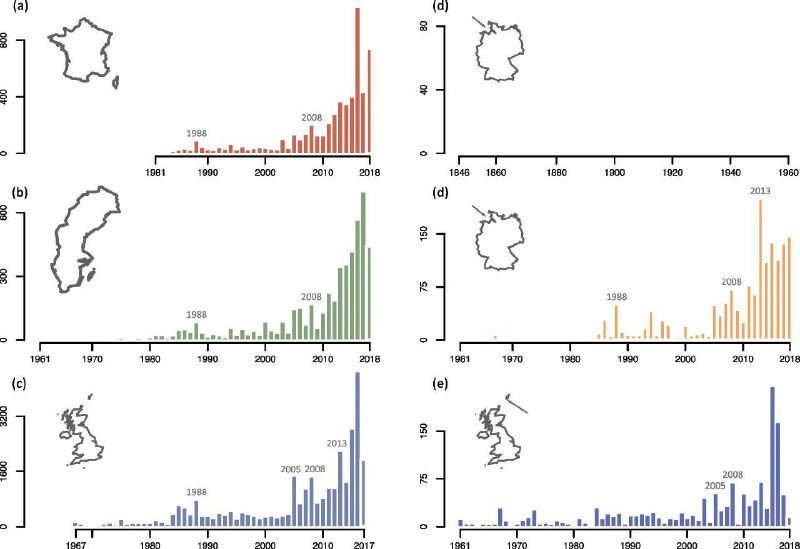
Рис. 2. Общее число зарничек, отмеченных осенью в а. Франции, b. Швеции, c. Великобритании, и на двух орнитологических станциях, где пролётных птиц учитывают издавна и очень точно, большими рыбачинскими ловушками, на о. Гельголанд, ФРГ (даны 2 панели, чтобы показать наблюдения с 1846 г.), и на о. Фэйр в Шотландии

Это «расписание» встреч зарничек в Европе, по-видимому, указывает общее направление движения птиц из Сибири на запад-юго-запад по всей Европе [10], при некоторых региональных различих. Птицы, перемещающиеся с востока на запад, отловлены Thorup [27] при кольцевании на о. Christiansø (Кристансё) в Дании (рис. 3). Зарничка, пойманная на Гельголанде осенью 2013 года, повторно отмечена на Лансароте в январе 2014 года, т.е. мигрировала на юго-запад [85] (рис. 3). Tonkin и Gonzalez-Perea [26] сообщили о повторных поимках конкретной окольцованной особи в Андалусии, Испания, в течение двух последовательных зим. Тем самым подтверждается, что хотя бы отдельные зарнички определяют место зимовки с высокой точностью и придерживаются его в разные годы. Хотя несколько повторных наблюдений [зарничек на зимовках] дают лишь отдельные свидетельства об экологии передвижения этого вида, они заполняют важные пробелы в наших знаниях о характере его пребывания в Европе.
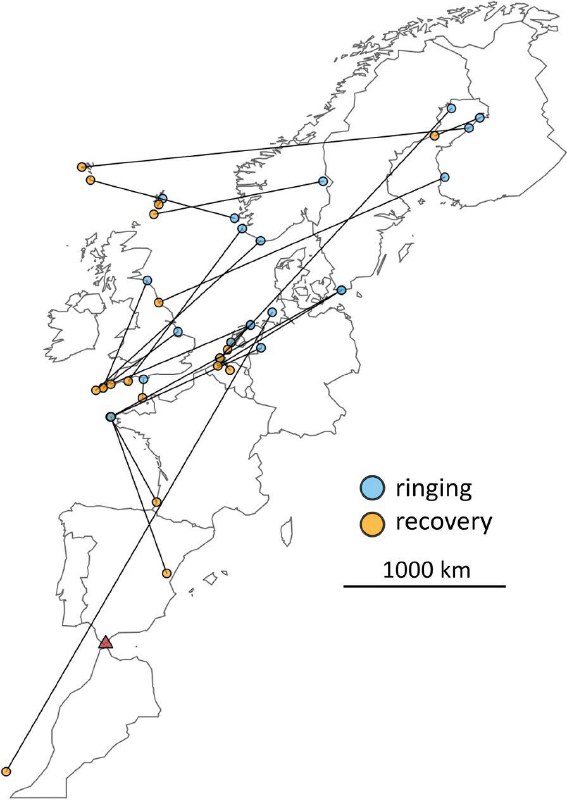
Рис. 3. Перемещения 22 зарничек между местами кольцевания (синий) и местами поимки той же осенью в Европе (оранжевый)
Дорожная карта по выявлению соотношения вкладов бродяжничества и миграции [в европейские встречи зарничек]
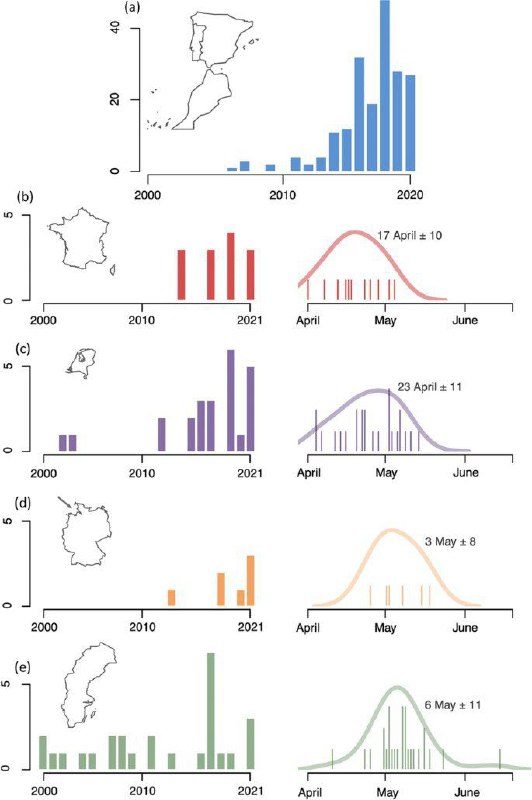
Рис. 4. Рост общего числа зимних регистраций зарничек в Испании и Марокко (а, декабрь-февраль) , на фоне роста числа их весенних регистраций во Франции (b), в Нидерландах (с), на Гельголанде d и в Швеции e, в сравнении с распределением сроков этих находок (правая панель). Учитывая, что зимние встречи из года в год идут в одних и тех же местах, в радиусе до 5 км, естественно предположить, что, успешно перезимовав, они весной движутся к местам гнездования. Цифры следует интерпретировать с осторожностью из-за изменения в интенсивности наблюдений и возможной предвзятости отчётности в базах данных гражданской науки.
Отталкиваясь от этой информации и идей табл.1, понимаешь необходимость большего числа доказательств для оценки эволюционного статуса зарничек в Европе, также как соотношения вкладов бродяжничества и миграции. Действительное обнаружение нового пути миграций требует показать, что часть зарничек, отмеченных в Европе осенью, успешно зимует, успешно размножается в видовом ареале, и их потомство также мигрирует в Европу, прежде чем вернуться для размножения (или что многие особи, гнездящиеся в данном районе внутри видового ареала — скажем, в наиболее западной его части — перезимовали в Европе, см. табл.1). Далее следует подтвердить то, что в 2000-х гг. выявлено для черноголовки: птицы, использующие новый миграционный маршрут, образуют самоподдерживающуюся популяцию, не зависящую полностью от притока бродяг в осенний период, а поддерживаемую собственным размножением. Такого точности фактических данных, по-видимому, трудно достичь в ближайшем будущем, но мы можем получить о том же некоторое представление благодаря другим подходам, изложенным в дорожной карте в табл.1.
Перспективы в изучении миграции птиц и заключение
Выводы
Статья представляет пеночку-зарничку моделью для понимания механизмов бродяжничества и его потенциальной значимости в эволюционном становлении новых миграционных путей у птиц. Хотя бродяжничество уже давно привлекает как исследователей, так и непрофессионалов, некоторые вопросы обсуждались в литературе и интерпретировались по-разному. Мы желаем, чтобы настоящий обзор стимулировал дискуссию о бродяжничестве и о том, действительно ли бродяги могут выступать в качестве первопроходцев в открытии новых миграционных маршрутов.
Мы также хотели бы подчеркнуть, что понимание распространения зарнички в Европе будет зависеть от сотрудничества многочисленных исследовательских групп, орнитологов, орнитологов, кольцевых станций по всей Европе, Северной Африке и Азии. Мы надеемся, что в рамках этого проекта возникнет сильная сеть, в будущем способная оказать помощь для запуска других аналогчных проектов.
Источник в Movement Ecology
Примечания
1«Лучшие» или «худшие» особи здесь — с «точки зрения» естественного отбора, т. е. более или менее приспособленные относительно тех изменений среды, чьё воздействие изучается в данной работе и в месте её проведения. Почти всегда в этих исследованиях фигурирует «качество» особи, «оцениваемое» отбором, для чего связывается с разными индивидуальными характеристиками: физическим состоянием, жирностью, уровнем тестостерона и пр. Последние, однако, предполагаются исследователем, догадки которого отнюдь не всегда счастливые. Даже при значимости связи нужно показать, что в её основе — причинность, а не корреляция, и остаётся возможность, что «качество» связано с другими параметрами индивидуальности, не учтёнными в данной работе. Так или иначе, «качество» особи — удобный технический термин, важно не «онтологизировать» его без обсуждения.
Рекомендуем прочесть
 Направления эволюции социальной организации животных
Направления эволюции социальной организации животных Острова в море возможностей: что определяет темпы и направления эволюции каротиноидной окраски у птиц?
Острова в море возможностей: что определяет темпы и направления эволюции каротиноидной окраски у птиц? Дивергентно-конвергентные механизмы в эволюции биоразнообразия
Дивергентно-конвергентные механизмы в эволюции биоразнообразия Теории биологической эволюции с позиций современного развития системной биологии
Теории биологической эволюции с позиций современного развития системной биологии «Гены урбанизации» и приспособление к средовому стрессу
«Гены урбанизации» и приспособление к средовому стрессу Быстрая эволюция в связи с урбанизацией: примеры
Быстрая эволюция в связи с урбанизацией: примеры Мини-рецензия на книгу Менно Схилтхёйзена «Дарвин в городе. Как эволюция продолжается в городских джунглях»
Мини-рецензия на книгу Менно Схилтхёйзена «Дарвин в городе. Как эволюция продолжается в городских джунглях» Групповые адаптации без группового отбора?
Групповые адаптации без группового отбора?
