

Когда прошло первое десятилетие XXI века, уважаемый ![]() nature_wonder “попросил некоторых ученых, имеющих lj, задуматься о десятилетии предстоящем (до 2020 года). И задал им следующий вопрос: какая тенденция, которая еще не всем видна, будет увеличивать свое влияние в ближайшие 10 лет? Это может быть другой способ думать об определенных вещах, какой-то метод работы, подход к проблеме, взгляд на действительность, другая оценка известного/неизвестного, уже существующая теория и т.д. Иными словами, что, на их взгляд, уже начинает меняться и в ближайшие годы станет заметным трендом в их области”. И опубликовал у себя их прогнозы.
nature_wonder “попросил некоторых ученых, имеющих lj, задуматься о десятилетии предстоящем (до 2020 года). И задал им следующий вопрос: какая тенденция, которая еще не всем видна, будет увеличивать свое влияние в ближайшие 10 лет? Это может быть другой способ думать об определенных вещах, какой-то метод работы, подход к проблеме, взгляд на действительность, другая оценка известного/неизвестного, уже существующая теория и т.д. Иными словами, что, на их взгляд, уже начинает меняться и в ближайшие годы станет заметным трендом в их области”. И опубликовал у себя их прогнозы.
Я, по склонности к многописанию и любви к подробностям, наваял много больше, чем нужно (тем более что писал во время каникул, на наблюдениях в Подмосковье, по вечерам больше заняться было нечем, а нестабильно работающий Интернет пресёк возможность втягивания в ЖЖ-дискуссии). Так что выкладываю полный текст; пп.1-7 касаются биологии, п.8 – гуманитарных наук и общественной жизни.
Надо сказать, что в прогнозах я максимально оптимистичен. Я всегда плохо умел оценивать «сопротивление материала и вязкость среды», поскольку как-то всегда полагал, что полагая некоторую идею хорошей, надо просто действовать в её пользу, честно и изо всех сил, а там уж как сложится.
————————————————————-
В общем случае можно сказать, что «идеи, которые набирают силу», связаны с поиском альтернатив в решении тех до сих пор не решённых общебиологических проблем, которые уже 30-40 лет как пытаются решить при помощи, условно говоря, «программы Гамильтона-Вилсона-Докинза» — радикального редукционизма, пытающегося понять разные эволюционные феномены (как возможны групповые адаптации? устойчивые системы вроде организма и популяции? и т.п.) «снизу вверх», через отбор «эгоистических» и «независимых» элементов, которые могут быть «склеены» им в некое общее целое, но каким-то метафизическим образом сохраняют эту самую «эгоистичность». Если в онтологии это радикальный редукционизм, то в методологии – радикальный индивидуализм. Эта естественнонаучная позиция имеет хорошее отображение в тех гуманитарных дисциплинах, которые имеют дело с поведением и взаимодействием людей, именно в социологии и в экономике, где получила преобладание примерно в эти же годы, и точно также исчерпывается, вызывает неудовлетворённость сейчас.
Сейчас уже понятно, что умножение исследований в рамках этой программы вместо ожидаемого «всеобщего ответа» принесло только множество «частных опровержений теории» и умножение объяснений ad hoc, вроде родственного отбора, от которых через некоторое время отказываются даже их творцы. См. «Kin selection died?» и С.Линке «Была ли социобиология «новым синтезом»?
На фоне растущего понимания, что соответствующая исследовательская программа не оправдала надежд, которые некогда на неё возлагались, и одновременного роста активности её сторонников, притязающих на всеобщность и единственность такого подхода, потихонечку будет расти и усиливаться противоположная идея целостности. Она
а) связана с натуралистическим пониманием структуры, функции и эволюции биосистем,
б) требует, чтобы субъекты эволюционного процесса (виды, популяции и т.п. системы меняющиеся во времени и приспосабливающиеся к среде обитания) прежде чем исследовались, были бы хорошо выделенными системами, с наличествующими механизмами внутренней связи и регуляции, превозмогающими «независимость-эгоистичность» поведения элементов или способными направлять её к нужному результату, то есть «морфологический подход» с систематикой здесь предшествуют эволюционным, популяционным или поведенческим исследованиям, задают им объекты и процессы, требующие причинного объяснения, а не пытаются их «увидеть» на основе выводов высокой теории;
в) основана на мысли о большей продуктивности рассмотрения эволюции «сверху вниз», от популяционных систем, обладающих целостностью и способных регулировать как перемещения и взаимодействия особей, так и распределение собственного генного пула между локальными группировками в целях достижения общей, а не локальной адаптации;
— на большей важности идеальных регуляторов перемещений и взаимодействий элементов системы надындивидуального уровня, будь то вид, популяция, онтогенез, создающий видовой морфотип, по сравнению с биолого-генетическими особенностями самих индивидов;
— на примате реляционных характеристик перед атомарными в анализе процессов отбора (который всегда остаётся индивидуальным, и никогда групповым);
— на идее сортировки особей по потенциям внутри популяционной системы за счёт внутренних регуляторных механизмов последней, увеличивающих «точность попадания» разных особей именно в ту среду, которая наиболее благоприятна для реализации их специфических поведенческих иных характеристик, как при тех объяснениях, что господствуют ныне, вместо нынешнего примата селекции особей по признакам, уже дефинитивным (1 и 2, PS). За счёт этого локальная адаптация сопрягается с ростом общей устойчивости системы, а не противоречит ему.
Иными словами, идеи целостности и системоцентризма (концентрирующегося на популяционной регуляции в понимании вида и на онтогенетической регуляции — в понимании биологической формы) будут усиливаться и умалять нынешнее торжество радикального редукционизма и методологического индивидуализма.
В частности, можно надеяться, что она позволит решить те нерешённые проблемы теории эволюции, о которых я писал ранее — или хотя бы понять в какой области стоит искать решение, в том числе позволит разобраться в соотношении селекционных и номогенетических процессов в эволюции разных групп (уверен, что оно будет неуниверсальным). Далее пп.1-8 конкретизируют им раскрывают пп. а)-в), 1-7 для биологии, 8 для гуманитарных наук, имеющих дело с человеческим поведением и взаимодействиями людей (социология, психология, экономика). Всё это даёт надежду на решение тех самых «нерешённых проблем эволюционной теории», про которые мы с marina_fr писали когда-то. В нынешней ситуации очевиден застой и/или топтание на месте; может, смена парадигмы поможет?
1. Идея радикального редукционизма в этологии, популяционной и эволюционной биологии, будучи радикально высказанной после короткого периода торжества начинает отрицать самоё себя. Это видно уже по попыткам отказаться от отдельных частных концепций, вроде родственного отбора, которые явно не подтверждаются фактами. Не выдержали они и притязаний на всеобщность.
Скажем, явно не оправдываются утверждения Бэйтмена и Трайверса об универсальности различий инвестирования разных полов в половое поведение и заботу о потомстве, на котором покоятся господствующие сейчас представления о половом отборе. Какой пол инвестирует больше, какой меньше (и соответственно, самки конкурируют за внимание самцов, а самцы выбирают, или наоборот), определяется конкретной «конструкцией» социальной системы разных видов и филогенетической историей группы, определяющей не общие, а локальные причины перехода от одной «конструкции» к другой. См. пример про брачное доминирование у разных видов пёстрых дятлов: у видов с доминированием самца (большой, сирийский, и т.д.) в период весеннего возбуждения и образования пар самцы конкурируют за внимание самки, а она «натравливает» их друг на друга, если пара уже образовалась, если нет – спокойно ждёт исхода их борьбы. У видов с доминированием самок (белоспинный, трёхпалый, в меньшей степени малый пёстрый) всё наоборот – самки ожесточённо конкурируют за внимание самцов, которые спокойно ждут исхода борьба. При столь резких различиях соотношение полов среди той части популяции, что активно участвует в занятии территории и токовании на них, т.е. в конкуренции за партнёра, во всех случаях 1:1 и нормой отношений является моногамия с заботой о потомстве обоих родителей. Отклонения в сторону сдвоенного гнездования, классической полиандрии или полигинии встречаются только в исключительных случаях и либо в слишком поздних кладках, либо при сильных демографических нарушениях.
Таких фактов много, и их обобщение заставляет Roughgarden et al. (2006) отказываться от идеи полового отбора в пользу более общей идеи отбора социального.
![The types of ornaments and weapons produced by sexual selection (top row) are also produced by non-sexual social selection (bottom row). The red epaulettes of red-winged blackbirds (Agelaius phoeniceus, top left) are used in territorial defence during the breeding season [32], resulting in enhanced mating success; the colourful crowns of golden-crowned sparrows (Zonotrichia atricapilla, bottom left) help win contests over food in winter [33], and winter social dominance correlates with enhanced survival in other bird species. Bird beaks are used as weapons by many birds to defend territories that serve to attract mates, including lark buntings (Calamospiza melanocorys, top centre) [34]; beaks are also used as weapons by offspring in species such as laughing kookaburras (Dacelo novaeguineae, bottom centre; photo Sarah Legge), wherein siblicide increases food resources for the aggressor, resulting in improved survival [35]. Horns and antlers are favoured by sexual selection in male insects and mammals such as elk (Cervus canadensis, top right), where they function as weapons used in fights over females [36]; female beetles (Onthophagus sagittarius) use their horns to fight over resources their offspring require [37] (bottom right; photo Sean Stankowski). All photos by Bruce Lyon unless credited otherwise.](http://www.socialcompas.com/wp-content/uploads/2015/01/F3.large_.jpg)
The types of ornaments and weapons produced by sexual selection (top row) are also produced by non-sexual social selection (bottom row). The red epaulettes of red-winged blackbirds (Agelaius phoeniceus, top left) are used in territorial defence during the breeding season [32], resulting in enhanced mating success; the colourful crowns of golden-crowned sparrows (Zonotrichia atricapilla, bottom left) help win contests over food in winter [33], and winter social dominance correlates with enhanced survival in other bird species. Bird beaks are used as weapons by many birds to defend territories that serve to attract mates, including lark buntings (Calamospiza melanocorys, top centre) [34]; beaks are also used as weapons by offspring in species such as laughing kookaburras (Dacelo novaeguineae, bottom centre; photo Sarah Legge), wherein siblicide increases food resources for the aggressor, resulting in improved survival [35]. Horns and antlers are favoured by sexual selection in male insects and mammals such as elk (Cervus canadensis, top right), where they function as weapons used in fights over females [36]; female beetles (Onthophagus sagittarius) use their horns to fight over resources their offspring require [37] (bottom right; photo Sean Stankowski). All photos by Bruce Lyon unless credited otherwise.
То есть, огрублённо, в зависимости от своих задач, требующих производства тех или иных генных продуктов с тем или иным напряжением, организм «выбирает сам», какие генетические тексты «считать» отдельностью, с какой интенсивность и «тщанием» их прочитывать, что делается включением/выключением регуляторов генных сетей. И магистральное направление эволюции на молекулярном уровне – это отнюдь не отбор на уровне самих генов, а подбор оптимальной комбинации регуляторов под разные задачи, или перестройка системы регуляторов в связи со сменой задач, как это хорошо аргументируют Суслов и Колчанов. Отсюда следует, что изменения на уровне организмов через «эгоизм» отдельных генов и отбор невозможны даже в теории, как полезная фикция — просто потому, что соответствующие регуляторы направлены только в одну сторону, от организма к генам, но не наоборот, как стрелочки направления матричного копирования в Центральной догме молекулярной биологии.
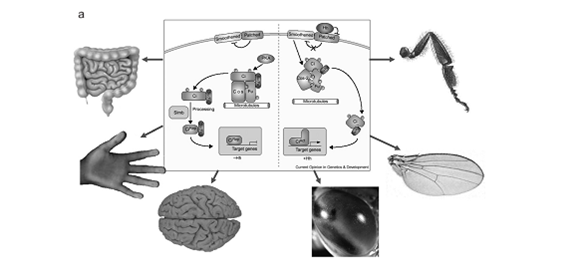
Примеры трех генных сетей, гены которых формируют консервативные паттерны коэкспрессии у разных таксонов – регулоги: а – генная сеть Hh-каскада (по: Nybakken, Perrimon, 2002); б – генная сеть циркадного ритма (по: Badiu, 2003); в – генная сеть морфогенеза цветка (по: Honma, Goto, 2001; Theissen, Saedler, 2001).
Следовательно, мы возвращаемся к старой идее генов-модификаторов и отбора классического эволюционизма (для которой центральным был отбор фенотипов, да не независимых, а связанных взаимодействиями внутри популяции в целостную систему, а не на уровне генов), но на «повышенном основании». В чём «повышенном»? Благодаря 40 лет развития «программы Гамильтона-Вилсона- Докинза» мы знаем, как «эгоистичные» мутации или морфологические изменения могут «рушить и портить» групповые адаптации, вроде «уменьшения честности» видовых сигналов и снижения устойчивости популяционной структуры, уменьшения связанности индивидов социальными связями. Но постепенно всё больше становиться понятно, что никаким сочетанием таких «эгоизмов» групповой адаптации не получишь – социальность возникает лишь развитием предшествующей, менее развитой социальности, то же относится и к популяционной структуре и т.п. Типа целое только от целого.

Далее, сейчас всё больше проясняется, что центральное для этой программы понятие итоговой приспособленности неадекватно, ибо единицей, учитываемой отбором, являются не «эгоистичные гены», а организм как целое. Последний умеет управлять своими генами ничуть не хуже, чем мышцами и печенью, хотя и при помощи несколько иных сигналов – поведенческих стимулов, опосредованных каскадом изменений в переменных состояния, заданных концентрациями гормонов, нейромедиаторов и т.п. Также как подразделённая популяция управляет своим генофондом, изменяя концентрации разных генотипов в разных локальных группировках так, чтобы максимизировать устойчивость системы в целом и «точность воспроизводства» специфической структуры системы, что хорошо показано в работах Ю.П.Алтухова с учениками.
Далее, ген, в отличие от организма, не является дискретной единицей, тождественной самой себе, в разные моменты своего существования их нельзя выделять как отдельности, согласно обычным классификационным процедурам биологической систематики. А виды и отдельные особи можно – в силу известного факта эквифинальности онтогенеза и наличия механизмов, поддержания целостности, обеспечивающих устойчивость и дискретность дикого типа, у генов же ничего такого нет и в помине.
Лучше всего эту мысль пояснить это на примере идеи мема, который слеплен «по образу и подобию» «эгоистичного гена» социобиологов. Почему всякий мем – это не отдельность, а условность? Потому что в культуре нет «естественных границ», которые бы делили её на элементы, этаких «швов» или «надрезов». Кто какие элементы выделит в какой ситуации – это определяется позицией (классовой, идеологической и т.п.) выделяющего. То же самое относится и к устойчивости/бесперебойности/помехозащищённости последующих процессов социальной трансляции этих элементов, о чём убедительно писал М.К.Петров.
Соответственно, говорить о мемах без указания «позиции» выделяющего бессмысленно. Разные группы с разных позиций выделяет и будет воспроизводить разные наборы элементов и будут бороться друг с другом за «каналы воспроизводства», чтобы пропустить в будущее ключевые символы собственной идеологии, но не пропустить идеологию оппонента. Сейчас постепенно становится понятно, что то же самое верно и в отношении генов.
2. Поскольку «программа Гамильтона-Вилсона-Докинза» интеллектуально привлекательна – стройна, логична, красива, последовательна, как всякая замкнутая концепция, её легко принять и отказываться от неё не хочется, то приведение её в соответствие с новыми данными, на которые я ссылаюсь по тексту, позволит понять её действительно полезную роль – не онтологии и не теории, а аналитического инструмента и нулевой гипотезы.
То есть если задать себе вопрос: откуда столь высокая популярность идей «эгоистичных генов» и «отбора генов», высказанных людьми, которые никогда исследований генов не проводили (в то время как проводивший исследования отбора и динамики генофондов в популяциях Алтухов придерживается идей противоположного рода), почему многие биологи так держатся за их концепцию, несмотря на множество частных опровержений, то единственный разумный ответ будет то, что они предложили хорошую методологическую идею.
Соответственно, надо продумать как следует, в чём действительно она состоит, и продумав, сменить способ её использования – не как рабочей гипотезы, которая должна подтверждаться в исследованиях, а как гипотезы нулевой, которую надо опровергать, чтобы убедиться в том, что перед нами «хорошо выделенная система», с которой происходит нечто существенное в плане функционирования и/или эволюции.
Действительно, если радикальный редукционизм-индивидуализм из гносеологии перенести в онтологию, то его честное и последовательное применение соответствующих идей можно ликвидирует сами объекты изучения социологии, с одной стороны и биологии, с другой: общество в первом случае и индивид как объект отбора + вид (популяционная система) как субъект эволюционного процесса во втором. Что выглядит более чем странным, ведь всякое исследование социальной истории или биологической эволюции начинается с того, что «натуралисты», относящиеся к соответствующим дисциплинам (историки/антропологи в первом случае и натуралисты-систематики во втором) должны соответствующие объекты «открыть в природе». То есть увидеть, выделить и отделить от других объектов того же ряда, поставить в исторический и средовой контекст, иными словами, произвести классификацию.
И только потом приходят теоретики — социологи или эволюционисты — и начинают интерпретировать, какого рода факторы и механизмы были существенны для данного процесса развития. И, если теоретически одарены, сравнив свой результат с данными других авторов, могут сделать интересный скачок в сторону того, какие факторы и механизмы существенны для биологической эволюции/социальной истории вообще.
То есть «программа Гамильтона-Вилсона-Докинза» (с присовокуплением Мэйнарда Смита с Трайверсом) не исчезнет совсем, но сохранится как полезный приём – или умственный эксперимент, — позволяющий допущением предельной «эгоистичности частей» «проверить» соответствующую биосистему «на излом»? Сохранит ли она хороший устойчивость и целостности при такой интенсивности «обмана» и такой концентрации «обманщиков», справятся ли с ними существующие механизмы регуляции и контроля? То есть нечто вроде крэш-теста на тему, выживет ли эволюционирующая система в процессе эволюции, не утратит ли самотождественности в каждый данный момент и способности к преемственным изменениям, без которых эволюция невозможна. А вот направления и скорость эволюционирования будут определяться существенно другими причинами, — именно теми, которые приверженцы этой программы просто отказываются рассматривать как феномены, считая эпифеноменами, в то время как скорей всего всё точно наоборот.
3. Будет набирать силу «морфологический подход» к стохастическими системам, прежде всего к популяциям, социуму и виду как системе популяций, определённым образом структурированной в пространстве ареала и отграниченной от аналогичных популяционных систем, составляющих другие виды. Это даст новую жизнь исследованиям процессов регуляции, так что мы сможем больше узнать как образом система управляет своими элементами — вроде бы независимыми индивидами, чтобы иметь возможность устойчиво воспроизводить присущий ей специфический паттерн системной структуры вопреки возмущениям извне и напряжениям изнутри. Это позволит распространить на популяцию и социум те представления о структуре, архетипе, форме и гомологии, которые оказались исключительно плодотворны для идеалистической морфологии. До сих пор популяция априори полагается «плохо выделенной системой», то есть по умолчанию предполагается, что:
— особи в популяции при расселении перемещаются стохастически, ну, в крайнем случае следуют направляющим линиям экологического характера,
— популяция структурируется только физическими барьерами или границами подходящих местообитаний, поэтому популяционная структура вида, и в том числе пространственно-этологическая структура, не имеет собственной формы – «будет то, что получится» в стохастике перемещений и взаимодействий особей, движения которых мыслятся подобными траекториям молекул идеального газа, такая и получится. Соответственно, в однородной среде такая популяция не сможет реализовать никакой устойчивой и видоспецифичной структуры, а пространственные и социальные связи особей (скажем, моделируемые клеточными автоматами), образуют нечто вроде «морозных узоров», форма которых существенно зависит от начальных условий образования и уровня средовой нестабильности случайного характера.
В последние годы накапливается всё больше наблюдений, показывающих, что это идеализация не только не соответствует действительности, но, напротив, в природе всё точно наоборот:
— популяция разных видов обладают «морфологией», в том плане, что популяционная структура, реализуемая в гетерогенном пространстве ареала типологически определена. То естьобладает специфической формой отношений между особями-элементами – пространственных, социальных и биотопических, которая [форма] узнаваема и отличима от аналогичной формы отношений других видов, даже в случае, когда из-за высокого уровня нестабильности среды популяционная структура реализуется не вполне полно или не совсем точно. Здесь, здесь и здесь.
Иными словами, популяционная структура вида допускает такое же «морфологическое» описание и также требует применения сравнительного метода в анализе траекторий своего развития, как и анатомия представителей этого вида. Я даже думаю, что все утверждения эпигенетической концепции эволюции М.А.Шишкина – Шмальгаузена – Уоддингтона столь же приложимы к устойчивому воспроизводству «типа» популяционной структуры вида в пространстве ареала, вопреки демографическим флюктуация внутри и нестабильности среды вовне воспроизводящейся популяционной системы, как и к онтогенезам, воспроизводящим видовую норму в той же самой череде поколений.
— Популяция не ни исчерпывается стохастикой перемещений особей, ни мозаикой группировок (поселений, демов), «запертых» на «островах» подходящих местообитаний (поскольку все естественные местообитания всегда мозаичны, расселение всякого вида носит островной характер, и связано с «архипелагом» подходящих биотопов). Устойчивое воспроизводство видоспецифической структуры отношений придаёт популяции целостность и, действительно, разрозненные популяционные группировки объединены устойчивым потоком нерезидентных особей, постоянное «давление» которого
I. обеспечивает непрерывный отбор резидентов, так чтобы на разных «позициях» в социальной системе – и доминантов, и подчинённых и среднеранговых особей – с высокой вероятностью оказывались индивиды, по своим качествам наиболее соответствующие «позиции», а все прочие вытеснялись и в составе того же потока направленно перемещались в другие группировки – такие, где их индивидуальные качества будут лучше всего соответствовать социальной среде (см. про сортировку особей по потенциям между популяциями, из которой следуют групповые адаптации вида вцелом).
II. устанавливает границы между разными популяциями даже в однородной среде, позволяет популяции структурироваться в однородных местообитаниях, не имеющих физических барьеров, делает переселения особей между группировками, направленные на повышение приспособленности, детерминированным процессом, определенным образом ориентированным в пространстве ареала (которое в свою очередь, становится столь же анизотропным, как, например, яйцеклетка перед началом дробления или бластула в начале дифференциации, продолжая сравнение с онтогенезом). На эту тему см. работы Н.А.Щипанова по мелким млекопитающим.,
III. связывает в одно целое разные части популяции или разные географические популяции, обитающие в разнородных условиях, позволяет им сопряжённым образом реагировать на разнонаправленные средовые тренды в ареале каждой из них. Этим максимизируется устойчивость системного целого и устойчивость воспроизводства специфической пространственно-этологической структуры вида (за счёт некоторого снижения совершенства локальных адаптаций в каждой из частей последнего, не без того). Действительно, специально поставленные опыты в подразделённых ящичных популяциях дрозофилы, а затем и в природных подразделённых популяциях показали, что популяция как целое управляет перемещениями своих элементов – особей разных генотипов, ориентируя их таким образом, чтобы определённое (резко неравное) распределение носителей разных генотипов между разными группировками в подразделённой популяции было устойчивым, а не смазывалось. См. эту же статью Ю.П.Алтухова и его книгу «Генетические процессы в популяциях». М.: ИКЦ Академкнига, 2003.
То есть уже и сейчас новое понятно, надо лишь «иначе сложить кусочки головоломки», которых накоплено натуралистами уже более чем достаточно. А для этого надо менять концепцию, что, как думается, будет постепенно происходить по мере разочарования биологов в геноцентрической концепции эволюции и идее итоговой приспособленности как универсальной валюты для оценивания «платы и выигрыша» от разных «эволюционных решений». Хотя метод, предполагающий такую оценку и требующий её – он безусловно хорош и представляет собой одно из крупнейших достижений «программы Гамильтона-Вилсона-Докинза», в этологии уж как минимум.
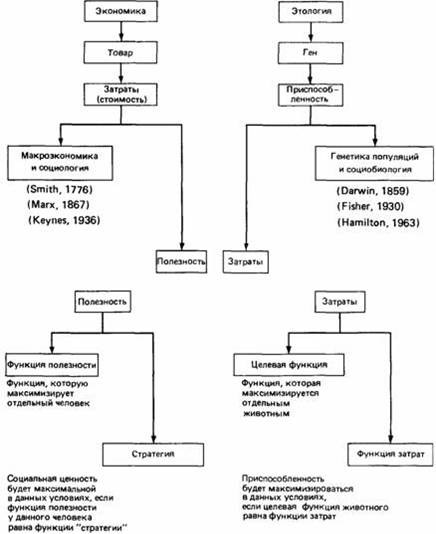
Параллель между понятиями экономики и этологии. Вверху. Общая схема, на которой указаны фамилии исследователей, внесших наибольший вклад в развитие этих областей науки, и даты публикации их основных работ. Внизу. Детализированная схема в приложении к отдельному человеку или животному. (McFarland, Houston, 1981.).
4. В связи с успехами «морфологического подхода» к популяционным системам можно предполагать ренессанс биологической концепции вида, которая нынче находится в сильном забвении. Но это отдельная необщая тема, которую дальше развивать здесь не стану.
5. Поскольку радикальный редукционизм не смог объяснить эволюционное происхождение групповых адаптаций (или его объяснение требует настолько большой «цены элиминации«, которую ни один вид не платит в реальности), а вроде бы оппонирующая концепция группового отбора явно неверна, можно ожидать усиления идеи сортировки особей по потенциям вместо отбора по признакам в популяции, понимаемой «морфологически», как струткурно дифференцированное целое п.3.
А это, в свою очередь, усилит позиции идеи, согласно которой более продуктивно рассматривать эволюцию, «сверху вниз»: от вида к популяциям, от популяций к особям и от особей – генам, нежели наоборот. И это понятно: если в «хорошо выделенной» системе целое управляет своими частями, и специфический паттерн системной структуры умеет воспроизводиться несмотря на «возмущения», вносимые независимостью элементов, стохастикой их перемещений внутри системы и т.п., ввиду того, что управляющие воздействия в системе направлены «сверху вниз», то и эволюция будет направлена точно также.
В силу такой асимметрии управления и регуляции между целым и элементами (или же между пространственными частями, вроде пространственных подразделений в популяции) селективные воздействия на уровне целого обязательно будут «передаваться» на уровень элементов, причём в усиленном и мультиплицированном виде, поскольку на это будут работать уже существующие механизмы поддержания целостности и внутри системной регуляции. Понятно, что это относится и к организму, и к популяции, и к онтогенезу, в процессе которого создаётся видовой морфотип. А вот селективные воздействия на уровне элементов вряд ли будут передаваться на уровень системного целого и вызывать изменения последнего, поскольку уже существующие системы поддержания целостности будут «работать» против соответствующих изменений, гасить и нейтрализовывать их; в крайнем случае, система погибнет, но не изменится – а именно направленные и необратимые изменения составляют те элементарные эволюционные явления, разные причинные объяснения которых мы стараемся предложить, оттеснив конкурирующие.
То есть из всего того, что там известно на тему, как популяция управляет перемещением и взаимодействием особей, организм – своими генами, а онтогенез – становлением видовой морфологии через взаимодействие частей и взаимную регуляцию клеточных пластов, потихоньку становится ясно, что
— изменения на уровне популяционной структуры или системы взаимодействий между особями в популяции, передаваясь на уровень особей, «тянут за собой» вполне эволюционные изменения сперва в физиологии, а затем и в морфологии индивидов.
— изменения на уровне элементов системы, если они увеличивают приспособленность в каких-то частных обстоятельствах биологии вида, включаются в структуру нормы, увеличивая скрытый фонд разнообразия, или же просто нейтрализуются.
То есть в силу вышесказанного эволюцию гораздо продуктивней понимать «сверху вниз», в направлении уже существующих управляющих воздействий от целого к элементам системы, чем в направлении противоположном. Тем более что смысл эволюции мы видим и в направленном преобразовании этих систем (филетическая эволюция), и в их разделении (диверегенция) и во всё большем обособлении друг от друга при увеличении их целостности/управляемости внутри себя (прогресс), а набор элементов с их свойствами и «валентностями» остаётся примерно тем же (гены, индивиды и т.п.).
Думаю, что это понимание будет постепенно усиливаться, проясняться и конкретизироваться. Критическим здесь, наверное, является соотношение наследуемости и управляемости, в тех случаях когда первая и вторая находятся не в гармонии, а в контрапункте (и надо сказать, что таких случаев большинство). Если управляемость поведения особей (их перемещений и взаимодействий друг с другом) регуляторными взаимодействиями со стороны социума и популяции значимо выше, чем генетическая детерминация структур поведения, реализуемых и используемых в соответствующих перемещениях и взаимодействиях, вышеописанное рассмотрение будет более продуктивно, чем противоположное ему.
Если же наоборот, популяция или социум будут «плохо выделенной системой», и их рассмотрение будет неинтересно даже как полезной фикции, нужно будет рассматривать просто «россыпь особей», взятых исследователем в некой выборке, и даже не думать, что это отображение некоторого системного единства. Но я полагаю, что таких случаев не найдётся вовсе: как люди не независимы, а живут в обществе, и конкурируют не друг с другом, а за «улучшение позиции» в существующей социальной структуре, «места в которой» с их требованиями к человеку в известном смысле реальней самих человеческих качеств и способностей – настолько, что последние можно считать не существующими «в себе», так особи живут в популяциях. Поэтому последние эволюционируют не независимо, а связанно друг с другом, так что все изменения, производимые отбором, передаются по внутрисистемным связям и дают мультиплицирующий эффект.
6. Будет дальше развиваться и получать новые подтверждения крайне продуктивная идея, что наблюдаемая поливариантность жизненных циклов у разных видов позвоночных (а может быть, и беспозвоночных) животных есть результат трейд-оффа между размножением и самосохранением (телесным ростом). Первое рассчитано на большой выигрыш, но связано со столь же большим риском снижения витальности (через снижение иммунитета в связи с брачной активностью, и сугубым воздействием специфических рисков последней). См. пару интересных статей М.П.Мошкина про сабж, 1 и 2.
Соответственно, искомые «регуляторы оптимального соотношения» между первым и вторым устанавливаются на уровне организма (он выбирает), которые направляются и «поправляются» воздействиями внутрипопуляционного характера, связанные с социальными отношениями внутри группировок, и «давлением» потока нерезидентов между группировками, связывающими их в целостную систему и только и ищущими возможности занять территорию/приобрести партнёра, вытеснив резидентов с соответствующих вакансий. Последнее создаёт мощное внутрисистемное давление отбора, превосходящее эффект аналогичной конкуренции в собственно группировках. Т.е. усиление этой идеи, исключительно интересной самой по себе, будет «работать» на усиление более общей идеи целостности, также как прочие частные примеры.
7. Потихоньку станет всё более ясно, что в передаче управляющих воздействий от системного целого к элементам, благодаря чему последние перестают быть «независимыми-эгоистичными», а их перемещения и взаимодействия делаются регулируемыми и направленными, важнейшую роль играет идеальное, объективно существующее в природе (1 и 2). Собственно, без последнего подобного рода системная регуляция представляется совсем невозможной, и это понимание, думаю, будет набирать силу. В популяционной системе (или в социуме) это «идеальные формы» видовых сигналов или «идеальная структура» отношений внутри видоспецифической социальной организации. В наблюдениях этологов много раз показано, что конкретные телодвижения возбуждённых особей при взаимодействии «должны укладываться» в набор соответствующих идеальных форм (с соответствующими изменениями «формы» телодвижения сообразно изменениям ситуации), чтобы быть эффективными – в смысле, устойчиво разделять исходно «равноправных» особей на выигравшего и проигравшего с соответствующей дифференциацией ролей и «общепризнанностью» возникшей асимметрии для прочих членов социума. Нарушения такого соответствия ведут к срыву взаимодействия и к потере всех уже сделанных затрат времени и энергии на него у обоих особей. То же самое верно и для «идеальной структуры» отношений между особями в социуме: она относится к видовому уровню, индивиды ей «только подчиняются», и по отношению к конкретным результатам взаимодействий отдельных индивидов это идеальное выступает как «реальность высшего порядка», успешно подчиняющая их себе, но не наоборот.
К тому же кругу идей относится понимание генетического кода как своего рода языка, использование которого позволяет «компетентному читателю» генетических «текстов» строить дефинитивный фенотип, которое было развито Б.М.Медниковым. Думаю, оно тоже будет получать всё большее распространение и популярность.
Наконец, если на секунду вернуться к взаимодействиям особей в популяции, к социальному поведению животных в сообществах, то когда мы видим прогресс внутренней регуляции и в той и в другой системе, мы всегда сталкиваемся с семантизацией телодвижений, с превращением действий, составляющих демонстрацию, из стимула в знак, в сигнальном эффекте демонстраций повышается роль передачи информации идеального характера, реакция на которую предполагает свободу выбора, и падает роль «принудительного» воздействия стимулов и т.п.
Про это подробнее см. “Коммуникация животных: от стимула к символу”. Это именно те процессы, которые этологи называют ритуализацией и эмансипацией сигнала от мотивационной подосновы; легко видеть, что это «прогресс идеального» в соответствующих системах –
а) увеличение отграниченности форм, информации и значения от чисто материальных событий, действий и воздействий индивидов,
б) увеличение подчинённости телодвижений, производящих знак (или взаимодействий, воспроизводящей отношения) задаче воспроизводства соответствующих «идеальных форм» и ничему более,
в) нарастание роли идеального как регулятора в соответствующей системе, прекращающего стохастику / независимость элементов в соответствующей системе, и претворяющего энергию последнего в кооперативный эффект.
8. Пара слов о небиологических «идеях, набирающих силу». Думаю, всякое существенное изменение представлений о биологической эволюции и других общебиологических концепций имеет свой отклик в социальной жизни – и в идеологиях, и в социальных практиках. Поскольку в современную эпоху идеология всё больше «онаучивается» — чтобы убеждать, должна меньше взывать к эмоциям, больше прибегать к рациональному доказательству, уметь убеждать что «новости с переднего фронта науки на её стороне», то есть эволюционировать от слепой веры к научному мировоззрению.
«Программе Вилсона-Докинза» в биологии в экономике соответствовали неолиберализм, а в подходе к человеку – геноцентризм и биологизаторство. То есть по мере распространения соответствующих идей среди «образованной публики» подавляющее большинство человеческих качеств и способностей считались врождёнными, биологически обусловленными хотя бы в своей основе. Соответственно, главной задачей педагогики, психологии, образования и воспитания, которые до этого считались Bildung, человекоделанием, вылепливанием личности из биологического организма, в последние 30-40 лет стали считаться тестирование и оценка для удобства селекции.
По мере отказа от этой исследовательской программы в пользу противоположной идеи целостности-системоцентризма будет распространяться противоположная идея социальной обусловленности и социальной зависимости человеческих способностей и человеческих качеств. Согласно этой точке зрения «биологическое», «гены» представляет лишь «сырьё», субстрат для развития, некоторое количество структур поведения и морфоструктур, как таёжный охотник оставляет продукты, соль и спички в избушке тем, кто придёт вслед за ним. Но вот совершенству использования этих структур для разрешения разных ситуаций – интеллектуальных, социальных, личностных (как «складывать» из них инструменты социального труда? Чтобы они не только были действенными, но развивали тебя самого) люди учатся у родителей, учителей, друзей, окружения и прочей социальной среды.
С т.з. последней человеческая личность – это не более чем точка пересечения общественных отношений и взаимодействий, формируемая ими как в колонии сифонофор столоны, появляющиеся в разных местах, закономерно формируются в разных специализированных зооидов. Тем более что сейчас, с открытием зеркальных нейронов, уже понятна та психобиологическая инфраструктура, которая опосредует социальные влияния и через которую социальное формирует личность с её качествами, то есть, по выражению Иоахима Бауэра «делает из психологии биологию». См. подробнее 1 и 2.
То есть нас ждёт возвращение педагогики с психологией, обучения и воспитания на прежнее место «делателей человеков», но уже на повышенном основании, обогащённом знанием многих новых биолого-генетических влияний – которые будут понимать не как формирующие и детерминирующие факторы, а как факторы уязвимости или односторонности развития.
В экономике и социальной жизни, скорей всего, вызовет поворот от рыночного фундаментализма к социальному государству. Во-первых, окажут действия сами образовательные изменения: если отношение преподавателей, родителей, учителей к следующему поколению сдвинется к «селекционного» на «развивающее» (а значит, и отношение этого следующего поколения к себе, их требования к другим, самооценку). Во-вторых, подействуют те демографические изменения, которые активно идут сейчас и завершаться к тому времени. Коротко говоря, в «третьем мире» завершится первый демографический переход, в «первом мире» — второй. И по завершении того и другого на любом из уровней квалификации работников просто не будет тех лишни рабочих рук, которые можно будет выкинуть в число безработных, чтобы меньше платить тем, кому приняли.
То есть возможность экономических действий либеральным способом – увеличение прибыли за счёт селекции и элиминации, позволяющих сэкономить на зарплате работников – резко сократится в силу объективных демографических причин. Придётся получать дополнительный выигрыш за счёт повышения трудоспособности и квалификации работников за счёт всего общества (учить, лечить и переучивать их за госсчёт) и повышать их мотивацию тем, что они не только зарабатывают деньги для себя, но и делают полезное дело для общества, через ограничение права собственности и рабочий контроль.
То есть нарастание демографических, социальных (которые просматриваются в нынешних общеевропейских столкновениях по поводу прав профсоюзов, образования и неподконтрольности действий правительства при формальной демократии) и экологических проблем уже в 2030-2040 гг. не позволит сэкономить ни на рабсиле и на экологической компенсации разрушений, производимых во время хозяйственной деятельности. Первую придётся учить-лечить, а второе восстанавливать за госсчёт и это даст мощный стимул к обобществлению.
Тем более к этому времени уже станет явственно, что более либеральные общества, экономически выигрывают у более солидарных (и более традиционных обществ – социалистических-то уж почти не осталось), но проигрывают им чисто дарвиновскую конкуренцию. А это заставит первые «сравнять счёт» новой социализацией общества и государства, изменением в сторону «большей социалистичности», чтобы избежать «нового средневековья». Так после Первой мировой войны в экономику США, Англии, Франции, до этого полностью либеральную, вошли плановые начала, и это резко повысило уровень жизни, физическое состояние, и т.п. характеристики простонародья (см. подробное описание у Э.Хобсбаума в “Эпохе крайностей. Короткий ХХ век”. Постепенное становление социального государства в 1935-1950-х гг. было следующей ступенью, сейчас появляется острая необходимость в третьей.
***
В заключение оговорюсь: писал не обо всех идеях из сферы компетенции, которые мне интересны и за которыми слежу, но лишь о тех, которые привлекают внимание, которые мы поддерживаем или отрицаем, которые связаны с развитием наших «любимых теорий». То есть мои собственные предпочтения задают ограниченность и пристрастность взгляда, но плюсов от честного указания собственной позиции всегда больше, поскольку такая позиция есть у всех, и она ограничена у всех примерно в одинаковой степени.
Рекомендуем прочесть
 Марихуановое наступление продолжается
Марихуановое наступление продолжается Как социальная помощь уходит самым богатым
Как социальная помощь уходит самым богатым Кыргызстан 2020: протесты рабочих и трудовые права
Кыргызстан 2020: протесты рабочих и трудовые права Синдром темпа жизни (pace-of-life syndrome, POLS): эволюция концепции
Синдром темпа жизни (pace-of-life syndrome, POLS): эволюция концепции Уроки сгоревшей Австралии
Уроки сгоревшей Австралии Экоцид в Амазонии: как Макрон и французские банки «спасали» леса Бразилии
Экоцид в Амазонии: как Макрон и французские банки «спасали» леса Бразилии Взрыв в Бейруте и «летящие гуси»
Взрыв в Бейруте и «летящие гуси» Заместитель Собянина по строительству Хуснуллин признался, что работает для застройщиков.
Заместитель Собянина по строительству Хуснуллин признался, что работает для застройщиков.
