

Гелада Theropithecus gelada
Резюме. Тщательный обзор литературы о том, что такое социальная иерархия, каковые её проявления и адаптивный смысл. Обязательно надо читать всем любителям поп-этологии, чтобы не повторять глупости про «примативность», не воспринимать ранги — будь то альфа, бета или омега -как индивидуальные характеристики. Уже в 1980-х гг. было понятно, что не только у приматов, но даже у хищных, копытных и хоботных (а также врановых, ткачиков, тимелий, попугаев и других «социальных» видов птиц) самый крутой доминант не в состоянии сам, своей собственной силой, поставить себя «царём горы» и тем более удержаться на ней. Нужно умение дружить с равными себе индивидами, создавать коалиции, особи в которых не только поддерживают друг друга, но и связаны неагонистическими взаимодействиями. Нужно быть «популярным» у будущих подчинённых, и уметь превзойти по «популярности» текущего доминанта — часто это более важное условие его замены, чем собственные усилия свергающих, особенно у антропоидов. Именно необходимость коалиций и «популярности» для эффективного продвижения в социальной иерархии тренирует у соответствующих видов птиц и млекопитающих, что 10 годами позднее этологи назовут маккиавеллиевским интеллектом.
Гольцман М.Е.
Введение
Содержание
- 1 Введение
- 2 1.Проблемы определения феномена «доминирования»
- 3 2.Логический статус доминирования
- 4 Связь между доминированием и агрессией
- 5 Адаптивное значение доминирования
- 5.1 5.1. Взаимовлияние связи на связь
- 5.2 5.2. Зависимость доминирования от места и времени взаимодействий
- 5.3 5.3. «Право обладания»
- 5.4 5.4. «Престиж»
- 5.5 5.5. Отношения доминирования-подчинения и структура стада японских макаков
- 5.6 6.1. «Структура внимания»
- 5.7 6.2. Управление движением группы
- 5.8 6.3. Взаимные чистки и «обыскивания»
- 5.9 7.1. Прямая, позитивная связь между доминированием и частотой спаривания
- 5.10 7.2. Очередность копуляций и репродуктивный успех
- 5.11 7.3. Общая и дифференциальная способность к оплодотворению
- 5.12 7.4. «Общий» репродуктивный успех особи и «альтернативные» репродуктивные стратегии
- 6 Заключение
В классических исследованиях поведения, проведенных в начале века, вновь были сделаны попытки описать и оценить социальное доминирование — феномен, к тому времени известный лишь из работ энтомологов XIX века. Интерес к проблеме возник после работ норвежского биолога Торлайфа Шьелдеруп-Эббе, исследовавшего структуру взаимоотношений в группах домашних кур и уток (Schjelderup—Ebbe, 1922а, б; 1923, 1935).
Т. Шьелдеруп-Эббе обнаружил, что взаимоотношения в группах строго упорядочены в единую иерархическую систему доминантных рангов. Каждая особь либо доминирует над партнером, либо подчиняется ему. Эти взаимоотношения были названы «правом клевка» («реск order»), так как доминирующей особью в паре именовали ту, которая в стычках чаще клевала противника. Собственно говоря, клевки становились все более редкими в течение нескольких дней после формирования группы, но зато устанавливалось «право клевка», на основании которого делился насест и регламентировался доступ к корму. Таким образом, был сделан вывод, что при ограниченном ресурсе доминирующая особь получает его в первую очередь — вывод, вызвавший большой резонанс в экологии.
В 30—40-е годы эти исследования получили дальнейшее развитие в различных направлениях, Конрад Лоренц в своих известных работах 30-х годов описал значение рангового порядка в стае галок. Он обнаружил, что птицы высшего ранга, агрессивно реагируют на тех партнеров, статус которых лишь немногим уступает их собственному, но миролюбиво относятся к галкам низших рангов. При спаривании самцы выбирают самок ниже своего ранга и низкоранговая самка может резко повысить свой статус, если станет «супругой» высокоранжированного самца (Lorenz, 1931, 1935).
Уордер К. Олли и его сотрудники в Чикагском университете провели большую серию работ по исследованию факторов, определяющих доминантные отношения. Они изучали и гормональные детерминанты (Alle, Collias, 1938, 1940; Alle et al., 1939; Alle et al., 1940) и роль индивидуального опыта ( Ginsburg, Alle, 1942; Guhl, Alle, 1944; Guhl et al., 1945) и значение самого феномена доминирования для репродуктивного успеха и выживания особи (Alle, 1942), В 1949 году появился сообщающий труд коллектива авторов во главе с У.К. Олли и Альфредом Е. Эмерсоном «Принципы экологии животных», в котором иерархия доминантных рангов, основанная на «праве клевка» рассматривается как основной механизм социальной организации животных (Alle et al., 1949).
В этот же период начинают разрабатываться методы статистического анализа факторов, определяющих результаты взаимодействий и устойчивость социальных связей (Collias, 1943; Landau, 1951а,b, 1965), а в дальнейшем появляются и математические модели социальных структур, основанных на доминантных связях (Landau,1968, Chase, 1974). В подавляющем большинстве названных работ исследовали поведение птиц (как правило, домашних), но в эти же годы началось изучение доминирования и у млекопитающих (Zuckerman, 1932; Harlow, Yudin, 1933; Maslow, 1936 а,в; Maslow, Flanzbaum, 1936; Winslow,1938).
Эти работы в основном велись на приматах, но общее отношение к феномену доминирования было таким же, как и в исследованиях домашних кур: так, если Т. Шьелдеруп-Эббе (1922 а) пишет, что
«из любых двух птиц… одна обладает неизменным превосходством над другой» и «является привилегированной особью, деспотом или тираном, она обладает силой и может использовать ее как пожелает»,
то к подобному же выводу приходит и С. Цукерман (Zuckerman, 1932) в книге «Социальная жизнь обезьян»:
«Физические потребности удовлетворяются настолько, насколько высок доминантный ранг. Доминирование определяет число самок, которыми может обладать самец, а при нехватке корма — количество пищи, которое достается обезьянам…».
На основании подобных описаний были сделаны попытки более строго определить понятие «доминирующего» и «подчиненного» животного. Так например, согласно А. Мэслоу (Maclow, 1936а) доминирующим животным называется такое,
«которое может действовать (при половом, пищевом, агрессивном и социальном поведении), не считаясь с действиями своих партнеров. Подчиненное животное — такое, чьи действия (в половом, пищевом, агрессивном и социальном поведении) вызываются, модифицируются, ограничиваются или ингибируются действиями партнеров».
Описанное явление производило впечатление единого цельного феномена, благодаря которому одно или несколько животных в группе получают приоритет во всех жизненных ситуациях. Эта одномерность дала основание постулировать единый причинный фактор, вызывающий весь «поведенческий синдром доминирования» — побуждение «доминировать» (Maslow, 1936а, в; Maslow, Flanzbaum ,1936).
Феномен доминирования отчетливо проявлялся в группах животных, содержащихся в зоопарках и исследовательских центрах, а с развитием полевых исследований стало очевидно, что доминирование — это не порождение искусственных условий. Было ясно, что избирательность в выживании и в репродуктивном успехе явно зависит от «социальных» факторов. А главным из них, организующим очередность доступа к жизненно важным ресурсам, а, следовательно, лежащим в основе социальных структур, представлялись доминантные отношения.
При описании доминирования приматологи столкнулись с рядом трудностей, например, из-за переплетения сексуальных и агрессивных тенденций в поведении, проявлявшихся как в неотчетливости взаимоотношений, так и в возможности различного толкования внешне подобных поз и актов. Например, С. Цукерман и А. Мэслоу обнаружили, что как павианы, так и макаки используют гомосексуальные садки как форму «ритуализированной агрессии», а «позу подставления» самки, приглашающей к спариванию, — в качестве сигнала покорности и умиротворения. В определенной мере проблемы дифференцировки поведенческих актов возникали из-за несовершенства техники описания и сама концепция доминирования в течение почти четверти века фактически оставалась неизменной.
Кратко представления этих лет можно резюмировать следующим образом. Феномен доминирования не является артефактом содержания в клетках, он характерен для социального поведения животных и в естественных, и в экспериментальных условиях.
Его структура вполне очевидна: доминирование основано на агрессивных взаимодействиях, таких как удары, атаки, укусы и связанные с ними угрозы. Когда животные вступают в открытую борьбу, одно из них может быть изгнано, искалечено или даже убито и, таким образом, в буквальном смысле, появляются победитель и побежденный.
Оценка экологического значения доминантных связей базировалась на представлении, что доминирование ранжирует членов группы в соответствии со способностям каждого из них адаптироваться к данным условиям среды. Таким образом, ранжирование выделяет наиболее приспособленных, жизнеспособных, обеспечивая их лучшую индивидуальную и репродуктивную выживаемость и, следовательно, — их успех в процессе естественного отбора. Иерархия доминантных рангов рассматривалась как основа социальной структуры, тем более, что жестко организованные сообщества казались наиболее приспособленными. Например, об их преимуществе свидетельствовали изящные эксперименты, проведенные А.М. Гулом и У.К.Олли (Guhl, Alle, 1944). Исследователи сравнили разные группы кур. В одних группах еженедельно удаляли доминанта, заменяя его чужой особью, чтобы дезинтегрировать группу, в жизнь других групп (контрольных) — не вмешивались. Оказалось, что контрольных группах агрессивные стычки происходят реже, яйценоскость птиц, выше, интенсивнее потребляется корм. Из этого был сделан вывод, что организованные группы обладают преимуществом, благодаря стабильной социальной среде и, следовательно, отбор должен им благоприятствовать?
В 50-60-е годы диапазон исследований, прямо или косвенно связанных с изучением доминирования, еще более увеличивается. Разрабатываются новые направления поисков психологических, психофизиологических и физиологических коррелятов доминирования. Одно из наиболее мощных направлений возникло в результате прочного вхождения в экофизиологию концепцию стресса (Селье, 1960). Предположение о том, что стресс-это основной фактор, перестраивающий физиологию подчиненных особей и таким образом «забивающий клин» между ними и доминантом, завоевало большую популярность не только у экспериментаторов, но и у полевых экологов, анализирующих механизмы регуляции численности популяции (Christian, 1968, 1980; Шилов, 1977).
Проблемы, связанные с теорией «группового отбора», в этих работах не обсуждались.
Однако при расширении диапазона исследуемых условий эта благополучная картина стала нарушаться. В конце 60-х — начале 70-ос годов необходимость пересмотра существующих представлений стала очевидной для многих приматологов, у которых уже накопился достаточный опыт наблюдений за межиндивидуальными связями животных в естественных сообществах. Негативная реакция на широкое использование концепции доминирования основывалась на следующих доводах:
(1) концепция доминирования основана на выявлении определённых корреляций между поведенческими параметрами, а при увеличении разнообразия исследуемых ситуаций эти корреляции становятся все менее заметными;
(2) иерархические структуры в наиболее очевидной форме проявляются в условиях неволи или в каких-то критических ситуациях и, возможно, не являются внутренне присущими данной группе, а лишь отражают специфику условий, в которых она находится.
Были предприняты разнообразные попытки установить теоретическую ценность концепции доминирования и анализирующие структуру, функции и эволюционное значение связанного с ней биологического феномена. Результаты этих исследований показали, что мы имеем дело с явлением гораздо более сложным, чем предполагалось раньше. Многие прежние представления, связанные с этой концепцией, оказались несостоятельными, но новой целостной системы понятий пока не появилось.
Современное состояние концепции доминирования обсуждалось в ряде работ: T.E.Rowell (1966,1967,1972, 1974); J.S.Gartlan (1968); D. van Kreveld (1970); S.M.Richards (1974); G.J.Syme (1974); E.O.Wilson (1975); T.H.Clutton-Brok, Harvey (1976); J.M.Deag (1977); R.A.Hinde (1974. 1978); L.S.Bernstein (1970, 1981); D.Dewsbury (1982). К рассмотрению возникших при этих обсуждениях проблем мы сейчас и переходим.
1.Проблемы определения феномена «доминирования»
Прежде чем рассматривать доминирование, его, видимо, было полезно определить. Хотя во многих работах исследователи разбирали причины возникновения доминантных отношений, их функции и эволюционное значение, дать сколько-нибудь полное и универсальное определение доминирования оказалось сложной проблемой, Ее постановке (но не решению) была посвящена недавняя статья И. Бернстайна (Bernstein, 1981), в обсуждении которой приняли участие 28 авторов. Однако сколько-нибудь жесткой платформы для формулирования определения не создано.
Трудность формулировки структурного определения прежде всего связана с тем, что под доминированием подразумевает не какой-либо поведенческий или структурный, параметр, а взаимовлияния между актами разных животных.
Когда речь идет о поведении, то мы можем определить его структуру по набору регистрируемых действий: ударов, укусов и прочее*, но когда мы рассматриваем доминирование, то приходится определять его как «относительное право» на выполнение определенных действий. Для оценки этого права важен не столько сам факт появления каких-то актов, сколько влияние действий одного животного на поведение другого.
В определении доминирующего и подчиненного животного А.Мэслоу (см. Введение) старался подчеркнуть только этот аспект — направление влияния. Но его определение не может претендовать на точность. Во-первых, при более строгом подходе ясно, что действия любого животного находятся под социальным контролем, и следовательно, могут «ингибироваться, ограничиваться и вызываться» партнерами по группе. Во-вторых, как было показано в дальнейших исследованиях, доминирование не сводится к подавлению поведения партнера: часто в поддержании этих отношений наиболее активен подчиненный, а не доминант ( Howell, 1966, 1974).
На практике исследователи исходят из частных определений, говорящих о результатах или непосредственных причинах процесса в конкретной обстановке. Выявление доминирования производится двумя основными способами: (1) анализом «конкурентной борьбы» между двумя или несколькими противниками и (2) анализом результатов «спонтанных» агонистических взаимодействий, возникающих в группе. Поэтому часто используются функциональные определения, специфические для этих двух ситуаций. Как правило, подразумевается, что доминирование обеспечивает «приоритет в доступе к источникам ресурса, в результате успешных атак, преследований или «вытеснения», которые происходят сейчас или были в прошлом» (Morse, 1974). Однако функции и причины, на описании которых строится *это определение, в основном связаны с вопросами «почему» и «зачем», а наш анализ был бы продуктивнее, если бы мы могли определить «что» за процесс мы имеем в виду.
В большинстве случаев под доминированием подразумевают определенные достаточно устойчиво сохраняющиеся формы асимметрии во взаимоотношениях.
«Понятие доминирования-подчинения применимо при описании дисбаланса взаимодействий у пары животных, если такие формы дисбаланса устойчиво проявляются и у других пар» (Hinde, 1978, с. 36).
Однако при необыкновенном разнообразии форм такой асимметрии и отсутствии единого, объясняющего их возникновение принципа, круг рассматриваемых взаимоотношений Р. Хайнд ограничивает только теми, которые исходно устанавливаются в результате агрессивных действий или «метакоммуникативных» сигналов, ассоциирующихся с ними». Это ограничение (помимо того, что постулирует если не сам механизм, то направление его поиска), еще более подчеркивает связь асимметрии во взаимоотношениях с «силовой» обусловленностью неравномерного распределения ресурсов. Однако это ограничение не позволило достаточно отчетливо определить искомый феномен, главным образом потому, что разные его параметры могут не коррелировать между собой. Поэтому определение часто сужают, сводя его к «асимметричному агонистическому взаимодействию» (Bernstein, 1981) или в более полной форме — к «стабильной асимметрии в агонистическом поведении двух или более животных» (Oewsbuff, 1982).
В этом определении обозначены три свойства доминирования:
1) доминирование основано на проявлении агонистических действий, 2) признаком доминирования является асимметричность этих действий, 3) доминирование установлено тогда, когда асимметрия стала стабильной. Если мы обнаружим существование такой асимметрии, то можем искать корреляцию с приоритетом в доступе к различным ресурсам. Этой корреляции может и не быть, так как приоритет — это важное, но не обязательное свойство доминантных отношений.
Предполагается, что это определение доминирования, как и прочие, полезно прежде всего эмпирически:
«доминантность полезна как сжатый термин, служащий для указания, что исход агонистических и конкурентных взаимодействий между двумя животными предсказуем на некотором практически достижимом уровне определенности» (Howell, 1974).
Однако и это определение не решает проблем. Во-первых, ни один из трех поясняющих его терминов не является достаточно определенным. Во-вторых, доминантный ранг (то есть эмпирическая оценка различий между животными, основанная на стабильной асимметрии отношений) может не коррелировать с приоритетом в доступе к ресурсам. Д. Дьюсбери считает, что это затруднение легко обойти — в таких случаях ранг просто маловажен для дележа рассматриваемых ресурсов. Однако сама борьба за ресурс может принимать стабильную форму агонистических взаимодействий. И направленность их может не совпадать при различных ситуациях. Третий момент, который Д. Дьюсбери считает маловероятным, — отсутствие корреляции между самими показателями доминирования, то есть между разными признаками агонистических взаимодействий. Однако и это допущение справедливо лишь до определенных пределов (см. раздел 5).
Отсутствие сколько-нибудь полного определения (описания) понятия доминирования является очень существенным моментом в его сегодняшнем использовании. С одной стороны очевидно, что мы имеем дело с чрезвычайно важным биологическим феноменом, если судить по «вездесущности» его проявления: мы можем достаточно четко формулировать частные задачи, чтобы считать понятие «доминирование» полезным эмпирическим термином, и такие задачи достаточно разнообразны и многочисленны, чтобы термин был необходим. С другой стороны, следует помнить, что при неясных и фрагментарных сегодняшних представлениях о биологическом процессе — основе доминирования, мы не можем говорить о полноценном общем определении, поиск которого превращается в «совершенно стерильное интеллектуальное упражнение» (Seyfarth, 1981).
Эмпирическое функциональное определение доминирования перестанет быть операциональным, если мы найдем общий принцип формирования стабильной асимметрии в поведении животных, частным отражением действия которого является этот феномен[1]. Мы пытались найти подход к постановке этой проблемы (Гольцман, 1977), но его обоснование [через идею «стимуляции подобного подобным» в поведении участников, т.ч. первоначальное повторение действий друг друга переходит в «соревновательность» в плане увеличения экспрессии этих действий, пока один из «состязающихся» не сдастся; тогда поведение обоих стен взаимно комплементарным, а роли — дифференцированными. См. современное развитие этой идеи М.Е.Гольцманом] представляет собой самостоятельную тему, выходящую за рамки настоящей статьи.
Отметим еще один существенный терминологический момент. Методологически важно разделять понятия доминантных отношений, возникающих у пары особей (так называемое диадное взаимодействие) и иерархию доминантных рангов (Hinde, 1978; Bernstein, 1981). Эти понятия не только не равнозначны, но и не всегда жестко связаны.
Доминантные отношения обозначают асимметрию, существующую во взаимоотношениях данной пары особей и подразумевают, что эта асимметрия возникла в результате влияния одной особи на другую. Иерархия доминантных рангов — один из аспектов описания структуры группы. Доминантный ранг отражает положение особи относительно ее партнеров и является не абсолютно присущим ей свойством, а ее положением на искусственной шкале.
Существование доминантных отношений еще не означает, что должна существовать иерархия доминантных рангов. То есть, если в группе существуют доминантные отношения, то не обязательно особей можно ранжировать в линейном порядке. Если А доминирует над В, а В над С, то из этого еще не следует, что А также доминирует над С. Это замечание является важным уточнением терминов. Дело в том, что во многих работах говорится о значении иерархий, например, для регулирования размножения или доступа к источникам ресурсов, но на самом деле авторы делят особей на две категории: доминантов и подчиненных, чаще всего подразумевая «имущих» и «неимущих». При этом об иерархии, как таковой, речи не идет.
Поскольку доминантные отношения могут проявляться в самых различных формах поведения, а ранжирование можно произвести лишь по одному параметру, то в зависимости от числа параметров и корреляции между ними можно выделить одну или несколько иерархических систем доминантных рангов. Абсолютная корреляция между разными системами может быть найдена лишь в исключительных случаях.
2.Логический статус доминирования
Эта проблема непосредственно вытекает из первой. Ряд исследователей считает, что доминирование является лишь промежуточной переменной (Hinde, 1974; 1978; Hinde, Datta, 1981; Baenninger, 1981; Brain, 1981). P. Хайнд (1978) так поясняет эту мысль:
«Предположим, что А бьет В больше, чем В бьет А; С бьет D больше, чем D бьет С. Кроме того, А обладает приоритетом в доступе к корму перед В, а С— перед D. В обыскивает А больше, чем А обыскивает В, a D обыскивает С больше, чем С—D, и т.д.
Мы можем упростить наше описание, рассматривая доминирование — подчинение как «промежуточную переменную» ( McCorquodale, Мееhl, 1954), которая предсказывает направление ударов, приоритет в доступе к пице и в обыскивании. Короче, А доминирует над В, а С — над D.
Если мы выделяем три или более независимых переменных, влияющих на отношение доминирования — подчинения (например, размеры, пол, знакомство с условиями), то использование промежуточной переменной даст экономию при описании, так как три независимых переменных могут быть связаны с тремя зависимыми девятью возможными связями, а через промежуточную переменную — шестью. Конечно, этот подход полезен лишь настолько, насколько зависимые переменные коррелируют между собой».
Однако интерпретация связей между независимыми, промежуточными и зависимыми переменными затрудняется существованием обратных связей между ними. Вследствие этого И. Бернстайн (1981) считает, что доминирование можно рассматривать не только как промежуточную, но и как независимую или как зависимую переменную, что определяется конкретной постановкой задачи. В частности, доминантный статус в иерархии может рассматриваться как независимая переменная.
Обладает ли концепция доминантности объяснительной силой или является лишь формой описания определенных корреляций между характеристиками поведения, которые мы наблюдаем? На этот счет существуют противоположные мнения (см., например, Bernstein, 1981; Candland, Ноег, 1981; Hinde, Datta, 1981). Эта проблема тоже возникла из-за отсутствия структурного определения. Если доминирование является гипотетической конструкцией, показывающей статистическую связь между зависимыми переменными, то ее нельзя использовать для объяснения этих характеристик. Сама статистическая корреляция ничего не может сказать о природе зависимых переменных — она ограничивается лишь оценкой степени их согласованности (Candland, Ноег, 1981). Отсюда логически вытекает заключение С. Альтманна:
«Доминирование — это наше изобретение, а не наше открытие» (Altmann, 1981).
-
Связь между доминированием и агрессией
Одно из положений классической концепции доминирования гласит, что доминирование устанавливается в результате агрессивных действий. В обзоре, посвященном социальному доминированию, У.К. Олли пишет:
«Может быть полезным определить то, что я подразумеваю под социальной иерархией. Термин «иерархия» применим к любому социальному ранговому порядку, устанавливаемому с помощью прямой борьбы, угроз, пассивного подчинения или некоторой комбинации этих поведенческих актов. Индивидуум, удерживающий первое место в иерархии, является доминантом, или господствующим животным в группе» ( Аllе, 1952).
Здесь возникают три проблемы. Если доминантные отношения целиком основаны на таких поведенческих актах, как прямые физические воздействия, и на оценке определенных физических черт — «метакоммуникативных сигналов», связанных с вероятностью агрессии и подчинения, то не означает ли это, что они являются лишь эпифеноменом агонистических взаимодействий? В таком случае мы можем искать их причины, но не можем говорить об их следствиях.
«Доминантные связи не имеют ни влияния, ни эффектов, ни функций, ни адаптивного значения. Они не могут влиять на успех при спаривании, на доступ к ресурсам, порядок передвижения или другие аспекты социального поведения или биологических процессов, хотя могут коррелировать с ними и прогнозировать их появление. Из этого следует, что вопрос о функции и адаптивном значении доминантных связей бессмыслен, при том, что соответствующие вопросы об агонистическом поведении правомочны… Существуют ли доминантные связи? Да, в уме и в записях наблюдателей. Подобно улыбке Чеширского кота, они абстрактны. Важны ли доминантные связи? Безусловно, но для исследователей, а не для животных» (Altmann, 1981).
На это утверждение можно возразить, что в концепцию доминирования включены, кроме самой агрессии, еще по крайней мере два механизма: оценки и обучения. Доминирование обозначает прежде всего направленность взаимодействий в связях между особями, а не количество элементов агрессивного поведения между ними (Hinde, 1978). Отношения доминирования-подчинения подразумевают установление такой связи, в которой открытая агрессия уменьшена или совсем исчезла. Таким образом, направленность устанавливается не только благодаря агрессии (или ассоциирующимся с ней сигналом), но и благодаря другим параллельно действующим механизмам, позволяющим «оценивать» (или «переоценивать») возможности противника и «обучаться» результатам действий. Однако следует учитывать, что и в сам процесс агрессии включены механизмы «оценки» и «обучения». Таким образом, разделение агрессии и доминирования; вопрос, решаемый лишь эмпирически.
Вторая проблема связана с тем, что, выявляя доминантные связи даже только на основе агонистических взаимодействий, мы далеко не всегда обнаруживаем линейное распределение рангов. Как правило, такие системы включают разнообразные «петли». Причина этого прежде всего в том, что рассматриваемые пары животных никогда не находятся в «социальном вакууме». Множество различных форм внешних социальных воздействий влияют на поведение партнеров.
И последняя проблема, на которой мы остановимся в этом разделе. Рассматривая доминирование как результат агрессивных действий, можно сделать вывод, что именно доминант подавляет или «модифицирует» поведение подчиненного. Между тем, в определении взаимоотношений между ними более активную роль могут играть подчиненные животные. Т. Рауэлл (Rowell, 1966), впервые подчеркнувшая этот аспект доминантных отношений, пишет:
«… иерархия поддерживается или выражается главным образом в демонстрациях подчинения и… именно благодаря низко ранжированным животным сохраняются различия в ранговых положениях… Именно подчиненные животные непрерывно следят за доминантом и активно поддерживают иерархическую структуру, так что доминирующим без особой натяжки можно назвать животное, которое «не задумывается перед тем, как что-нибудь сделать» во время социальных конфликтов» (Rowell, 1966).
Исходя из этого, Т. Рауэлл приходит к выводу, что термин «иерархия соподчинения» точнее описывает наблюдаемую картину, чем «иерархия доминирования» ( Rowell, 1974). Т. Рауэлл также указывает, что и для наблюдателя поведение, связанное с подчинением, часто значительно удобнее для описания, чем поведение доминанта. Некоторые нейроэндокринные корреляты ранга также легче найти у подчиненных животных. Эти наблюдения были подтверждены в ряде исследований (см. например,
Chance, 1967; Fox, 1971; Rowell, 1971; Deag, 1977; Lockwood, 1979). Именно поэтому в определении доминирования часто подчеркивается само установление асимметрии взаимоотношений, а не его причина.
В целом можно сделать вывод, что при применении концепции доминирования в описании поведения необходимо опираться на достаточно большой набор показателей поведения обоих партнеров и одновременно учитывать влияние внешней «социальной среды».
-
Адаптивное значение доминирования
Необходимо разделять два вопроса: адаптивно ли явление доминирования, и является ли доминант самым адаптированным животным в группе? На наш взгляд, постановка второго вопроса хотя и часто встречается, не вполне корректна. Как нельзя определить самого «сильного» или самого «умелого» из людей (очевидно это будет зависеть от поставленных перед испытуемыми задач), так же, видимо, нельзя найти и наилучшим образом адаптирующееся животное, если не понимать под адаптацией приспособление к частной ситуации. Даже физиологические «функциональные» системы одного организма, несравненно жестче интегрированные между собой, чем особи в популяции, достигают требуемого результата различными меняющимися в зависимости от ситуации средствами.
При обсуждении доминирования как результата адаптивной эволюции обычно выделяют три аспекта: (а) адаптивность социальной системы доминантных рангов; (б) адаптивность поведенческих механизмов, обеспечивающих установление доминантных связей; (в) адаптивность статусов доминанта и подчиненного. Какой именно аспект подчеркнут в той или иной работе, зависит обычно от того, какую точку зрения разделяет автор — сторонников группового или индивидуального отбора.
Если сформулировать точку зрения сторонников группового отбора, то конкурирующие группы, популяции и виды следует рассматривать как такой же материал для естественного отбора, как и отдельные организмы, и поэтому появляется возможность дать простое объяснение целому ряду сложных явлений (иерархическим системам, территориальности, авторегуляции плотности популяции и пр.) в социальном поведении животных: ход эволюции социальных систем можно рассматривать как аналогичный традиционному пути эволюции организмов. Однако при этом общая теория группового отбора сталкивается с важными теоретическими проблемами (Майр, 1974).
Если же ограничиться предпосылками теории отбора особей, то возникают проблемы при попытках объяснить развитие сложных социальных феноменов, исходя только из индивидуальной стратегии особей, направленной на достижение максимальной репродуктивной выживаемости.
В начале 60-х годов появилось несколько фундаментальных работ, обосновывающих каждую точку зрения, интенсивная разработка которых продолжается и сейчас.
Теория функций социального поведения, исходя из предпосылок группового отбора, в 1962 г. была развита В. Винни-3двардсом в книге «Расселение животных в связи с их социальным поведением» (Wynne—Edwards, 1962). Эта прекрасно написанная монография оказала большое влияние как на экологические, так и на этологические исследования.
В. Винни-Эдвардс считает, что
«… социальная иерархия или «право клевка» и их эквиваленты — это обычное и важное следствие «условной конкуренции[2]» Их функция — автоматически, как только возникает соответствующая ситуация, дифференцировать особей на «имущих» и «неимущих»… Например, если запасы корма использованы вплоть до какой-то установленной нормы, то «лишние» особи должны подчиниться закону условной конкуренции и не вступать в борьбу за ресурсы даже под угрозой неминуемой гибели. Эта система поддерживается, исходя из интересов популяции или вида, и безжалостно подавляет временные интересы отверженных, которые «приговариваются» к голодной смерти еще в период обилия корма» (Wynne—Edwards, 1962, с 18).
При таком рассмотрении на популяционном уровне можно выделить следующие основные функции иерархии доминантных рангов:
1) Понижение уровня агрессии благодаря упорядочению растре деления «условных наград». Эту функцию можно проиллюстрировать и экспериментом А. Гула и У. Олли (см. Введение), а также множеством данных, показывающих,что при формировании новых групп из незнакомых друг с другом животных, первоначальная вспышка агрессивных реакций угасает с одновременным установлением доминантных связей. Чем длительней период формирования доминантных отношений, тем дольше продолжается агрессивная борьба (напр. Struhsaker, 1967; Шилов, 1977);
2) Регулирование доступа «менее ценной» части популяций к ограниченным источникам ресурсов. Иерархия
«… не только может исключить часть популяции из размножения, но и ограничить доступ к корму, если его не хватает. В зависимости от обстоятельств, особи, находящиеся на нижних ступенях иерархической лестницы либо вообще исчезают, либо сохраняются как неучаствующий в конкуренции резерв, если это позволяют ресурсы» (Wynne—Edwards, 1963).
3) Авторегуляция плотности популяции. В поддержании популяционного гомеостаза иерархия по В. Винни-Эдвардсу в сущности является регулятором, механизмом, избавляющим от лишних особей. Этот регулятор непрерывно меняет свою функцию в зависимости от популяционного пресса, с одной стороны, и емкости биотопов — с другой (Wynne—Edwards, 1962, Шилов, 1977);
4) Согласно теории Дж. Кристиана (Christian, 1970) иерархия выявляет ту часть популяции, в которой повышена тенденция к миграции при высокой численности. Эту часть составляют подчиненные животные, тогда как доминирующие образуют «хорошо организованное» ядро популяции, которое остается в уже освоенной местности (см. также Calhoun, 1971; Sade, 1980).
Эти выводы о функциях иерархии сделаны из предпосылки, что естественный отбор происходит не только среди особей, но и среди конкурирующих групп и популяций животных.
«Иерархия как система взаимоотношений имеет бесчисленные варианты у представителей разных видов и разных систематических групп, аналогично тому, что мы обнаруживаем в отношении нервной и кровеносной систем. Подобно этим системам, иерархия подвергается адаптивным эволюционным изменениям под давлением естественного отбора» (Wynne—Edwards, 1978).
Таким образом, В. Винн-Эдвардс рассматривает иерархию как «орган» социальной группы, адаптивно меняющийся в процессе эволюции. Т. Клаттон Брок и П. Харви (1976, 1978) выдвигают возражение против указанного подхода на том основании, что в этом случае
«… иерархия рассматривается как поведенческая черта, тогда как она не более, чем «статистическое следствие компромисса, в который вступает каждая особь при конкуренции за корм, полового партнера или другие ресурсы. Каждый компромисс адаптивен, но это не значит, что, адаптивна их статистическая суммация (Williams, 1966)» ( Clutton — Brock, Harvey, 1976).
В сущности, это не возражение, а постулирование другой точки зрения, основанной на том, что структура доминантных отношений рассматривается как результат взаимовлияния особей в борьбе за повышение индивидуальной приспособленности. Две основные проблемы, возникающие при этом: каким образом такая «статистическая суммация» приобретает структурированный вид (то есть проблема самоорганизации связей в сообществе и внешних факторов, которые на нее влияют) и как объяснить те черты поведения, которые «невыгодны» для особи, но, видимо, полезны для существования группы. Эти проблемы неоднократно рассматривались в литературе (например, Haldane, 1932; Шмальгаузен, 1939).
На современном этапе их обсуждения большую роль сыграли работы В Л. Гамильтона (Hamilton, 1963, 1964), разработавшего концепцию «отбора родичей» и Дж. Мэйнарда Смита (Maynard Smyth, 1974), предложившего систему понятий для анализа процессов конкурентной борьбы.
Доминантные отношения между двумя конкурирующими партнерами рассматриваются как суммарный показатель (и регулятор) их индивидуальных оценок полезности ресурсов и потенциальных затрат на их получение. Каждый ресурс (половой партнер, корм или жизненно необходимое пространство) обладает определенной ценностью для выживания особи, но конкурентная борьба за него связана с затратами энергии и риском получить ранение, то есть в конечном итоге со снижением приспособленности. Исходя из этого, можно найти «экономическую» основу для ответа на вопрос: выгодна в данной ситуации конкурентная борьба или нет? Если возможная выгода велика, а цена невысока, то особь вступает в конкурентную борьбу. Если же, потенциальные затраты существеннее, чем польза от победы, то от борьбы лучше воздержаться и попытаться найти ресурс в другой раз или в другом месте (Emlen, 1973; Maynard Smyth, 1974; Parker, 1974). Вследствие того, что индивидуальные способности к конкурентной борьбе различны и индивидуальные оценки ресурсов не совпадают, в отношениях между конкурентами возникает асимметрия.
Можно выделить три типа асимметрии (Maynard Smyth, Parker, 1976):
(1) асимметрия из-за «стоимости» ресурса, когда для одного из конкурентов победа важнее, чем для другого;
(2) асимметрия в способностях к борьбе или в возможностях удержать ресурс, возникающая в результате различий между особями, например, в силе, опыте или в размерах; и
(3) безотносительная асимметрия, не связанная ни с относительной стоимостью, ни с различиями в способностях к борьбе (например, преимущество владельца территории).
Таким образом, перед вступлением в борьбу противники должны оценивать асимметрию в заинтересованности в победе и соотношение своих возможностей. Способности производить такую оценку, по-видимому, адаптивны, а надежные признаки, по которым она может быть сделана, полезны и для подающего, к для воспринимающего сигнал (Dawkins, Krebs, 1978).
Если легко различимой асимметрии нет, то между незнакомыми животными разгорается конкурентная борьба, в ходе которой соперники обучаются взаимной оценке. Таким образом, доминантные связи могут устанавливаться на основе обучения оценке асимметрий и индивидуального распознавания. Большое значение при этом имеет использование надежных признаков, коррелирующих с асимметрией. Если эти признаки выработаны в процессе отбора, то они могут быть надындивидуальными характеристиками, связанными с полом, возрастом, размерами и агрессивностью животных. Такие признаки, используемые для оценки социального статуса, описаны у приматов (Altmann, 1962), псовых (Schenkel, 1947; Fox, 1971), копытных (Walther, 1977: Geist, 1978, Соколов, Данилкин, 1981), грызунов (Гольцман с соавт., 1977) и представителей других групп.
Существенное влияние на рассмотрение процесса конкурентной борьбы оказали простые модели, основанные на теории игр. Дж. Мэйнард Смит доказал, что в популяции может существовать эволюционно-стабильная стратегия поведения, то есть такая стратегия, при которой (если она принята большей частью особей популяции) никакая другая из возможных стратегий не обеспечивает столь же высокой репродуктивной приспособленности. Так, например, если большинство животных отвечает мирной условной демонстрацией на демонстрацию, а на открытую агрессию реагирует дракой, то они не будут вытеснены ни «голубями», которые только демонстрируют, ни «ястребами», сразу вступающими в бой.
Кроме того, Дж.Мэйнард Смит обсудил возможность появления сбалансированной стратегии, когда сохраняется возможность нескольких решений одной и той же проблемы. Вероятности их появления соотносятся в определенной сбалансированной пропорции и любое нарушение, уменьшающее частоту проявления одной стратегии будет приводить к компенсирующему отбору животных, использующих эту стратегию. Таким образом, может существовать ряд альтернатив, при которых, например, максимальный репродуктивный успех при заданных индивидуальных характеристиках достигается без агонистического доминирования.
Так, И. Бернштейн (1981) приводит пример из сообщения Т. Клаттон-Брока на Международном этологическом конгрессе 1977 г. о «клепто-репродуктивной» стратегии у самца благородного оленя. Самец оленя успешно спаривался с самками, пока хозяин гарема отвечал на вызовы других самцов. Пользуясь этой тактикой, самец мог оставить не меньше потомства в разных стадах, чем защищающий своих самок владелец гарема. Более того, «гастролер» не рисковал получить какие-либо повреждения от схваток с соперниками, а значит, мог дольше сохранять свои силы для размножения. Такое поведение может например, приносить успех молодым животным, не достигшим расцвета сил. Хотя на подобные отклонения от «нормы» пока обращали недостаточно внимания (однако см. Le Boeuf, 1974; Hausfater, 1975; Packer, 1979; Rubenstein, 1980) они представляют собой скорее правило, чем исключение.

Благородный олень Cervus elaphus, самец
Конечно, модели, подобные моделям Дж. Мэйнарда Смита, упрощены и для анализа реальных ситуаций неприменимы (см., например, Caryl, 1981), однако они привлекли особое внимание исследователей к проблеме поведенческого полиморфизма, то есть существования набора поведенческих альтернатив, а это заставляет искать новые подходы к анализу социальных структур (Bateson, Klopfer, 1981). Таким образом, доминирование, если пользоваться терминологией Дж. Мэйнарда Смита, можно рассматривать как эволюционно стабильную стратегию поведения, но не единственную, а одну из многих возможных.
Третий подход к оценке адаптивности доминирования сводится к сопоставлению биологических особенностей статусов доминанта и подчиненного (Alcock, 1975; Wilson, 1975; Hausfater, 1975; West—Eberhard, 1975; Hrdy, 1976; Jones, 1980; Slater, 1981). Если исходить из того, что при конкурентных отношениях доминирующие животные чаще получают «приоритет в доступе» к источникам ресурсов или добиваются репродуктивного успеха, то преимущества, связанные со статусом доминанта, представляются очевидными.
При обсуждении биологии подчиненных особей обычно придают принципиальное значение ряду факторов, компенсирующих подчиненное положение. Такими факторами считают следующие.
-
Подчиненное состояние может быть лишь временным явлением на жизненном пути особи. Например у большинства видов доминантный статус в той или иной степени коррелирует с возрастом, а у многих животных варьирует на протяжении одного репродуктивного сезона.
-
У социальных видов подчиненные особи делят все преимущества группового образа жизни (например, Hamilton, 1971 а, в). Например, у некоторых видов они лучше защищены от хищников, эффективнее добывают корм, легче находят полового партнера и реже гибнут от случайных причин, чем если бы они жили поодиночке.
-
Подчиненная особь может успешно адаптироваться к поведению доминанта и получать доступ к ресурсу, не вступая в агрессивную борьбу с ним[3]. Как заметил по поводу макаков-резусов Дж. Кауфманн (1967):
«Некоторые обезьяны своей навязчиво демонстрируемой покорностью обеспечивают себе прекрасную жизнь».
Подчиненные особи играют ключевую роль в дисперсии генов между популяциями, если справедливо предположение, что в основном именно они становятся мигрантами (Wilson, 1975; Шилов, 1977).
Как мы уже отметили (с. 85), Дж. Кристиан ( Christian, 1970) придает подчиненным животным еще большее значение. Если бродячие особи чаще становятся пионерами, осваивающими новые места обитания, то, именно они испытывают эффективность новых адаптаций. Однако вполне справедливо замечание Э.О. Уилсона:
«… эта гипотеза привлекательна, но она остается спекуляцией. Столь же легко построить модель, в которой: наибольшую ценность для эволюции будет представлять установившийся центр популяции. В центре мы находим наибольшее генетическое разнообразие. Местообитания здесь наиболее продуктивны и экологически разнообразны. Тут существуют наиболее плотные и стабильные популяции с наиболее разносторонними социальными взаимодействиями между животными» (Wilson, 1975, с. 290).
-
Концепция «совокупной приспособленности», основанная на теории отбора родичей, дает основание полагать, что подчиненные особи в группе, даже если они исключены из размножения, могут оказаться в более выгодном положении в отношении пресса отбора, чем особи, ведущие одиночную жизнь, так как могут содействовать выживанию своих ближайших родственников.
Е. Вильсон (Wilson, 1975) приписывает доминантному порядку еще ряд специальных свойств, определяющих роль доминирования в социальной организации. Кратко эти свойства таковы:

Cervus elaphus, оленуха
(1) Принцип ксенофобии. Принцип заключается в том, что, при установившейся системе взаимоотношений, появление чужой особи грозит нарушением балансу связей в группе.
Е. Вильсон объясняет этим резко агрессивную встречу «чужаков» у большинства исследованных видов. Этот тезис представляется нам довольно сомнительным, поскольку подобная ксенофобия может и не быть следствием доминантного порядка.
(2) Мир в результате сильного контроля. Во многих сообществах (особенно приматов) доминирующие особи могут силой подавлять драки между членами группы, однако, здесь также следует отметить, что животные, играющие «контролирующую роль», не всегда являются доминантами.
(3) Тяга к повышению социальной силы. У многих видов млекопитающих самцы на протяжении жизни занимают множество промежуточных статусов, пока не достигают положения доминирующего самца в репродуктивной группе. Например, это характерно для многих видов макаков, тонкотелов, павианов, ушастых тюленей, копытных;
(4) Социальная инерция. Становление организации в группе включает многие формы обучения и привыкания, поэтому в целом такая стабилизированная система обладает определенной инерцией. В сущности именно инерция является основой стабильности социальных структур и возникновения сложных форм взаимосвязей в группе, проявляющихся, например, во влиянии связи на связь или же «престиже» доминанта, которые мы будем обсуждать в следующем разделе.
(5) Иерархия групп. Объединение партнеров может резко повысить их социальную мощь. Поэтому объединенные действия «клик», «кланов», «временных союзов» в борьбе за положение внутри группы являются обычным явлением по крайней мере у приматов.
Мы провели разделение взглядов сторонников группового отбора и отбора особей, чтобы четче сформулировать различия в постановке проблемы. Однако ясно, что в принципе могут идти одновременно оба процесса и основная трудность заключается в оценке соотношения их влияния. Большинство исследователей считает, что групповой отбор имеет меньшую селективную силу, чем ему придавал В. Винни-Эдварс. В то же время, некоторые эффекты группового отбора получили возможное объяснение благодаря теории «отбора родичей», разработанной В.Д. Гамильтоном (Hamilton, 1963, 1964) почти одновременно с публикацией идей В. Винни-Эдвардса (1962). Зачастую между понятиями группового отбора и отбора родичей (хотя последний можно рассматривать как форму индивидуального отбора) трудно провести границу[4].
Е. Вильсон, в монографии которого применение идей отбора родичей к исследованию сообществ животных является центральной темой, считает, что
«чистый отбор родичей и чистый отбор демов являются полюсами, связанными градиентом, при все увеличивающихся наборах родственных особей» (Wilson Е., .1975).
Подобную же оценку этих двух процессов можно найти и у Д.С. Вильсона (Wilson D.S., 1975)
«… традиционные концепции группового и индивидуального отбора, по-видимому, являются двумя крайностями одного континуума».
Макак-резус Macaca mulatta

- Проблема выбора параметров для оценки доминирования
При описании любого феномена выбор параметров имеет большое значение, но когда этот феномен столь ненадежно определен как явление «доминирования», этот выбор становится решающим. Описание и анализ доминирования велся в двух основных и взаимосвязанных направлениях: 1) экспериментальный анализ поведения в «конкурентных ситуациях» и 2) непосредственные исследования социальных взаимодействий в искусственных и естественных группах животных.
Предпосылкой для развития первого направления были данные о том, что средством для установления доминантных отношений являются агрессивные взаимодействия. Так как непосредственная регистрация естественно возникающих агрессивных действий требует больших затрат времени и не всегда возможна, были разработаны методики, позволяющие провоцировать агрессивные стычки в так называемых «конкурентных ситуациях».
Эти методики были положены в основу большой серии экспериментальных исследований, которые интенсивно ведутся до сих пор. В связи со спецификой подхода, авторы этих работ определяют доминирование как
«приоритет в доступе при приближении (например, к пище или к воде) или в уходе при избегании (например, электрического шока), которым одно животное обладает перед другим» (van Kreveld, 1970).
Чтобы определить приоритет, используют два основных методических приема. Типичные примеры применения первого из них (моделирование ситуации ограниченного доступа к источнику корма) можно найти в исследованиях A.H.Maslow (1936 а,с), R.H.Bruce (1941), W.T.James (1949), P.L.Baenninger 1970), D.L.CIark, J.E.Dillon (1974). Так, в работе P. Брюса показателем доминирования у крыс было поведение в конкурентной борьбе за корм или воду. В этих опытах двум голодным животным одновременно открывали доступ к корму (или к воде) и та крыса, которая на большой срок оттесняла другую, считалась доминирующей. Впоследствии подобную методику применяли многие авторы, модифицируя различные условия эксперимента: вид подкрепления, длительность депривации, продолжительность эксперимента и т.п. Некоторые авторы в аналогичных опытах в качестве показателя доминирования использовали не общее время контроля над доступом к корму, а другие показатели, — например, количество съеденной пищи (Hoyenga, Rowe, 1969).
Другим широкораспространенным методическим приемом являются тесты, построенные по принципу «все или ничего» (Syme, 1974), В этих тестах двух соответствующим образом мотивированных (например, голодных) животных заставляют бороться за возможность выполнения действия, приводящего к одноразовому подкреплению. Так, при работе с грызунами часто применяют «трубу доминирования» (Lindzey et al., 1961; Schumsky, Jones, 1966; Messeri et al., 1975) — узкую трубу, в которую с двух сторон одновременно запускают животных. Чтобы достигнуть предварительно обусловленного подкрепления, каждый из конкурентов стремится вытолкнуть противника назад.

Хотя исходная предпосылка при разработке различных методических приемов состояла в том, что все проявления приоритета в группе отражают единый феномен, оказалось, что порядки доминирования, выявляемые разными способами, зачастую плохо коррелируют между собой. Так, в ряде работ обнаружена слабая корреляция между системами доминирования, определяемыми при применении разных видов подкрепления (Boice et al.,1969; Syme, Pollard, 1972). Например, К. Гэртнер с соавт. (Gärtner et al., 1981) сообщает, что порядок доминирования, выявленный в тестах при конкуренции за спаривание у лабораторных крыс, не коррелировал с другим, выявленным на основе тестов при конкуренции за воду. В других работах обнаружена слабая корреляция между системами доминирования в тестах с одинаковым подкреплением, но учитывающих разные поведенческие акты (например, разные инструментальные реакции) (Syme, 1974).

Параллельно выяснилось, что устойчивые картины, найденные при использовании одного теста с одним типом подкрепления, справедливы лишь для постоянных периодов депривации. Повышение мотивационного уровня у подчиненных животных приводит к усилению их конкурентоспособности ( Bruce, 1941; Hsaio, Schreiber, 1968j Syme, Pollard, 1972; Иванов, 1980). Кроме того, исследования показали, что взаимодействия в конкурентных ситуациях при экспериментальных ограничениях не эквивалентны агрессивным взаимодействиям, «спонтанно» возникающим в группах, которые эти опыты призваны моделировать (Winslow, 1938; Baenninger, 1970; Drews, 1973; Gage, 1978).
Д. Бентон с соавт. (Benton et al., 1980) исследовали степень корреляции разных параметров, используемых для оценки доминирования у лабораторных мышей. В их опытах «доминирование» определялось следующими способами:
(I) выявляли особей, принимающих позу подчинения;
(II) проводили подсчеты числа ран на хвостах у противников в конце периода наблюдений;
(III) измеряли время, проведенное возле бутылочки с водой в первые две минуты ее предъявления после периода депривации;
(IV) сравнивали результаты ссаживания членов группы со «стандартными» противниками в «трубе доминирования»;
(V) сопоставляли результаты подсаживания незнакомых «стандартных» противников в камеру, освоенную одним из членов группы (тест на «территориальную агрессию»);
(VI) анализировали результаты ссаживания испытуемых самцов с самками в состоянии эструса;
(VII) сравнивали результаты взвешивания различных желез испытуемых самцов в конце серии экспериментов.
Мыши, доминировавшие в группе (I тест), имели мало ран на хвосте, больше дрались в тесте «защиты территории» и проявляли повышенный интерес к рецептивным самкам. Однако факторный анализ корреляционных матриц не выявил какого-либо общего фактора во II — VII тестах, который можно было бы обозначить как доминирование. Результаты тестов при конкуренции за воду и опыты с «трубой доминирования» не коррелировали с результатами первого теста. Измерение веса желез тоже не дало возможности различить доминирующих и подчиненных животных. Д. Дрюс и К. Дики (Drews, Dickey, 1977) провели аналогичные исследования с лабораторными крысами и обнаружили подобное же отсутствие корреляции результатов различных тестов.
Таким образом, оказалось, что результаты применения текстов с «конкурентными ситуациями» при выяснении внутригрупповой структуры доминантных отношений истолковывать весьма не просто, и в дальнейшем периодически повторялись попытки более четко установить границы их применимости и обосновать методические процедуры (van Кreveld, 1970; Richards, 1974; Syme 1974; Benton et at.,1980; Иванов, 1980).
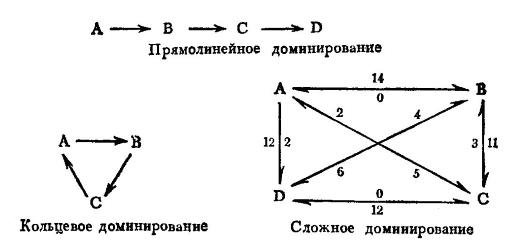
Системы ранжирования в разных популяционных группировках. Буквами обозначены отдельные особи, цифрами — число случаев доминирования. (По Наумову и Карташеву, 1979.) Это стандартный взгляд, критикуемый в данной статье
В целом можно сделать вывод, что наиболее взаимосвязана группа параметров, основанных на спонтанных агонистических действиях, которые можно обозначить как «агонистическое доминирование» (Bernstein, 1981; Brain, 1981). Параметры, основанные на разрешении «конкурентных ситуаций» часто не коррелируют ни с ними, ни между собой.
Второе направление исследований социального доминирования — анализ взаимоотношений в естественных и искусственных группах животных. Если в рамках ранее описанного подхода производится оценка доминантных отношений в изолированных парах специально подобранных животных в одном или нескольких тестах, моделирующих стандартные ситуации, то при втором подходе исследуются взаимоотношения в целой группе животных, так что в центр внимания попадают иерархические системы доминантных рангов. Естественно, что анализ социальных влияний в этом случае резко усложняется из-за большого количества участвующих животных и из-за разнообразия взаимодействий между ними.
Следует подчеркнуть изменения, происшедшие в представлении о значении и структуре иерархии доминантных рангов. В ранних работах считалось, что доминантный ранг отражает степень господства во всех областях социальной жизни, что его выявление можно произвести к любому удобному параметру и что иерархия доминантности является основой социальных систем. В современных работах иерархия доминантных рангов используется в основном в качестве описательного понятия, отражающего одну из многих характеристик сообщества. Это ограничение вызвано прежде всего частым отсутствием корреляций между группами параметров, связанных с «агонистическим поведением», с конкурентными взаимодействиями, с «положительными» социальными контактами и с репродуктивным поведением. Исходя из этого, описание иерархической системы доминантных рангов часто ограничивают только первой группой параметров. Было показано, что такие иерархические структуры можно выявить у многих видов млекопитающих, хотя в большинстве случаев их изучали в сообществах приматов (Alexander, Bowers, 1969; Tokuda, Jensen,1969; Bernstein, 1969, 1970,1976; Hinde, 1974; Rowell, 1974; Hausfater, 1975; Dunbar, 1980; Jones, 1980, 1981), и что в некоторых сообществах иерархические структуры могут оставаться стабильными в течение нескольких месяцев, а иногда и лет ( Bernstein, 1969, Eaton, 1976; Seyfarth, 1976, 1978).
Обычно описание иерархических систем ведется на основании оценки взаимоотношений каждого члена группы с каждым из его партнеров при последовательном переборе всех возможных сочетаний пар. Иногда применяются и суммарные показатели, такие как отношение числа побед к общему числу взаимодействий. Выбор такого показателя, также как и метода статистической обработки, влияют на выявленный порядок ранжирования, однако общая картина в целом, как правило, сохраняется достаточно постоянной. В качестве примеров описания иерархических структур мы рассмотрим результаты двух работ, в которых иерархические системы доминантных рангов были выделены с помощью применения многофакторного анализа при большом наборе параметров.
В первой из них (Lockwood, 1979) были проанализированы результаты трехлетней регистрации поведения волков в семи группах. Использовали широкий спектр показателей — от «эмоциональности», определяемой в экспериментах, до физических характеристик животных и разнообразных параметров взаимодействий (всего 27 переменных). По результатам были построены корреляционные матрицы и методами факторного анализа выделены наборы связанных переменных, объединяемых общим гипотетическим «фактором». Из пяти выделенных таким образом групп наибольшее количество переменных связал фактор, условно названный «доминантностью». Основными переменными в этой группе были: направленность агрессии (процент агонистических взаимодействий, в которых данная особь была инициатором); результаты конкуренции за корм; вес тела; направленность поз и актов «подчинения» и частота оставления запаховых меток.
Интересно, что фактор «агонистическое поведение» оказался отделен от фактора «доминантности», так как показатель абсолютного уровня агрессивных форм поведения оказался слабо связан с последним. Таким образом, было подтверждено, что при определении доминирования основное значение имеет направленность, а не частота агонистических актов. Высокая корреляция между переменными, традиционно связываемыми с понятием «агонистического доминирования», позволила сделать вывод, что эта концепция полезна для описания поведения волков. В то же время автор подчеркнул, что речь идет о суммарных оценках, иными словами, если какое-то животное в группе не маркирует или не достигает успеха в конкурентной борьбе за корм, то у нас еще нет оснований для вывода, что данная особь находится на нижних ступеньках иерархической лестницы:
«каждый волк — это индивидуальность, и каждого из них необходимо рассматривать в полном спектре поведенческих характеристик».

Аналогичный подход был применен и для анализа структуры группы шимпанзе в Арнемском зоопарке (Noe et al., 1980). Было выделено 15 показателей, большинство из которых оказались связаны с агонистическими и конкурентными отношениями. Связи между этими показателями анализировали с помощью нескольких разных статистических приемов, основанных на факторном и кластерном анализе. Оказалось, что можно выделить по крайней мере три главные группы переменных:
(1) «конкурентное доминирование», связанное с поведением в двух типах конкурентных ситуаций: при «социальной конкуренции» за контакт с партнером (такой, например, как игра с детенышем) и в случае «пространственной конкуренции» во взаимодействиях типа «подход — избегание»;
(2) «агонистическое доминирование» — фактор, связывающий самую большую группу переменных: разнообразные угрозы, погони, акты «подчинения» и бегство;
(3) «демонстрация превосходства» — фактор, объединяющий специфические демонстрации и серьезные драки. Второй и третий факторы оказались теснее связанными между собой, чем с первым. Эта картина была подтверждена и результатами кластерного анализа.
Слабая корреляция между первой группой параметров и двумя остальными обусловлена тем, что в «неагонистических конкурентных ситуациях» (1-й фактор) самцы обычно уступают взрослым самкам, хотя доминируют над ними при агрессивных стычках (2-й и 3-й факторы). Однако, если рассматривать только взаимодействия между взрослыми самцами или между взрослыми самцами и неполовозрелыми самками, то оказывается, что все параметры доминирования коррелируют между собой. Таким образом, можно сделать вывод, что выявление иерархических доминантных рангов при точном определении параметров и правильно выбранном методе сбора материала является удобным методом описания структуры определенных форм взаимоотношений.
Однако следует учитывать, что выявление взаимосвязанных переменных еще не означает, что в исследованных группах иерархия имеет линейную форму, которую можно было бы ожидать, исходя из предположения, что она устанавливается на основе агонистических взаимодействий независимо в каждой паре животных (Chase, 1974). Р. Локвуд в вышеописанной работе проверил линейность рангового порядка в исследуемых стаях волков и пришел к выводу, что показатели линейности далеко не достигали установленного критерия (табл. 1),
Таблица 1
Линейность агонистического рангового порядка в стаях волков
| Стая (год) | Коэффициентлинейности | Стая (год) | Коэффициентлинейности |
| I (1972) | 0,10 | IV | 0,20 |
| I (1973) | 0,30 | V | 0,20 |
| I (1974) | 0,30 | VI (1972) | 0,65 |
| II (1973) | 0,50 | VI (1973) | 0,17 |
| III | 0,69 | VII | 0,54 |
Примечание: критерием линейности является достижение коэффициентом величины 0,9 (по Lockwood, 1979)
Ниже мы рассмотрим факторы, обусловливающие отсутствие линейности иерархических систем и сложность сети взаимоотношений «доминирования-подчинения «.

5.1. Взаимовлияние связи на связь
Методы, используемые при анализе социальных структур, рассчитаны прежде всего на выявление связей между каждой парой особей в группе и, как правило, мало пригодны для выявления тех влияний, которые оказывают отношения животного А с В на его взаимоотношения с С. Между тем, в реальном сообществе (и не только приматов, на которых подобные эффекты в основном продемонстрированы), эти «связи второго порядка» имеют очень большое значение (Hinde, 1974). Одно из наиболее часто описываемых проявлений такого эффекта лежит в основе так называемых «зависимых рангов».
Исследуя поведение японских макаков М. Каваи (Kawai, 1965) показал, что, кроме основного ранга, зависящего только от собственных возможностей животного, можно выделить и «зависимый ранг», то есть ранг, определяемый социальными связями и умением использовать социальную среду.
Например, самец, защищающий определенную самку, в соответствии с действенностью своей опеки повышает ее ранг. Даже высокоранговые самки уступают в конкурентных ситуациях самке низкого ранга, если поблизости находится ее защитник (Kawai, 1965; Takahata, 1982). При конфликтах в стаде японских макаков самцы вожаки берут под защиту самок, детенышей и подростков и наказывают агрессоров (Watanabe, 1979).
Сходный эффект наблюдается и при возникновении так называемых «коалиций» — объединение животных примерно одного ранга, резко увеличивает их социальный вес (например, Watanabe, 1979; Dunbar, 1980). В исследованиях Т. Струзакера ( Struhsaker, 1967) показана распространенность коалиций в стадах зеленых мартышек (Cercopithecus aethiops). Примерно в 20% всех интенсивных стычек, происходящих в группе, вовлекалось по несколько животных, приходящих на помощь зачинщикам. Такие коалиции могут действовать как довольно стабильные единицы и возникновение их может приводить к глубоким перестройкам группы. Так, в небольшой колонии яванских макаков (М. fascicularis) доминирующая самка (в момент ослабления связи с самцом) была свергнута объединившимися подчиненной самкой и ее старшей дочерью, после этого «переворота» произошла реверсия рангов потомков обеих самок (Chance et al., 1977). Сейчас накопилось множество аналогичных примеров, показывающих как животные могут активно использовать социальную среду для повышения своей «социальной силы» (Deag, Crook, 1971; Gil mo re,, 1977, Taub, 1980).

Макак яванский или крабоед Macaca fascicularis
Умение оценивать и использовать социальные связи в значительной мере зависит от индивидуального опыта. Показательны эксперименты, проведенные в Калифорнийском университете. Были сформированы две группы макаков-резусов. Одна состояла из обезьян, выращенных в изоляции с «искусственными матерями» (муляжами), члены другой группы росли со своими матерями и сверстниками. В обеих группах экспериментаторы искусственно создавали конкуренцию за пользование бутылочкой с водой. Оказалось, что если выращенные в изоляции обезьяны формируют относительно стабильный порядок доступа к бутылочке, то у «социально опытных» этот порядок изменчив и зависит от ситуации.
Изменчивость обусловливается прежде всего умением использовать отношения между партнерами. Например, обезьяна могла заставить владельца бросить бутылочку, обратившись за помощью к доминирующему животному, якобы для «разрешения агрессивного конфликта» или просто отобрать ее, объединившись с кем-нибудь из компаньонов. Она могла также отвлечь владеющего бутылочкой доминанта просьбой защитить ее в специально спровоцированной ею самой драке и воспользоваться бутылочкой в этот момент (Anderson, Mason, 1978).
«Зависимый ранг» и умение пользоваться социальной ситуацией могут влиять на формирование «основного ранга». Эта тенденция прослеживается и в процессе установления социального статуса детенышей у макаков и павианов (Кawаmura, 1965; Sade, 1972; Seyfarth, 1976; Berman, 1980, 1982; Schulman, Chapais,1980). Например, у японских макаков и макаков-резусов социальный статус дочери зависит от положения матери. При этом ранги взрослых дочерей связаны обратной зависимостью с их возрастом: при достижении самкой четырехлетнего возраста она приобретает ранг, непосредственно следующий за рангом матери, то есть поднимается по иерархической лестнице выше своих сестер. Обычно это объясняют её более тесной связью с матерью, которая больше помогает младшей из взрослых дочерей. Предполагается, что такую стратегию обусловливает самый высокий репродуктивный потенциал у самой молодой из взрослых самок (Schulman, Chapais, 1980).
Однако такое объяснение, видимо, сильно упрощает дело и феномен нельзя обосновать коалицией матери с младшими дочерьми (Hinde, 1974; Sade, 1980). Некоторые авторы указывают, что при анализе драк между сибсами нельзя сказать, что младший получает больше помощи от матери (Kaplan, 1977; Sade, 1980), Еще удивительнее то, что в некоторых случаях (в искусственной колонии на о. Кайо Сантьяго) молодые самки достигают своего предсказуемого ранга через несколько лет после смерти матери (Sade, 1980), что совсем не укладывается в общепринятое объяснение. Поскольку при выкармливании чужого детеныша он так же, как и ее собственный, обычно принимает ранг приемной матери, ясно, что генетические механизмы здесь не при чем. Д. Сейд предполагает, что в первый год жизни происходит какое-то обучение, напоминающее импринтинг, результаты которого проявляются только тогда, когда животное (в данном случае самка) взрослеет (Sade, 1980).
Трудности при анализе становления статуса молодой самки макака связаны прежде всего с проблемой взаимовлияния социальных связей (см., например, Hinde, Spencer-Booth, 1967; Harlow, 1969; Simonds, 1974; Cheney, 1978; Berman, 1980; 1982). С момента появления детеныша его взаимоотношения с матерью складываются под влиянием присутствия других животных и, в первую очередь, «теток» внимание которых он постоянно привлекает. Поэтому, например, когда мать и детеныш живут в группе, то мать меньше отпускает его от себя, чем если бы они жили в изолированной клетке (Hinde, Spencer-Booth,1969). Насколько мать разрешает другим самкам общаться с детенышем, зависит от ее отношений с ними. Чаще всего она позволяет это своим взрослым дочерям и самкам из своего клана. При этом действенность ее протестов в определенной степени зависит от ее взаимотношений с самцами. Отношения самца с данной самкой могут повлиять и на взаимоотношения матери с этой самкой, а в конечном итоге — и на взаимоотношения матери с детенышем (Hinde, 1974).
У макаков-резусов кланы как единицы дифференцируются по своему иерархическому статусу и в высокоранжированных кланах детеныши более изолированы от контактов с чужими особями, что также накладывает свой отпечаток на дифференциацию систем социальных связей (Berman, 1980, 1982). Социальные влияния могут быть настолько дифференцированы и сильны, что в двух искусственных колониях на островах в Пуэрто-Рико самки из высокоранжированных кланов раньше достигают половой зрелости, чем самки из средне- и низкоранжированных кланов (Drickamer, 1974; Sade et al. (l976).
5.2. Зависимость доминирования от места и времени взаимодействий
Различия между социальными организациями у животных, г держащихся тесной группой на одном участке обитания и на индивидуальных или на семейных территориях, обладатели которых «доминируют» в стычках с пришельцами, часто не столь уж отчетливо выражены (Fisler, 1969; Kaufmann, 1971). Как правило, возможности формировать социальные связи весьма пластичны и изменение экологических условий влечет за собой изменение социальной организации. Например,
М. Кларк и В. Галеф (Clark, Galef, 1977) описали, как агути (Dasyprocta punctata), обычно ведущие одиночный образ жизни в периоды нехватки корма, формируют доминантную иерархию в местах его концентрации.

Агути Dasyprocta punctata
П. Лейхаузен (Leyhausen, 1965, 1971), рассмотревший такие случаи и тщательно изучивший взаимоотношения в группах домашних кошек, разработал схему, ставшую теперь классической. Он пришел к выводу, что существует дихотомия социальной иерархии: одна иерархическая система — система «относительной иерархии», права доминирования в которой зависят от места и времени встречи (например, одни животные могут использовать тропы утром, другие — вечером и в это время они доминируют над всеми, оказавшимися на их пути), вторая иерархическая система — это жесткая «абсолютная иерархия». возникающая лишь в определенных местах (например, у кормушек, мест отдыха и т.п.).
Обе эти иерархические системы могут действовать одновременно, а баланс между ними определяется экологическими факторами, в первую очередь плотностью популяции. «Взаимодействие этих двух систем и возможность смещения центра тяжести от одной системы к другой, лежит в основе способности многих видов вести как одиночную территориальную жизнь, так и создавать небольшие и даже довольно большие группы» (Leyhaysen, 1965).

Японский макак Macaca fuscata
5.3. «Право обладания»
Когда самец шимпанзе добывает редкое лакомство — мясо, остальные члены стада окружают его, выпрашивая свою долю. Доминирующие животные ведут себя при этом так же, как и остальные, не пытаясь отобрать добычу (Teleki, 1973; Лавик-Гудолл, 1974). Аналогичное явление было описано не только у приматов, например, о нем сообщают Л.Д. Мич ( Mech, 1970) на основании наблюдений за волками и М. Бибен (Biben, 1982) — за южноамериканскими лисицами майконгами (Cerdocyin thous), Г. Куммер, выделивший этот феномен и посвятивший ему ряд работ (Kummer, 1973; Kummer et al., 1970, 1974; Bachmann, Kummer, 1980), обозначил его как «право обладания». Г. Куммер показал, что у гамадрилов (Papio hamadryas) «право обладания» распространяется и на самку как на объект конкуренции и существенно модифицирует отношения доминирования—подчинения.

Майконг, или лисица-крабоед Cerdocyon thous
В экспериментальных условиях самцы, предварительно информированные о том, что самка связана с другим самцом, не пытаются захватить ее, даже если доминируют над ее «обладателем». В первых работах Г. Куммера этот феномен расценивался лишь как результат права самца собственника, также как в случае права на добычу. Однако в его последней работе (совместно с К. Бахманном) было показано, что на поведение самца конкурента влияет не только его отношение к самке или отношение к ней «владельца» но и отношение самки к ним обоим. Следовательно, в этом случае «право обладания» усложняется влияниями связи на связь. Свои результаты авторы объяснили с точки зрения оценки асимметрии в стоимости борьбы и ожидаемой награды (см. раздел 4).
Если самец знает, что самка предпочитает не его, а своего «владельца», то захват ее при большом риске не удержать в дальнейшем, оказывается чрезвычайно высокой ценой, учитывая «стоимость» борьбы. Если же соперник информирован, что самка предпочитает его, то он атакует «владельца» и помогает ей освободиться от него. Такое объяснение вполне логически обосновано, хотя в сущности терминологический аппарат Дж. Мэйнарда Смита здесь не столь уж полезен, так как благодаря введению понятий «цены» и «затрат» мы в данном случае не приближаемся сколько-нибудь ощутимо к реальной оценке ни того, ни другого. Более того, злоупотребление этой терминологией способствует появлению сверхупрощенных «математических» объяснений сложных биологических феноменов. Например, в одном из исследований поведения павианов анубисов (Packer, 1979) феномен, сходный с вышеописанным, объясняется тем, что если для спаривающегося в данный момент самца вероятность отцовства будет равна 1/n (где n — число самцов, которые спаривались с самкой до него), то для любого последующего эта вероятность будет уже 1/n+1, и следовательно возникает асимметрия в «цене»- для второго самца самка уже представляет меньшую ценность.
«Право обладания» в значительной мере объясняет стабилизацию отношений между самцами обладателями гаремов и «холостяками». Это иллюстрируется и экспериментом, проведенным японскими приматологами в стаде диких гелад (Theropithecus gelada): возвращенный в стадо после двухнедельного отсутствия, гаремный самец не смог вытеснить «холостяка», быстро занявшего его место и установившего за период его отсутствия связь с самками, хотя до этого бывший «владелец» успешно и многократно отстаивал свой гарем (Mori, 1979).
5.4. «Престиж»
В группе с установившимися связями одной из важнейших определяющих характеристик доминирования является «престиж», Несмотря на свою очевидную роль, этот феномен в сущности совершенно не исследован, хотя ссылки на него можно найти во многих работах. Например, Ф. де Ваал описывает случай, когда в группе яванских макаков старый самец — вожак однажды был сильно ранен своим сыном, субдоминантам своей группы. Вполне возможно, что эта рана была нанесена при защите, а не при нападении. Тем не менее старый вожак в этот день несколько раз демонстрировал свое подчинение молодому самцу и оба были очень «осторожны» в своих взаимоотношениях. Молодой самец по размерам почти вдвое превосходил отца и обладал необходимыми физическими данными для господства в группе. Несмотря на это, никакой реверсии рангов не последовало — молодой самец даже не пытался ее произвести. Это никак нельзя объяснить тем, что он не был агрессивен. Напротив, в эти напряженные дни оба самца были сильно озлоблены, но агрессию свою они направили не друг на друга, а на остальных членов группы. Молодой самец присоединялся к отцу и они вдвоем терроризировали группу. Ф. де Ваал объясняет причины этой переадресованной агрессии тем, что старый самец боялся своего сына, а сына удерживал «престиж» отца (Waal, 1975).

Гамадрилы Papio hamadryas: самец, самка, детёныш
Такие случаи отнюдь не являются редкостью. C.Southwick, и M.R.Siddiqi (1967) описали как смертельно израненный собаками вожак в стае макаков резусов легко привел в подчинение другого самца из стаи
«… старый самец с трудом поднял голову и лишь слегка пригрозил, что тут же заставило периферийного самца удалиться».
До тех пор пока доминирующий самец не умер, стая передвигалась приспосабливаясь к его возможностям.
В большом стаде японских макаков в Орегонском региональном центре по исследованию приматов самец высшего ранга по кличке Эроухед сохраняет свое положение с тех пор, как стадо завезено в Орегон в 1964 г. У Эроухеда нет клыков, одного глаза и он один из самых маленьких самцов в стаде. Тем не менее никогда не видели посягательств на его авторитет как альфа-самца. Более того, когда Эроухед атакует других самцов, они никогда не отвечают ему тем же, а просто пытаются вырваться на свободу, По впечатлению исследователей, отношение членов стада к Эроухеду и другим самцам старшего возраста определяется «уважением» к ним (Eaton, 1976; Rasmussen, 1981).
Об аналогичных наблюдениях в группе свинохвостых макаков (M.nemestrina) сообщал И. Бернстайн (1969): старый самец, несмотря на утрату глаза и трех клыков, оставался доминантом, хотя не был ни самым крупным, ни самым агрессивным самцом в группе. Примеры доминирования старых животных над физически более мощными молодыми можно найти в работах многих исследователей (например, Yamada, 1966; Kummer, 1968; Furuja, 1969).

Свинохвостый макак или лапундер Macaca nemestrina
5.5. Отношения доминирования-подчинения и структура стада японских макаков
В одном из предыдущих разделов мы показали, что направленность агрессивных взаимодействий оцениваемая по комплексу взаимосвязанных параметров, является реальной и стабильной характеристикой групповых взаимодействий. Эта характеристика безусловно очень важна, хотя и не может рассматриваться как основная. Так как связи этой характеристики с другими не универсальны, мы проиллюстрируем ее реальное место в оценке структуры группы конкретным примером, дав описание организации стада японских макаков — животных, у которых эта характеристика проявляется в наиболее яркой форме и социальное поведение которых исследовалось очень интенсивно.
Основу типичного стада японских макаков составляют несколько кланов родственников по материнской линии, объединяемых группой «центральных» самцов (Koyama, 1967, 1970). Каждый клан состоит из взрослых самок — близких родственниц и их потомства — молодых самцов и самок. Самцы остаются со своими матерями и материнским кланом обычно до 3-4 лет, после чего постепенно покидают стадо. Таким образом, в стаде всегда находится много низкоранжированных самцов, которые либо собираются его покинуть, либо присоединились к нему и держатся на периферии стада (Sugiyama, 1976). Благодаря тому, что значительная часть самцов активно вытесняется из центра на периферию, стадо сохраняет довольно стабильную пространственную структуру. Такая «концентрическая» упорядоченность стада — результат сложных социальных взаимодействий и пространственная конфигурация каждого стада во многом зависит от сети индивидуальных взаимоотношений «индивидуальных характеров» центральных самцов и самок. «Периферийные» самцы постоянно делают попытки проникнуть в центр стада, но при этом наталкиваются на агрессивные реакции самок, которые тут же поддерживаются атаками кого-нибудь из центральных самцов. Если центральные самцы агрессивны, а самки избегают контактов с периферийными самцами, то стадо сохраняет строгую форму.
Если самцы агрессивны, но самки толерантны к периферийным самцам, то последние могут проникать в центр стада, хотя стараются держаться подальше от центральных самцов и стадо уже теряет свою определенную форму. Если же центральные самцы не агрессивны, то периферийные самцы получают еще большую свободу действий, хотя и избегают только затевать конфликты неподалеку от центральных самцов (Yamada, 1971).
Повышение ранга периферийного самца — процесс малоизученный. Однако ясно, что он определяется многими факторами, в том числе и рангом материнского клана (Koyama, 1967), Из общих соображений можно ожидать, что агрессивность, размеры, темперамент, гормональное состояние и индивидуальный опыт имеют большое значение. Однако эти факторы не оказывают прямого влияния на ранг. Эксперименты и наблюдения в Орегонском приматологическом центре показали, что хотя демонстрация клыков играет важную роль в самозащите низкоранговых самцов во время стычек, для достижения и сохранения высокого ранга наличие клыков не имеет значения (Alexander, Hughes, 1971; Eaton, 1976). Так, Г. Итон (1976) пишет:
«Парадоксально, но доминирующие самцы занимают вершину иерархии не благодаря способности драться или физическим характеристикам, таким как размеры тела. Эроухед (альфа-самец) — один из самых маленьких взрослых самцов, у него нет клыков и он потерял один глаз… Сублидерам — Бруно и Перфекту также недостает клыков, а Грейтер Тан (самец 2 ранга) достиг своего положения до того, как его клыки полностью прорезались. Если размеры и клыки не играют роли в достижении и поддержании высокого доминантного ранга, то на чем же основано «право клевка» у макаков?»
Иерархия доминантных рангов прослеживается и внутри клана самок (Kawai, 1965; Kawamura, 1965). Хотя эти отношения между самками менее ярко выражены, чем среди самцов, они очень стабильны. Кроме того, ранг самки определяет и ранг ее детеныша, особенно дочери.
Между центральными самцами, составляющими ядро стада, прямые взаимодействия могут отсутствовать, и интеграция стада в значительной мере происходит благодаря тому, что каждый центральный самец взаимодействует с несколькими кланами, а члены каждого клана поддерживают связи с несколькими самцами. Таким образом, косвенные связи через центральных самцов объединяют разные кланы, а кланы, в свою очередь, объединяют между собой центральных самцов. Связи между центральными самцами и кланами могут ежегодно изменяться: один и тот же центральный самец может в разные годы поддерживать отношения с разными кланами (Grewal, 1980).
В основе этих связей лежат «личные дружеские привязанности» самца к избранным самкам. Привязанности формируются по-разному, они могут вырастать из половой связи или возникать у самки под влиянием близости самца с ее родственницами (особенно матерью или дочерьми). Влияние центральных самцов на поведение кланов в значительной мере определяется тем, насколько велика его «дружеская компания» в каждом клане, что в конечном итоге определяет его влиятельность в стаде и социальную силу как вожака при передвижении стада.
В компании, формирующейся вокруг самца, между самками разных кланов возникают дополнительные связи, так как эти самки больше времени проводят рядом и чаще обыскивают друг друга. Таким образом, все стадо оказывается объединено «социальными узами». Градиент силы этих уз примерно таков: самая сильная связь соединяет матерей с дочерьми, затем следуют связи между родичами (в первую очередь между сибсами), затем связи самца с группирующимися вокруг него самками.
Мы не случайно назвали компании самца с самками дружескими, так как «личные привязанности» самца и группирующихся вокруг него самок формируются на «дружеской», а не на сексуальной основе. Хотя они могут возникать и из половой связи, после формирования «дружбы» половые отношения полностью прекращаются (Takohata, 1982). Таким образом, подгруппа самца не является репродуктивной единицей, что принципиально отличает ее от групп с одним самцом в стадах гамандрилов (Kummer, L968) или гелад (Crook, 1966, Kawai, 1979). Фактически высокоранговые самцы, поддерживающие связи со многими самками, не всегда имеют высокий репродуктивный успех в сезон спаривания (Taka- hata, 1982). Дело в том, что сложная концентрическая структура стада (центральные самцы, кланы, периферийные самцы) в этот период прекращает функционировать.
В сезон спаривания подгруппы распадаются, самки перестают следовать за центральными самцами и пользоваться предоставляемой ими защитой. «Рецептивные» самки далеко удаляются от своего клана в поиске избираемого самца. Это ослабление социальных сил, действующих вне сезона размножения, разрушает стену между центральной частью стада и его периферией. Даже самцы низких рангов могут в этот период производить специфические демонстрации и преследовать самок. Хотя зависимость между рангом самца и его поведением в этот период существует, она очень сложна и корреляции между разными показателями могут отсутствовать ( Enomoto, 1981).
Мы привели стадо японских макаков в качестве примера сложного сплетения разнообразных социальных связей не потому, что у других млекопитающих хищных, копытных или грызунов) сообщества примитивнее. Вероятно, на сегодняшний день примитивнее наше впечатление об этих сообществах. Для выявления описанных социальных связей у макаков понадобились непрерывные и интенсивные тридцатилетние наблюдения японских приматологов. Это уникальный случай столь длительного изучения одних и тех же естественных групп. Кроме того, приматы вообще являются одним из главных объектов в исследованиях социального поведения, поэтому при изучении японских макаков был доступен большой объем сравнительного материала по близким видам и на вооружении исследователей был хороший набор разнообразных методик.
- Система доминантных рангов и «положительный» аспект социальных связей
Конечно, деление связей на «положительные» и «отрицательные» операция сугубо условная. В каждом взаимодействии можно обнаружить источник и положительной и отрицательной стимуляции, влияющей и «на себя» и на партнера. В отношении доминирования проблема возникает потому, что если оно основано на агонистических действиях, то доминантный порядок может лишь ограничить действие рассеивающих группу отрицательных факторов (например, агрессивные контакты), но сам по себе не может обеспечить никаких положительных связей.
Между тем, есть определенная связь между доминированием и поведением «связывающим» группу. Проявляется она и в феномене более активной роли в поддержании отношений доминирования-подчинения подчиненного животного, чем доминирующего (см. раздел 3), и в ряде особенностей поведения доминанта, некоторые из которых сейчас мы рассмотрим.
6.1. «Структура внимания»
В ряде случаев найдена связь между доминированием и так называемой структурой внимания. Описание этой характеристики социальных взаимоотношений в группе было разработано в большой серии работ (Chance, 1967; Virgo, Waterhouse, 1969; Chance, Jolly, 1970; Chance,Larsen, 1976; Keyerne et al, 1978). Основным показателем при этом подходе служит «доля внимания», которую каждое животное получает от партнеров. Внимание может определяться различными путями — от длительности фиксации взглядов на данном животном до продолжительности и частоты обысканий (этот показатель мы рассмотрим отдельно).
М. Чейнц и К. Джолли (1970) показали, что у макаков, гелад и павианов внимание членов группы «центростремительно» направлено на главного самца, и предложили использовать эту характеристику в качестве параметра доминирования[5]. Если при ссаживании незнакомых животных агрессивные взаимодействия постепенно замещаются отношениями доминирования-подчинения, то параллельно идет и распределение направленности внимания. Причинные факторы, лежащие в основе этого процесса, не ясны, но было показано, что когда «структура внимания» хорошо выражена, она может быть связана с различными характеристиками взаимоотношений между членами группы, ролью доминирующего самца и с экологическими факторами (см. например, Emory, 1975, 1976; Lockwood, 1979).

Чакма или медвежий павиан, Papio ursinus
Структура внимания тесно связана еще с двумя феноменами — управлением движением группы (лидерством) и с «обыскиванием» партнеров.
6.2. Управление движением группы
Часто (хотя далеко не всегда) животное, доминирующее в агонистических взаимодействиях, также управляет и передвижением группы, В подавляющем большинстве случаев физическое принуждение здесь не причем, члены стада сами корректируют направление и скорость своего движения, ориентируя внимание на лидера или лидирующую группу. Например, у горилл группу ведет доминирующий самец. Остальные могут недалеко отходить в ту или другую сторону, но ждут дальнейшего передвижения самца. Перед началом общего движения группы самец принимает специфическую позу, после чего остальные гориллы следуют за ним (Schaller, 1963). В стае волков вожак в значительной степени управляет передвижением стаи и особенно ее действиями во время охоты (Mech, 1970).
В наиболее яркой форме совмещение черт лидера и доминанта обнаруживается у гамадрилов. В общей структуре стада гамадрилов можно выделить три уровня: стадо состоит из стай, а каждая стая включает несколько «односамцовых» групп, состоящих из взрослого самца с его гаремом, детенышей, и иногда — самца-подростка. Кроме того, в стаю входят несколько одиночных самцов. Обладатель гарема доминирует над самками и физически препятствует им далеко удаляться от. группы. В стаях часты союзы между двумя самцами, у одного из которых (или у каждого) есть свой гарем. Передвижение обычно инициируется младшим самцом, но сам он отходит недалеко, ожидая пока подтянутся к нему остальные члены группы. Однако, хотя старший самец редко открыто берет на себя роль лидера, движение самок в основном контролируется его действиями. Когда все стадо покидает место ночевки в скалах, различные лидирующие самцы пробуют выбирать разные направления движения, увлекая за собой свои гаремы. Стадо подобно амёбе ‘втягивает и вытягивает «псевдоподии» до тех пор, пока лидирующая группа окончательно не выберет маршрут (Kummer, 1970).
Ранжированный порядок следования группы гамадрилов (Papio hamadryas) (по Иванову, 2007). Идеализированная схема
Лидерство при движении обусловлено, не только тем, что доминирующие животные обладают достаточным жизненным опытом, чтобы избрать лучший путь, и не тем, что они физически способны заставить остальных подчиняться. Тот факт, что у других видов животных лидерство часто не связано с доминированием, говорит, что последний аспект (принуждение) не существен. Сравнительная оценка животными чужого индивидуального опыта также вряд ли исчерпывает проблему. Пока можно лишь сделать вывод, что возможности влиять без физического воздействия на движение группы различны у ее членов и часто в той или иной степени связаны с доминантным статусом.
6.3. Взаимные чистки и «обыскивания»
Взаимодействия этого типа, основанные на тактильной положительной стимуляции партнеров, широко распространены в разных отрядах млекопитающих. У многих видов обнаружено, что продолжительность и направленность этих действий отражают существование стабильных связей в группах. Подобно «фиксации взглядов» и «лидерству», взаимные чистки могут быть в определенной степени связаны с системой доминантных отношений.
Например, М. Simpson (приводится по Hinde, 1974) установил связь между обыскиваниями и отношениями доминирования-подчинения у шимпанзе в Национальном парке Гомбе (Танзания). В таблицах 2 и 3 дана идеализированная схема взаимосвязей между доминантным рангом, частотой и продолжительностью обыскиваний. Высокоранговые самцы обычно обыскивают чаще и у каждого самца выявляется тенденция обращать больше внимания на высокоранговых партнеров, чем на остальных. Кроме того, из таблиц следует, что каждая особь чистит каждого из партнеров примерно с такой же частотой, с какой партнер чистит ее. Анализ начала взаимодействий при обысканиях показал, что если особь очень низкого ранга взаимодействует с особью очень высокого, то инициатива обычно принадлежит низкоранговой особи,
У низкоранговых особей наблюдали тенденцию к более кратковременным чисткам партнера, чем у высокоранговых* и каждая особь чистит своего партнера тем дольше, чем выше его ранг (табл. 2). Кроме того, видно, что каждый самец обыскивает партнера высокого ранга дольше, чем этот партнер обыскивает его самого. Эти данные указывают, что обсуждаемые характеристики являются не простым следствием поведения во время самого акта обыскивания, а отражают более глубокие и длительно сохраняющиеся аспекты взаимоотношений.
Таблица 2
Связи между доминантным рангом и продолжительностью обыскиваний у шимпанзе (приводится по R.Hinde, 1974)

Tаблица 3
Связи между доминантным рангом и частотой обыскиваний у шимпанзе (приводится по R.Hinde, 1974)
 Р. Зейфэрт (1976, 1977) нашел, что у самок павианов существует связь между доминантным статусом и выбором партнера при обыскивании. Любая самка, в группе может «предложить» (приняв соответствующую позу) обыскать ее любой другой самке. В ответ на этот партнер может либо начать обыскивание, либо просто игнорировать «предложение». В последнем случае никаких особых реакций у просящей самки не бывает, она просто отходит в сторону. Таким образом, самка, которой адресовано «предложение», получает возможность решить, обыскивать ли ей партнера или нет. На основе ее действий наблюдатель может ранжировать этих партнеров по привлекательности каждого из них для другого. Результаты показали, что порядок предпочтения соответствовал порядку доминантных рангов в группе. С другой стороны, было показано, что большая часть обыскиваний направляется на животное смежного, но более высокого ранга, кроме того, если одна самка доминирует над другой, то первая может оттеснить вторую, когда та обыскивает какого-либо третьего партнера.
Р. Зейфэрт (1976, 1977) нашел, что у самок павианов существует связь между доминантным статусом и выбором партнера при обыскивании. Любая самка, в группе может «предложить» (приняв соответствующую позу) обыскать ее любой другой самке. В ответ на этот партнер может либо начать обыскивание, либо просто игнорировать «предложение». В последнем случае никаких особых реакций у просящей самки не бывает, она просто отходит в сторону. Таким образом, самка, которой адресовано «предложение», получает возможность решить, обыскивать ли ей партнера или нет. На основе ее действий наблюдатель может ранжировать этих партнеров по привлекательности каждого из них для другого. Результаты показали, что порядок предпочтения соответствовал порядку доминантных рангов в группе. С другой стороны, было показано, что большая часть обыскиваний направляется на животное смежного, но более высокого ранга, кроме того, если одна самка доминирует над другой, то первая может оттеснить вторую, когда та обыскивает какого-либо третьего партнера.
Р. Зейфэрт связал эти наблюдения, постулировав, что все самки в группе предпочитают обыскивать самку № 1, но самка № 2 оттесняет от нее остальных. Аналогично этому самка № 3 обыскивает самку N° 2, не подпуская нижестоящих и т.д. Иными словами, мы наблюдаем результирующую двух сил: привлекательности партнера, которая пропорциональна его рангу, и с другой стороны, конкурентной борьбы за контакт с ним. Эти данные нашли, подтверждение и в ряде последующих работ (см., например, Fairbanks, 1980; Seyfarth, 1980).
Связь, существующая между такими феноменами как «лидерство», «ориентация внимания», «связи при обыскиваниях» и доминантным статусом, говорит о том, что последний не может обусловливаться только отрицательным воздействием на партнеров (подавлением их активности), но связан по крайней мере с некоторыми формами положительной стимуляции, Этот аспект никак не отражен в сегодняшней концепции доминирования, что, на наш взгляд, указывает на ее неадекватность реальным событиям.
- Доминирование и репродуктивный успех
Поскольку доминантные отношения рассматриваются прежде всего как результат конкурентной борьбы за репродуктивную выживаемость, важной проблемой является исследование связи между отношениями доминирования-подчинения и репродуктивным успехом, У млекопитающих эта связь чаще всего обсуждается в отношении самцов. При этом вывод о ее существовании обычно базируется на двух тезисах:
(1) доминирующие самцы обладают приоритетом в доступе к самкам, спариваясь чаще и в наиболее благоприятные периоды;
(2) более частые спаривания, приуроченные к соответствующим периодам половых циклов самок, обеспечивают самцам избирательный репродуктивный успех.
Чаще всего иллюстрируется первый тезис. У многих видов млекопитающих мы обнаруживаем, что самцы различными способами ограничивают доступ соперников к самкам: формируя гаремы — у ластоногих (Мараков, 1974, Bartholomew, 1970), копытных (Соломатин, 1973; Баскин, 1976). хищных (Bertram, 1976) и приматов (Kummer, 1968 ; Crook 1966) или занимая соответствующее положение в группах со многими самцами ( Hausfater, 1975; Packer, i979), или защищая от конкурентов территорию, посещаемую самками (например, Walther, 1964; 1972, 1978; Estes, 1967,1969, 1974; Leuthold, 1977).
Однако монополизация самок не бывает абсолютной, и вывод о том, что приоритет в доступе к самкам доминирующих самцов всегда обеспечивает их повышенную репродуктивную выживаемость, неоднократно подвергался сомнению (например, Rowell, 1974; Hausfater, 1975; Kolata, 1976; Ralls, 1977). Прямая проверка этого предположения может быть основана лишь на точном установлении «отцовства», которое затруднительно провести даже в экспериментальных условиях. Пока нам известны лишь две работы, в которых доминантный ранг был связан с отцовством, установленным по «генетическим маркерам» в группе макаков-резусов. Тем не менее, работы в этом направлении уже начали проводиться, хотя пока дали лишь частные результаты (Duvall et al., 1976; Smith,1983).
Д. Дьюсбери (Dewsbury, 1982), тщательно рассмотревший современные данные по всей проблеме, пришел к выводу, что в целом значение доминантного статуса для репродуктивного успеха преувеличено, прямые свидетельства этой связи получить очень трудно и бытующие представления опираются на недостаточно репрезентативные и косвенные показатели. Таким образом, вывод о дифференциальном репродуктивном успехе животных разных социальных статусов редко бывает хорошо обоснованным.
Во-первых, наиболее употребимый показатель — связь между доминированием и частотой спаривания — должен определяться для каждых конкретных условий. При этом недостаточное соблюдение общих требований к сбору и обработке это- логического материала часто приводит к искажению реальной картины[6]. Во-вторых, как правило отсутствуют данные, позволяющие (1) адекватно интерпретировать репродуктивный успех в зависимости от очередности спариваний; (2) правильно оценивать влияние обшей и дифференцированной способности к оплодотворению; (3) правильно оценивать репродуктивный успех не по кратковременному преимуществу, полученному в данный момент, а по результатам, достигаемым на протяжении всей жизни особи. Рассмотрим каждый из этих моментов.
7.1. Прямая, позитивная связь между доминированием и частотой спаривания
В группах с несколькими самцами-производителями такая связь часто обнаруживается, но следует подчеркнуть, что бывает она не всегда и не является универсальной. В некоторых исследованиях корреляции между рангом и частотой спаривания не обнаруживается (см. например, Baldwin, 1968; Horwich, 1972).
Таблица 4
Связь между рангом и частотой копуляций (сводные данные, по Dewsbary, 1982)
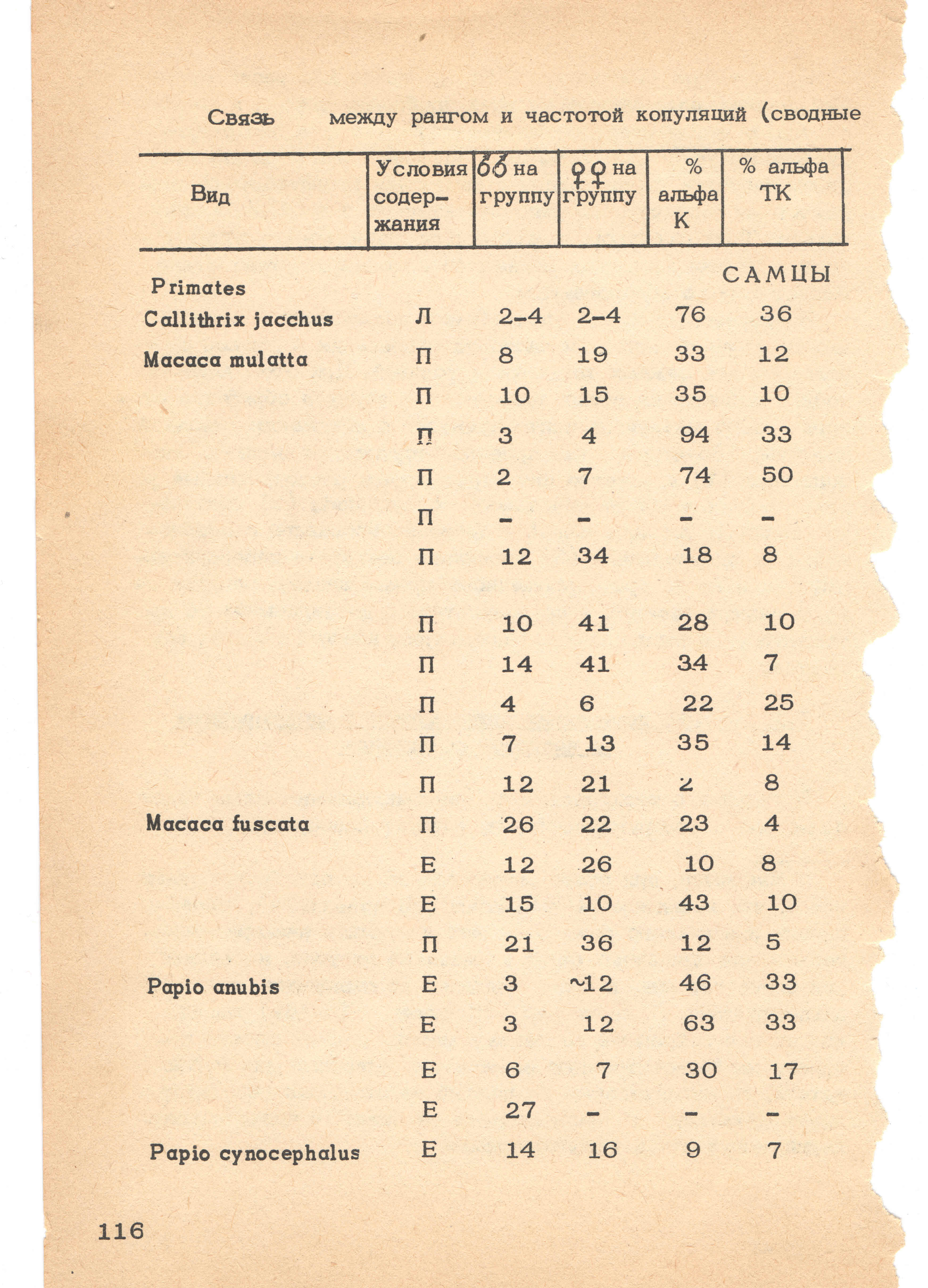
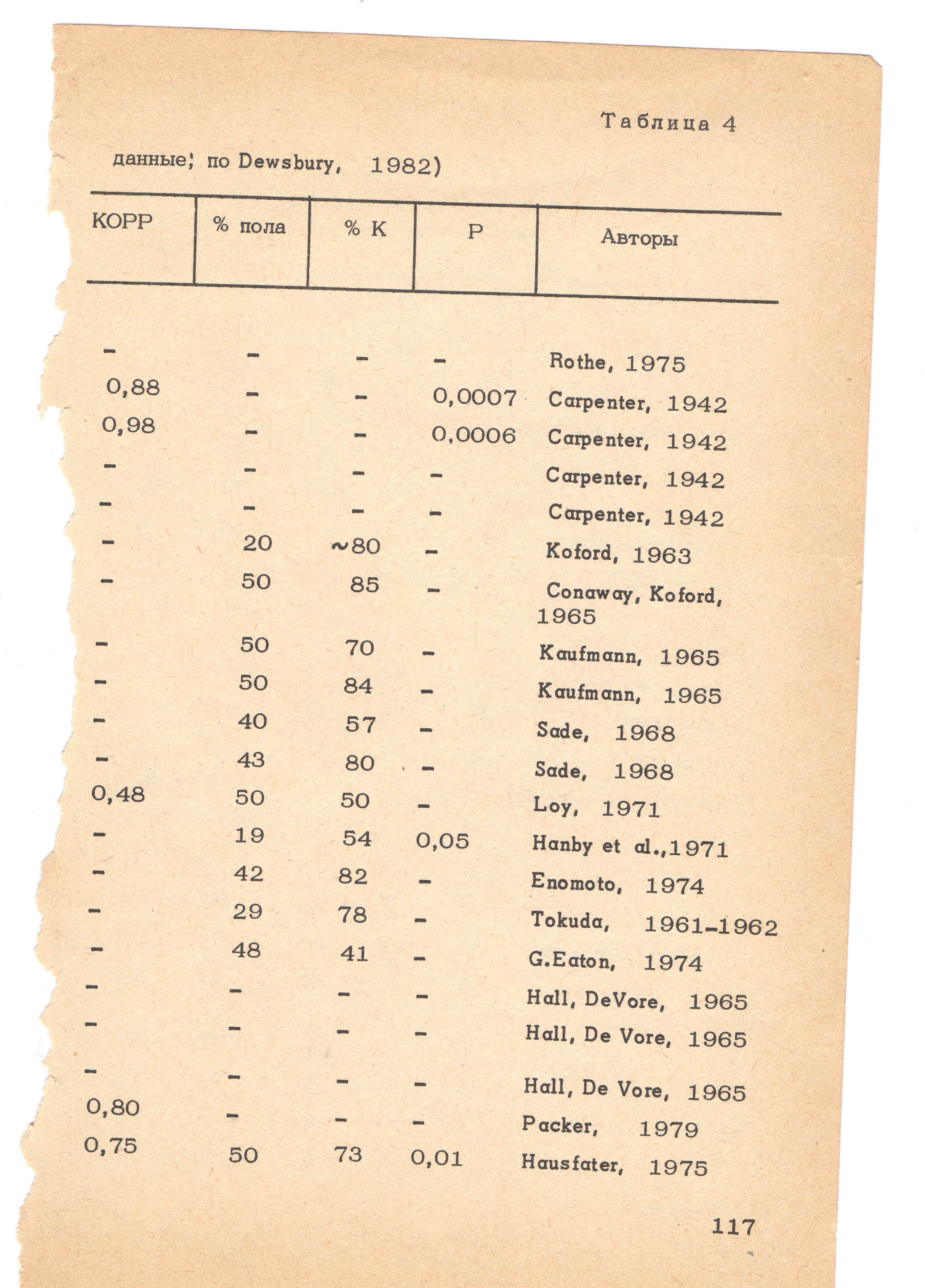
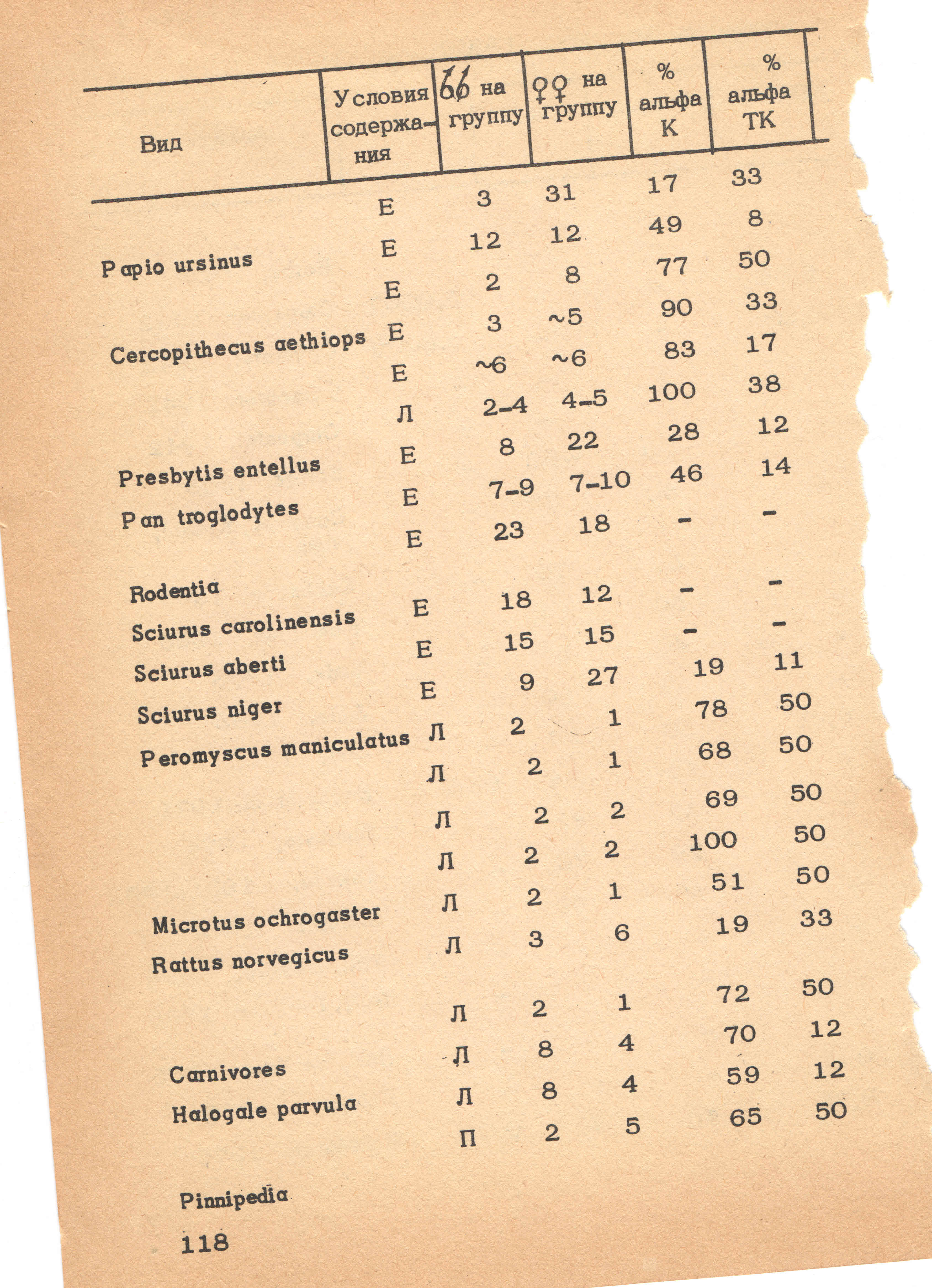

Даже у видов с наиболее отчетливо выраженной системой доминантных рангов, (например, у павианов и макаков, эта связь оказывается зависящей от многих условий. Так, например, Г. Итон (1974) сообщил о результатах многолетнего исследования связи между социальным статусом, агрессивностью и репродуктивным поведением в большом стаде японских макаков, живущих в загоне. Было показано, что половая активность самцов не коррелирует с их социальным статусом и агрессивным поведением. Кроме того, автор нашел, что вмешательство доминирующих самцов мало влияет на частоту спариваний подчиненных особей. В работе К. Пэкера (1979) эта корреляция обнаружена у павианов анубисов, но в работах Б. Смуте (приводится по Smuts, 1981) и С. Страм (1980) она не найдена. Методы регистрации и описания поведения в этих исследованиях были близкими, так что методологичекие различия в них не могут служить причиной расхождения результатов. Следовательно, можно сделать вывод, что даже у одного вида с резко выраженной формой доминантных отношений связь между доминантностью и успехом при спаривании может сильно варьировать (Smuts, 1981).
Д. Дьюсбери (1982) свел результаты значительного числа работ в таблицы, которые мы здесь приводим (табл. 4). Наиболее изменчива корреляция между доминированием и частотой спариваний у приматов и у грызунов, наиболее стабильна у копытных и у хищных. В целом же можно сделать вывод, что эта связь сложна, изменчива и требует специального анализа в каждом конкретном случае. Кроме того, нельзя переоценивать и сам факт ее существования — она является лишь косвенным показателем репродуктивного успеха.
7.2. Очередность копуляций и репродуктивный успех
У многих видов самка в период восприимчивости» может спариваться не с одним, а с несколькими самцами. Поведение самцов при спаривании часто зависит от их социального статуса. Исследователи оценивают успех при спаривании по таким показателям как очередность и частота копуляций, интервал между спариванием и периодом овуляции у самки. У ряда видов обнаружено, что доминирующие самцы спариваются первыми и чаще демонстрируют элементы полового поведения. У некоторых видов они спариваются преимущественно в те дни, когда наиболее вероятна овуляция, так что вывод об их избирательном репродуктивном успехе кажется довольно обоснованным.
Так по словам Б. ле Бефа и Р. Петерсона (Le Boeuf, Peterson, 1969) на лежбище морских слонов (Mirounga angustirostris) лишь 4% самцов «по-видимому оплодотворяют» 85% самок. Авторы отмечают, что эти данные учитывают лишь результаты первого спаривания, и вторичные спаривания с другими самцами из рассмотрения исключены. В то же время в конце периода эструса самки часто спариваются с периферийными самцами (Сох, Le Boeuf, 1977). У некоторых приматов (макаков и павианов) доминирующие самцы часто спариваются либо первыми, либо преимущественно на той стадии, когда у самки, происходит овуляция (например, Carpenter, 1942, Hall, de Vore, 1965; Hausfater, 1975; Packer, 1979). Например, у бабуинов (Papio cynocephalus) самка в эструсе спаривается с самцами на протяжении приблизительно недели. Обычно в этот период самку постоянно сопровождает какой-нибудь самец, причем в течение недели самцы могут несколько раз меняться. Г. Хаусфатер (1975) показал, что в исследуемой им популяции доминирующие самцы чаще спаривались на третий день после набухания у самки «половой кожи», то есть когда овуляция была наиболее вероятной. Аналогичные данные получил и К. Пэкер, исследовавший павианов анубисов (Packer, 1979).

Бабуин или жёлтый павиан Papio cynocephalus
В экспериментальных условиях показано, что хомячков Peromyscus maniculatus первое спаривание с эйякуляцией обычно принадлежит доминанту, так же как и большее число эйякуляций, в то время как подчиненный самец чаще эякулирует последним (Dewsbury, 1979), Картина распределения частоты копуляций у кистеухой белки (Sciurus aberti) в естественных условиях описана в работе Farentinos (1980). Самки этого вида рецептивны один раз в году. В этот день вокруг самки собираются несколько самцов. Спаривание длится от 9 до 13 часов и наиболее агрессивные самцы утверждают свой «приоритет в доступе» к самке. За этот период самка успевает спариваться с 2 — 6 разными самцами. Однако доминирующий самец спаривается не только первым, но и чаще остальных.
Во всех приведенных случаях различия в очередности копуляций доминирующих и подчиненных самцов явно существует. Однако интерпретация этих данных отнюдь не очевидна.
Когда самка в один период эструса спаривается с несколькими самцами, в силу вступает физиологическая «конкуренция» спермы самцов. Фертильность спермы в репродуктивных трактах самки сохраняется не один день. Динамика взаимодействий сперматозоидов из разных эйякулятов очень сложна, мало изучена и варьирует у разных видов.
Последовательность копуляций и их частота безусловно могут иметь значение, но чтобы оценить последствия мы должны располагать многими данными. Так, в экспериментальных условиях было показано, что у домовой мыши первая эйякуляция при спаривании одной самки с несколькими самцами дает преимущество при оплодотворении (Levine, 1967), но у золотистых хомячков (Oglesby et al., 1981) и прерийных полевок (Dewsbury, Baungardner, 1981) преимущество получают самцы, спаривавшиеся последними. У некоторых видов влияния порядка спаривания не обнаружено — например, у свиней (Sumption, Adams, 1961), серых крыс (Lanier et al., 197 9) и хомячков Peromyscus maniculatus (Dewsbury, Baumgardner, 1981). В экспериментальных конкурентных ситуациях с двумя самцами у лабораторных крыс больше потомков оставлял самец, эйякулировавший последним и большее число раз. У самца, производящего эйякуляцию первым, преимуществ в оплодотворении не было (Dewsbury, Hartung, 1980). Все сказанное справедливо и в отношении данных, полученных при исследовании бабуинов, анубисов — спаривания, приуроченные к овуляции, могут быть не единственными эффективными.

Анубис или оливковый павиан Papio anubis с добычей
7.3. Общая и дифференциальная способность к оплодотворению
Самцы с разными генотитами могут отличаться как по обшей плодовитости, так и по относительной способности оплодотворять самок в конкурентных ситуациях, когда их сперма в половых путях самки встречается со спермой других самцов. Анализ этого явления возможен сейчас лишь в отношении лабораторных животных (см. например, Lanier et al., 1979); так что значение этого феномена еще только предстоит выяснить (Dewsbury, 1982).
7.4. «Общий» репродуктивный успех особи и «альтернативные» репродуктивные стратегии
Проблема роли доминантного статуса в установлении половых связей рассматривается во многих работах. Значительно меньше внимания уделяется вопросу, насколько статус и репродуктивный успех могут варьировать на протяжении жизни одного индивидуума. Хотя эта проблема пока еще мало изучена, полученные данные позволяют утверждать, что различия между возможностями самцов в установлении половых связей не столь уж очевидны, даже при полигамии с жестким ограничением числа самцов, имеющих доступ к самкам. Проиллюстрируем это несколькими примерами.
Еще недавно структура лежбища северного морского котика (Callorhinus ursinus) рассматривалась как относительно неизменная на протяжении сезона спаривания. Считалось, что захват относительно небольшим числом секачей репродуктивных участков обеспечивает им оплодотворение подавляющего большинства самок. Однако последние работы показали, что структура лежбища претерпевает непрерывные изменения со сменой гаремных секачей. Колоссальный энергетический расход самца, участвующего в драках и покрывающего многих самок, приводит к быстрой замене его другим секачем со свежими силами. Исследования, проведенные на одном из лежбищ о. Медного (Урилье лежбище), показали, что за основной период спаривания на гаремной территории лежбища совершается более, чем трехкратная смена секачей. Способы, время и продолжительность захвата территории и самок, место преимущественной активности самца, определяются его индивидуальными особенностями и возрастными «нормами».
Секачи принимают участие в размножении в возрасте от 7 до 14 лет и обладают разными физическими возможностями и опытом. Например, в 1980 и 1981 гг. на указанном лежбище о. Медного в размножении принимали участие 14,3% 7-летних секачей, 47,9% — 8-летних, 63,5% — 9-летних, 66,7% — 10-летних, 71,4% — 11-летних. Секачи 12-14 лет обзаводились гаремами поголовно. Время выхода секачей на лежбище на протяжении репродуктивного периода и продолжительность владения территорией также зависят от возраста, например, 7-летние удерживают свою территорию в среднем 5,7 дней; 8-летние — 12,9; 9-летние — 19,9; 10-летние — 16,4, а секачи 10 и более лет — 13,5 дней (Лыскин, Владимиров, 1982, а,б). Столь же динамичен режим использования лежбища сивучем (Eumetopias jubatus). На наиболее предпочитаемых участках секачи удерживаются в среднем около 9 дней, на менее привлекательных для соперников — 13. В местах с высокой конкуренцией секачи бывают вынуждены до 5 раз менять свой участок (Лисицына, 1976, 1982).

Кистеухая белка Sciurus aberti
Согласно типичной схеме социальной организации копытных, особенно африканских Bovidae (см„ например, Estes, 1974; Jarman, 1974; Owen—Smith, 1977), стада (гаремы) самок с молодняком широко перемещаются по обширным участкам. Часть взрослых быков устанавливает индивидуальные территории на участке обитания самок и владелец территории сопровождает самок, пока те пасутся в охраняемой им зоне. Таким образом, чем больше и выгоднее расположена территория самца тем больше времени он проводит с самками. Владельцы территорий происходят из стад «холостяков», объединяющих всех взрослых и молодых самцов. Четкая иерархия в этих стадах устанавливается в результате схваток с партнерами. Самцы высшего ранга становятся владельцами территорий.
Эта схема без учета индивидуального жизненного пути самца наводит на мысль о том, что явное деление самцов на»имущих» и «неимущих» приводит к жесткой избирательности при размножении. Однако такой вывод не обоснован. Например, Г. Дюбост и Ф. Фир (Dubost, Feer, 1981 ) показали, что у гарны (Antilope cervicapra) даже в условиях ограниченной свободы, жизненный путь самца совсем не прост. Стадо антилоп (35 взрослых самок и 13 самцов), живущее в огороженном парке, в основных чертах сохраняло свою естественную структуру. Самки с ягнятами использовали определенный участок пастбища, основную часть которого защищал от вторжения других быков альфа-самец. Кроме того, на периферии участка самок защищали свою территорию бета- и гамма-самцы. Эти три самца изолировали остальное стадо «холостяков» от стада самок. Поскольку альфа-самец являлся доминантом в линейной иерархической системе отношений, можно было ожидать, что повышение ранга происходит последовательно. Однако оказалось, что ни бета- ни гамма-быки никогда не становятся альфа. В свою очередь, смещенный альфа-бык никогда не занимает положения бета-особи. Во всех случаях альфа-самцами становились быки «средних» рангов (нижние ступени иерархической лестницы занимают молодые животные). Таким образом, были выделены две основные социальные стратегии у самцов: одна, направленная на достижение альфа-статуса, а другая — бета. Смещение альфа-самца сопровождалось полной перестройкой иерархической системы. За период исследования все взрослые быки (за исключением больных животных) по крайней мере однажды занимали альфа- или бета-положение.

Антилопа гарна Antilope cervicapra
В естественных условиях становление репродуктивной стратегии самца может происходить еще сложнее. Как показали исследования, М. Джармэн (Jarman, 1980) у антилопы импала (Aepyceros melampus) в национальном парке Серенгети успех территориального самца зависит от многих факторов: размера территории, привлекательности пастбища для самок, времени года, когда охраняется территория и длительности периода владения территорией. Интенсивность конкуренции за различные территории в разное время года варьирует и, следовательно, различна и энергия, затрачиваемая на их защиту. Быки импала почти всегда пытаются восстановить территорию в том же самом месте, где они имели ее до этого. Таким образом, первый выбор территории в значительной мере определяет выбор дальнейшей репродуктивной стратегии быка, а возможно и eго долголетие. Хотя в каждый данный момент времени лишь около трети взрослых самцов удерживает территории, относительно быстрая смена хозяев делает невероятым предположение об исключении из размножения сколько-нибудь существенной части популяции. Аналогичные данные можно найти и в работах по другим копытным (например, loody, Arnold, 1975).
Г. Хаусфатер (Hausfater, 1975),описавший у бабуинов связь между доминантным статусом самца и частотой спаривания в оптимальный для оплодотворения период полового цикла самки, в заключении работы пишет:
«… данные по общему репродуктивному успеху за всю прожитую индивидуумом жизнь настоящее время отсутствуют даже для отдельных видов приматов. Предположим, например, что каждый взрослый самец бабуин на протяжении своей жизни остается на каждой ступеньке иерархической лестницы столько же времени, сколько и все остальные. В этом случае, если рассматривать достаточно большие отрезки времени, все самцы будут обладать равным суммарным репродуктивным успехом. Даже если (что более вероятно) самцы отличаются по тому, сколько времени и в какой последовательности они занимают различные ранги, суммарный репродуктивный успех у них может остаться равным.
Например, в нашем исследовании самцы второго ранга обладали большим успехом, чем самцы пятого ранга, хотя последние могли дольше сохранять свой статус, чем самцы второго ранга. Таким образом, чтобы достичь данного уровня репродуктивного успеха, самец может либо долго занимать пятый ранг, при невысоком темпе размножения, либо короткое время занимать второй и более активно участвовать в размножении. Конечные же результаты могут быть одинаковыми… Очевидно, что и специфическая для данного статуса смертность тоже играет роль в определении общего репродуктивного успеха при этих различных стратегиях» (Hausfater, 1975, с. 140-141).
Конечно, мало вероятно, что суммарные репродуктивные успехи всех самцов в популяции равны, однако ясно, для того чтобы говорить об избирательности репродуктивного успеха, требуется учитывать множество факторов.

Антилопа импала Aepyceros melampus
Наконец, при сравнении результативности репродуктивных усилий самцов следует одновременно учитывать и поведение самок. Даже у тех видов млекопитающих, у которых доминантные отношения среди самцов играют большую роль в регулировании половых связей, выбор, производимый самкой, не менее существенен (см. например, Ewer, 1968; Wittenberger, 1980). В уже цитированной работе Р. Фарентинос (1980) по поведению кистеухих белок автор указывает, что во всех (11) наблюдавшихся случаях гона была отмечена экстремальная «неприступность» самок для альфа-самцов. Самки мешали им при половых контактах, отталкивали, убегали, демонстрировали угрозы и кричали, а иногда даже переходили к открытым атакам. Напротив, попытки спаривания некоторых подчиненных самцов у самок явно не вызывали протеста. Процент успешных копуляций от общего числа подходов был достоверно меньше у альфа-самцов, чем у подчиненных (Farentinos, 1972в).
«Особенно наглядна относительная легкость, с которой некоторые подчиненные самцы успешно спаривались с самками, на короткое время оторвавшимися от преследований альфа-самцов. Более того, при спаривании 4 самок я наблюдал, как, убегая от альфа-самца, самка сама приближалась к одному из подчиненных и подставлялась ему для копуляции. Чтобы подобные сексуальные предложения адресовались альфа-самцам, я не наблюдал ни разу» (Farentinos, 1980, с. 338).
Эти наблюдения затрагивают еще одну интересную, но малоизученную тему. В ряде работ были сделаны попытки найти связь между «манерами» поведения доминирующих и подчиненных животных и половым поведением (Lorenz, 1935; 1966; Yerkes, 1939b ). «Опорными точками» для постановки проблемы служат следующие факты:
(1) у млекопитающих самцы, как правило, доминируют над самками (Wilson, 1975);
(2) позы доминирования или самоутверждения и подчинения часто имеют определенный сексуальный контекст, при этом действия доминанта чаще включают элементы полового поведения самца, тогда как у подчиненного — самки ( Maslow, 1936);
(3) в половом поведении многих видов млекопитающих (как и других позвоночных), прослеживается связь между агрессивностью и половым поведением самца, и распространена та или иная степень «насилования» самок (Lorenz, 1935; Leyhausen, 1975);
(4) влияние со стороны подчиненных и доминирующих партнеров могут оказывать взаимное как маскулинизирующее, так и феминизирующее воздействие на поведение, что соответствующим образом отражается и на половой активности (Гольцман, 1977);
(5) получены данные, подтверждающие гипотезу Р. Йеркса (Yerkes, 1939a) о том, что усиление доминирования при половом поведении у самцов приматов способствует уменьшению зависимости спариваний от половых циклов самок (см., например, Nadler, 1977; Nadler, Miller, 1982). Мы не имеем возможности рассмотреть сейчас все эти моменты, но должны отметить, что тенденция к «сексуальному» доминированию тоже может быть одним из факторов, определяющих различия в стратегиях репродуктивного поведения разных самцов. Эта проблема пока еще не изучена и заслуживает внимания.
Заключение
В последние годы при использовании концепции доминирования возникают существенные трудности. В первую очередь это связано с тем, что неясны механизмы, лежащие в его основе. Дополнительные сложности порождает недостаточно четкое определение понятия «доминирование». В настоящее время этот термин используется двояким образом — «доминирование» в широком смысле обозначает всю совокупность социальных сил, которые обусловливают деление животных на «имущих» и неимущих»; термин «агонистическое доминирование» используется как описательное эмпирическое понятие, обозначающее устойчивую асимметрию в агонистическом поведении.
При использовании понятия «доминирование» в широком смысле возникает опасность неоднозначного толкования его разными авторами. Концепция при этом может превратиться в чисто умозрительную схему, подкрепляемую некорректно отбираемыми данными. С другой стороны, использование понятия «доминирование» в узком смысле, лишь как описательного термина, для обозначения феномена по эмпирически выделяемым критериям приводит к существенному обесцениванию этого понятия. Мы предполагаем, что решение возникающей проблемы должно состоять не в дальнейшем уточнении уже существующей концепции и не в подборе соответствующих фактов, которые позволили бы более точно определить границы применения понятия «доминирование» в данных конкретных условиях для данного вида животных. При росте наших знаний о биологии вида и специфике социальных связей эта задача будет беспредельно усложняться.
Более перспективна надежда, что со временем произойдет замена самого понятия «доминирование» более общей и адекватной концепцией. Такая концепция должна интерпретировать не только феномены, связанные с агрессивным поведением, но и все другие формы динамической асимметрии поведения. Для ее разработки, по-видимому, придется пересмотреть сам подход к анализу поведения. Он должен, в первую, очередь, опираться не на выявление конечных стадий взаимоотношений животных — «статусов» доминанта и подчиненного, а на хорошее описание кинематики процесса взаимовлияния животных друг на друга.
Примечания
[1]Стабильную асимметрию, подобную возникающей в агрессивных схватках, можно видеть в любых формах взаимодействий — половых, играх, чистках и пр. Ее отражением является и «комплементарность» поз и действий (например, садки и подставления и т.п.).
[2] Согласно теории В.Винн-Эдвардса, в популяциях животных прямая борьба за ресурсы (например, корм или половых партнеров), таящая опасность подорвать источник ресурсов, заменяется конкуренцией «по соглашению» (conventional competition) за «условную награду» (например, территорию). В процессе эволюции развиваются символические средства борьбы при условной конкуренции (например, демонстрации).
[3] Например К. Пекер (Packer, 1979) нашел факторы, облегчающие у анубисов (Papio anubis) доступ подчиненных самцов к самкам: (а) Установление самкой брачной связи до того, как это сделает один из доминирующих самцов, которые предпочитают вступать в брачную связь только на определенной стадии цикла самки. Таким образом, подчиненный заранее устанавливает «право обладания» самкой; (б) формирование коалиций, когда два или несколько самцов, объединившись, отбирают самку у «супруга», в этом случае им становится обычно инициатор коалиции, который может не доминировать надо своими помощниками; (в) специфический опыт «ухаживания» за самками; (г) постепенно ослабление отпора со стороны доминирующих самцов, которые не могут долго сохранять необходимую физическую форму (поддержание брачной связи требует большого энергетического расхода); (д) избирательность в выборе полового партнера как со стороны самцов, так и самок.
[4]Дж. Мэйнард Смит, посвятивший этой проблеме специальное исследование, приходит к выводу, что понятие «группового отбора» следует ограничить случаями, в которых единицами отбора являются группы. Для того, чтобы такой процесс мог иметь место необходимо, чтобы группы были способны «репродуцироваться» (например, расщеплением) и чтобы часть из них вымирала на эволюционных временах. Появление альтруистической черты (но не ее сохранение) требует, чтобы группы были небольшими или происходили от небольшого числа основателей Отбор родичей требует только того, чтобы родственники и жили недалеко друг от друга и в связи с этим могли оказывать влияние на шансы друг друга выжить и размножиться. В этом случае жизнь группами не обязательна (Maynard Smyth, 1976).
[5]Здесь можно еще раз подчеркнуть условность того, что мы считаем основным параметром доминирования — М. Чейнц и К. Джолли, например, постулируют, что основное — это направленность внимания.
[6]Например, при сборе материала по методу «регистрации всего, что возможно» («ad libitum«, Altmann, l974), наблюдатель значительно реже отмечает копуляции низкоранжированных или одиночных самцов макаков-резусов, поскольку они более скрытны и чаше держатся на периферии стада, чем доминирующие. Л. Дрикэмер (Drickamer, 1974а) нашел, что если в собранные по такому методу данные ввести поправку на «относительную заметность» животных для наблюдателя, то значительные на первый взгляд различия между доминирующими и подчиненными самцами в частоте копуляций могут стать недостоверными.
Итоги науки и техники. Сер. зоол. позв. М.: ВИНИТИ. – 1983. – Т. 12. – С. 71-150.
Рекомендуем прочесть
 Как Европа открыла для себя Московскую Русь, а потом закрыла
Как Европа открыла для себя Московскую Русь, а потом закрыла COVID-19 вскрывает правду
COVID-19 вскрывает правду «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Банковское дело и эволюция целей банковского регулирования
Банковское дело и эволюция целей банковского регулирования- «Люди из Черкас»: «ордынские казаки» и «служилые татары» лесостепного пограничья (конец ХIV-XV вв.)
- Социальная сегрегация в городах
- Про европейские ценности
 Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных
