

Бурлак С.А.
Аннотация. В статье суммируются данные, проливающие свет на время возникновения звучащей речи. Рассматриваются анатомические черты, с которыми различные исследователи связывали переход гоминид к членораздельной звучащей речи (опущенная гортань, величина базикраниального угла, толщина канала подъязычного нерва, форма и расположение подъязычной кости, подбородочный выступ, ширина позвоночного канала), и степень их релевантности для обоснования таких гипотез: наиболее релевантными оказываются низкое положение гортани (в сочетании со сравнительно небольшой длиной ротовой полости, что обеспечивает языку бóльшую подвижность в разных плоскостях и, тем самым, большие возможности для различения звуков при помощи артикуляции) и толщина позвоночного канала (свидетельствующая о расширении возможностей управления дыханием). Сопоставляются слуховые возможности как современных людей и человекообразных обезьян, так и ископаемых гоминид (по реконструкции на основе формы слуховых косточек), а также их возможности управлять дыханием (при помощи спинного мозга) и обрабатывать информацию (при помощи головного мозга). В обеспечении функционирования языка принимают участие не только зона Брока и зона Вернике, большое значение имеют лобные доли (в особенности, префронтальные формации коры).
В то же время само по себе увеличение объёма мозга в тех областях, где у современного человека находятся «речевые зоны», не является однозначным свидетельством в пользу наличия у соответствующего вида гоминид развитого звукового языка. Распознавание речи имеет комплексный характер, в нём участвуют мозговые структуры, распознающие не только отдельные смыслоразличительные признаки фонем, но и переходы от одной фонемы к другой, общий звуковой облик слова, языковой и внеязыковой контекст.
Рассматриваются также когнитивные приспособления, необходимые для формирования звучащей речи: способность к самоподдерживающемуся звукоподражанию, желание слышать понимаемую речь и интерпретировать звуки, произносимые окружающими, как знаки. Формирование всего этого комплекса признаков происходит лишь в гоминидной линии и не может быть вызвано одиночной макромутацией.
Всесторонний анализ данных позволяет сделать вывод, что звучащей речью обладал уже Homo heidelbergensis: у него отсутствуют горловые мешки (сильно уменьшающие влияние артикуляции на акустические характеристики звука), слух постепенно настраивается на лучшее распознавание тех частот, которые могут быть усилены или ослаблены при помощи артикуляции; у него же увеличивается толщина позвоночного канала. Но при этом нет оснований считать эту речь настоящим человеческим языком, поскольку многие аспекты языка связаны с лобными долями, которые у Homo heidelbergensis были меньше, чем у Homo.
В числе ключевых свойств человеческого языка, отличающих его от коммуникативных систем других видов, непременно называют членораздельную звучащую речь. В середине XX века с лёгкой руки американского психолога Олвина Либермана эта идея обрела форму изящного афоризма — Speech is special! (букв, «речь [видо]спе цифична»).
Человеческая речь — не просто издавание звуков, имеющих определенный смысл. Звуковая сторона речи имеет сложную, иерархически устроенную организацию [см., например: Кодзасов, Кривнова, 2001]. Из отдельных звуков — фонем — складываются слоги, из слогов — фонетические слова (они могут не совпадать с грамматическими словами, например, в -одно фонетическое слово, но два грамматических), из фонетических слов — фонетическим синтагмы (сочетания из нескольких фонетических слов без пауз между ними), а из них — фонетические предложения, или периоды. Непросто устроены и сами звуки. При помощи движений языка, губ, нижней челюсти, нёбной занавески, надгортанника, человек, изменяя резонансные свойства речевого тракта, ослабляет одни частоты получающегося в итоге звука и усиливает другие. Каждый гласный характеризуется своим собственным «узором» усиленных частот (формант). Согласные также имеют свои частотные максимумы и минимумы, но распознаются в значительной степени по тому влиянию, которое они оказывают на форманты соседствующих с ними гласных. Например, после заднеязычного согласного (г или к) у последующего гласного сближаются начальные точки контуров второй и третьей формант.
Вообще говоря, прямое отождествление человеческого языка и членораздельной звучащей речи не вполне верно, поскольку жестовые языки глухонемых ни в коей мере не являются «менее человеческими», чем языки устные. Вопреки распространённому заблуждению, жесты этих языков передают не отдельные буквы (хотя пальцевая азбука — дактилология — тоже имеется, прежде всего для передачи имен собственных), а целые слова (или морфемы — значимые части слов). Каждый жест-слово состоит из незначимых элементов — хирем, а из слов, как и в устном языке, составляются словосочетания и предложения [см.: Stokoe, 1960; Зайцева, 1993]. В них имеется грамматика (например, средства выражения множественного числа, разнообразных видовых различий и мн. др.), различные стили речи, на них можно вести диалоги и составлять монологические рассказы на любые темы (например, можно пересказать приятелю сюрреалистический мультфильм).
И всё же, по-видимому, человеческий язык формировался в первую очередь именно как язык устный — об этом говорят имеющиеся у Ното sарiens многочисленные приспособления для производства и восприятия членораздельного звука. Главное из них — гортань, расположенная ниже, чем у современных человекообразных обезьян. Низкое положение гортани открывает возможности для чёткого произнесения звуков человеческого языка, но при этом создаёт риск подавиться. Как пишет С. Пинкер, «до недавнего изобретения приёма Геймлиха попадание еды в дыхательные пути было шестой лидирующей причиной смерти от несчастного случая в Соединенных Штатах, уносившей шесть тысяч жизней в год» [Пинкер, 2004].
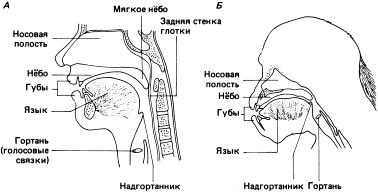
Схематическое изображение (сагиттальное сечение) головы и шеи А – взрослый человек; Б – взрослый шимпанзе. Строение гортани шимпанзе и ее расположение в верхней части голосового пути не позволяет им, в отличие от человека, с помощью языка менять конфигурацию глотки и осуществлять модуляцию звуков (по Lieberman 1968, с изменениями). Источник
Следует уточнить, что для членораздельной речи важно не положение гортани само по себе, а соотношение длины ротовой полости и длины глотки: у современного человека эти длины приблизительно одинаковы, так что части языка, расположенные во рту и в глотке, примерно уравниваются, и это даёт возможность чётко различать в произношении все звуки, включая «крайние» гласные — [i], [u] и [а] [Lieberman, 2002]. У других видов гоминид такой возможности, видимо, не было, поскольку они, как видно по сохранившимся черепам, обладали достаточно сильно выступавшими вперёд челюстями и, соответственно, длинной ротовой полостью. Чтобы уравновесить такую длину рта соответствующей длиной глотки, гортань должна была бы располагаться где-то в груди, что крайне маловероятно [ср.: Lieberman, 2002]. Впрочем, невозможность произнесения «крайних» гласных сама по себе не может свидетельствовать ни об отсутствии языка (язык мог в принципе быть и жестовым), ни даже об отсутствии членораздельной звучащей речи — просто, если таковая была, она должна была в большей степени полагаться на различия согласных звуков [Deacon, 1997].
Отметим, что у человеческих младенцев гортань тоже, как и у шимпанзе, расположена высоко (это позволяет одновременно сосать и дышать). Примерно к трём годам гортань опускается — и это приблизительно совпадает со временем полного овладения звуковой стороной языка. Впрочем, справедливости ради следует сказать, что положение гортани не остается неизменным в течение жизни не только у человека: по данным группы японских ученых, некоторое опускание гортани наблюдается и у шимпанзе [Nishimura et al., 2003].
О том, для чего нужно низкое положение гортани, существует по меньшей мере две гипотезы. Согласно одной точке зрения, оно необходимо только для членораздельной звучащей речи [см., например, Lieberman, 2002; Вишняцкий, 2004], поскольку дает возможность языку двигаться внутри речевого тракта как в горизонтальной, так и в вертикальной плоскости. Это позволяет создавать различные конфигурации ротовой полости и глотки независимо и тем самым сильно расширяет набор возможных фонем, различающихся по тому, на каких частотах звук усилен, а на каких, наоборот, приглушён. Согласно другой точке зрения, главная роль понижения гортани — это обеспечение возможности издавать более низкие звуки и тем самым создавать у слушателей впечатление, что говорящий имеет бóльшие размеры, чем на самом деле [Hauser et al., 2002. Р. 1574]. По-видимому, эта точка зрения принципиально неверна. Дело не только в том, что преувеличение собственного размера — слишком ничтожное приобретение для такой огромной «цены», как риск подавиться.
Главное, как кажется, то, что приматы а значит, по-видимому, и ранние гоминиды) — групповые животные с достаточно высоким уровнем интеллекта. Они долгие годы живут вместе, часто встречаются и хорошо знают друг друга «в лицо» — как показывают наблюдения, роль межличностных контактов в обезьяньем сообществе весьма велика [см., например, Бутовская, Файнберг, 1993]. В такой ситуации пытаться создавать ложное впечатление о размере (который виден невооружённым глазом и всей группе давно известен) просто бесполезно (показательно, что М. Хаузер и его соавторы в подтверждение своей точки зрения ссылаются на лягушек и птиц, которые производят коммуникативные действия на таких расстояниях и в такой среде, что размер того, кто издаёт звук, слушающему не виден).
Предположение, что понижение тембра голоса было необходимо в межгрупповых конфликтах (для того, чтобы издалека устрашить членов соседней группировки) тоже едва ли может претендовать на обоснованность: во-первых, такая задача должна была обусловить понижение гортани у взрослых особей мужского пола, но не у женщин и трёхлетних детей, для которых устрашение соседей неактуально, а во-вторых, слух человека настроен на преимущественное восприятие частот, слишком высоких для тех расстояний, на которых осуществляется межгрупповое общение (см. подробнее ниже). Таким образом, остается лишь одна возможность: низкое положение гортани как видовой признак — это одно из приспособлений для членораздельной звучащей речи.
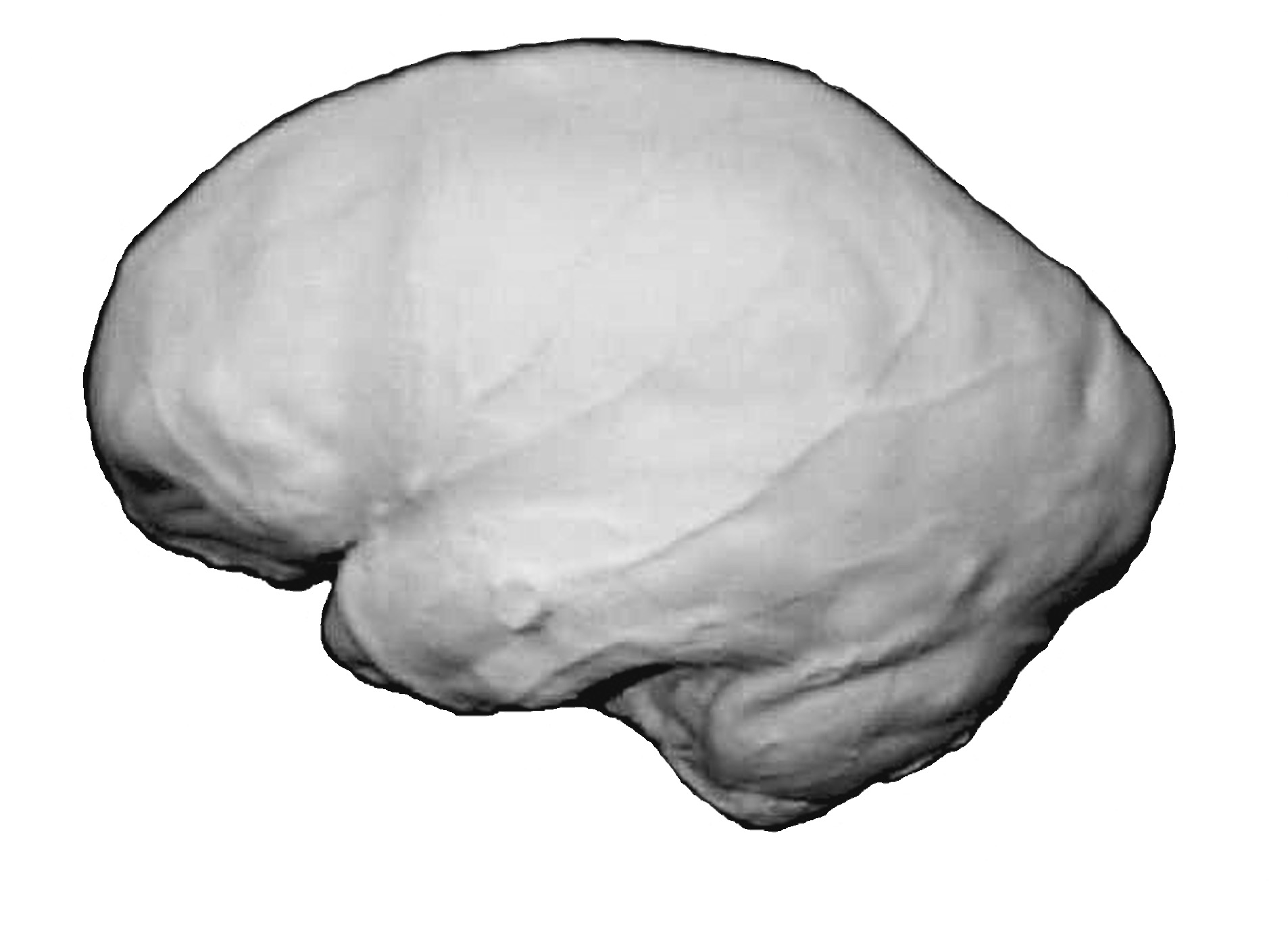
Эндокран Homo heidelbergensis (Брокен-Хилл)
Иллюстрация из книги: Дробышевский С.В. Эволюция мозга человека (анализ эндокраниометрических признаков гоминид). М., КомКнига, 2007
Сложность заключается в том, что гортань не содержит костей, а мягкие ткани не сохраняются, так что все имеющиеся в науке сведения о положении гортани у того или иного вида гоминид — это реконструкции, выполненные на основании предположения, что положение гортани связано с величиной базикраниального угла (угла основания черепа): чем сильнее изогнуто основание черепа, тем ниже расположена гортань [Lieberman, Crelin, 1971; Lieberman et al.,, 1972; Laitman, Crelin, 1976; Crelin, 1987]. У австралопитеков и Homo habilis изгиб основания черепа примерно такой же, как у современных человекообразных обезьян, у архантропов он выражен значительно сильнее — поэтому некоторые исследователи предполагают, что «первыми «говорящими» гоминидами были, возможно, Ното еrgaster» [Зубов, 2004]. Впрочем, исследования указывают на отсутствие (у современных людей) надёжной корреляции между величиной изгиба базикраниума и положением гортани [Leberman, McCarthy, 1999].
Но низкое положение гортани — далеко не единственное анатомическое приспособление, необходимое для языка. Большое значение для звучащей речи имеет точное управление мышцами языка. Движения всех этих мышц, кроме одной, обеспечиваются подъязычным нервом, канал которого расположен в толще затылочной кости между ярёмным отростком и мыщелком. Чем толще этот канал, тем, как считается, больше нейронов задействовано в управлении языком, поэтому на основании этого признака делаются предположения о наличии у того или иного вида гоминид звучащей речи: в частности, известно, что у представителей рода Ното этот канал как абсолютно, так и относительно больше, чем у человекообразных обезьян и австралопитековых [Кау et al., 1998]. Однако, по данным других исследователей, с учётом различий в размере языка толщина канала подъязычного нерва варьирует практически в одних и тех же пределах даже у человека и шимпанзе [Fitch, 2000. Р. 262; Lieberman, 2002. Р. 177]; кроме того, отсутствует корреляция между толщиной канала подъязычного нерва и числом аксонов, из которых этот нерв состоит [DeGusta et al., 1999].
В обеспечении членораздельной звучащей речи участвует подъязычная кость. У человека она расположена ниже, чем у других приматов, благодаря чему сильно расширяется спектр возможных движений глотки, гортани и языка друг относительно друга. Если бы подъязычная кость располагалась у нас иначе, мы были бы способны произносить не больше различающихся звуков, чем, например, шимпанзе. Как была расположена эта кость у других представителей клады человека — неизвестно, поскольку эта небольшая кость, не прикреплённая к остальному скелету, обычно не сохраняется. К настоящему времени исследователи располагают лишь очень немногими образцами подъязычной кости. Наиболее известны найденная в Израиле (пещера Кебара) подъязычная кость неандертальца [Аrensburg et al., 1989] и подъязычная кость гейдельбергского человека из Испании (район Атапуэрка, пещера Сима де лос Уэсос); кроме того, в пещере Сидерон в Астурии, близ Пилона, была найдена частично сохранившаяся подъязычная кость гоминида, принадлежавшего к неандертальцам (или гейдельбержцам) [Саbo et al., 2003].
Все эти кости, хотя и несколько отличаются по своему строению, чрезвычайно сходны с теми, которые можно наблюдать у современных людей [Зубов, 2004] (в частности, в них отсутствуют отверстия для горловых мешков, свойственных современным шимпанзе [Fitch, 2000]), и это даёт возможность высказать гипотезу, что голосовые аппараты неандертальца, гейдельбержца и современного человека анатомически чрезвычайно близки. Напротив, найденная недавно подъязычная кость афарского австралопитека оказалась такой же, как у шимпанзе [Аlemseged et al., 2006].
Не менее важен для использования звучащей речи тонкий контроль дыхания [МасLarnon, Hewitt, 1999]. Дело в том, что при речи, в отличие от нечленораздельного крика, воздух надо подавать на голосовые связки не сразу, а небольшими порциями — слогами. Это позволяет строить длинные высказывания на одном выдохе, перемежая его короткими вдохами в моменты значимых для смысла и/или синтаксиса пауз. В рамках одного такого высказывания можно произнести большое количество слогов — тем самым, возникает эволюционная задача снабдить эти слоги необходимым количеством различий, что позволит придать высказыванию большую информационную наполненность. Если бы воздух подавался на голосовые связки сразу весь, возможности изменения звучания в процессе одного выдоха-высказывания были бы крайне ограничены (читатель может убедиться в этом сам, попытавшись снабдить членораздельными изменениями звучания, скажем, вопль ужаса). Как следствие, в таком языке оказалось бы очень мало слов: слишком малые возможности варьирования звука не позволяли бы проводить большое число различий.
Кроме того, при произнесении фонем сближающиеся органы артикуляции ослабляют акустическую мощность — по-разному в разных случаях, так что при одинаковой силе воздушного потока, подаваемого на голосовые связки, одни звуки оказались бы настолько громче других, что заглушили бы последние (при восприятии наблюдается «маскирующий» эффект: тихий звук, непосредственно предшествующий громкому или следующий за ним, не распознаётся). Таким образом, речевое дыхание должно не только квантовать выдох на слоги, но и регулировать силу выдоха в рамках одного слога так, чтобы соседствующие звуки не заглушали друг друга. Как было показано ещё Н.И. Жинкиным [Жинкин, 1958] при помощи рентгенокимографи- ческой съёмки, это обеспечивается движениями диафрагмы: «в процессе речевого произнесения диафрагма на выдохе совершает резкие и отчётливо заметные вдыхательные и выдыхательные движения.
Она модулирует с определённой амплитудой на каждом речевом звуке, то поднимаясь вверх, то опускаясь вниз, при этом экспирация не прекращается» [Жинкин, 1958]. Например, при произнесении слова скалы «на слоге ска диафрагма вначале делает два движения вверх (ск), потом опускается на а. После этого идёт краткое нижнее слогостояние диафрагмы и новый слог лы, который начинается малым подъёмом диафрагмы на л и вторым, большим подъёмом на ы» [там же], при этом «в момент падения диафрагмы на слогоразделе вдоха не происходит» [там же]. Диафрагма иннервируется диафрагмальными нервами, отходящими от шейного отдела спинного мозга на уровне третьего, четвёртого и пятого шейных позвонков. В речевом дыхании задействованы также межрёберные мышцы, которые иннервируются из грудных спинно-мозговых нервов.
Таким образом, для эффективного управления дыханием при речи необходим достаточно широкий позвоночный канал. По имеющимся данным, у неандертальца и гейдельбергского человека этот канал был примерно таким же широким, как у неоантропа, тогда как у архантропа и даже у Ното antecessor - значительно ýже [MacLarnon, Hewitt,1999, 1999; Walker, Shipman, 1996; Дробышевский, 2004].
Иногда можно встретить утверждение о том, что значительную роль для членораздельной речи играет подбородочный выступ. Но это не вполне верно. Подбородочный выступ — это просто результат неравномерной редукции челюстей, происходившей в процессе эволюции человека. Другое дело, что при развитии речи мышцы языка совершали всё больше разнообразных тонко дифференцированных движений, и именно необходимость в прикреплении этих мышц, возможно, уберегла нижнюю челюсть от редукции. Более того, на ней возникли подбородочные ости и выступ. В становлении же членораздельной речи сыграл роль не подбородочный выступ как таковой, а изменение способа прикрепления подбородочно-язычной мышцы с мясистого на сухожильный. Впрочем, как отмечал В.В. Бунак, для развития членораздельной речи уменьшение размеров нижней челюсти сыграло положительную роль, поскольку «при быстрой смене артикуляции массивная нижняя челюсть и мускулатура создавали бы большую инерцию в работе речевого аппарата, основанной, как известно, именно на быстрой смене артикуляции» [Бунак, 1980].
Из всех гоминид подбородочный выступ имеется лишь у неоантропа, но у некоторых неандертальцев прослеживается определённая тенденция к его формированию [Зубов, 2004]. Присутствует в зачаточном состоянии подбородочный выступ, например, у неандертальца Кебара 2, обладателя сохранившейся подъязычной кости. Впрочем, для выводов о системе коммуникации, возможно, более показательно строение внутренней поверхности нижней челюсти [Дробышевский С.В., устное сообщение]: в её середине (в области симфиза) у человека имеются подбородочные ости (места прикрепления подбородочно-язычной мышцы); у обезьян же на этом месте ямка (поскольку эта мышца прикрепляется к кости не сухожилием, а мясистой частью). Челюсти ископаемых гоминид демонстрируют спектр переходных форм.
Анатомические изменения, связанные с развитием членораздельной звучащей речи, коснулись не только речевого аппарата. У человека иначе, чем, например, у шимпанзе, устроен слуховой анализатор. У нас имеется область лучшей слышимости в диапазоне от 2 до 4 кГц — именно на этих частотах сосредоточены значимые характеристики фонем. Шимпанзе же лучше всего слышат звуки частотой около 1 кГц-для них это очень важно, поскольку примерно такую частоту имеют их «долгие крики» (один из типов коммуникативных сигналов). Настройка слуха на повышенную чувствительность к высоким частотам происходила ещё у предков человека современного типа — у Homo heidelbergensis.
И. Мартинес и его коллеги исследовали слуховые косточки Homo heidelbergensis, найденных в Испании (местонахождение Сима-де-лос-Уэсос) и реконструировали, каким слухом обладали представители этого вида [Martinez et al., 2004]. Оказалось, что область лучшей слышимости в диапазоне от 2 до 4 кГц к этому времени уже начала формироваться, но ещё не установилась полностью, поскольку в степени её развития разные индивиды обнаруживают доcтаточно значительную вариативность.
Анализатор речевых звуков работает у человека чрезвычайно быстро (быстрее, чем распознаются неречевые звуки) — до 20-30, а при искусственном ускорении речи — до 40-50 фонем в секунду [Пинкер, 2004]. При этом люди могут проводить достаточно тонкие фонетические различия, например, мы способны не перепутать такие похожие звуки, как b и p (физически отличающиеся друг от друга тем, что колебания голосовых связок начинаются либо одновременно с тем, как разомкнутся губы, либо после этого).
Важным свойством человеческой коммуникации является то, что она подконтрольна воле, а не эмоциям (т.е. управляется структурами коры больших полушарий): чтобы заговорить, нам необязательно приходить в сильное возбуждение (это скорее помешает), надо лишь захотеть нечто сказать.
Главную роль в обеспечении функционирования языка играют две области левого (в норме у правшей) полушария: зона Брока и зона Вернике. Зона Вернике, соседствующая с зоной визуального распознавания, хранит образы отдельных языковых элементов, зона Брока, прилегающая к премоторной коре, — программы обращения с ними. Но не менее важны и другие отделы мозга, в особенности лобные доли — они обеспечивают возможность подавлять лишние эмоции, а также сосредоточивать внимание на главном, отвлекаясь при этом от несущественных деталей [см., например, Дерягина, 2003]. В отсутствие такой возможности люди никогда не смогли бы распознать, например, какие элементы фонетической реализации звука несут смыслоразличительную нагрузку, а какие — нет. При поражениях лобных долей человек не теряет дара речи, но утрачивает возможность строить поведение по словесной инструкции [см., например, Шульговский, 2003]. Если поражены префронтальные отделы коры, больной может повторять слова и целые фразы, но не в состоянии самостоятельно высказать какую-либо мысль или задать вопрос.
Следует отметить, что в коре больших полушарий связаны между собой самые разные стороны восприятия одного и того же объекта: его внешний вид, запах и вкус (если он их имеет), звуки, называющие этот объект, звуки, производимые этим объектом (если он производит звуки), ощущение этого объекта в руке (если его можно взять в руку), представление о манипуляциях с ним и т.д. Словом, все то, что позволяет нам, видя (слыша, обоняя, осязая) этот объект, понимать, чего от него можно ожидать, что с ним можно (или даже нужно) делать, а чего нельзя [ср.: Calvin, Bickerton, 2000]. В хранение наших знаний о различных объектах оказываются вовлечены те отделы мозга, которые регулируют связанное с этими объектами поведение: например, в распознавании инструментов участвует премоторная кора, которая управляет рабочими движениями, а «при категоризации и назывании изображений животных, напротив, активируются прежде всего затылочно-височные области, ответственные за сложные формы зрительной обработки и восприятие движения» [Величковский, 2006].
Это оказывается важным при распознавании звучащей речи. В мозге, как показывают данные мозгового картирования, существуют специальные участки, предназначенные для обработки речевых звуков (отличные от тех, которые используются для распознавания звуков неречевых) [Hickok, Poeppel, 2000; Poeppel, 2001; Trout, 2001;Vouloumanos et al., 2001]. Эти участки позволяют обнаруживать различные простые характеристики акустических событий: наличие звучания на определенной частоте, увеличение энергии звука, уменьшение энергии звука, скорость изменения энергии звука, повышение частоты, понижение частоты и некоторые другие [Кодзасов, Кривнова, 2001]; различные комбинации показаний детекторов складываются в смыслоразличительные признаки фонем (набор которых уникален для каждой фонемы).
Но человек, воспринимая и понимая речь, не распознаёт фонему за фонемой, складывая их потом в слова (подобно компьютеру, побуквенно распознающему отсканированный текст), распознавание речи устроено значительно сложнее. Во-первых, от звука к звуку в потоке речи имеются вполне акустически заметные переходы (поэтому, если звуки в слоге поменять местами, человек услышит не слог, произнесённый наоборот, а бессмысленную абракадабру — за счёт того, что привычные ему правила перехода от звука к звуку не будут соблюдены). Формантные переходы между соседствующими звуками нередко позволяют людям «услышать» нужный звук даже в том случае, когда он не был реально произнесён [Sinnott, Saporita, 2000], — и человек вполне может не осознать, что вместо, скажем, Он — человек ответственный услышал …чек отвесный. Во-вторых, звуки речи встречаются — если не брать экспериментов — в словах, а «информация, достаточная для опознания слова по звуковому облику, включает в себя его общую длину, просодический контур, несколько гласных и согласных звуков, следующих друг за другом в определенном порядке» [Бурлак, Старостин, 2005].
Кроме того, слова употребляются в высказываниях, а высказывания — в тех или иных жизненных ситуациях, тем самым количество «контекста» (как языкового, так и внеязыкового) увеличивается. К распознаванию звуков речи может подключаться также зрительный анализатор, чему свидетельством известный «эффект Мак-Гурка»: если дать человеку прослушать слог ba и при этом показать ему губы, произносящие ga, он, автоматически сделав соответствующую поправку, воспримет услышанное как слог da [McGurk, MacDonald, 1976] (разомкнутые губы не могли произносить b, а шум на тех частотах, которые характерны для b, можно с некоторым напряжением принять за d, но никак не за g). Все это позволяет людям понимать друг друга даже при наличии ошибок.
Всем этим анатомо-физиологическим приспособлениям сопутствуют приспособления когнитивные: дети приходят в мир с желанием обнаруживать слова, т.е. интерпретировать звуки, произноcимые окружающими, как знаки. Желание слышать понимаемую речь столь велико, что подчас заставляет человека обнаруживать слова в шумах природы (например, песня птицы чечевицы описывается обычно как «Витю видел?»). Даже новорожденные младенцы предпочитают звуки речи (включая такие, которые они не могли различать в утробе) неречевым звукам, похожим на речевые по своему тембру и ритмике [Vouloumanos, Werker, 2004a; 2004b].
Большое значение для развития речи имеет способность к звукоподражанию: притом, что люди, в массе своей, достаточно плохие имитаторы (далеко не каждый способен адекватно изобразить пение синицы, мяуканье кошки или правильно воспроизвести даже несложную мелодию), подражание речевым звукам даётся им гораздо лучше: уже к трём-четырём-пяти годам дети научаются правильно произносить все согласные и гласные своего родного языка, воспроизводить тоны (в тех языках, где они есть), интонационную структуру разных типов предложений и т.д. Важно, что звукоподражание это является самоподдерживающимся: детям, овладевающим языком, не нужно специальное поощрение за выученные элементы коммуникативной системы.
Формирование всего этого комплекса признаков начинается лишь в гоминидной линии. У современных человекообразных обезьян высоко расположена гортань (что позволяет есть и дышать / издавать звуки практически одновременно), имеются горловые мешки, иначе, чем у человека, крепится к нижней челюсти подбородочно-язычная мышца, отсутствует произвольная регуляция дыхания (зато имеется возможность издавать звуки как на выдохе, так и на вдохе), звукопроизводство подконтрольно эмоциям, а не воле (и управляется подкорковыми структурами мозга), слуховой анализатор настроен на преимущественное распознавание звуков только на низких частотах, желание обнаруживать слова, видеть за звуками имена всего сущего не отмечается даже в языковых проектах, способности и склонности к звукоподражанию нет.
При этом у обезьян имеются гомологи зоны Брока и зоны Вернике [Зорина, Смирнова, 2006], соответствующие человеческим не только по своему расположению, но и по клеточному составу, а также по входящим и исходящим нейронным связям; кроме того, эти области — как у человека, так и у человекообразных обезьян — соединены между собой пучком волокон (это было показано как отечественными, так и зарубежными исследователями [Кочеткова, 1973; Galaburda, Pandya, 1982; Deacon, 1997]). Гомолог зоны Брока «отвечает» за автоматические комплексные поведенческие программы, осуществляемые мышцами лица, рта, языка и гортани, а также за координированные программы действий правой руки [Greenfield, 1991; Rizzolatti, Arbib, 1998]. Гомолог зоны Вернике (и соседние участки мозга) используются для распознавания звуковых сигналов, а также для того, чтобы различать сородичей по голосу.
Кроме того, «различные подобласти этих гомологов получают данные от всех областей мозга, задействованных при слушании, ощущении прикосновения во рту, языке и гортани и областях, где сливаются потоки информации от всех органов чувств» [Пинкер, 2004]. Таким образом, обнаружение на эндокранах ископаемых гоминид развитых выпуклостей в области зоны Брока и зоны Вернике не может свидетельствовать о наличии у соответствующего вида членораздельной звучащей речи [ср.: Tobias, 1996].
При этом у Ното sapiens весь этот комплекс признаков закреплён в геноме, и, поскольку появление такого сложного комплекса признаков в результате единственной макромутации совершенно невозможно, это означает, что перед видом- предком задача приспособления к членораздельной звучащей речи уже стояла (и основу вида-потомка положили те, кто смог её успешно решить).
В качестве «гена речи» нередко рассматривается ген FОХР2, расположенный на седьмой хромосоме. Люди, имеющие дефектный вариант этого гена, страдают специфическим расстройством речи (англ. SLI), затрагивающим как фонетическую, так и грамматическую составляющие языка [см., например, Бурлак, 2011. С. 255, 256]. Кроме того, у них несколько нарушен волевой моторный контроль за мышцами рта (в области как отдельных движений, так и их последовательностей) — им сложно, например, по команде высунуть язык или несколько раз подряд сомкнуть и разомкнуть передние зубы [Alcock et al., 2000]. Как показали исследования, он является геном-регулятором высокого уровня (т.е. регулирует активность других генов-регуляторов) [Konopka et al., 2009]; он экспрессируется в различных отделах мозга, в частности, влияет на характер нейронных связей между корой больших полушарий и базальными ядрами, повышая синаптическую пластичность в них [Reimers-Kipping et al. 2011], что является чрезвычайно существенным для возможности выучивать последовательности действий [Jin, Costa, 2010]. [У певчих птиц, где видовая песня выучивается молодыми самцами от взрослых, ген FOXP2 вовлечён в регуляцию этого процесса.$ Вообще, обучение пению у 3-х групп птиц, сформировавших его независимо друг от друга — певчих воробьиных, попугаев и колибри — показывает параллелизм с организацией человеческой речи как на уровне морфологии (отделов мозга, вовлечённых в «запись» и «воспроизведение»), так и на уровне генов.]
Ген FОХР2, по всей видимости, был мишенью отбора в гоминидной линии: со времени разделения предков людей и шимпанзе в этом гене произошли две замены, и обе они являются несинонимичными [Enard et al., 2002]. Как показали недавние исследования, таким же, как у нас, вариантом этого гена обладали и. неандертальцы [Krause et al., 2007]. Тем самым, велика вероятность, что «человеческие» мутации в этом гене произошли уже у общего предка неандертальца и Ното sapiens, т.е. у Homo heidelbergensis, расширив его возможности как в области моторного контроля за органами артикуляции, так и в области автоматизации последовательностей действий, что особенно важно для тех, кто может произносить длинные, многосложные реплики и перед кем, соответственно, стоит задача наделять эти реплики необходимым количеством различий между соседствующими элементами.
Важное значение для определения времени формирования членораздельной звучащей речи имеет недавнее исследование Б. де Бура [de Boer, 2011], направленное на выявление функции горловых мешков. Эти образования имеются у современных шимпанзе (и имелись, судя по строению подъязычной кости, у афарских австралопитеков), но отсутствуют у людей (а также у неандертальцев и Homo heidelbergensis). Построив модель речевого резонатора с горловыми мешками и без них, он показал, что при наличии горловых мешков во-первых, резонансы речевого тракта смещаются ближе друг к другу, а во-вторых, появляются дополнительные резонансы и антирезонанcы — причём появляются независимо от производимой артикуляции. Из этого сразу видна отрицательная роль горловых мешков для членораздельности речи. Во-первых, если все области усиления звука приближены друг к другу, это значит, что звуки получаются более похожими один на другой, тогда как для членораздельной речи необходимо, наоборот, чтобы звуки достаточно сильно различались.
Увеличение различающихся на слух звуков позволяет иметь коммуникативную систему с большим числом знаков (и тем самым, с большими выразительными возможностями). Во-вторых, наличие резонансов и антирезонансов, независимых от производимой артикуляции, сильно сужает возможности произвольного варьирования производимого звука. Такая задача актуальна для обезьян, которые, имея высоко расположенную гортань, могут есть и вокализировать одновременно: при наличии горловых мешков еда, находящаяся во рту, не мешает издавать необходимые звуки. Но членораздельная речь обусловливает противоположную задачу: при помощи органов артикуляции, доступных волевому управлению, обеспечить как можно большее количество различий в звучании. Ещё одна функция горловых мешков — понижение высоты звука.
Эта задача также актуальна для обезьян, которые используют звуковую коммуникацию для общения с сородичами, находящимися сравнительно далеко и скрытыми густой листвой тропического леса (в общении на близком расстоянии более существенную роль играет мимика, жесты, позы и разнообразные прикосновения) и — в связи с этим — имеют слуховой анализатор, настроенный на преимущественное распознавание низкочастотных звуков. Но для гоминид, обитающих в полуоткрытых и открытых ландшафтах, эта задача постепенно теряет актуальность. Слуховой анализатор Нотo демонстрирующий формирование дополнительной области лучшей слышимости на высоких частотах, свидетельствует, что на первый план у гоминид постепенно выходило звуковое общение на близком расстоянии.
Всё это позволяет в общих чертах представить себе картину формирования звучащей речи в ходе эволюции. Первоначально основными носителями намеренно передаваемой сигнальной информации у гоминид, как и у современных приматов, вероятно, были жесты — они подчинены волевому контролю и могут использоваться для создания сигналов аd hoc (сигналов, создаваемых по ходу дела, у которых ни форма, ни смысл не являются врождёнными). Звуки же могли использоваться только в качестве эмоционального дополнения. Но когда объем манипулятивной активности гоминид возрос, в частности, за счет всё более частого изготовления и применения орудий, сочетание обычной и коммуникативной деятельности стало затруднено [Бутовская, 2004]: руки не могли одновременно делать орудия и знаки, мозгу приходилось выбирать, какой сигнал посылать на руки, какую информацию обрабатывать — от практических движений или от сигнальных (подобные затруднения легко смоделировать, попытавшись говорить и одновременно с этим жевать жвачку).
Это, по-видимому, привело к эффекту замещения — сигнал из мозговых структур, управляющих коммуникацией, стал подаваться не только на руки, но и на органы звукопроизводства (и зона Брока, соседствующая с премоторной корой, получила в своё распоряжение контроль над вокализацией). Такое замещение могло быть облегчено тем, что у приматов управление ротовым аппаратом и управление руками связаны между собой, поскольку эти органы вместе задействованы в питании, груминге и т.д. [см., например, Pollick, de Waal, 2007].
Как было показано ещё советским археологом-экспериментатором С.А. Семёновым, самое примитивное «рубящее орудие возникало после двух-трёх ударов оббивки» [Семёнов, 1968], а для получения отщепа с острым краем бывает достаточно и одного (сходные результаты были получены и позднее, Н. Тотом и К. Шик, в том числе, в экспериментах, где орудия олдувайского типа изготавливал бонобо Канзи [Schick, Toth, 1993]); ашельская же технология предполагает гораздо большие трудозатраты: орудие обрабатывается со всех сторон, при этом сначала производится грубая оббивка каменным молотком, а затем края ретушируются с использованием мягких отбойников из кости или рога (подробнее о технологиях изготовления палеолитических орудий см. [Inizan et al., 1999; Schick, Toth, 1993]).
Тем самым, с развитием технологии производство орудий требовало всё большего времени. Если при этом необходимо было коммуницировать, то в выигрыше оказывались те, кто мог ещё до начала собственно передачи информации (т.е. использования значащего, подконтрольного воле жеста), по звуку общего возбуждения или привлечения внимания догадаться (хотя бы в какой-то степени) о том, что будет сообщено. В принципе такое не невозможно: так, человек, слыша обращение к себе по имени, может по интонации предугадать часть смысла будущего сообщения — намерен ли говорящий просить его о чём-то, угрожать ему, стыдить, подозвать к себе, сообщить о каком-то поразившем его событии и т.д.; при становлении речи у ребёнка овладение интонацией происходит до овладения словами [Лепская, 1997, Цейтлин, 2000].
Соответственно, отбором будет поощряться всё более вариабельный исходный сигнал и всё более точное «угадывание» другими особями по этому сигналу смысла планируемого сообщения. В этом случае информационная нагрузка переместится на звуковой канал, использование же прочих каналов редуцируется. При этом увеличится важность для коммуникации высоких частот, поскольку общение происходит на близком расстоянии.
Вероятно, «архантропы вследствие массивности челюстей могли произносить только небольшое число различных выкриков» [Бунак, 1980]. Поскольку у них не было возможности произносить длинные высказывания (вследствие недостаточных анатомо-физиологических средств для управления дыханием), они могли общаться при помощи голофраз (однословных высказываний), возможно, как современные дети, не столько описывая таким образом те или иные ситуации, сколько выражая свои эмоции по их поводу [Лепская, 1997].
У Homo heidelbergensis ситуация сильно изменилась: увеличенный позвоночный канал свидетельствует о том, что они были в состоянии произносить реплики из нескольких слогов, сочетая при этом по-разному артикулируемые звуки. Это имеет смысл только при наличии возможности вкладывать в звучание максимум различий (максимизируя тем самым передаваемую информацию). И действительно, отсутствие в подъязычной кости отверстий для горловых мешков, а также настройка слуха на высокие частоты показывают, что приспособления для лучшего различения звуков при помощи артикуляции коснулись как звукопроизводства, так и звуковосприятия. Вероятно, это имело генетическую подоснову, обеспеченную мутациями в гене ЕОХР2. Таким образом, можно утверждать, что в основе коммуникации гейдельбергского человека лежала звучащая речь, в которой различия между звуками обеспечивались при помощи артикуляции. Косвенным свидетельством этого может служить и увеличение у Homo heidelbergensis базикраниального угла (по сравнению с тем, что было у архантропов).
Но всё же, по-видимому, эта речь ещё не была настоящим человеческим языком. Для языка очень важна способность делать выводы из нескольких посылок одновременно, сосредоточивать внимание на главном, отвлекаясь от несущественного (в том числе это касается и чисто звуковых различий), держать в оперативной памяти достаточно много единиц, чтобы мочь обобщить синтаксические правила, определённые на длинных предложениях. Всё это обеспечивают лобные доли коры больших полушарий головного мозга, которые у гейдельбергского человека были гораздо меньше, чем у Ното sapiens [см. Дробышевский, 2007].
Итак, анатомически современный человек, появившийся не менее, чем 195 ± 5 тыс. лет назад [McDougall et al., 2008 ], уже, по-видимому, пользовался настоящим человеческим языком. Но база для этого, овладение членораздельностью звучащей речи, была заложена ещё предшествующим видом, Homo heidelbergensis, на несколько сотен тысячелетий раньше.

Благодарность
Работа выполнена при финансовой поддержке гранта РФФИ № 11-06-12036-офи-м-2011.
Вестник МГУ, серия ХХIII. «Антропология». 2012. №3. C.110-120
Рекомендуем прочесть
 Подъём религиозной правой в США 1970-х-80-х гг.
Подъём религиозной правой в США 1970-х-80-х гг. «Круги равных» и «вертикали власти»: мономеры человеческой социальности
«Круги равных» и «вертикали власти»: мономеры человеческой социальности Рождение «глобального потепления»
Рождение «глобального потепления» Да начнутся Игры
Да начнутся Игры Куба не подошла для «твиттерной революции»
Куба не подошла для «твиттерной революции» Евграф Каленьевич Дулуман. Существовал ли Иисус Христос?
Евграф Каленьевич Дулуман. Существовал ли Иисус Христос? Мираж оптимальности: внедрение математических методов в экономику как ответ на проблемы реформы управления промышленностью 1957 года
Мираж оптимальности: внедрение математических методов в экономику как ответ на проблемы реформы управления промышленностью 1957 года Сжечь ведьму! Или российские доброхоты о мигрантах.
Сжечь ведьму! Или российские доброхоты о мигрантах.
