

Белоспинный дятел осматривает место кормления
Свойства древесины и выдалбливание гнездовых дупел
В продолжение темы дятлового долбления
C.А.Фетисов
Данная статья – вторая в серии моих работ о гнездовых дуплах дятлов (Фетисов 2017). В ней изложены материалы, показывающие каким образом дятлы выдалбливают свои гнездовые дупла в деревьях, с какими проблемами они при этом сталкиваются и как решают эти проблемы, а также почему большинство дупел устраивается, как мы знаем, в стволах деревьев с «мягкой» древесиной или стволах фаутных деревьев.
Возможности дятлов при долблении древесины
Содержание
Давно известно, что дятлы – весьма специализированная группа лесных птиц, хорошо приспособленная к лазанию по деревьям и долблению древесины. В частности, благодаря адаптированному для долбления клюву они способны не только успешно кормиться на стволах и ветвях деревьев, но и выдалбливать гнездовые дупла в древесине. Хотя, конечно, не все виды дятлов имеют равные возможности и навыки к долблению, потому что среди них встречаются птицы разной величины и их клювы обладают, соответственно, разной мощностью (ср.,например, желну, или чёрного дятла Dryocopus martius и малого пёстрого дятла Dendrocopos minor).
Показателен в связи с этим размер щепок, который остаётся на местах кормёжки разных видов дятлов. Так, желна долбит во время кормёжки даже твёрдую смолистую древесину живых елей Picea sp. (рис. 1а) и сосен Pinus sp. При этом он не только долбит клювом, но также пользуется им как рычагом, отламывая крепкие щепки сырой древесины толщиной до 2-3 мм и длиной 10-20 см (Формозов 1974). Щепа, выброшенная желной после долбления дупла, бывает несколько мельче «кормовой»: её размеры от 3 до 11 см в длину и от 1.0 до 1.8 см в ширину. Ещё меньше, длиной 2-4 см, строительные» щепки можно найти под дуплами большого пёстрого дятла Dendrocopos major (Иноземцев 1986), а у малого пёстрого дятла они не превышают в длину 1 см и в некоторых случаях напоминают крупные опилки (Птушенко, Иноземцев 1968).
Некоторые виды дятлов кормятся почти исключительно путём долбления (рис. 1а-г) и поэтому освоили эту технику в совершенстве, в то время как у других видов дятлов основные способы добывания корма совсем не связаны с долблением. Не случайно В.И.Осмоловская и А.Н.Формозов (2009) разделили всех дятлов на 2 группы. К первой они отнесли «долбящих» дятлов, для которых долбление служит основным способом добывания корма – желну, трёхпалого Picoides tridactylus и белоспинного дятлов Dendrocopos leucotos, а ко второй группе – «малодолбящих», в число которых включили, кроме большого пёстрого, ещё седого Picus canus, зелёного Picus viridis, малого и среднего Dendrocopos medius пёстрых дятлов.

Рис. 1. Возможности дятлов при долблении древесины. а – кормовые ниши желны Dryocopus martius в живой ели; б – кормовые ниши желны в усыхающей чёрной ольхе; в – ствол чёрной ольхи после кормёжки белоспинного дятла Dendrocopos leucotos; г – щепа ольхи на месте кормёжки белоспинного дятла; д — щепа осины под дуплом большого пёстрого дятла Dendrocopos major (щепа слева – при долблении дупла снаружи, щепа справа – при долблении дупла внутри). Фото автора.
 Чтобы долбить древесину, человек издавна использует специальный инструмент – долото или заточенную стальную пластину для выдалбливания в древесине различных отверстий, пазов и тому подобных пустот.
Чтобы долбить древесину, человек издавна использует специальный инструмент – долото или заточенную стальную пластину для выдалбливания в древесине различных отверстий, пазов и тому подобных пустот.
Пользуясь долотом, человек может как расщеплять, так и перерубать древесные волокна – в зависимости от того, что требуется согласно заданной им программе обработки древесины. С подобной же целью – долбления древесины (в первую очередь, для добывания живущих в ней насекомых) – дятлы начали использовать клюв. Это привело к тому, что почти все они имеют теперь прямые долотообразные, твёрдые, с продольными гранями клювы, которые дятлы и вгоняют, как долото, в древесину сильными и точными движениями мышц головы и шеи (рис. 2)1.
Основной набор приёмов долбления клювом у дятлов таков, что большинство из них эффективно используют клюв лишь для расщепления или размочаливания древесных волокон (с последующим отрывом их клювом), а также для откалывания или отламывания краевых древесных волокон, но не перерубания их клювом.

Рис. 2. Белоспинный дятел Dendrocopos leucotos, долбящий ствол живой ивы. Национальный парк «Себежский», апрель 2013 года. Фото автора.
Сравнение клюва дятла с долотом основано, вероятно, на некоторых похожих движениях, производимых этими двумя «инструментами», а главное, на сходных задачах и результатах их применения, хотя, конечно, возможности клюва, особенно в подрубании древесных волокон и расщеплении их с торца, а не вдоль волокон, несравненно более ограничены, чем у стального долота. Так, К.Н.Благосклонов (1972) даже утверждает, что дятлы не могут раздолбить древесину, если её волокна расположены горизонтально. На самом деле они, конечно, способны раздолбить и горизонтально расположенные стволы и ветки, но только при условии, что у них будет возможность расположить своё тело параллельно тем сукам или стволам деревьев, которые они облюбовали для долбления (рис. 3).
Как считалось ранее, толстые кости черепа дятлов амортизируют такие удары. По последним данным (Zhao et al. 2014), почти вся энергия от ударных нагрузок распределяется по телу дятла, и только 0.3% энергии удара клювом достаётся на долю мозга.

Рис. 3. Белоспинный дятел Dendrocopos leucotos, долбящий горизонтальный сук ивы. Для долбления горизонтального сука дятел меняет вертикальное положение тела на горизонтальное и начинает долбить древесину вдоль растительных волокон. Национальный парк «Себежский», февраль 2014 года. Фото автора.
Однако при долблении древесины всегда приходится считаться и с тем, что представляет собой сама древесина: в частности, какой древесной породе она принадлежит и в каком состоянии на момент долбления находится. Ведь совсем не случайно на обработку разных сортов древесины человек тратит разное по продолжительности время и прикладывает разные усилия. По той же причине дятлы никогда не выдалбливают дупла во всех деревьях подряд, а выбирают для долбления только те деревья, которые им под силу. В этом не трудно убедиться путём анализа многочисленных литературных данных, касающихся мест расположения гнездовых дупел разных видов дятлов. Так, в зависимости от географического положения района, типа и возраста леса и других условий состав древесных пород с дуплами дятлов может заметно изменяться, но в целом все виды дятлов предпочитают выдалбливать гнездовые дупла в определённых породах деревьев с «мягкой» древесиной, часто уже достигших фаутного состояния (Гладков 1951; Севастьянов 1962; Благосклонов 1968; Митяй 2009; и мн. др.).
Давно замечено, например, что если снаружи дятлы могут долбить почти любую древесину, то внутри дупла – только мягкую и гнилую (Благосклонов 1952); к тому же щепа, отколотая дятлом внутри дупла, в среднем заметно меньше по размерам, чем щепа, отколотая при долблении дупла снаружи (рис. 1д). Помимо того, при выборе деревьев длгнездования дятлы предпочитают обычно осину Populus tremula – возможно потому, что её ударная твёрдость значительно меньше, чем у берёзы Betula sp., ели и сосны, хотя торцевая твёрдость осины при статических нагрузках оказывается даже выше указанных пород (Зимин 1968). В связи с этим нельзя не затронуть и других свойств древесины разных пород деревьев, так или иначе способствующих или, наоборот, препятствующих долблению дятлов и даже сводящих на нет их возможности выдолбить себе гнездовое дупло.
Свойства древесины в местах выдалбливания дятлами гнездовых дупел
В данном разделе я не буду подробно останавливаться на разных свойствах древесины, хотя эта тема очень хорошо разработана ещё в 1930-1950-е годы в статьях многих отечественных ботаников и специалистов по древесиноведению: А.Т.Вакина, С.И.Ванина, В.Е.Вихрова, Л.А.Иванова, В.П.Крайнева, А.И.Кузнецова, В.Е.Москалёвой и др. Многие сведения обобщены в книге В.Ф.Раздорского (1955) «Архитектоника растений» и других сводках. Здесь для нас важны два момента, важные с точки зрения оценки, во-первых, возможности и процесса образования в деревьях естественных дупел и, во-вторых, податливости разных сортов древесины (с её неодинаковой твёрдостью и другими свойствами у разных видов деревьев) для долбления её дятлами.
В обыденной жизни и технике древесиной называют обычно внутреннюю часть дерева, лежащую под корой. Ей присущи многие механические, физические и химические свойства (прочность, плотность, влагопроводность, теплопроводность и др.), но наиболее существенным для нашего рассмотрения является твёрдость или, иначе говоря, способность древесины сопротивляться внедрению в неё более твёрдого тела (в нашем случае клюва дятла). В зависимости от твёрдости древесины все виды деревьев принято делить на 3 группы: виды с мягкой древесиной (ель, ива Salix sp., конский каштан Aesculus sp., липа Tilia sp., ольха Alnus sp., осина, пихта Abies sp., сосна, тополь Populus sp.); виды с твёрдой древесиной (берёза, бук Fagus sp., вяз Ulmus sp., груша Pyrus sp., дуб Quercus sp., клён Acer sp., лиственница Larix sp., рябина Sorbus sp., яблоня Malus sp., ясень Fraxinus sp.); виды с очень твёрдой древесиной (белая акация Robinia pseudoacacia, граб Carpinus betulus, самшит Buxus sp.).
Твёрдость древесины неодинакова даже в разных частях одного и того же дерева: например, она выше в прикомлевой части любого ствола, а также в сучках и наростах, зачастую (например, в капах) со свилеватым строением древесины. Сучки – это основания ветвей, заключённые в древесине ствола дерева. По форме их делят на круглые, овальные и продолговатые; по степени срастания с окружающей древесиной – на сросшиеся, частично сросшиеся, несросшиеся и выпадающие; по состоянию древесины самих сучков – на здоровые, загнившие и гнилые. Здоровые сучки обычно в 2-3 раза твёрже окружающей их древесины, а полностью сгнившие сучки превращаются в бурую, легко растирающуюся в порошок массу, напоминающую нюхательный табак (т.н. табачные сучки). Наряду с сучками, свилеватость древесины, характеризующаяся нарушением параллельного расположения волокон дерева по отношению к продольной оси его ствола, и некоторые другие её пороки (наклон волокон, завиток, крень) могут существенно препятствовать выдалбливанию дятлами гнездовых дупел и даже вынуждать их бросать уже начатые ими дупла и ограничивать тем самым выбор мест, благоприятных для постройки дупел.
В связи с этим – как во время кормёжки на деревьях путём долбления древесины, так и во время выдалбливания дупел – все виды дятлов на практике приобретают конкретный опыт определения твёрдости (и, соответственно, доступности) древесины – и на разных участках ствола и ветвей одного дерева и у разных видов деревьев, свойственных тому географическому региону, в котором они находятся.
Отчего же зависит твёрдость древесины? В живых деревьях она появляется в основном в результате одревеснения клеточных оболочек после пропитки их лигнином (Раздорский 1955). При этом древесина хвойных пород деревьев содержит 27-30% лигнина и даже до 34% у сосны, а древесина лиственных пород – лишь 18-27%. Помимо того, в древесине всех деревьев имеется своя «арматура», которую можно сравнить по типу со структурой железобетона. У хвойных пород деревьев функции «арматуры» выполняют такие структуры, как трахеиды, поэтому древесина у них сравнительно однородна, хотя и у хвойных в центре стволов паренхима более мацерирована, т.к. снаружи находятся более молодые и живые клетки (Чавчавадзе 1979). У лиственных же пород подобные функции выполняют древесные волокна (т.н. либриформ), и древесина их более дифференцирована. Обычно древесные волокна занимают от 1/3 до 2/3 объёма древесины, и от этого напрямую зависит её твёрдость. У пород с «мягкой» древесиной этот объём минимален, потому что стенки древесных волокон у них заметно тоньше. А у некоторых видов (например, ив и тополей) древесина становится несколько мягче ещё и с возрастом самих деревьев.
Однако какой бы твёрдостью ни обладала древесина любого дерева, рано или поздно (из-за старения, после механических или биологических повреждений, в результате заболеваний деревьев и т.д.) она утрачивает все свои свойства, включая твёрдость. Чаще всего древесина теряет их после воздействия на неё дереворазрушающих грибов и бактерий, проникающих в неё через трещины, ходы насекомых и другие пороки ствола и ветвей (рис. 4).
Паразитирующие на деревьях грибы, в первую очередь широко известные трутовики, далеко не «всеядны». Каждый из них, как и любой другой живой организм, существует только в определённых для него условиях влажности, температуры и даже освещённости. Из-за этого они выбирают, как правило, однотипные породы деревьев: одни – лиственные (ложный трутовик Phellinus igniarius, берёзовая губка Piptoporus betulinus, дубовая губка Daedalea quercina), другие – хвойные (еловая Phellinus chrysoloma и сосновая Ph. pini губки и др.), а узкоспециализированные виды поселяются вообще только на определённых видах растений, как, например, ложный осиновый трутовик Phellinus tremulae, паразитирующий лишь на осине2.
В отличие от последнего, ложный трутовик Phellinus igniarius развивается на живых деревьях, отмерших стволах и пнях многих видов лиственных деревьев. Он имеет несколько специализированных форм, поражающих осину, берёзу, ольху и другие лиственные породы, причём в насаждениях всех возрастов, но по мере старения древостоев поражённость ложным трутовиком увеличивается иногда до 100%.

Рис. 4. Механические (а-б) и биологические (в) повреждения древесины, способствующие её заражению дереворазрушающими грибами. а – трещина ствола дерева; б – место сломанного и выпавшего сука; в – ходы жуков-древоточцев, вскрытые желной Dryocopus martius. Национальный парк «Себежский», 2015-2016 годы. Фото автора.
Поскольку трутовики питаются древесиной, они постоянно нуждаются в источниках (в частности, углеродного) питания, которые находятся в деревьях первоначально в нерастворимом состоянии. Для того, чтобы перевести необходимые для их питания соединения в растворимое состояние, эти грибы выделяют определённые экзоферменты, расщепляющие древесину. На конечной стадии поражения деревьев дереворазрушающими грибами в них образуется гниль и древесина утрачивает свои структуру и свойства.

Рис. 5. Примеры больных и мёртвых деревьев, ставших доступными для долбления дятлами. а, в – мёртвые гнилые стволы берёзы со следами кормёжки на них белоспинного дятла; б – мёртвый ствол сосны, ошкуренный желной. Национальный парк «Себежский», 2015-2016 годы. Фото автора.

Рис. 6. Примеры естественных дупел в стволах деревьев. а – дупло на месте заросшего механического повреждения в берёзе, занятое осами; б – дупло на месте морозобойной трещины в чёрной ольхе, частично вскрытое дятлами; в – дупло на месте выпавшего сука в иве, заселённое лазоревкой Parus caeruleus. Национальный парк «Себежский», 2014-2016 годы. Фото автора.
Так, при действии грибных ферментов только на целлюлозу в древесине возникает красная и бурая гниль. В результате древесина становится рыхлой и хрупкой, теряет свою упругость и твёрдость (рис. 5) и начинает легко крошиться или растрескиваться на мелкие призматические кусочки. По сравнению со здоровой древесиной, в ней легко происходит «защепистый, или раковистый» излом, поэтому она плохо противостоит ударному изгибу и другим механическим нагрузкам (Ванинская 1965а,б), включая долбление её дятлами; от ударов она не раскалывается, а легко разламывается на куски.
При разложении же грибами и целлюлозы и лигнина – например, плоским трутовиком Ganoderma applanatum или сосновой губкой – в древесине образуется коррозионная, или белая гниль (Раздорский 1955), после чего такая древесина становится мягкой, волокнистой и часто расслаивается по годичным кольцам. Возникшая гниль может распространяться по всему стволу дерева и проникать в толстые ветки. В таких случаях в поражённых стволах могут образоваться естественные дупла (рис. 6), причём на дне таких дупел часто формируется своеобразная рыхлая подстилка из гниющей древесины, которая может служить местом для размещения яиц для птиц – дуплогнездников.
В словаре Ожегова слово «дупло» обозначает полое пространство в стволе дерева; в других словарях и Большой советской энциклопедии – пустота в дереве, выгнившее нутро пня, пустота, отверстие в стволе, образовавшееся на месте выгнившей древесины и т.п. Обычно под дуплами понимают частично закрытые полости в стволах или ветвях деревьев, возникающие чаще в стволах старых или мёртвых деревьев и формирующиеся довольно медленно естественным путём. Возникновению дупел способствуют сырой и влажный климат, близость водоёмов и заболачивание местности, когда деревья оказываются в воде и начинают усыхать, грибковые заражения и гниение древесины. Особенно часто дупла образуются в лиственных деревьях и гораздо реже – в хвойных (Лес России 1995). Иногда живые дуплистые стволы деревьев заселяют насекомые, например муравьи-древоточцы, которые прокладывают внутри свои ходы на глубину до 10 м.
Придерживаясь годичных колец дерева, они выгрызают более мягкие слои, возникающие во время быстрого весеннего роста, и активно способствуют дальнейшему разрушению древесины. Естественные дупла имеют самую разнообразную форму и размеры, а также расположение относительно поверхности земли, ствола дерева или его ветвей, поэтому далеко не все из них пригодны для заселения дятлами.
Например, в дуплах хвойных деревьев с зачастую неровным дном яйца дятлов могут быть легко повреждены об острые выступы древесины. Очень большие дупла мало соответствуют строению гнездовой чаши, и яйца в них раскатываются. К тому же с увеличением размеров летка и самого дупла в него могут проникать не только многие враги, но и конкуренты дятлов, также нуждающиеся в убежищах для гнездования. Хотя некоторую часть первоначально непригодных для них дупел дятлы, несомненно, могут в значительной степени «доделать» и превратить в пригодные для себя (например, расширить леток или даже полость дупла) и использовать их потом в качестве убежищ и даже гнездовых дупел. Именно так дятлы, видимо, и поступали в то время, когда гнездились в основном в естественных дуплах деревьев и, вероятно, постоянно вступали в конкурентные отношения за естественные дупла с другими дуплогнездниками3.

Рис. 7. Сухой ствол сосны Pinus sylvestris с дуплом желны Dryocopus martius (внизу) и дуплом большого пёстрого дятла Dendrocopos major (вверху), нижняя часть летка которого раздолблена дятлами. Национальный парк «Себежский», 2016 год. Фото автора.
Научившись же выдалбливать собственные дупла, дятлы не только ослабили конкурентную борьбу за места гнездования, но и получили возможность осваивать новые пространства, где естественные дупла отсутствовали вовсе. Правда, наряду с этим у них появилась другая необходимость – поиск для выдалбливания дупла деревьев с мягкой древесиной или фаутного древостоя, которая была продиктована, в первую очередь, ограниченными возможностями дятлов к долблению живых деревьев, особенно с твёрдой древесиной. Именно недостаток приемлемых мест для гнездования до сих пор приводит к возникновению довольно жёсткой конкуренции за пригодные места для устройства гнездовых дупел между дятлами как разных, так и одного вида, особенно если плотность их поселения велика, а доля фаутного древостоя на их гнездовых участках незначительна.
Не случайно мы встречаем иногда в лесу деревья, в стволах которых находится сразу по нескольку дупел дятлов, подчас разных видов (рис. 7). Так, в литературе есть сообщения о том, что можно встретить всевозможные комбинации дупел большого пёстрого дятла, малого пёстрого дятла, желны и зелёного дятла, расположенных по два и даже по три в одном дереве (Благосклонов 1939). В частности, известен случай, когда в одном и том же стволе одновременно размножались большой пёстрый и трёхпалый дятлы (Иванчев 1996а). В Псковской области в одном гнилом осиновом пне также было найдено жилое и брошенное дупла трёхпалого дятла и прошлогоднее дупло большого пёстрого дятла (Бардин 2003). В Московской области в половине сухих буреломных осин находили по 2-3 и даже по 5 дупел дятлов в каждом стволе (Благосклонов 1972). Удобные для устройства дупел деревья нередко используются дятлами в течение ряда лет; на некоторых из них бывает по 5-6 дупел желны, большого пёстрого и белоспинного дятлов, выдолбленных в разные годы. Особенно это типично для желны, большая величина которой дополнительно затрудняет выбор подходящего для неё ствола дерева (Благосклонов 1968).
Приёмы, используемые дятлами при долблении древесины
Каковы же способности поведения (приёмы и навыки) дятлов, позволяющие выдалбливать им свои гнездовые дупла? В принципе, набор их не так уж велик и полностью повторяет тот, который они используют при добывании корма путём долбления. Более того, у разных видов дятлов основные приёмы долбления чрезвычайно схожи. Об этом свидетельствуют визуальные наблюдения за той техникой долбления, которую дятлы используют как во время добывания пищи, так и при выдалбливании дупел, а также осмотры шероховатостей на стенках и дне гнездовых дупел, указывающих на направление ударов клювами на тех стадиях постройки дупел, когда движения долбящей птицы скрыты от визуального наблюдения. Очевидно, одинаковые приёмы долбления обусловлены как сходным строением клювов у всех дятлов, так и сходным строением той древесины, которую они долбят, а также и тем, что подобное поведение было свойственно их общим предкам.
На первой стадии долбления, когда древесина ещё не тронута клювом, перед дятлом встаёт, вероятно, самая сложная задача: как проникнуть внутрь неё, если учесть, что его клюв (в отличие от долота или стамески) мало приспособлен для перерубания древесных волокон.
Вместо этого дятел начинает расщеплять и откалывать определённые древесные волокна, нанося вдоль них клювом прямые и косые удары разной силы (рис. 8-9), пока не «размочалит» их до такого состояния, когда данный кусочек древесины полностью теряет свою целостность (рис. 11а-б), а какая-то часть волокон при этом может быть даже перебита клювом.

Рис. 8. Долбление большим пёстрым дятлом вертикального ствола. а – прямой удар клювом, расщепляющий (прорубающий) кору (древесину); б – косой удар клювом, откалывающий (прорубающий) кору (древесину). Национальный парк «Себежский», апрель 2014 года. Фото автора.
Нанося прямой удар клювом, рассекающий растительные волокна вдоль их расположения в древесине (рис. 8а, 9а), дятел не отклоняет головы от вертикальной плоскости тела, и вся сила его удара направлена на глубокое проникновение клюва в древесину и «прорубание» внутрь ствола. Однако при этом мне ни разу не удалось наблюдать, чтобы дятел наносил рассекающий удар клювом поперёк растительных волокон, поэтому отрубание волокон клювом происходит, скорее, лишь случайно. Используя косой удар клювом с поворотом головы относительно вертикальной плоскости тела и направленный под углом к растительным волокнам, дятлу обычно уже после нескольких таких ударов удаётся отбить кусок коры или щепку (рис. 8б, 9б) или расщепить и отделить друг от друга часть волокон, которые, однако, могут оставаться поначалу скреплёнными со стволом одним или обоими своими концами (рис. 11б). Иногда такого же результата дятел добивается и путём прямых ударов клюва, особенно если этому способствует положение его тела и головы относительно периферийных частей ствола.

Рис. 9. Долбление белоспинным дятлом горизонтального ствола. а – прямой удар клювом, расщепляющий (прорубающий) древесину; б – косой удар клювом, откалывающий древесину. Национальный парк «Себежский», февраль 2014 года. Фото автора.

Рис. 10. Выламывание (а-б) и отдирание (в) белоспинным дятлом отщеплённых им кусочков древесины (коры) от ствола. Национальный парк «Себежский», февраль-апрель 2014 года. Фото автора.
Вторым приёмом долбления является откалывание или отламывание дятлом отдельных древесных волокон (щепочек) от ствола клювом. В частности, при описании этого приёма у чёрного дятла К.Н.Благосклонов (1968) замечал, что «подсовывая клюв сбоку и поворачивая голову, желна, как долотом, отщепляет у сырой осины щепки длиной до 15 см». А.Н.Формозов (1974) писал, что желна «не только долбит клювом, но также пользуется им как рычагом, отламывая крепкие щепки сырой древесины толщиной до 2-3 мм и длиной 10-20 см». Другие дятлы тоже отламывают отщеплённые ими кусочки дерева боковыми движениями клюва, пока отдельные щепочки не отделятся от ствола или не сломаются и не превратятся каждая в две щепочки, связанные с пластом других волокон только одним из своих концов (рис. 11а-б).

Рис. 11. Результаты долбления дятлами мёртвой древесины. а – «размочаленный» белоспинным дятлом кусочек ствола берёзы; б – отколотые косыми ударами клюва и сломанные белоспинным дятлом щепочки на стволе ивы; в – кормовая ниша трёхпалого дятла в трухлявом сосновом пне. Фото автора.
Только после этого дятлы применяют свой третий приём: захватывают каждую из таких щепочек клювом, как клещами, и отдирают или выламывают её окончательно из цельного ствола (куска) древесины (рис. 10). Именно таким образом, вовсе не отрубая древесные волокна, а меняя раз за разом места своих прямых и косых ударов клювом, размочаливая (рис. 11а), отщепляя, отламывая (рис. 11б) и отдирая отдельные щепочки от ствола, дятел постепенно расширяет объём образующейся в стволе дерева полости (рис. 11в).
Таким образом, долбление у дятлов – непростой и довольно трудоёмкий процесс, и большинство видов способны раздолбить далеко не любую, тем более живую, древесину, а предпочитают для этого фаутную древесину. Это особенно наглядно на тех этапах долбления, когда при углублении начатой гнездовой камеры дятел вынужден раздалбливать древесину не вдоль её волокон, а с их торца, превращая её отдельные участки в подобие сосновой шишки, раздолбленной в «кузнице» большим пёстрым дятлом. После такого приёма долбления только захват раскрытым клювом отщеплённых и торчащих в виде «ёжика» кусочков древесины и дальнейший их отрыв (отламывание, отрывание, скорее выщипывание) представляется наиболее эффективным для продолжения работы, особенно если учесть, что дятел при этом едва помещается в ограниченном пространстве своего незаконченного дупла
и, видимо, с трудом использует там «традиционные» для него приёмы долбления. И конечно, в такие моменты ему важно иметь податливый субстрат для долбления, в связи с чем он и выбирает обычно стволы деревьев с утратившей свои первоначальные свойства мёртвой или загнившей древесиной.
Выдалбливание дятлами гнездовых дупел
Перед устройством гнездового дупла дятлы приступают к поиску подходящего для этого ствола дерева (нужного им диаметра, с мягкой древесиной, а ещё лучше усохшего или поражённого грибами) и конкретного места на нём. Чаще всего они, в частности, белоспинный дятел (Иванчев 1996б; Бутьев, Фридман 2005а; и др.), выдалбливают дупла там, где гниль ближе всего подходит к поверхности ствола. Для этого дятлы выстукивают приглянувшийся им ствол и, видимо, по звуку определяют минимальную толщину стенки плотной древесины (Севастьянов 1962, 1964). На подобные поиски может уйти не один день,
особенно если учесть, что в природе дятлы нередко бросают начатое ими дупло и начинают новое, иногда на том же дереве (Новиков и др. 1963), а также могут долбить несколько дупел одновременно (Книстаутас, Люткус 2010; Митяй 1985).
В Ленинградской области, например, один самец большого пёстрого дятла обследовал для этого более 15 стволов деревьев, на которых начинал долбление, но потом бросал и искал новые стволы. Другой самец 4 дня долбил дупла одновременно в двух стволах, расположенных в 50 м один от другого. В результате он выдолбил два дупла (глубиной 11 и 17 см), однако бросил оба и выдолбил третье. Третий самец выдолбил себе дупло только в четвёртом дереве (Гавлюк 1972; Митрофанов, Гавлюк 1976). В том же регионе самка белоспинного дятла, активно делавшая новое дупло и почти полностью влезавшая в него, бросила его и выдолбила другое, в нескольких десятках метров от прежнего (Фёдоров 2011). Такие случаи происходят у дятлов постоянно. В Архангельской тайге на 81 дупло большого пёстрого дятла пришлось 75 брошенных: 14 дупел – в живых деревьях, 16 – в сухих, 45 – в пнях (Севастьянов 1962).

Рис. 12. Внешний вид летков в дуплах дятлов. а-в – летки в дуплах большого пёстрого дятла (леток в начале выдалбливания; леток с «порожком»; леток на месте выгнившего сучка (следуя положению бывшего сучка, этот леток уходит внутрь дупла не горизонтально, а в сторону и вниз); г – леток в дупле белоспинного дятла, устроенный под трутовиком; д-е – летки в дуплах желны (леток в начале выдалбливания и леток в готовом дупле). Национальный парк «Себежский», 2014-2016 годы. Фото автора.
Для того, чтобы яснее понять, отчего дятлы начинают выдалбливать, но потом всё же бросают свои незавершённые гнездовые дупла, нужно представить себе весь процесс выдалбливания дятлами дупел. Выдалбливание любого нового дупла начинается с летка. Если дупло устраивается в живом дереве, то первыми долблению подлежат кора дерева и его лубяные волокна, а уже потом древесина (рис. 12а). Иногда, правда, дятлы могут поступить и иначе: в деревьях с твёрдой древесиной они начинают выдалбливать леток дупла как правило на месте выгнившего сучка (рис. 12в, 14б), реже морозобойной трещины, а также на участке ствола, поражённого трутовиком (рис. 12г). Как бы то ни было, в конце постройки летки дупел некоторых видов дятлов более или менее достоверно можно отличить друг от друга по их размерам и форме4, и такие материалы будут представлены мной в следующей статье.
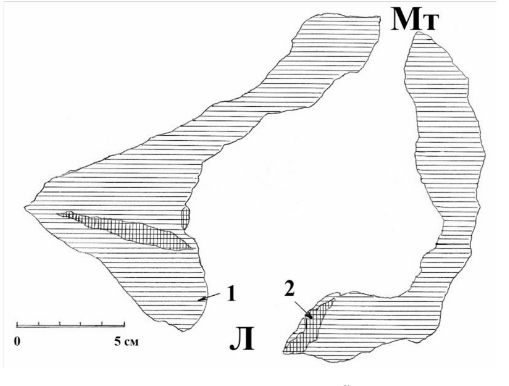
Рис. 13. Схема поперечного спила через гнездовое дупло белоспинного дятла на уровне летка. Л – леток; Мт – морозобойная трещина, 1 – древесина ствола усыхающей чёрной ольхи; 2 – внутренние сучки. Верховья реки Псковы, 1981 год. Рисунок автора.
Наряду с этим практически каждый леток имеет свои особенности, связанные, в первую очередь, с тем местом на стволе и породой и состоянием того дерева, в котором он выдолблен. Например, в одном (правда, единственном) известном мне случае леток в гнездовом дупле белоспинного дятла в Псковской области имел «подсветку» с противоположной стороны ствола, где находилась небольшая морозобойная трещина (рис. 13). Тем не менее, дятлы успешно вывели в этом дупле птенцов в 1981 году.
Гораздо чаще летки в дуплах, например большого пёстрого дятла, бывают устроены на месте бывших сучков (рис. 12в, 15б) и имеют нередко более округлую форму или бо́льшие размеры, чем аналогичные летки в сухих стволах сосны (рис. 12б). В свою очередь, леток в сосне, изображённый на рисунке 12б, снабжён так называемым «порожком», т.е. лоткообразным углублением в его нижней части. Считается, что дятлы специально раздалбливают нижний край летка в виде желобка для удобства влезания в дупло (Иванчев 2005а) и меньшего попадания в дупло дождевой воды (Митрофанов, Гавлюк 1976)*. Но тогда почему же «порожек» имеется далеко не во всех дуплах дятлов, да и какое принципиальное значение он имеет для этих птиц при влезании и вылезании их из дупла, если посмотреть на эти движения у большого пёстрого дятла на рисунке 14а-в?
* В частности, «порожка» никогда не бывает в летках, устраиваемых на месте выпавших сучков, а также в гнилых стволах деревьев с отмершими и ломкими древесными волокнами

Рис. 14. Влезание в дупло с «порожком» и вылезание из него большого пёстрого дятла. Псковский район, 1981 год. Фото автора.
На мой взгляд, дело тут совсем в другом. Если вспомнить приёмы долбления дятлом древесины, описанные выше, легко представить себе, как дятел сначала расщепляет, потом отламывает клювом или, по крайней мере, надламывает им, как рычагом, крайние щепочки и, наконец, отрывает их клювом, как щипцами, от ствола. Такая схема, однако, всегда нарушается, когда дятел приступает к долблению нового (ещё цельного) куска древесины, от которого невозможно отщепить «крайние» щепочки, которые можно было бы отломить боковым движением клюва. В связи с этим, расщепив или размочалив первые волокна (щепочки), дятел не может сломать и даже надломить их клювом, потому что они попрежнему остаются зажатыми с боков соседними (ещё цельными) волокнами, поэтому он вынужден отдирать такие щепочки от ствола не надломленными. Но это, в свою очередь, приводит к тому, что некоторые (живые и гибкие) волокна не отрываются там, где обычно бывает сломана отрываемая щепочка, а образуют «задир» древесины в нижней части летка.
Дятел мог бы избежать этого в том случае, если бы подрубал древесные волокна в нижней части летка, но он этого никогда не делает, да и его клюв для этого не приспособлен. Тем не менее, при продвижении дятла внутрь ствола каждый «задир» начинает закономерно укорачиваться в связи с тем, что его длину ограничивают лежащие ниже нетронутые части ствола*. Возможно, по той же причине большинство летков у дятлов получаются при долблении не цилиндрической, как принято считать, а иной, как правило, неправильной конусовидной формы, постепенно сужаясь кверху по мере их долбления (рис. 15а, 17, 19).
*Помимо того, после образования первой, пусть даже небольшой, полости в стволе (после выдирания первых щепочек), у дятла появляется возможность не только расщеплять, но и отламывать клювом «крайние» щепочки, поэтому задиры в древесине не идут в стороны, а начинают уменьшаться, создавая форму жёлоба.
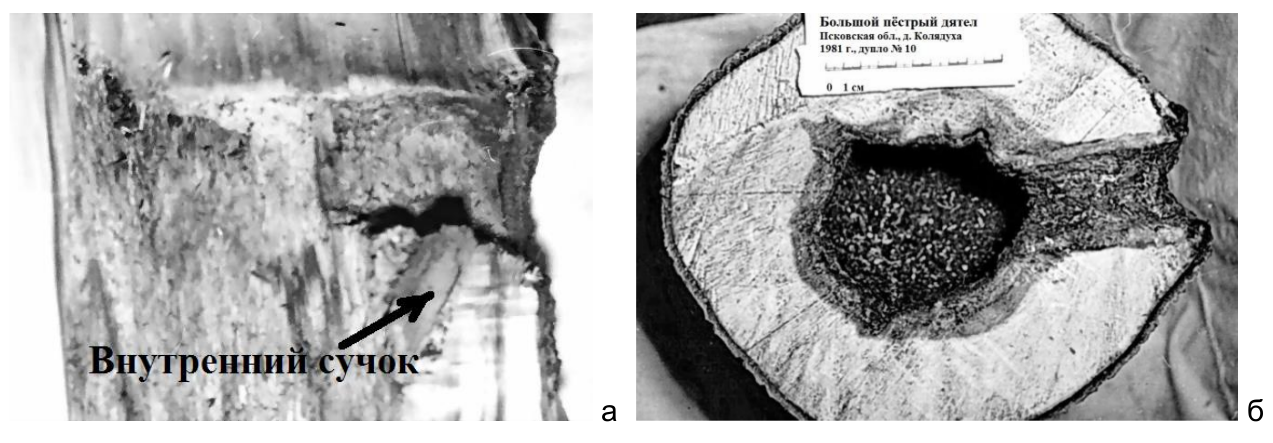
Рис. 15. Вертикальный (а) и горизонтальный (б) распилы через летки больших пёстрых дятлов в живых, но загнивших осинах. Прогнившая древесина выглядит на обоих рисунках более тёмной. Псковский район, верховья реки Псковы, июнь 1981 года. Фото автора.
Свойственными дятлам приёмами долбления древесины при большом размере тела птицы объясняется, по-видимому, особенная форма летка у желны. В отличие от других дятлов, она имеет иногда почти прямоугольную форму, но чаще, однако, форму вытянутого и не всегда правильного овала, и никогда не бывает округлой, особенно в нижней части летка (рис. 12е). Скорее всего, мощные удары клювом и крупные отщепляемые желной щепки не позволяют ему аккуратно осуществлять плавный переход с одного места долбления на другое, следуя точно по окружности. Хотя не следует думать, что у других видов дятлов формы летков имеют абсолютно круглую форму и одинаковые, характерные для каждого вида размеры. Примером тому могут служить форма и размеры летка в одном из дупел большого пёстрого дятла в усыхающей берёзе в верховьях реки Псковы в Псковской области, которые удалось отобразить в мае 1981 года года путём съёмки разных частей летка на кальку в их натуральную величину (рис. 16).
На данном примере хорошо видно, что дятел не всегда долбит древесину «как хочет», а зачастую, вероятно, «как сложится» в зависимости от её конкретных особенностей, а если «не сложится», то дятел бросает начатое дупло и приступает к поиску более удобного места. В случае же успеха дятлы «работают» над летком пока он не начнёт соответствовать размеру взрослой птицы (Благосклонов 1969). На первом этапе долбления в качестве первоначального «шаблона» для примерки большие пёстрые дятлы, например, используют свою голову, регулярно «заглядывая» в начатое дупло, а затем устраняя лишнюю древесину в наиболее узких участках летка, оказывающую, по-видимому, наибольшее давление на оперение головы. На втором этапе, не обнаружив особых недоделок в летке, они начинают подолгу отщипывать крошечные кусочки древесины и проверять размеры и округлость летка путём «многочисленных наклонов внутрь дупла своей “плечевой зоны” тела» (Симкин 1977).
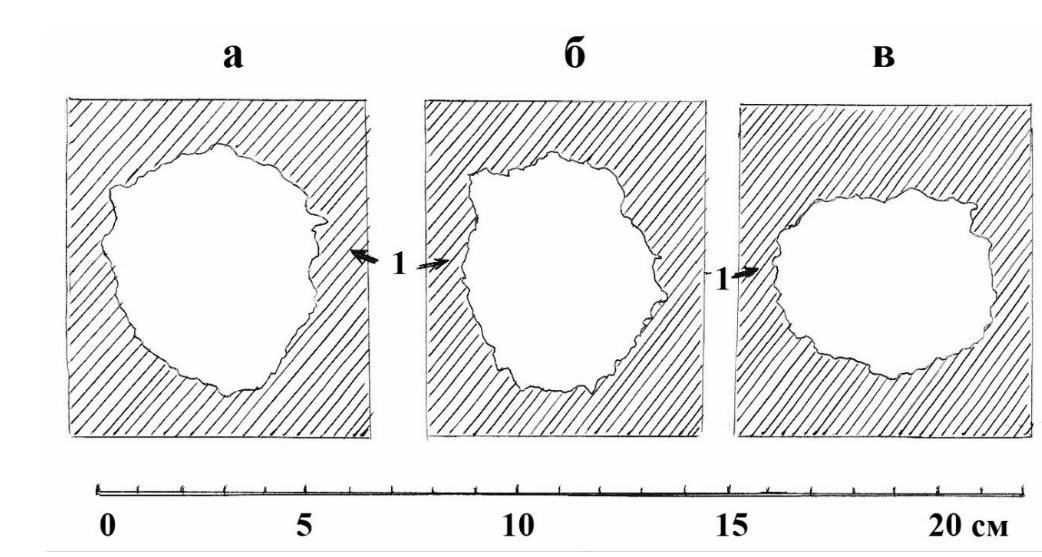
Рис. 16. Форма и размеры разных частей одного из летков большого пёстрого дятла на разном удалении их от поверхности ствола. а – на коре дерева; б – под корой дерева; в – на переходе летка в гнездовую камеру; 1- древесина вокруг летка. Рисунок автора.
Особого упоминания в отношении разных возможностей долбления древесины заслуживает, конечно, малый пёстрый дятел. Из-за своей малой величины и слабого клюва его строительные способности так ограничены, что он чаще других дятлов вынужден искать себе значительно прогнившие деревья, чтобы преодолеть в них наружный (более твёрдый) слой древесины, хотя и его гнездовые дупла находят иногда в таких деревьях с твёрдой древесиной, как дуб, вяз и груша (Иванчев 1995а).
Выдалбливая леток, все дятлы находятся снаружи дупла, т.е. в довольно «комфортных» для долбления условиях. Совсем другая картина наблюдается, когда они переходят к выдалбливанию гнездовой камеры. Именно тогда, помимо тесноты в едва начатом дупле, дятлы вынуждены (судя по результатам их долбления на рисунке 17) продвигаться не только поперёк, но и вдоль ствола (т.е. почти вниз головой) и раздалбливать древесину не своим традиционным способом – вдоль древесных волокон, а в основном с торца, а также вырывать при этом отколотые или размочаленные волокна, сидя в летке и не имея достаточно устойчивой опоры для тела, поэтому им особенно трудно преодолевать на своём пути не только встречающиеся в этой зоне внутренние сучки (рис. 18а), но даже участки ствола со здоровой и менее податливой для долбления древесиной (рис. 18б).

Рис. 17. Вертикальный распил через гнездовое дупло большого пёстрого дятла. Псковская область, верховья реки Псковы, 1981 год. Фото автора.
Поскольку после долбления на стенках дупла остаются многочисленные «шипы», представляющие собой не до конца отломанные или оторванные дятлом кусочки древесины, торчащие в определённых направлениях на расстояния до 5-6 мм, в данном дупле – внутри выделенной фигуры на фото – по разному характеру «шероховатости» на стенках дупла можно заметить границы того «минидупла», которое дятел выдолбил, сидя в летке. По своим размерам оно соответствует примерно размерам дятла, позволяет ему разворачиваться и продолжать долбление, сидя уже в дупле, в вертикальном положении.
Но главное, что именно на этой стадии (выдалбливания свода гнездового дупла и перехода летка в гнездовую камеру) дятел в первый раз оценивает возможность размещения в выбранном им стволе дерева всей гнездовой камеры. В большинстве случаев, по-видимому, ствол признаётся пригодным, если площадь загнившей в нём или податливой для долбления древесины соответствует хотя бы минимальным размерам гнездовой камеры (по ширине), а для этого производящая долбление птица должна как минимум залезть и развернуться в этом «минидупле» (рис. 17). Вертикальные и горизонтальные распилы начатых дятлами дупел показывают, что на самом деле площадь загнившей древесины внутри ствола напротив выдолбленного летка далеко не всегда соответствует необходимым размерам будущей гнездовой камеры (рис. 18б). Поэтому на этом дальнейшее долбление прекращается.
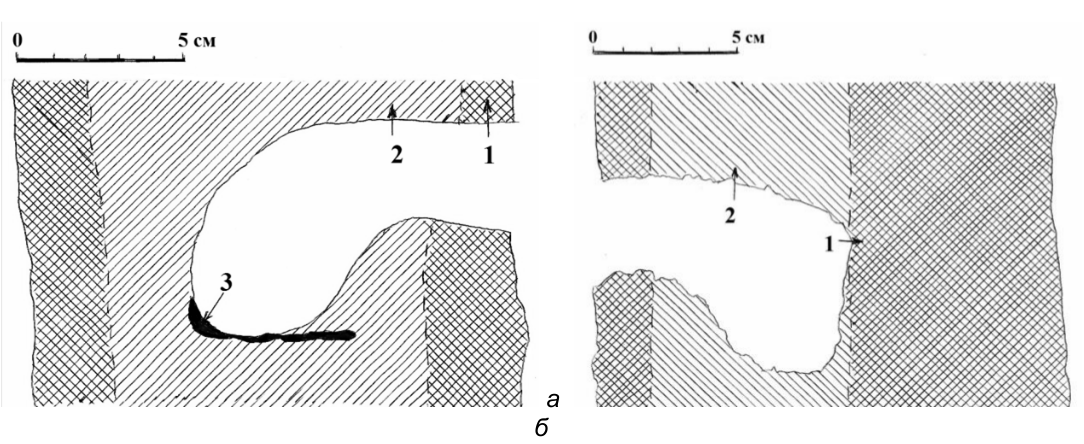
Рис. 18. Вертикальные распилы начатых и брошенных малым пёстрым дятлом дупел. а-б – начатые и брошенные дупла: 1 – здоровая древесина; 2 – загнившая древесина; 3 – остатки внутреннего сучка. Псковская область, верховья реки Псковы, 1981 год. Рисунок автора (на основе кальки с распилов дупел).
Тот факт, что при долблении гнездового дупла дятлы вынуждены постоянно считаться с распространением в стволе разных по твёрдости участков древесины, наглядно доказывается тем, что границы полостей их дупел во многих стволах деревьев практически совпадают с границами загнившей древесины, даже если сами дупла получаются при этом не оптимальной для дятлов, а зауженной и даже «щелевидной» формы (рис. 19). Вполне понятно, что на дне узких дупел будет трудно разместиться сразу нескольким подросшим птенцам (рис. 20).
Большой пёстрый дятел, например, в период роста птенцов иногда даже расширяет дупло, раздалбливая его клювом и выбрасывая из него щепу (Птушенко, Иноземцев 1968). Но дятлы довольствуются и узкими дуплами, и это косвенно свидетельствует о том, что в ряде случаев они явно испытывают дефицит в нужных для устройства дупел деревьях. Более того, о подобном дефиците свидетельствуют и те случаи, когда дятлы выбирают для выдалбдивания гнездовых дупел недостаточно толстые, хотя и удобные для долбления стволы с прогнившей или мягкой древесиной, а в результате стенки их дупел оказываются иногда чрезвычайно тонкими, что напрямую сказывается не только на прочности стволов и, соответственно, самих дупел, но, вероятно, и на температурном режиме внутри таких дупел. Сравнительно часто дупла с тонкими стенками встречаются у белоспинных дятлов (рис. 21А-в), но их можно найти и у других дятлов, например, у седых (рис. 21г-д).
Однако это, как правило, приводит к тому, что время от времени, в периоды сильных ветров, стволы деревьев с такими дуплами не выдерживают напора и ломаются по месту нахождения дупел (рис. 22).
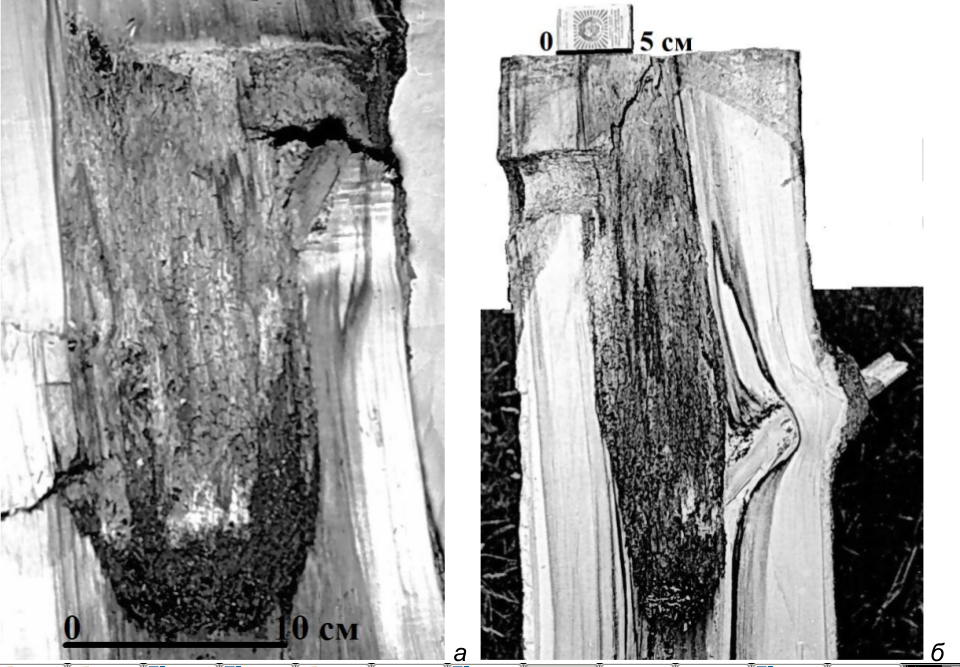
Рис. 19. Вертикальные распилы через гнездовые дупла большого пёстрого дятла в живых осинах. а – дупло нормальной формы (в стволе с обширной площадью загнившей древесины); б – «щелевидное» дупло (в стволе с небольшой площадью загнившей древесины). Здоровая древесина на фото выглядит гораздо светлее загнившей. Верховья реки Псковы, 1981 год. Фото автора.
В других случаях, когда объём гнилой древесины в стволе дерева достаточно обширен, дятлы (в частности, малые пёстрые) при встрече внутренних сучков или крепких участков древесины нередко имеют возможность обойти их стороной. В результате они не бросают начатого ими дупла, но форма окончательной гнездовой камеры бывает у них весьма замысловатой (рис. 23). Поэтому поперечные срезы дупла малого пёстрого дятла редко имеют правильную круглую форму, что бывает чаще в тонких осинах (диаметром до 12 см); в толстых стволах полости дупла очень разнообразны по форме (Иванчев 1995а).

Рис. 20. Расположение птенцов в дуплах дятлов. а – Dendrocopos major (Псковская область, 1981 год, фото автора); б – Dryobates pubescens medianus (фото A.A.Allen, по: Bent 1939).
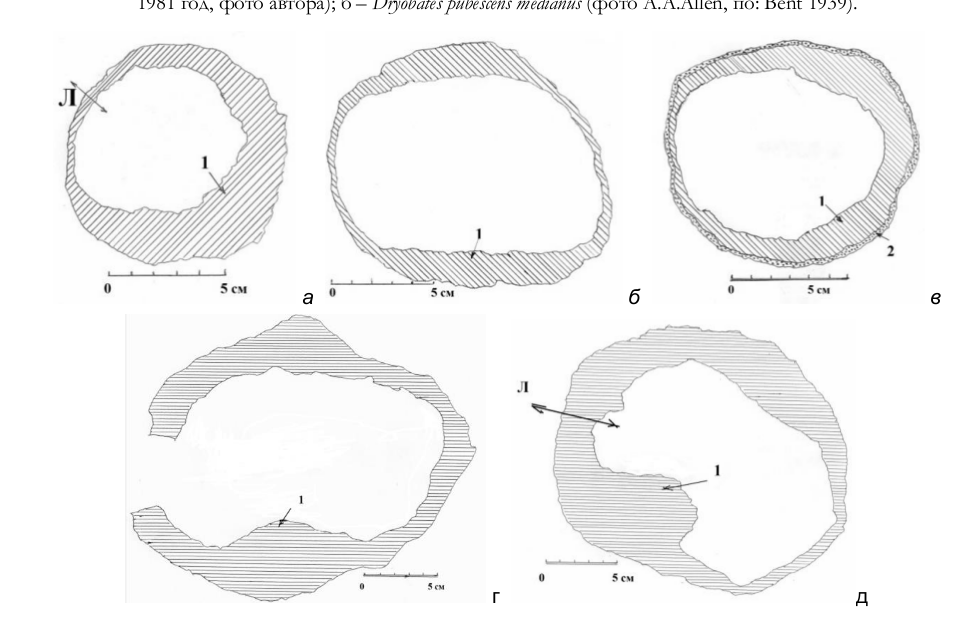
Рис. 21. Горизонтальные распилы через гнездовые дупла белоспинного (а-в) и седого (г-д) дятлов. Спилы на уровне: а – нижнего среза летка; б – середины гнездовой камеры; г – летка; в, д – лотка. Л – направление летка. 1 – древесина дерева. Псковский (а-в) и Себежский (г-д) районы Псковской области, 1981 (а-в) и 1986 (г-д) годы. Рисунки автора.
В зависимости от обстоятельств весьма разными по форме бывают дупла и у других видов дятлов. Так, одно из гнездовых дупел большого пёстрого дятла в сосне, показанное мне В.А.Фёдоровым в 1988 году в Себежском Поозерье, имело форму плоской фляги с шириной не более 8 см, «прилепившейся» на периферии огромного, но лишь частично загнившего ствола диаметром 45 см на уровне летка; леток в нём располагался на месте выгнившего сука и вёл внутрь дупла не перпендикулярно к поверхности ствола, а наискось – в сторону и вниз.

Рис. 22. Ствол ивы с гнездовым дуплом белоспинного дятла, сломанный порывом ветра. Квадратами на фото помечено место слома ствола ивы и место положения гнездового дупла. Национальный парк «Себежский», 2014 год. Фото автора.
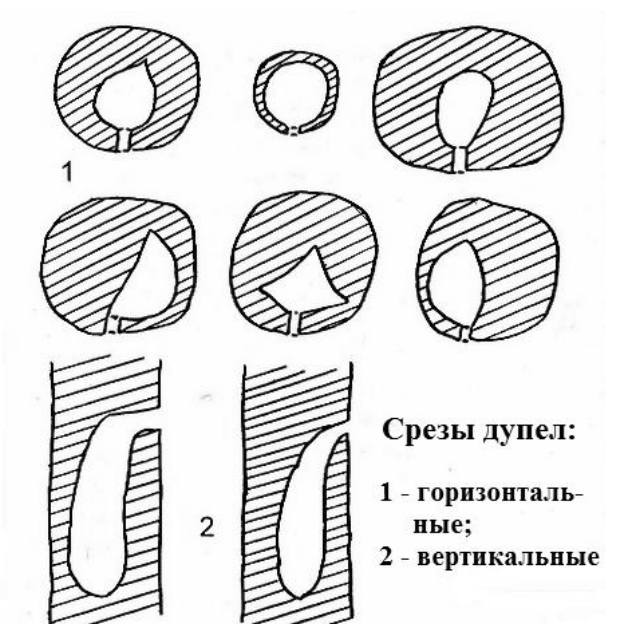
Рис. 23. Схемы разрезов через дупла малого пёстрого дятла. По: Иванчев 1995а.
Возвращаясь к рисунку 17 и анализу разных картин следов долбления, остающихся на стенках дупла, можно отметить, что примерно всю вторую половину или даже последние 2/3 гнездовой камеры дятлы выдалбливают, сидя уже не в летке, а в дупле, в более удобном для долбления положении, и приобретают всё большую свободу для движений по мере увеличения объёма камеры. В некоторых случаях на стадии завершения гнездовой камеры большие пёстрые дятлы, например, вновь приобретают способность раздалбливать подгнившие внутренние суки (рис. 19б). В других случаях, например, седые дятлы (рис. 21д) и белоспинные (рис. 24) не могут раздолбить какой-то участок древесины даже на уровне своего будущего лотка, и он получается неправильной формы, к тому же весьма неудобной для расположения в нём насиживающей яйца птицы.
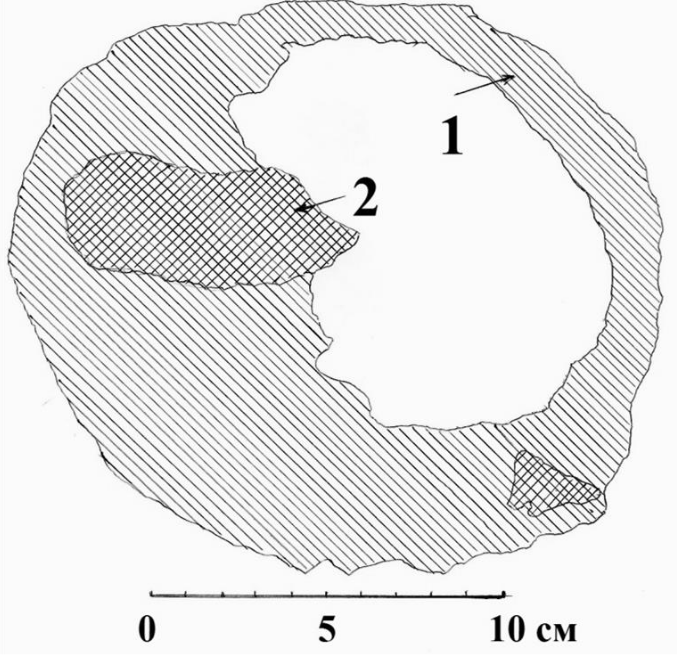
Рис. 24. Горизонтальный распил на уровне лотка через дупло белоспинного дятла в чёрной ольхе. 1 – обычная, слегка загнившая древесина дерева; 2 – участки с твёрдой древесиной. Псковская область, Себежское Поозерье, 1988 год. Рисунок автора.
Тем не менее, рано или поздно дятлы завершают выдалбливание гнездовой камеры. Этот процесс знаменуется последними «работами» на дне дупла, которое приобретает чаще всего вид более или менее ровной площадки или слегка вогнутого лотка (рис. 19а-б). Однако в некоторых дуплах подобная деятельность может оказаться почему-то незаконченной, поэтому поверхность дна дупла в них остаётся совершенно непригодной для насиживания кладки яиц (рис. 17, 25).
Такое поведение дятлов можно объяснить, по-видимому, несколькими причинами. Первая – это какие-то неожиданно возникшие препятствия для дальнейшего долбления (внутренние суки, свилеватая древесина и пр.), с которыми дятлы не могут справиться. Однако часто долбление дупел, например в сильно прогнивших осинах, завершается в такой древесине, которую можно выщипывать даже пальцами, и несмотря на это, дно дупла остаётся малопригодным для насиживания яиц. Вторая причина – это сроки откладки яиц. Если выдалбливание дупла по каким-то причинам сильно затягивается, самка (в отличие от самца) может начать устраивать лоток независимо от состояния дна дупла и действия самца. Но главная причина, скорее всего, заключается в том, что дно дупла в любом его состоянии, если только оно не устроено в совершенно гнилой мягкой древесине, не пригодно для размещения на нём яиц, потому что его поверхность равномерно покрыта шипами длиной 5-6 мм, остающимися после долбления на всей его поверхности. Поэтому практически в каждом гнездовом дупле дятлы вынуждены устраивать специальную выстилку на дне дупла, формируя лоток из древесной трухи или щепы (рис. 15б, 17, 19а-б, 25, 26), о чём уже подробно говорилось в предыдущей статье (Фетисов 2017). В дополнение к ранее изложенным сведениям подчеркну, что формирование лотка в гнездовых дуплах свойственно как европейским, так и американским видам дятлов, т.е. носит общий характер, и такие примеры проиллюстрированы на рисунке 26.
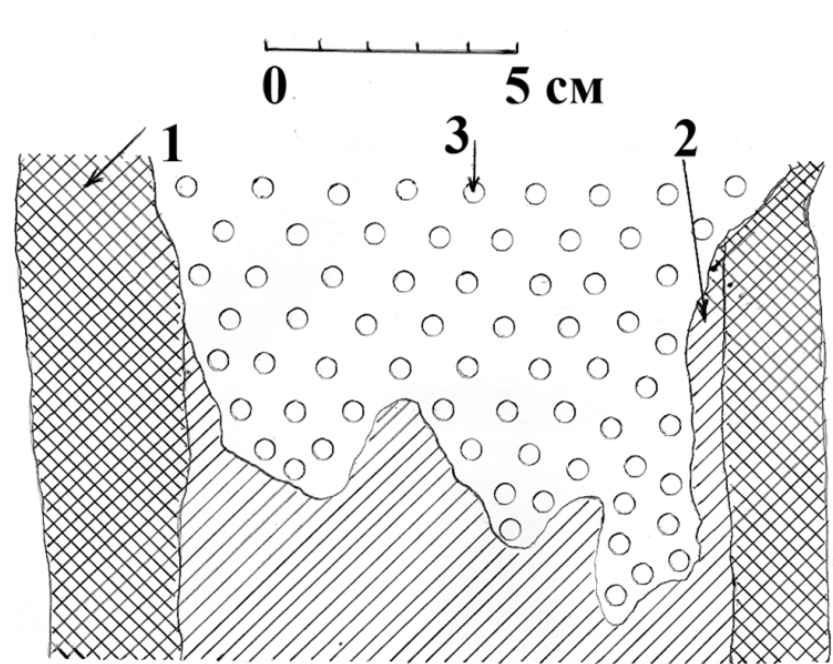
Рис. 25. Вертикальный распил на уровне лотка через гнездовое дупло большого пёстрого дятла в живой загнившей осине. 1 – нормальная древесина; 2 – гнилая древесина; 3 – лоток (выстилка) из древесной трухи. Псковская область, верховья реки Псковы, 1981 год. Рисунок автора.
Однако даже на этом выдалбливание гнездового дупла у дятлов не заканчивается. Как известно, материал для выстилки дна дупел дятлы могут приносить в дупла извне (Н.А.Зарудный – по: Гладков 1951; Ковшарь и др. 1978; Афанасьев 2014; Фетисов 2017) или откалывать для этого мелкие щепочки («щепу») со стенок внутри дупел (Иванчев 2005а,б; Головань 2006; и др.). По данным В.П.Иванчева, дятлы получают щепу путём раздалбливания вполне определённых частей дупла.
Желна, белоспинный, средний и большой пёстрый дятлы делают это путём долбления преимущественно свода дупла, изредка боковых стенок; седой и трёхпалый дятлы – боковых стенок и изредка верхней части дупла; малый пёстрый дятел – боковых стенок в местах расположения наиболее прогнившей древесины. По моим наблюдениям, дятлы делают это чаще всего в узких дуплах, на стенках которых почти никогда не остаётся гнилой древесины, и поэтому, уцепившись лапами за край летка с внутренней стороны дупла, они выщипывают мелкие кусочки такой древесины в районе свода дупла, где им удобнее всего до неё дотянуться (рис. 27б). В широких же дуплах гнилая древесина нередко остаётся на стенках свежевыдолбленного дупла и её бывает, вероятно, достаточно для выстилки дна того же дупла.
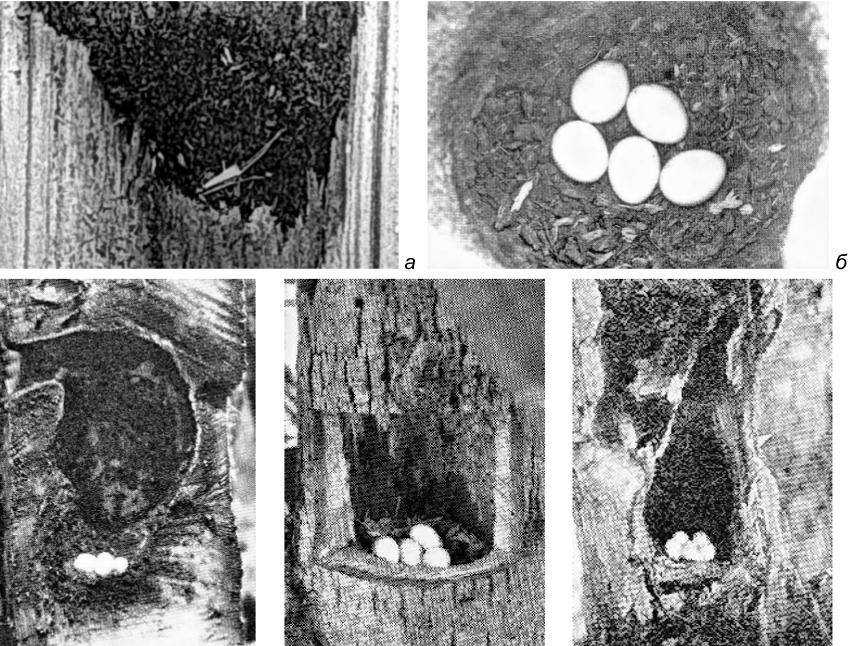
Рис. 26. Выстилка в лотках гнездовых дупел дятлов. а – Dendrocopos major (фото автора); б – Colaptes auratus (фото A.A.Allen, по: Bent 1939); в – Centurus uropygialis (фото M.F.Gilman, по: Bent 1939); г – Melanerpes erythrocephalus (фото A.D.DuBois, по: Bent 1939); д – Dryobates nutalli (фото W.M.Pierce, по: Bent 1939).
С началом откладки яиц строительство гнездового дупла у дятлов не заканчивается. В течение всего периода насиживания как самцы, так и самки добавляют на дно дупла «щепу», добытую с внутренних стенок гнездовой камеры (Митрофанов, Гавлюк 1976; Иванчев 1993, 1994; 2005а,б). Малый пёстрый дятел, например, после появления яиц выдалбливает мелкие воронкообразные углубления в основном в боковых стенках дупла или же, следуя распространению гнилой древесины, раздалбливает боковые стенки (Иванчев 2005б), в результате чего дупла этого вида имеют весьма разнообразное строение (рис. 23).
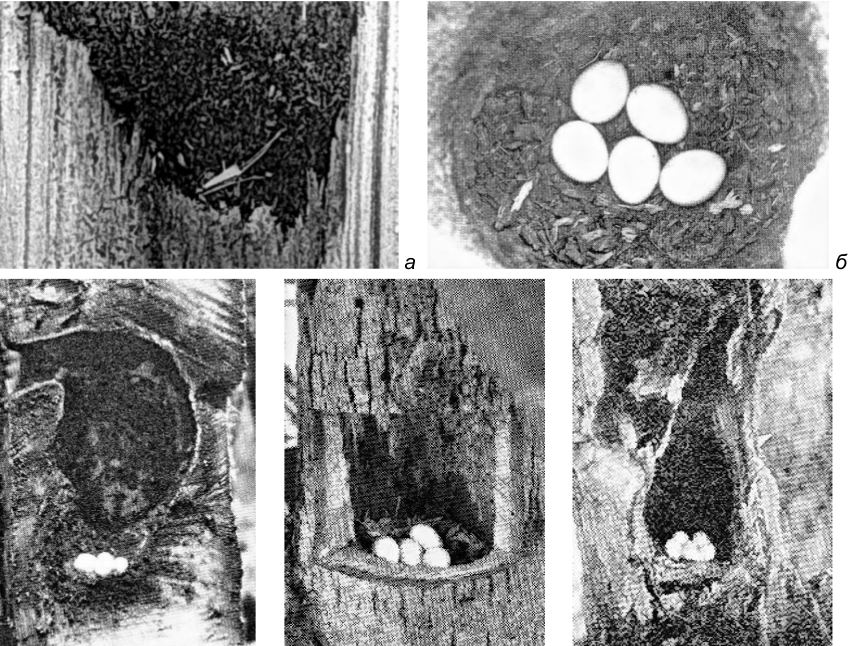
Рис. 27. Вертикальные распилы через гнездовые дупла больших пёстрых дятлов в живых загнивших осинах (в районе свода дупла). а – одно из дупел с широкой гнездовой камерой и обычным сводом дупла; б – дупло с узкой гнездовой камерой и раздолбленным сводом дупла. 1 – кора; 2 – живая здоровая древесина; 3 – гнилая древесина. Псковская область, верховья реки Псковы, 1981 год. Рисунок автора.
Правильной формы дупла часто бывают у зелёного дятла (Благосклонов 1968; Гладков 1951), который, поселяясь в старых дуплах большого пёстрого и седого дятлов, добывает щепу для выстилки дупел и значительно раздалбливает для этого боковые стенки (Иванчев 2005в), «подгоняя» также эти дупла под собственные размеры, необходимые для нормального расположения насиживающей кладку птицы. Однако даже в стволах с достаточным объёмом податливой для долбления древесины дятлы редко выдалбливают дупла правильной колбообразной или грушевидной формы, как об этом иногда пишут в литературе. Это связано с тем, что в дуплах такой формы взрослым и птенцам придётся подниматься к летку не по вертикальной, а по отрицательно наклонённой поверхности. Поэтому полости гнездовых камер дятлов представляют собой чаще всего «трубы» разного диаметра с боковым входом вверху (рис. 19а-б). При наличии выбора дятлы предпочитают устраивать дупла снизу в наклонённых стволах, облегчая себе в какой-то мере передвижение внутри таких дупел.
Интенсивность выдалбливания дятлами дупел
Интенсивность выдалбливания дятлами дупла и сроки его долбления зависят одновременно от многих факторов: породы и состояния дерева, климатических условий, активности птицы, её возраста, фактора беспокойства (Митрофанов, Гавлюк 1976) и, возможно, других.
Так, в стволе липы сирийские дятлы Dendrocopos syriacus выдалбливали и удаляли за один день около 300-320 см3 древесины; ежедневно углубляя дупло в среднем на 6-7 см, и вся работа занимала у них 5-7 дней. В тополе же долбление шло значительно медленнее, и одно дупло было готово только на 21-й день (Марисова, Бутенко 2013). В неповреждённых сердцевидной гнилью осинах в Окском заповеднике чёрные дятлы за день углубляли дупло на 1.0-3.5 см, а в поражённых грибами – на 3.3-5.5 см (Иванчев 1995б).
В сырые и холодные дни интенсивность работы дятлов значительно снижается (Поливанов 1981). Так, в условиях холодной и затяжной весны большим пёстрым дятлам требуется на 3 дня дольше, чтобы закончить дупло, а в период сильных дождей с порывистым ветром они могут прерывать долбление на 3-4 ч и даже на сутки (Гавлюк 1972; Митрофанов, Гавлюк 1976).
По мере выдалбливания дупла чёрные дятлы выбрасывают щепу по нескольку раз подряд, максимально – до 63 порций за один «сеанс», меняя при этом поведение в зависимости от фазы завершения строительства (Иванчев 2005г). Большие пёстрые дятлы работают в начале строительства в первой половине дня, затрачивая на долбление чуть более 1 ч. В последующем они начинают долбить и во второй половине дня, затрачивая на это в общей сложности 2 ч 40 мин (Поливанов 1981).
Малые пёстрые дятлы долбят за день обычно 5-8 раз по 7-28 мин, но в конце строительства могут долбить непрерывно иногда по 30-58 мин. Общее время, затрачиваемое ими на долбление дупла в конечные этапы гнездостроения, у них также увеличивается и занимает более 42% светлого времени суток (Иванчев 1995а). Седой дятел, подолбив несколько минут, выбрасывает щепки обычно 18-32 (до 70) раз за «сеанс» и, немного посидев после этого в летке, вновь приступает к долблению (Иванчев 2005д). За день работы седые дятлы углубляют дупло на 3.5-5.0 см (Иванчев 2001).
Продолжительность выдалбливания дятлами дупел
Продолжительность выдалбливания дятлами дупла, как и интенсивность долбления, зависит в первую очередь от того, насколько удачно выбрано место для дупла, хотя вместе на этот показатель воздействуют и другие факторы: погодные условия, ранние или поздние сроки размножения, степень участия брачного партнёра и т.д. К сожалению, такие факторы не всегда учитываются и подобные наблюдения редко приводятся в литературе. Тем не менее, некоторые сведения об этом всё же есть. У зелёного дятла на выдалбливание дупла уходит от 11 дней (Иванчев 1995в) до 20 дней (Книстаутас, Люткус 2010). Седой дятел затрачивает на выдалбливание одного дупла 8-19, в среднем по 12.5 сут (Иванчев 2005д), по данным из Сумской области – 5-8 (Афанасьев 1996), в Приднепровской лесостепи – 13-20 сут (Митяй 1985). У желны строительство дупла продолжается, по данным разных авторов, 10-25 дней (Гладков 1951), в частности, в Ленинградской области – около 2 недель (Мальчевский, Пукинский 1983), в Рязанской области – 11-16 сут (Иванчев 1995б), в Приднепровской лесостепи (n = 7) – 17-23 сут (Митяй 1983, 1985).
Большой пёстрый дятел затрачивает на постройку гнездового дупла около 2 недель (Костин 1983). В Белгородской области на это уходит обычно всего несколько дней (Новиков и др. 1963), в Ленинградской области – 7-15 (Митрофанов, Гавлюк 1976) или 10-12 дней (Гавлюк 1972), в Приднепровской лесостепи – 9-12 дней (Митяй 1985), в Якутии 13 дней (Андреев 1974), в Рязанской области – 5-20, в среднем 11.2 сут (Иванчев 1998). В вольере Ленинградского зоопарка один самец выдолбил дупло в куске ствола дерева с прогнившей древесиной всего за 3 дня (Маклашова, Смирнов 1979). У белокрылого дятла процесс строительства дупла длится 7-8 дней, но одно дупло дятлы выдалбливали 17 дней (Абдреимов 1974, 1975). У сирийского дятла Dendrocopos syriacus на постройку одного дупла в загнившей вербе ушло 6 дней, а другого, в средневозрастной осине – 8 дней (Афанасьев 2014). По другим данным, на выдалбливание дупла ему требуется от 5 до 21 дня (Марисова, Бутенко 2013; Бутьев, Фридман 2005б). Продолжительность выдалбливания дупла среднего пёстрого дятла Dendrocopos medius достигает 12-18 (Митяй 1985), иногда более 20 дней (Косенко, Кайгородова 2003). На выдалбливание дупла обычно уходит около 20 дней, на его отделку – ещё 8 дней (Blume 1968; Cramp 1985).
Одна самка белоспинного дятла в Окском заповеднике построила ночёвочное дупло в сильно прогнившем дубе за 8 дней (Иванчев 1997). Малый пёстрый дятел затрачивает на выдалбливание своего гнездового дупла, по разным данным, не менее недели: в Приднепровской лесостепи – 12-17 дней (Митяй 1985), в Южном Приморье – 7-8 дней, но в одном случае около месяца (Поливанов 1981). Наиболее интенсивное выдалбливание наблюдается за 1-2 недели, а его завершение – за 1-2 дня до откладки первого яйца (Прокопов 2011).
В завершение хочу от всей души поблагодарить за помощь в работе своих друзей В.И.Голованя, И.В.Ильинского и И.Г.Милевского, а также моего бывшего полевого руководителя Ю.Б.Пукинского. Без их участия в полевых условиях большинство рисунков, касающихся распилов гнездовых дупел дятлов, вряд ли увидели бы свет.
Литература
Абдреимов Т. 1974. Особенности гнездовой жизни некоторых птиц в тугаях низовьев Аму-Дарьи // Материалы 6-й Всесоюз. орнитол. конф. М., 2: 3-4.
Абдреимов Т. 1975 . К биологии размножения равнинного белокрылого дятла из низовьев Аму-Дарьи // Вестн. Каракалпак. фил. АН УзССР 4 (62): 33-35.
Андреев Б.Н. 1974 . Птицы Вилюйского бассейна. Якутск: 1-311.
Афанасьев В.Т. 1996 . К биологии малоизученных видов дятлов северной части Сумской области и сопредельных районов // Працi Украiн. орнiтол. товариства 1: 40-44.
Афанасьев В.Т. 2014 . Сирийский дятел Dendrocopos syriacus в Сумском Полесье // Рус.орнитол. журн. 23 (993): 1295-1297.
Бардин А.В. 2003 . Находка гнезда трёхпалого дятла Picoides tridactylus в окрестностях Печор // Рус. орнитол. журн. 12 (228): 750-752.
Благосклонов К.Н. 1939. О значении дупел дятлов в лесном хозяйстве // Сб. научн. студенч. работ. М., 9: 68-82.
Благосклонов К.Н. 1952. Охрана и привлечение птиц, полезных в сельском хозяйстве. М.: 1-259.
Благосклонов К.Н. 1968 . Гнездовые деревья дятлов в различных типах леса // Орнитология 9: 95-102.
Благосклонов К.Н. 1969 . Гнездостроительные адаптации лесных птиц и развитие дуплогнездности // Научн. докл. высш. школы. Биол. науки 3: 13-22.
Благосклонов К.Н. 1972 . Охрана и привлечение птиц. М.: 1-240.
Бутьев В.Т., Фридман В.С. 2005а. Белоспинный дятел Dendrocopos leucotos (Bechstein, 1803) // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: 383-397.
Бутьев В.Т., Фридман В.С. 2005б. Сирийский дятел Dendrocopos syriacus (Hemprich et Ehrenberg, 1833) // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: 360-371.
Ванинская Ю.М. 1965а. Изменения в строении древесины сосны и ели при поражении дереворазрушающими грибами // Экология древесных растений. Минск: 23-32.
Ванинская Ю.М. 1965б. Влияние механических нагрузок на анатомический характер разрушения поражённой грибами древесины хвойных пород // Экология древесных растений. Минск: 54-59.
Гавлюк Э.В. 1972. Экологическая характеристика птиц-дуплогнездников и полуддуплогнездников в Лужском районе Ленинградской области // Питание, размножение и генетика животных. Л.: 108-123.
Гладков Н.А. 1951. Отряд Дятлы Picariae или Piciformes // Птицы Советского Союза. М. 1: 547-617.
Головань В.И. 2006. Территориальное распределение и численность дятлов на двух модельных площадках на юго-западе Ленинградской области // Рус. орнитол. журн. 15 (306): 19-23.
Зимин В.Б. 1968. Факторы, влияющие на размещение гнёзд дятлов в Карельской тайге // Конф. молодых биол. Карелии. Тез. докл. Петрозаводск: 112-113.
Иванчев В.П. 1993. Видовые особенности биологии размножения дятлов как основа методов поиска их гнёзд // Рус. орнитол. журн. 2, 2: 215-221.
Иванчев В.П. 1994. Биология гнездования большого пёстрого дятла Dendrocopos major в
Окском заповеднике // Рус. орнитол. журн. 3, 2/4: 303-318.
Иванчев В.П. 1995а . Биология гнездования малого пёстрого дятла в Окском заповеднике // Тр. Окского заповедника 19: 140-158.
Иванчев В.П. 1995б. Места расположения, устройство дупел и гнездостроительное поведение желны Dryocopus martius в Окском заповеднике // Рус. орнитол. журн. 4, 3/4:97-102.
Иванчев В.П. 1995в . Особенности биологии зелёного дятла у южных пределов ареала // Тр. Окского заповедника 19: 101-114.
Иванчев В.П. 1996а. Случай совместного гнездования в одном дереве большого пёстрого и трёхпалого дятлов // Орнитология 27: 289-290.
Иванчев В.П. 1996б. Распространение, численность и экология белоспинного дятла Dendrocopos leucotos в Европейской части России // Рус. орнитол. журн. 5, 3/4: 117-128.
Иванчев В.П. 1997. Выдалбливание белоспинным дятлом Dendrocopos leucotos дупла для ночёвки зимой // Рус. орнитол. журн. 6 (30): 3-4.
Иванчев В.П. 1998. Гнездостроительная деятельность большого пёстрого дятла // Современная орнитология. М.: 157-180.
Иванчев В.П. 2001. К экологии седого дятла в Окском заповеднике // Орнитология 29: 155-161.
Иванчев В.П. 2005а. Отряд Дятлообразные – Piciformes // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: 281-283.
Иванчев В.П. 2005б. Малый пёстрый дятел Dendrocopos minor (Linnaeus, 1758) // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: 401-412.
Иванчев В.П. 2005в. Зелёный дятел Picus viridis Linnaeus, 1758 // Птицы России и сопредельных регионов: Совообразные – Дятлообразные. М.: 298-305.
Иванчев В.П. 2005г. Желна Dryocopus martius (Linnaeus, 1758) // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: 319-327.
Иванчев В.П. 2005д. Седой дятел Picus canus Gmelin, 1788 // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: 309-319.
Иноземцев А.А. 1986. Отряд Дятлообразные (Piciformes) // Жизнь животных. М., 6: 338-359.
Книстаутас А., Люткус А. 2010 . Материалы по биологии зелёного дятла Picus viridis на юго-востоке Литвы // Рус. орнитол. журн. 19 (552): 342-343.
Ковшарь А.Ф., Жуйко Б.П., Пфеффер Р.Г., Белялов О.В. 1978 . Некоторые орнитологические находки в Заилийском Алатау // Биология птиц в Казахстане. Алма-Ата: 115-119 (Тр. Ин-та зоол. АНКазССР. Т. 38).
Косенко С.М., Кайгородова Е.Ю. 2003 . Особенности экологии среднего пёстрого дятла в Деснянском Полесье // Орнитология 30: 94-105.
Костин Ю.В. 1983 . Птицы Крыма. М.: 1-240.
Лес России: энциклопедия. 1995. М.: 1-447.
Маклашова Т.Г., Смирнов О.П. 1979. Об удачном размножении больших пёстрых дятлов в неволе // Экология гнездования птиц и методы её изучения. Тез. конф. молодых учёных. Самарканд: 151-153.
Мальчевский А.С., Пукинский Ю.Б. 1983. Птицы Ленинградской области и сопредельных территорий: История, биология, охрана. Л. 1: 1-480.
Марисова И.В., Бутенко А.Г. 2013. Материалы к распространению и экологии сирийского дятла Dendrocopos syriacus на Украине // Рус. орнитол. журн. 22 (871): 1069-1074.
Митрофанов П.Н., Гавлюк Э.В. 1976. К вопросу о биологии и поведении большого пёстрого дятла // Биология питания, развития и поведения птиц. Л.: 101-111.
Митяй И.С. 1983. Расширение ареала чёрного дятла на Украине // Вестн. зоол. 4: 86-87.
Митяй И.С. 1985. Дятловые Приднепровской лесостепи. Автореф. дис. … канд. биол. наук. Киев: 1-26.
Митяй И.С. 2009. Дятлы лесостепи бассейна Днепра // Рус. орнитол. журн. 18 (499): 1255-1260.
Новиков Г.А., Мальчевский А.С., Овчинникова Н.П., Иванова Н.С. 1963. Птицы «Леса на Ворскле» и его окрестностей // Вопросы экологии и биоценол. 8: 9-118.
Осмоловская В.И., Формозов А.Н. 2009. Очерки экологии некоторых полезных птиц леса: Дятлы // Рус. орнитол. журн. 18 (476): 575-605.
Познанин Л.П. 1947. Приспособительные особенности строения скелета дятлов // Докл. АН СССР 58, 4: 713-715.
Поливанов В.М. 1981. Экология птиц-дуплогнездников Приморья. М.: 1-171.
Прокопов А.С. 2011. Экология размножения дятловых равнинной тайги Западной Сибири // Рус. орнитол. журн. 20 (698): 2102-2103.
Птушенко Е.С., Иноземцев А.А. 1968. Биология и хозяйственное значение птиц Московской области и сопредельных территорий. М.: 1-461.
Раздорский В.Ф. 1955 . Архитектоника растений. М.: 1-431.
Сагитов А.К., Рустамов М. 1974. Новые данные об экологии гнездования горного белокрылого дятла (Dendrocopos leucopterus leptorhynchus) // Материалы 6-й Всесоюз. орнитол. конф. М., 2: 124-125.
Салихбаев Х.С. 1956. Позвоночные животные лесных насаждений Узбекистана // Тр. Инта зоол. и паразитол. АН УзССР 8: 131-193.
Севастьянов Г.Н. 1962. О распределении гнёзд-дупел в Архангельской тайге // Зоол.журн. 38, 4: 589-595.
Севастьянов Г.Н. 1964. Распределение и основные черты биологии хозяйственно ценных птиц в лесах Европейского Центра. Автореф. дис. … канд. биол. наук. М.: 1-19.
Симкин Г.Н. 1977. Групповое поселение большого пёстрого дятла // Орнитология 13: 134-145.
Фетисов С.А. 2017. О гнездовых дуплах и эволюции гнездостроения у дятлов Picidae: Что считать гнездом у этих птиц? // Рус. орнитол. журн. 26 (1405): 575-590.
Фёдоров В.А. 2011 . Случай повторного гнездования белоспинного дятла Dendrocopos leucotos в Юнтоловском заказнике (Санкт-Петербург) // Рус. орнитол. журн. 20 (656): 949-952.
Формозов А.Н. 1974 . Спутник следопыта. М.: 1-319.
Чавчавадзе Е.С. 1979. Древесина хвойных. Л.: 1-190.
Ширская М.Н. 1938. Возникновение сердцевидной гнили осины // Лесное хоз-во (В защиту леса). М., 1: 47-49.
Bent A.C. 1939. Life histories of North American woodpeckers. Washington: 1-334. (Bull.Smithsonian Inst. 174.).
Blume D. 1968. Die Buntspechte. Wittenberg-Lutterstadt: 1-112.
Cramp S. (Ed.) 1985. The Birds of the Western Palearctic. Oxford Univ. Press, 4: 1-960.
Zhao Dan Zhu, Wei Zhang, Cheng Wei Wu. 2014 . Energy conversion in woodpecker on successive peckings and its role on anti-chok protection of brain // China Tech. Sci. 57, 7: 1269-1275.
Русский орнитологический журнал 2017, Том 26, Экспресс-выпуск 1444: 1966-1967
Примечания
1С другой стороны, клюв дятла – более совершенное орудие, чем долото, как это обычно принято считать в литературе, и не имеет аналога с инструментами человека. В частности, у долбящих дятлов затылочное отверстие сдвинуто вниз, отчего их голова и шея приближаются по конструкции к молотку, частью которого служит и расширенный клюв, а череп не только укрепляется в целом, но упрочняются и его соединения с клювом (Познанин 1947). Помимо того, клюв может работать одновременно как пинцет или щипцы. Поэтому он более эффективен, чем простое долото, тем более при использовании его в таком тесном и замкнутом пространстве, какое представляет собой выдолбленное лишь наполовину гнездовое дупло.
2Именно осина чаще других подвержена возникновению в ней сердцевидной гнили (Ширская 1938), и этим объясняется её повышенная дуплистость.
3По наблюдениям в Крыму (Костин 1983), большой пёстрый дятел и в наше время часто устраивает своё дупло в твёрдых породах деревьев на месте выгнившего сука и расширяя уже существующее дупло. Белокрылый дятел Dendrocopos leucopterus также использует уже имеющиеся естественные дупла, выдалбливая в них лишь нужное им входное отверстие и слегка расширяя дно дупла (Салихбаев 1956; Сагитов, Рустамов 1974).
4Как видно на рисунке 12е, леток в дупле желны не всегда может иметь строго прямоугольную форму, хотя этот отличительный признак от летков всех других видов дятлов приводится во многих определителях.
Рекомендуем прочесть
 Про философию биологии
Про философию биологии Ещё раз об инстинктах
Ещё раз об инстинктах Ритуализация «по Благосклонову» и её сопряжение с «тинбергеновской»
Ритуализация «по Благосклонову» и её сопряжение с «тинбергеновской» Что не так с социобиологией?
Что не так с социобиологией? «Город как заказник»: (пока?) упускаемая возможность
«Город как заказник»: (пока?) упускаемая возможность Так кто всё-таки сэкономил на мозгах?
Так кто всё-таки сэкономил на мозгах? Болит ли голова у дятла?
Болит ли голова у дятла? Большой пёстрый дятел: зачем всю зиму охранять территории?
Большой пёстрый дятел: зачем всю зиму охранять территории?
