

Лесная мышь Apodemus uralensis
Я писал, что биологизаторство как научная теория (или объяснительная схема) в этологии тем, что «не примечает слона»(часто закрывая глаза и отворачиваясь от соответствующих феноменов, распространённых настолько широко, что «лезут в глаза»). Или даже пары «слонов» .
Во-первых, это формообразующая роль социального влияния, и её дериват на уровне пространственно-этологической структуры — примат системности над адаптивностью. Поэтому особи, связанные социальной связью (превращающей их в компаньонов по Лоренцу), вкладываются в её поддержание больше, чем в индивидуальную приспособленность; что важнее всего, они сперва вкладываются в первое, а во второе – только потом и лишь при условии успешности первого. См. «Что не так с социобиологией?«
Во-вторых, это способность отношений, соединяющих особей в сообщество (пространственно-этологической или социальной структуры), сохранять устойчивость, самотождественность и воспроизводиться при радикальных изменениях демографии, персонального состава сообщества и мотивации особей. Это т.н. демографическая и физиологическая стохастичность; бывает и средовая; относительно всех них структура связей сохраняет устойчивость, что хорошо видно при экспериментальных попытках её «расшатать», особенно не уменьшая актвности. И чем «социальнее» вид, тем устойчивость больше, см. исследования С.А.Шиловой (1993) на разных видах грызунов и В.С.Громова (2000), в основном на песчанках.
Фармакологические изменения агрессивности и социальная структура: С.А.Шилова, 1993
Содержание
«Современная биологическая литература непрерывно пополняется оригинальными и обобщающими работами, которые демонстрируют важнейшее значение социальных форм поведения млекопитающих в жизни, становлении и динамике популяций. Не вдаваясь в детальный анализ этих интересных сведений, отметим лишь, что сейчас хорошо известна роль основных форм социального поведения мелких млекопитающих преимущественно грызунов) в поддержании оптимальной популяционной плотности и стабильности внутрипопуляционных группировок.
Среди основных поведенческих реакций зверьков, определяющих популяционный гомеостаз, многие авторы придают большое значение агонистическим формам поведения, проявлению прямой и ритуализированной агрессии.
Поскольку роль социальных взаимодействий животных в поддержании популяционного гомеостаза ни у кого не вызывает сомнений, логично предположить, что нарушение таких реакций может вызвать необратимые последствия, которые в итоге будут препятствовать осуществлению нормальных популяционных функций и приведут к необратимому падению численности.
Перечисленные факты привели нас к мысли, что направленное изменение некоторых важных поведенческих актов в популяциях грызунов может оказаться одним из способов искусственного снижения численности интересующих нас видов.
Оговоримся сразу, что мы хорошо представляем сложность получения определенных результатов в предлагаемом направлении исследований и далеки от мысли рассчитывать на немедленное внедрение в практику соответствующих рекомендаций.
В качестве средств, нарушающих поведенческие реакции животных мы выбрали некоторые фармакологические препараты, преимущественно нейролептики и антидепрессанты, изменяющие уровень агрессивности. Ранее некоторые из этих препаратов (резерпин и др.) были предложены в качестве прямого дератизационного средства, в больших дозах вызывающего быструю гибель лабораторных мышей в результате релаксации мускулатуры. Несмотря на то, что прямое уничтожение грызунов в данном случае не соответствовало нашим целям, мы использовали для дальнейшей работы ориентировочные дозировки препаратов и описание клинической картины их проявления. К тому же уже было известно, что некоторые нейролептики, в частности аминазин, используются в животноводстве и звероводстве для снижения агрессивности некоторых животных (Шитый, 1966; и др.).
Поскольку интересующее нас направление исследований в доступной нам литературе не описано, мы приводим ниже лишь собственные данные, полученные в нашей лаборатории в 1977—1983 гг. (Shilova, 1991).
В качестве объектов исследований нами использованы естественные популяции некоторых мелких млекопитающих, населяющих высокогорные и аридные зоны (Тува, Калмыкия): монгольская пищуха, монгольская и полуденные песчанки, длиннохвостый суслик. Серия экспериментов выполнена на домовой и лесной мышах в вольерах, имитирующих естественные поселения этих зверьков в жилищах.
Основной метод исследования — регистрация пространственного размещения и важнейших форм поведения у животных в естественных условиях, получивших индивидуальные метки черным урзолом Д. За время полевой работы в Туве и Калмыкии помечено 537 зверьков, проведено 1445 ч наблюдений и составлено 2220 этограмм.
Монгольская пищуха
Монгольская пищуха характеризуется ярко выраженным индивидуальным использованием территории — как самцами, так и самками. Со второй половины лета, после расселения молодняка, каждый зверек имеет свой участок, границы которого он охраняет от соседей. Соответственно, сущность адаптивного поведения монгольских пищух заключается в высоком уровне агрессивности, который способствует полной изолированности индивидуальных участков. Высокая агрессивность монгольских пищух — основной фактор, способствующий поддержанию территориальной структуры популяций в условиях дефицита кормов.
 Эксперимент на монгольских пищухах проведен нами в естественных условиях юго-западных районов Тувы, где зверьки заселяют участки высокогорной щебнистой степи. Плотность пищух достигает здесь 20— 30 шт. на 1 га.
Эксперимент на монгольских пищухах проведен нами в естественных условиях юго-западных районов Тувы, где зверьки заселяют участки высокогорной щебнистой степи. Плотность пищух достигает здесь 20— 30 шт. на 1 га.
Опытный участок площадью 1 га был расположен на южном склоне каменистого ущелья. К середине июля иерархическая структура опытной семейной группировки полностью установилась и произошло окончательное распределение зверьков по индивидуальным участкам, границы которых они энергично охраняли. Многочисленные наблюдения за пищухами, проведенные в предыдущие сезоны и на аналогичных территориях, показали, что к середине лета, после расселения молодняка второго выводка, группировки пищух обладают большей устойчивостью и сохраняются до весны следующего года.
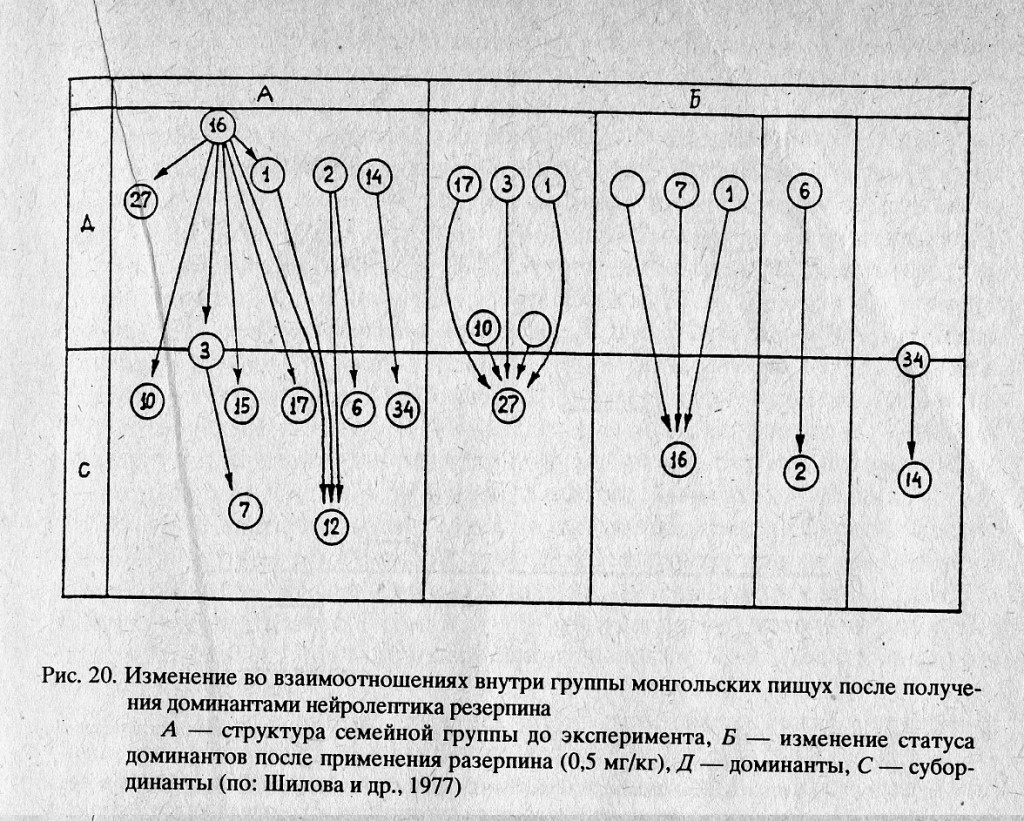
С целью снижения агрессивности пищух применен резерпин. Дозировка препарата в 0,5 мг/кг вызвала заметные изменения в групповых отношениях зверьков. До начала опыта на участке регулярно встречалось 14 пищух, из которых доминантное положение занимали 2 взрослых самца (№№ 16, 27), две взрослые самки (№№ 1, 2) и частично одна молодая самка первого выводка (№ 14). Наибольшей агрессивностью во всей группировке отличался взрослый самец № 16.
Препарат получали все зверьки, занимающие доминантное положение и четыре субдоминанта. Остальные пищухи препарат не получали, что позволило проследить взаимоотношения между контрольными и опытными зверьками.
После получения препарата внешних изменений в состоянии зверьков не наблюдалось. Они продолжали кормиться, грелись на солнце и подавали сигналы. Однако существенно изменилась их способность охранять территорию. Доминанты, получившие препарат, потеряли иерархическое превосходство и стали подвергаться нападениям своих низкоранговых соседей (рис. 20; Шилова и др., 1977). В зависимости от индивидуальных особенностей зверьков иерархическое положение пищух, получивших резерпин, изменялось по-разному. Среди пяти доминантов, населявших площадку, только одна взрослая самка продолжала активно охранять свой участок, хотя и в этом случае наблюдалась тенденция к ограничению границ ее территории молодыми зверьками. Интересно, что две пищухи сумели восстановить свое доминантное положение через 2 дня после окончания действия препарата.
Старый самец № 16, занимавший на площадке наивысший ранг, пострадал сильнее всех. После получения препарата он подвергался нападению трех своих соседей и, наконец, был вообще изгнан с участка. Молодая самка № 14, занимавшая доминантное, но не слишком прочное положение, стала подвергаться нападению соседей и до конца опыта так и не восстановила своего ранга.
В результате ослабления охраны границ индивидуальных участков доминирующими зверьками и выселения некоторых из них за пределы семейной территории на опытном участке в течение августа происходило достаточно хаотичное перемещение пищух из одной норы в другую, что никогда не наблюдается в этот период в нормальных условиях.
Поскольку строгое распределение индивидуальных участков зверьков во второй половине лета является, как мы говорили, основой их адаптивного поведения и приспособлением к переживанию суровых зимних условий, можно предположить, что описанные выше нарушения могли значительно ухудшить условия существования популяции. Действительно, нарушение принципов нормального функционирования популяционной группы ухудшило ее существование и сказалось на численности: весной 1977 г. на участке сохранилось всего два зверька, хотя предыдущей осенью, до начала эксперимента, территорию населяло 30 животных.
Аналогичные данные мы получили при воздействии на поселение монгольских пищух нейролептиком галоперидолом. Препарат в дозировке от 0,5 до 1,5 мг/кг при многократном введении вызывал также значительное ослабление тенденции к охране границ индивидуальных участков, что повлекло за собой существенное изменение территориального распределения зверьков (рис. 21). Произошло смещение границ индивидуальных участков, проникновение на площадку большого количества ’’пришельцев”.
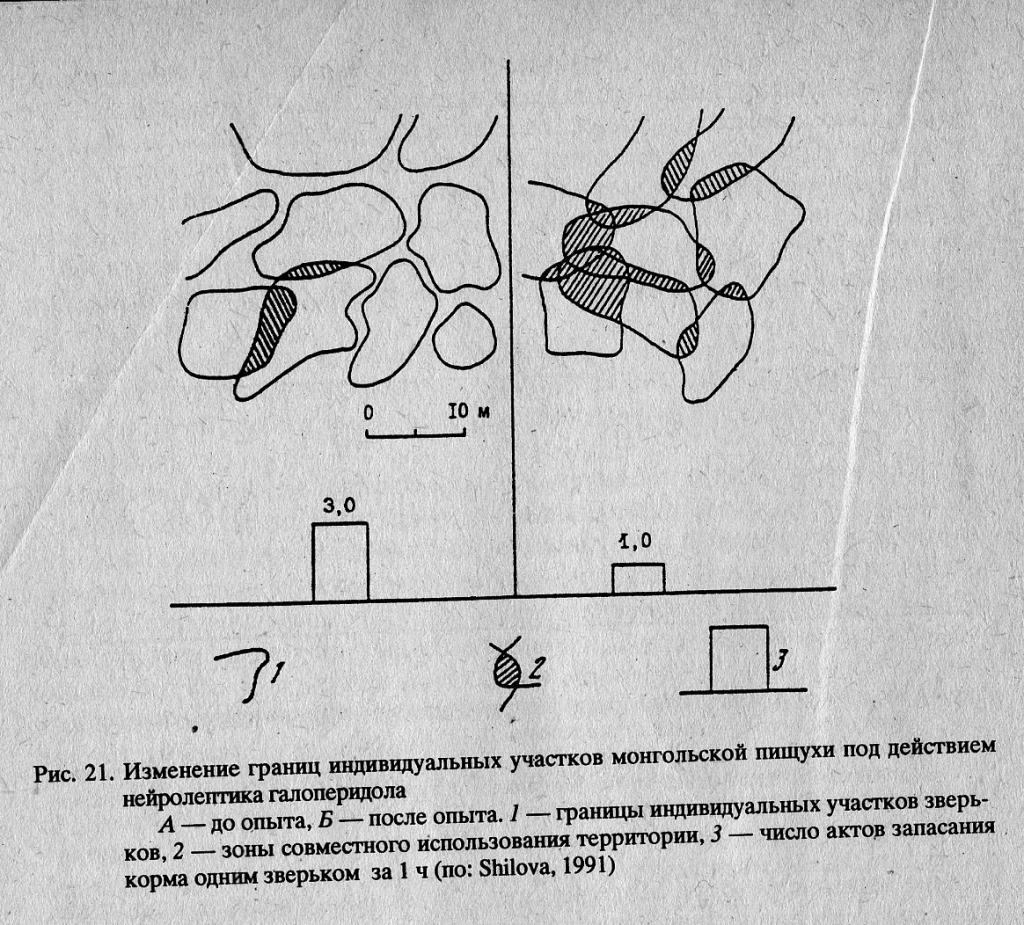
Вследствие хаотичного использования территории у хозяев участка значительно сократилось время запасания корма — одного из важнейших поведенческих актов в период подготовки к зиме. В июне 1978 г. на территориях, обработанных в предыдущий период галоперидолом, пищухи отсутствовали.
Таким образом, у вида, для которого оптимальная популяционная структура зависит в основном от агрессивных взаимодействий, применение нейролептиков вызвало нарушения, сопровождающиеся падением численности.
Следует, однако, отметить, что многие поведенческие акты пищух оказались достаточно стабильными. Так, мы пытались проследить, насколько повышение индивидуальной агрессивности между самками и молодыми зверьками может ускорить процесс расселения молодняка. С этой целью был использован парнат — один из антидепрессантов, вызывающий резкий подъем агрессивных контактов у белых мышей в экспериментальных условиях. Парнат применен в дозировках 2—10 мг/кг на опытном участке, населенном монгольскими пищухами.
У зверьков уровень агрессии значительно возрос (до введения препарата за 1 ч наблюдений — 0,5 агрессивных контактов; после введения — 2,4). Однако расселение молодняка на экспериментальном участке произошло в те же сроки, что и на контроле (через 18—20 дней после выхода на поверхность). Территориальные взаимоотношения зверьков после получения парната не изменились.
Монгольская песчанка
Пространственно-этологическая структура монгольских песчанок характеризуется семейно-групповым образом жизни. Семейная группа обычно совместно использует территорию и состоит из взрослого самца, одной или двух взрослых самок и их подросших детей. Внутри семейной группы образуются тесные социальные контакты, поддерживающие стабильность группировки. Выражено распределение поведенческих функций между особями: некоторым животным (особенно взрослым самцам) принадлежит ведущая роль в охране территории, маркировке участков и т.д. Семейная группа обладает высокой стабильностью социальных связей. Территория семьи охраняется от соседних группировок. В охране территории принимают участие все члены группы (Громов, Попов, 1979; Громов, 1981; Орленев, Перепадов, 1981).
В первом варианте наших опытов под наблюдением находилось две семейные группы монгольских песчанок, заселяющие участие злаково-полынной степи вблизи старых животноводческих строений. Одна семейная группа (1 самец, 2 самки, 6 молодых песчанок) получала резерпин. Вторая семейная группа (взрослый самец, самка и 5 молодых зверьков) служила контролем.
При дозировке препарата в 1,3 мг/кг у песчанок стали проявляться клинические симптомы отравления. У всех без исключения членов опытной семьи наблюдался птоз (опущение век), зверьки подолгу неподвижно сидели у своих нор, слабо реагируя на окружение.
Однако несмотря на ярко выраженное индивидуальное отклонение в поведении песчанок, внутрисемейные отношения зверьков не изменились. Это явление мы объясняем тем, что при описанном типе пространственно-этологической структуры (совместное использование территории членами одной семьи, более мягкие формы междусемейных контактов), снижение агрессивности не может определить существенных нарушений во внутрисемейной группировке.
Аналогичные результаты мы получили, применяя транквилизатор амизил (0,5—1,0 мг/кг). Экспериментальная семейная группа песчанок также занимала участок в злаково-полынной степи и состояла из самца-доминанта, взрослой самки, самца-субдоминанта и молодых зверьков.
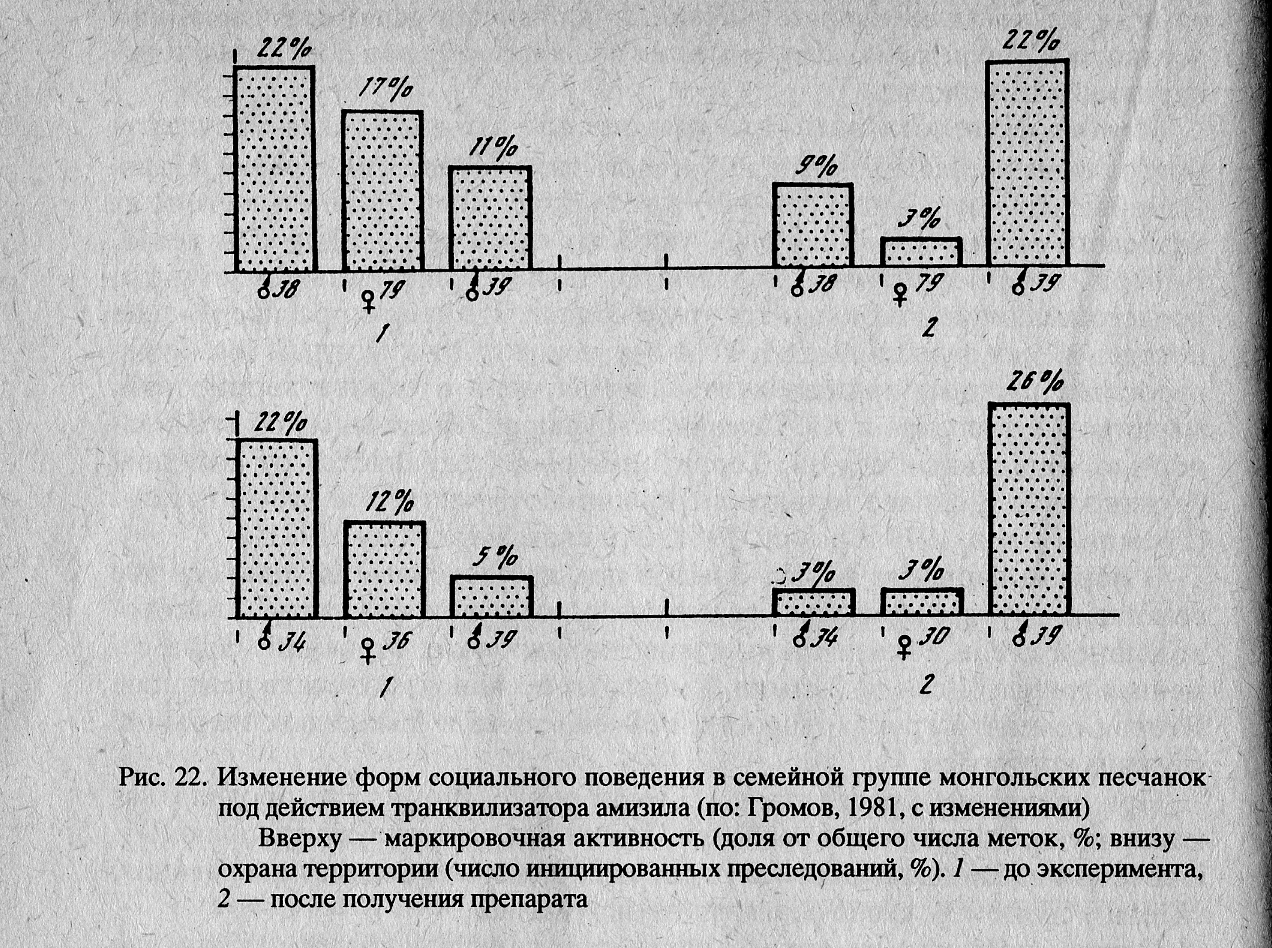
В результате действия амизила у самца-доминанта и у взрослой самки существенно нарушились две основные функции, способствующие охране территории: преследование соседей и мечение границ семейного участка.
После введения препарата маркировочная активность самца-доминанта снизилась в 3 раза, функция преследования — в 8 раз. У самки, соответственно, в 6 и 4 раза (Громов и др., 1981). Однако произошло перераспределение социальных функций. В охране границ семейного участка стали принимать участие другие члены группы (рис. 22). В результате этого явления общие границы и структура группы полностью сохранились.
Иная картина наблюдалась при экспериментах с семейными группами монгольских песчанок, имеющих наиболее простую структуру (один самец, одна-две взрослые самки) (Громов, Попов, 1979).
Нейролептик галоперидол был применен в двух таких семейных группах, территории которых соприкасались (дозировка препарата — 0,1—5,0 мг/кг). После получения галоперидола общее число агрессивных межсемейных взаимодействий между самцами и самками значительно упало. Функции охраны территории осуществлять было некому. Соответственно, произошло значительное перераспределение территории между зверьками.
Полуденная песчанка
Как уже говорилось, по сравнению с монгольской песчанкой полуденная обладает слабой социальной активностью. По нашим данным, частота социальных взаимоотношений в семейных группах монгольских песчанок в 8—10 раз выше, чем в поселениях полуденных песчанок (наблюдения в природе; Шилова, Громов, 1983). В основе специфики использования территории у полуденных песчанок лежит индивидуальная привязанность особи к определенному участку (Попов, 1981; Чабовский, 1986).
Эксперимент по воздействию галоперидолом на полуденных песчанок проведен нами осенью 1983 г. в Калмыкии на участке, где в течение трех предыдущих лет были детально изучены пространственное распределение и социальные отношения зверьков (Шилова, Дервиз и др., 1983). Площадь экспериментального участка составляла 1 га, на котором постоянно жили 37 песчанок, образуя три группировки. В течение 3 недель зверьков отлавливали в живоловки, где приманкой служил хлеб с растительным маслом, содержащим определенную навеску препарата, которую зверьки, попавшие в ловушки, съедали. Таким образом, 30 песчанок регулярно получали галоперидол в дозировке до 2,5 мг/кг.
Известно, что в качестве критерия интенсивности социальных контактов зверьков можно использовать показатель парных поимок в живоловки (Getz, Carter, 1980; Novak, 1983; Cassing, 1986 и др.). Наши многолетние отловы полуденных песчанок в живоловки показали, что в обычных ситуациях парные поимки этих зверьков практически не встречаются, что еще раз подчеркивает низкий уровень их социальных контактов.
Совершенно иная картина наблюдалась после получения зверьками галоперидола: на контрольной территории за 3 года наблюдений общее число поимок полуденных песчанок в живоловки составило 1177, парных поимок — 1 (0,08%). На опытном участке за месяц наблюдений получено 573 поимки, из которых парные составляли 22 (3,83%). Следует отметить, что в живоловки совместно ловились не только молодые зверьки, но чаще всего взрослые аборигены площадки — как самцы, так и самки. Соответственно, этот показатель в эксперименте возрос почти в 50 раз.
Однако несмотря на то, что галоперидол существенно облегчил социальные контакты между полуденными песчанками, принципы территориального распределения экспериментальной группы не изменились. Зверьки всех возрастных и половых категорий с удивительным постоянством продолжали встречаться в тех же местах, где они жили до начала опыта. Этот факт еще раз подчеркивает, что территориальное распределение полуденных песчанок мало зависит от характера их социальных взаимодействий.

Длиннохвостый суслик
Пространственно-этологическая структура длиннохвостого суслика изучена нами в Туве (Шилова и др., 1979). На обследованных территориях длиннохвостый суслик образовывал очаговые поселения, приуроченные к выровненным участкам межгорий с преобладанием злаково-полынных ассоциаций. Наиболее высокая плотность отмечалась возле старых строений животноводов (до 20—30 зверьков на 1 га). Для длиннохвостого суслика характерно совместное использование территории всеми зверьками, входящими в группировку.
Этологическая структура внутрипопуляционных группировок характеризуется четко выраженной иерархией. Для определения рангового положения зверька в нашем эксперименте высчитывалось число побед и поражений данной особи. Разность принимали за показатель иерархической значимости зверька.
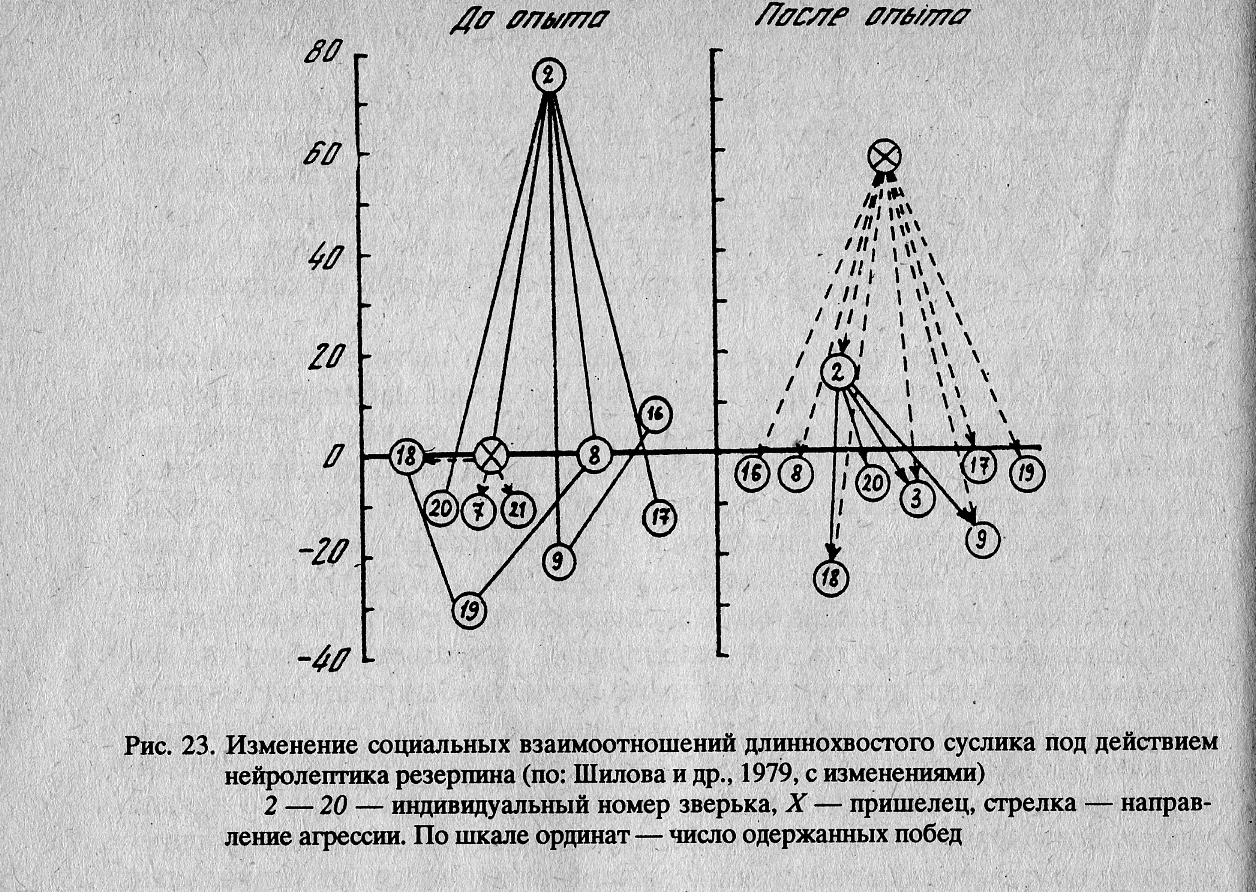 Рис. 23, а характеризует положение в группировке с конца июля. Этологическая структура в этот период отличалась упорядоченностью взаимоотношений. Во главе группировки находился один доминант, функции которого заключались в изгнании пришельцев и в контроле за поведением других зверьков, входящих в группировку. Остальные члены группировки занимали подчиненное положение. Агрессивные контакты между ними наблюдались крайне редко, а между особями, находящимися на близких иерархических уровнях, не наблюдались совсем.
Рис. 23, а характеризует положение в группировке с конца июля. Этологическая структура в этот период отличалась упорядоченностью взаимоотношений. Во главе группировки находился один доминант, функции которого заключались в изгнании пришельцев и в контроле за поведением других зверьков, входящих в группировку. Остальные члены группировки занимали подчиненное положение. Агрессивные контакты между ними наблюдались крайне редко, а между особями, находящимися на близких иерархических уровнях, не наблюдались совсем.
Поскольку приведенные выше данные показали значительную роль агрессивности в формировании внутрипопуляционных группировок длиннохвостого суслика, в качестве воздействующего агента на популяционную структуру был выбран резерпин, снижающий агрессивность зверьков.
В первом варианте опытов зверьки получили препарат 1 раз перед началом расселения молодняка. В результате мы имели возможность проследить взаимоотношения зверьков — членов группировки со зверьками-пришельцами, не подвергавшимися действию резерпина, и, следовательно, выяснить роль агрессивного поведения во взаимоотношениях с ’’чужими” зверьками, а также значение агрессивности для процесса расселения молодняка.
Одноразовое применение препарата вызвало серьезные последствия для группировки (Шилова и др., 1979). В первую очередь крупные изменения произошли в этологической структуре (рис. 23а, б). После применения резерпина снижение агрессивности зверьков — членов группировки не повлияло на общий принцип построения структуры, однако доминант (самец № 2) потерял свое высокое ранговое положение в группировке и его место было занято пришельцем (немеченный зверек), не получавшим препарат. Подобной смены доминанта в этот период на контрольных участках не наблюдалось. Смещение доминанта пришельцем было вызвано заметными изменениями в поведении. Так, до начала опыта доминант, замечая на своей территории чужого зверька, бежал к нему, издавая характерные агрессивные сигналы и после непродолжительной драки (или вовсе без нее) изгонял пришельца с участка. После применения резерпина этот самец, едва увидев незнакомого зверька, спешил скрыться в ближайшей норе. Аналогичным образом вели себя и другие бывшие хозяева территории.
Изменения в поведении сусликов, населяющих участок, вызванные действием резерпина, привели к тому, что одна и та же кормовая территория стала использоваться как бы в две смены. Зверьки, получившие резерпин, выходила на поверхность тогда, когда поблизости не было других сусликов. До начала опыта пришельцы, как правило, не успевали кормиться на территории установившейся семейной группировки, так как немедленно подвергались нападению доминанта. После применения резерпина они получили такую возможность.
Таким образом, после применения резерпина общий принцип построения иерархической структуры группировки длиннохвостого суслика сохранился, но произошли значительные изменения в расположении отдельных особей на ступенях иерархической лестницы. Так, самец № 2, занимавший ранее доминантное положение, теперь сам оказался в подчинении у пришельцев, хотя по отношению к другим членам группы он сохранял свое превосходство. До применения резерпина и на аналогичных контрольных территориях мы не наблюдали никаких отклонений в поведении зверьков. Этологическая структура внутрисемейных группировок сохранилась до момента залегания сусликов в спячку.
Следует, однако, отметить, что несмотря на снижение агрессивных контактов на опытном участке, расселение молодняка все же произошло. Это говорит о том, что агрессия, видимо, не является единственным фактором, определяющим этот процесс.
Описанные изменения в пространственно-этологической структуре после искусственного снижения агрессивности говорят о большом значении этой формы поведения в поддержании гомеостаза популяционной структуры. Произошедшие после применения резерпина изменения в характере использования территории длиннохвостым сусликом могут иметь неблагоприятные последствия для особей, входящих в группировку.

Домовая мышь
Синантропные популяции этого вида характеризуются приуроченностью семейной группировки к определенной небольшой территории, часто ограниченной одним помещением. Самцы-доминанты обладают высокой степенью агрессивности по отношению к членам других группировок. Для домовой мыши активная защита индивидуального участка — видоспецифический фактор, понижающий уровень стресса самцов.
В качестве агента, снижающего агрессивность самцов домовой мыши, был применен аминазин (Каменов, Золотарев, 19806). Препарат в дозе 1,9 мг/кг был введен перорально зверькам перед ссаживанием их на нейтральной территории. До начала эксперимента среднее число агрессивных контактов зверьков за 1 мин у 10 пар составило 1,85 ± 0,76. После введения препарата агрессивность самцов резко снижалась (0,02 ±0,01), различия с контролем статистически достоверны (р < 0,01). Полученные данные позволили применить аминазин для выяснения вопроса о реакции «неагрессивных» самцов на ’’высокоагрессивных” зверьков, подвергшихся воздействию препарата. После получения препарата в количестве 3,8 мг/кг агрессивность доминантов значительно упала (5,85 ± 1,84 до начала опытов, 1,79 ± 0,66 при эксперименте). Однако ’’неагрессивные” самцы, занимавшие ранее подчиненное положение, не использовали сложившейся ситуации и не пытались занять доминирующего положения. Через неделю агрессивность доминантов восстановилась (5,14 ± 0,37).
При введении аминазина (3,8 мг/кг) зверькам-доминантам, освоившим свою территорию, наблюдалось лишь временное снижение агрессивности по отношению к ’’пришельцам”. Сложившиеся ранее территориальные и иерархические отношения не нарушались.

Лесная мышь
Серия экспериментальных работ по изменению поведенческих реакций лесной мыши выполнена нами на Звенигородской биостанции Московского университета совместно с Ю.М. Смириным (Смирин, Шилова и др., 1985; Смирин, Шилова, 1989). Как показали ранее исследования Ю.М. Смирина (1977, 1979) и других авторов (Gumell, 1977; Montgomery, 1989; и др.), в отличие от домовых мышей у лесной мыши в стабильных группах принципы деспотического доминирования отсутствуют. Функции охраны территории, занятой группировкой выполняют все члены группы. Группы, освоившие определенную территорию, обладают большой устойчивостью. Между членами группы вне периода размножения агонистические взаимоотношения отсутствуют. Все члены группы проявляют крайнюю степень агрессивности в отношении интродуцированных мышей. В закрытом помещении интродукция пришельца всегда кончается его убийством. Лесная мышь — удобная модель для выяснения вопроса 6 том, насколько повышение (или снижение) двигательной активности и, соответственно, увеличение (или падение) частоты контактов может изменить уровень внутригрупповой или межгрупповой агрессии. Соответственно, в экспериментах были использованы два альтернативных фармакологических препарата: аминазин, снижающий двигательную активность, и сиднокарб, увеличивающий подвижность зверьков (Альтшуллер и др., 1974).
В первом варианте опытов изучено действие перечисленных выше препаратов на стабильную группу лесных мышей в зимний период (Смирин, Шилова и др., 1985). До начала эксперимента течение 4 мес. (период наблюдений) между зверьками в группе агонистические взаимоотношения полностью отсутствовали. Инициаторами дружелюбных контактов (скучивание, назо-назальные, назо-анальные, чистка) были, как правило, все члены группы; доминантных особей не выделялось. В отношении к интродуцированным лесным мышам (взрослые самцы и самки) все члены группы проявляли ярко выраженную агрессию: число агрессивных взаимодействий с ’’пришельцами” составляла 90,0% всех контактов, а время, затраченное на погони и схватки, — 48,5% от всего периода активности.
В ноябре все члены группы получили аминазин в дозировке 5 мг/кг на каждого зверька. После получения препарата активность зверьков значительно упала. Соответственно, частота контактов с ’’пришельцами” снизилась почти в 6 раз. Время, затраченное на погони и схватки, составляло 3,8% от периода активности. Но нам важно отметить, что снижение двигательной активности лесных мышей под действием аминазина не повлияло на уровень агрессивности группы по отношению к ’’пришельцу”. Несмотря на сокращение общего числа контактов с интродуцированным зверьком 83,3% из них составляли нападения и схватки. Через 5 суток подсаженный зверек погиб.
Таким образом, несмотря на значительное сокращение количеств контактов аборигенов группы с пришельцем, агрессивность членов группы лесных мышей по отношению к чужаку не исчезла и не заменилась другими формами поведения.
В следующем опыте зверьки получили препарат сиднокарб (100 мг/кг). Характер индивидуального поведения лесных мышей под действием препарата значительно изменился: заметно обострилась реакция и внешние раздражители и вследствие возбуждения и высокой двигательной активности интенсивность контактов внутри группы возросла более чем 3 раза. Однако увеличение числа контактов между зверьками-аборигенами не повлекло за собой возникновение внутригрупповой агрессия Из 84 зарегистрированных в этот период внутригрупповых контактов агрессивных взаимодействий не было.
При подсадке в помещение нового самца лесной мыши интенсивность контактов аборигенного населения с пришельцами под действием сиднокарба значительно возросла: за первые 2 ч наблюдений было за регистрировано 164 контакта; 70% времени, проведенного зверьками вне убежищ, пришлось на агрессивные взаимодействия с пришельцами (в контроле 48,5%). Все контакты носили агрессивный характер.
Таким образом, группировки лесных мышей, освоивших определенную территорию, обладают большой устойчивость) и проявляют высокую степень агрессивности в отношении зверьков, шатающихся проникнуть на освоенную ими площадь. Агрессивность в отношении пришельцев проявляют все члены группировки. Интенсивность контактов с пришельцем коррелирует с индивидуальной подвижностью особей.
Снижение активности группировки путем применения нейролептика резко сократило число контактов с пришельцами, значительно уменьшилось время агрессии, направленной на пришельца, но не снимало агрессивности как ведущей формы поведения в отношении интродуцированного зверька Агрессивные контакты в отношении пришельца не сменяются другим формами поведения, что еще раз подчеркивает высокую устойчивость ведущих поведенческих актов животных в сложившихся группировках.
Повышение активности членов группировки вызывает значительное увеличение количества и времени агрессивных контактов в отношения пришельца, но не влияет на взаимоотношения особей внутри группировки что, видимо, также обусловлено высокой степенью устойчивости популяционных систем».
Популяционная экология как основа контроля численности мелких млекопитающих. М.: Наука, 1993. С.140-155.
* * *
Изложенные выше данные показывают, что интенсивность воздействия фармакологических средств на популяционную структуру вида коррелирует с уровнем сложности пространственно-этологических параметров. Чем проще пространственная и социальная организация вида, тем легче разрушаются внутрипопуляционные группировки. Так, среди изученных нами видов наиболее простая социальная организация свойственна монгольской пищухе. Ведущей формой взаимоотношений с соседями у этого вида является агонистическое поведение, что способствует строгому разграничению индивидуальных участков. Соответственно, снятие агрессивности у монгольских пищух с помощью нейролептиков (резерпин, галоперидол) повлекло за собой существенные нарушения пространсгвенно-этологической структуры.
В семьях монгольских песчанок существенные изменения территориального поведения, вызванные искусственным снижением агрессивности, происходят в том случае, если группа обладает простой структурой и никто не может заменить доминанта в период нарушения его поведенческих функций. В более сложных семьях монгольских песчанок при резком искусственном изменении социальных форм поведения происходит перераспределение поведенческих функций между особями, вследствие чего популяционная структура сохраняет свою целостность.
Сравнительно простое строение имеет социальная структура поселений длиннохвостого суслика (защиту территории и упорядочение внутригрупповых отношений осуществляет один самец-доминант). Соответственно, снижение агрессивности доминанта способствует использованию территории семейной группы зверьками-пришельцами, что существенно нарушает нормальный характер жизнедеятельности хозяев участка.
С другой стороны, применение нейролептиков не оказало существенного влияния на внутрипопуляционные группировки домовой мыши. После снятия агрессивности у самца-доминанта, низкоранговые особи не претендовали на его роль, а доминант мог восстановить свое положение даже через неделю после начала эксперимента. Известно, что домовым мышам свойственна популяционная структура, построенная по принципу деспотического доминирования, в которой поддержание гомеостаза определяется целым набором разнообразных социальных взаимодействий.
В популяционных группах лесной мыши, несмотря на значительное увеличение частоты внутригрупповых контактов, агрессивность не проявилась, и группа, соответственно, сохранила свою целостность. Скорее всего, факторы, определяющие возникновение (или падение) агрессивности в группе, зависят от комплекса разнообразных причин, которые, как, впрочем, и другие важнейшие формы социального поведения, еще мало известны. Можно надеяться, что эксперименты по направленному изменению поведенческих актов различных видов животных могут способствовать дальнейшему развитию этих исследований.
Экспериментальное воздействие на агрессивное поведение: В.С.Громов, 2000
«Принимая во внимание, что агрессивное поведение имеет важное значение как средство регулирования территориальных отношении у животных и, по сути, является одним из основных этологических механизмов индивидуализации пространства, можно полагать, что искусственное подавление или стимуляция агрессивных реакции может оказать деструктивное или, наоборот, эффективное влияние на сложный комплекс территориального поведения и привести к нарушению динамического равновесия сложившейся пространственной структуры внутрипопуляционных группировок. Экспериментальные исследования такого рода, с одной стороны, важны для более углублённого и всестороннего изучения феномена территориальной агрессии, а с другой стороны, позволяют вести поиски эффективных средств и методов управления поведением животных.
Один из эффективных способов воздействия на агрессивное поведение животных связан с применением фармакологических психотропных препаратов. К настоящему времени детально разработана методология этолого-фармакологических исследований агрессивного поведения, синтезированы и опробованы многие препараты как агрессивного, так и агрессогенного действия (Вальдман, 1980, 1982; Вальдман, Пошивалов, 1984; Судаков, 1984; Пошивалов, 1986).
В их исследованиях объектом для подобного рода экспериментов служили монгольские песчанки, обладающие ярко выраженным территориальным поведением, а в качестве психотропных средств были выбраны транквилизатор галоперидол и антидепрессант парнат.
1. Опыт с транквилизатором галоперидолом
Цель опыта с галоперидолом, проведенного в полевых условиях, заключалась в попытке изменения территориальных отношений монгольских песчанок путем подавления агрессивной реакции резидентов на «чужаков» (Громов, Попов, 1979) Известно, что галоперидол подавляет мотивированную агрессию у грызунов (Буров, Салимов, 1975). Эффективные дозировки галоперидола находятся в интервале 0.1-5.0 мг/кг, при этом в дозе 0.2 мг/кг галоперидол проявляет себя как транквилизатор, а свыше 2.0 мг/кг — как нейролептик с выраженным седативным эффектом (Вальдман и др., 1976). Существенным эффектом действия галоперидола является нарушение адекватности поведения: вначале это проявляется в замедленной реакции на изменение ситуации, а по мере увеличения дозы нейролептика обнаруживается неспособность к завершению целенаправленных действий (Каткова, 1975).
В качестве подопытной была выбрана группировка монгольских песчанок на кошарном поселении в составе двух семейных групп (1 самца, 3 самки), одиночной самки A66G и одиночного самца №50 (рис. 39). Следует отметить, что для кошарных поселений монгольских песчанок характерна одна особенность, отличающая их от других локальных поселений этого вида, которая заключается в существенном ограничении пространственных и кормовых ресурсов. К самым оптимальным с этой точки зрения необходимо отнести места скопления помета внутри загонов для скота, перемешанного с остатками зерна, которые имеют ограниченную площадь, не превышающую 1-1.5 га. Ближайшие участки степи за пределами территории кошары с плотными щебнистыми почвами практически непригодны для заселения. В связи с этим кошарные группировки песчанок обычно состоят не более, чем из 2-3 семей.
Мигранты, попадающие на кошарное поселение с установившейся территориальной структурой, или особи, по тем или иным причинам покидающие семейные группы, вынуждены занимать худшие участки обитания на периферии кошарной группировки, мало пригодные для длительной эксплуатации. Такие особи долго не задерживаются на кошарных поселениях и, в конечном итоге, либо погибают, либо отправляются поиски более оптимальных мест обитания.
Возвратимся, однако, к эксперименту с галоперидолом, для проведения которого на территории двух семейных групп в составе двух самцов (№37 и №45) и трех самок (№25, №39 и №40) ежедневно раскладывали хлебные брикеты размером 2x2x0.5 см с суспензией галоперидола в растительном масле. Песчанки получали необходимую дозу препарата, съедая один брикет. Опыт длился с 21 июля по 30 августа. Дозировку препарата по ходу эксперимента увеличили с 0.1 до 5.0 мг/кг (рис. 38). Контролем служила группировка монгольских песчанок на аналогичном кошарном поселении в составе самцов, 3 самок и 3 выводков молодых зверьков. Результаты эксперимента представлены графически на рис. 38-40.
Графики на рис. 38 отражают уровень агрессивных взаимодействий между песчанками на экспериментальном и контрольном участках. Кривые 1 и 2 на рисунке сглажены методом скользящей средней и отражают динамику агрессивных контактов по ходу эксперимента. После получения препарата в дозировке 0.1-0.5 мг/кг у подопытных песчанок наблюдалось постепенное увеличение числа агрессивных взаимодействий (рис. 38, 1), затем, при увеличении дозы до 1.0 мг/кг, последовал резкий спад и далее, на фоне действия препарата в дозировке 2.0-5.0 мг/кг, снижение числа территориальных конфликтов между взрослыми особями практически до нуля. Вместе с тем, число нарушений границ семейно-групповых участков подопытных песчанок со стороны особей-соседей оставалось в среднем постоянным на протяжении всего эксперимента и, следовательно, падение уровня агрессивных контактов нельзя было связать с уменьшением числа нарушений границ. На контрольном участке уровень агрессивных контактов хотя и был более низким, но достаточно стабильным на протяжении всего периода наблюдений (рис. 38, 2).
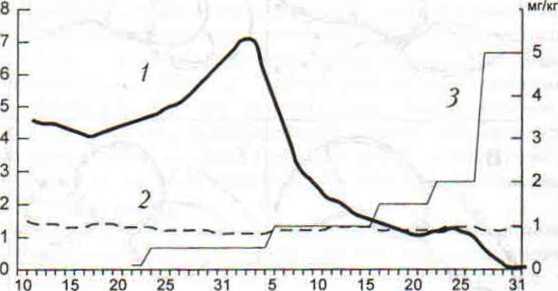
Рис. 38 Динамика агрессивных взаимодействий монгольских песчанок на экспериментальном (1) и контрольном (2) участках. Левая шкала — число агрессивных контактов за 1 час наблюдений, правая шкала — доза препарата (3).
Анализ поведения каждой из подопытных песчанок показал, что в результате действия препарата резко снизилась частота агрессивных контактов у трех зверьков (№45, №39 и №25), регулярно поедавших брикеты с галоперидолом. Однако, у самки №40 и самца №37, также регулярно получавших препарат, аналогичные изменения в поведении отсутствовали. Возможно, эти два зверька получали меньшую дозу препарата, не полностью съедая брикеты, либо оказались индивидуально более устойчивыми к его действию.
Интересен факт, что по ходу эксперимента в первую очередь снизилось число агрессивных контактов между особями-соседями, тогда как по отношению к другим членам группировки у подопытных песчанок до определенного периода сохранялась устойчивая агрессивная реакция. Возможно, увеличение порога агрессивной реакции под влиянием небольших (транквилизирующих) доз галоперидола оказалось достаточным для предотвращения территориальных конфликтов между знакомыми особями, отношения которых основаны преимущественно на ритуализованных формах поведения. Для угнетения агрессивной реакции на зверьков, не являющихся ближайшим соседями, потребовались значительно большие, седативные дозы препарата.
Параллельно с динамичными изменениями агрессивной реакции подопытных песчанок, по ходу эксперимента изменилась территориальная структура группировки. На рис. 39, А отражена картина пространственного размещения семейно-групповых и индивидуальных участков обитания песчанок до начала опыта, где они разделены четко определенными и охраняемыми границами. После получения минимальных доз галоперидола ситуация резко изменилась (рис. 39, Б): подопытные зверьки стали преследовать чужаков не только на своей территории, но и на соседних участках. Вследствие этого границы между семейно-групповыми территориями и индивидуальными участками самок стали неопределенными. Именно к этому периоду относится резкое повышение числа агрессивных контактов, показанное на рис. 38.
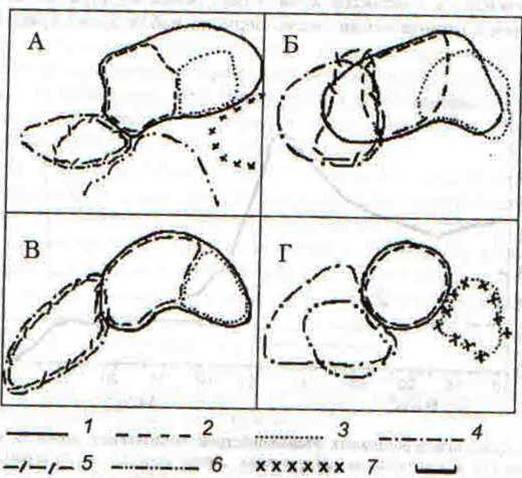
Рис. 39. Пространственное размещение границ семейно-групповых и индивидуальных территорий монгольских песчанок на экспериментальном участке А — до начала опыта; Б. В, Г — в I, II и III периодах наблюдений, соответственно 1 — самец №45, 2- самка №40, 3 — самка №25, 4 — самец №37, 5 — самка №39. б — самка №66, 7 — самец №50. Масштаб 10 м.
Описанные изменения в поведении песчанок легко объясняются эффектом малых доз галоперидола. В норме хозяин участка, как об этом уже неоднократно говорилось, нападает на любую чужую особь, изгоняя ее за пределы охраняемой территории. Во время преследования оба зверька могут оказаться на участке обитания преследуемого, а в этой ситуации они меняются ролями. Под влиянием небольших доз галоперидола адекватность поведения нарушается и реакция песчанок на изменение ситуации замедляется. Преследование прекращается не сразу, а спустя некоторое время, пока действие стимулов внешней среды окончательно не затормозит реакцию преследования у одной особи и не вызовет ответную реакцию у другой. Подобная дисфункция поведения может быть также результатом психостимулирующего эффекта малых доз галоперидола. Таким образом, под влиянием небольших доз транквилизатора нарушение адекватности поведения происходило на фоне снижения порога агрессивной реакции. В итоге это не могло не привести к дестабилизации сложившейся территориальной структуры.
С увеличением дозы препарата частота агрессивных взаимодействий песчанок снизилась, и территориальные отношения вновь стабилизировались (рис. 39, В), что объяснятся седативным, т.е. успокаивающим, действием более высоких доз галоперидола. Однако, стабилизация территориальных отношений произошла в качественно иной ситуации, когда изменились и прежние границы участков обитания, и первоначальный состав семейных групп. В результате у некоторых особей изменилось исходное территориальное положение в группировке. Так, например, самец №37 и самка №39 были вытеснены зверьками №45 и №40 с некоторой части своей территории, а самка №25 уступила часть своего участка самке №40.
Дальнейшее увеличение дозы галоперидола привело к еще большему усугублению ситуации (рис. 39, Г): семейная группа самца №45 распалась, и самка №25 присоединилась к одиночному самцу №50, участок обитания которого находился на периферии группировки, так что сформировалась новая пара. Самка №39 к концу опыта вообще исчезла с площадки наблюдений, а ее место в группе самца №37 заняла самка №66, которая по ходу эксперимента предпринимала активные попытки вытеснить самку №39 с занимаемого ею участка, и, когда последняя исчезла, обосновалась на ее территории на пару с самцом №37.
К концу опыта напряженность территориальных конфликтов снизилась до минимума Преследования приобрели односторонний характер, схваток практически не наблюдали, уменьшилось число “противостояний». Песчанки медленнее реагировали на появление незнакомых особей, иногда адекватная реакция вообще отсутствовала Преследования зачастую прекращались без видимых причин. Эти дисфункции поведения можно объяснить значительным увеличением латентного периода агрессивной реакции и увеличением ее порога в результате действия максимальных доз галоперидола.
Из пяти подопытных песчанок, наиболее существенные нарушения в поведении произошли у самки №39. Число агрессивных взаимодействий, инициированных этой особью, резко снизилось уже в начале эксперимента. Под давлением конкурентов самка была вынуждена переселиться со своей территории на пустующую нору, расположенную на периферии участка наблюдений, где у нее родились детеныши. Через некоторое время она вместе с выводком перешла в другую нору, ближе к своему бывшему участку обитания, но и здесь ей не удалось закрепиться надолго, так как вскоре ее окончательно вытеснила самка №66, с которой остались и детеныши самки №39. К середине эксперимента значительные отклонения в поведении стали проявляться и у самца №45: общая активность этого зверька заметно снизилась, особенно число агрессивных контактов с особями-соседями интенсивность маркировки территории.
Нами проведена количественная оценка некоторых форм агонистического поведения песчанок на протяжении эксперимента (Громов, Попов, 1979). В качестве критерия была выбрана величина, отражающая относительную частоту проявления той или иной формы территориальной агрессии в общем наборе поведенческих реакций животного. Этот критерий обеспечивает возможность сравнения изменений в поведении зверьков с различным уровнем активности и независимо от изменения активности каждого зверька в течение опыта.
Для сравнения результатов были выделены три периода в соответствии с увеличением дозы препарата: I — 0.1-0.4 мг/кг, II — 0.5-1.0 мг/кг, III — 2.0-5.0 мг/кг Необходимо отметить, что во II периоде зверьки №37 и №39 были вытеснены с той части своей территории, на которой находились брикеты с галоперидолом, вследствие чего потребление препарата ими было нерегулярным, особенно в III периоде.
Анализ динамики поведения песчанок в течение опыта показал, что во II периоде у самцов №45 и №37 значительно снизилась относительная частота преследований (различия достоверны, 9.17 < Х2 < 21,26, 0.003<р<0.0001). В III периоде частота преследований у самца №45 оставалась по прежнему низкой, а у самца №37 резко возросла, что, можно объяснить значительным снижением эффекта препарата из-за нерегулярного поедания брикетов с галоперидолом этим зверьком на заключительной фазе эксперимента (рис. 40, А). У самок №25 и №39 относительная частота преследований и ритуальных пограничных демонстраций («противостояний) также резко снизилась в III периоде (различия достоверны, 17.89<χ2<26.44, р=0.0001), но у самки №40 существенных изменений в территориальном поведении, кроме снижения частоты «противостояний», не произошло, несмотря на регулярное поедание брикетов с галоперидолом (рис. 40, Б, В).
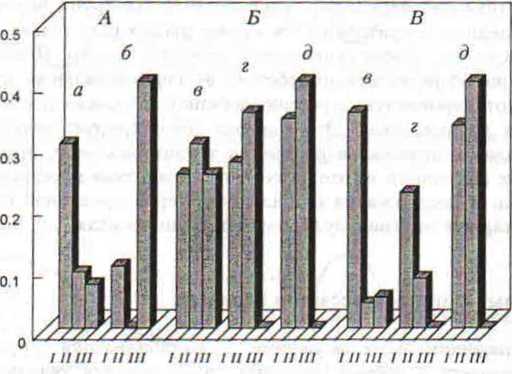
Рис. 40. Изменение относительной частоты преследований (A, Б) и ритуальных пограничных демонстраций (В) у самцов (A) и самок (Б, В) монгольской песчанки под влиянием галоперидола. По вертикали — относительная частота инициированных контактов. I, II, III — периоды наблюдений; а — самец №45, б — самец №37, в — самка №40, г — самка №25, д — самка №39
Таким образом, можно констатировать, что цель эксперимента была успешно достигнута: у песчанок, регулярно получавших необходимую дозу галоперидола, существенно повысился порог агрессивной реакции, в связи с чем они оказались неспособными эффективно защищать занимаемые участки обитания, которые в результате были частично или полностью оккупированы конкурентами; существовавшие до эксперимента семейные группы распались и сформировались новые семейные пары Следует отметить, что действие галоперидола было избирательным: самка №40 оказалась устойчивой к препарату и сохранила за собой занимаемый участок обитания. Высокая устойчивость к действию галоперидола была обнаружена и у детенышей самки №25, вышедших на поверхность к концу эксперимента. Молодые песчанки из выводков самок №25 и №39, регулярно получавшие небольшие дозы транквилизатора, так как имели доступ к брикетам с галоперидолом, демонстрировали высокий уровень агрессивности ко всем незнакомым особям независимо от их пола и возраста.
Анализируя результаты проведенного эксперимента, следует констатировать, что семьи монгольских песчанок являются чрезвычайно устойчивыми образованиями, о чем уже говорилось в главе 2. То, что удалось получить в результате воздействия на поведение песчанок нейролептика галоперидола — распад семейных групп вкупе с перераспределением территории, — в норме происходит только в случае гибели взрослых особей-основателей семейной группы. Показательно, что того же эффекта можно добиться экспериментальным путем, подавляя мотивированную агрессию песчанок в сложившихся территориальных группировках. Это еще раз демонстрирует ведущую роль внутривидовой агрессии в регуляции территориальных отношений у животных как одного из этологических механизмов пространственной сегрегации и поддержания стабильности территориальной структуры в элементарных внутрипопуляционных группировках.
[Эксперимент также показал возможность перераспределения общегрупповых функций и социальных ролей в ответ на лекарственные воздействия, уменьшившие агрессивность, что и обеспечило наблюдаемую устойчивость социума. Хотя к концу эксперимента при максимальных дозах нейролептика социальная активность особей была значительно подавлена: время пребывания на поверхности сократилось на 34%, интенсивность маркировки упала в 2,7 раза, но общее число преследований животных из двух контрольных групп увеличилось в 1, 4 раза – при том, что у отдельных зверьков оно в 1,5 раза упало.
Дело в том, что чувствительность к галоперидолу взрослых особей оказалась существенно выше, чем у молодых. Взрослые демонстрировали общее угнетение маркировки, самка №8 также и охраны территории. Однако эффективность охраны все 35 дней эксперимента оставалась на достаточно высоком уровне именно потому, что снижение активности у взрослых особей компенсировалось адекватным повышением активности молодых зверьков.
Территориальные конкуренты – особи контрольных групп за время опыта оккупировали только незначительную часть пространства экспериментальной группы, а когда после опыта поведение её членов вошло в норму, участки все трёх групп восстановились в исходных границах.]
3.3.2. Опыт с антидепрессантом парнатом
В описанном выше эксперименте дестабилизация территориальных отношений и распад семейных групп явились результатом подавления агрессивной реакции подопытных песчанок. Теоретически можно представить, что тот же дестабилизирующий эффект может быть достигнут при использовании не депрессивных, а агрессогенных психотропных препаратов, поскольку увеличение частоты агрессивных контактов выше некоторого предела может спровоцировать распад семейных групп. Именно такая цель была поставлена в другом полевом эксперименте с монгольскими песчанками.
В качестве психотропных средств, повышающих агрессивность животных, применяют, в частности, антидепрессанты-ингибиторы моноаминоксидазы (МАО). При угнетении активности МАО задерживается расщепление катехоламинов, которые накапливаются в определенных центрах головного мозга. Антидепрессивный (агрессогенный) эффект ингибиторов МАО объясняется повышением содержания серотонина или норадреналина в соответствующих структурах центральной нервной системы (Ильюченок, 1972). Для проведения эксперимента на монгольских песчанках с применением агрессогенных психотропных средств в полевых условиях был использован антидепрессант парнат.
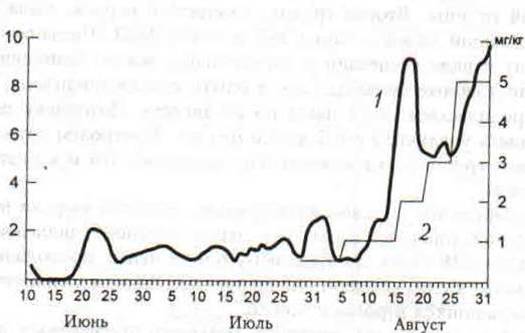
Рис.41 Динамика агрессивных взаимодействий монгольских песчанок в эксперименте с парнатом. Левая шкала — число агрессивных контактов за 1 час наблюдений (1), правая шкала -доза препарата (2).

Рас 42. Взаимное расположение семейно-групповых участков монгольских песчанок дo начала (А) и по окончании (Б) эксперимента с антидепрессантом парнатом. 1 – границы территории самца №3 и самки №4, 2- граница территории самца №5, 3- границы территории самки №22. Пунктиром показана осевая разметка площадки наблюдений. Масштаб 10 м.
При планировании эксперимента предполагалось подобрать семейную группу, состоящую из нескольких особей, и попытаться путем снижения порога агрессивной реакции песчанок спровоцировать распад семейной группы и изменить структуру территориальных отношений в сложившейся внутрипопуляционной группировке. Для проведения эксперимента было выбрано кошарное поселение монгольских песчанок в составе двух семей. Одна из них, включающая пару взрослых особей (самец №3 и самка №64) и шесть молодых зверьков в возрасте 1.5 мес., была выбрана в качестве основной подопытной группы. Вторая группа, соседняя с первой, была представлена семейной парой — самец №5 и самка №22. Песчанки получали препарат в виде суспензии в растительном масле, нанесенной на небольшие хлебные брикеты (как в опыте с галоперидолом). Эксперимент продолжался с 28 июля по 31 августа. Дозировку парната по ходу опыта увеличили с 0.5 до 5.0 мг/кг. Контролем служила та кошарная группировка монгольских песчанок, что и в опыте с галоперидолом.
К сожалению, в самом начале опыта молодые зверьки погибли неизвестной причине, возможно, из-за сильного ливневого дождя, прошедшего 28 июля. Эксперимент, тем не менее, продолжили, чтобы установить, каким образом антидепрессант парнат повлияет на поведение оставшихся взрослых особей.
График на рис. 41 отражает динамику агрессивных взаимодествий в экспериментальной группировке, из которого видно, что ходу эксперимента неуклонно возрастало число агрессивных контактов между двумя семейными парами. Территориальные конфликты песчанок приобрели напряженный и затянутый характер. К завершению эксперимента зверьки №5 и №22 вытеснили своих соседей части занимаемой ими территории и расширили собственную территорию (рис. 42).
Несомненный интерес представляют изменения в поведении самцов и самок в семейных парах, произошедшие по ходу эксперимента, которые можно продемонстрировать на примере самца №3 и самки №4. До получения препарата частота контактов между этими зверьками была относительно низкой, причем все взаимодействия были нейтральными или миролюбивыми (рис. 43, А). С первых дней эксперимента частота контактов многократно возросла (различия достоверны, Манна-Уитни U-тест, р<0.001), взаимодействия зверьков приобрели напряженный характер, в репертуаре поведения семейной пары появились ранее отсутствовавшие элементы ритуализованной агрессии, в основном боковые стойки (рис. 43, Б).
Поскольку в ходе эксперимента резко увеличилась частота территориальных конфликтов с соседней семейной парой, это не могло отразиться на поведении самца №3 и самки №4, агрессивность которых повысилась как под влиянием препарата, так и в результате территориальной экспансии особей-соседей. Приближение партнера в этой ситуации провоцировало демонстрацию боковой стойки, которая, как известно, используется песчанками в качестве средства подавления возможных агрессивных намерений со стороны партнера. Следует отметить, что оба зверька, несмотря на возросшую напряженность отношений, тем не менее, избегали открыто агрессивных взаимодействий (преследований, схваток и т.п.).
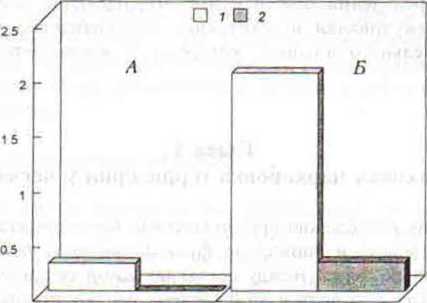
Рис. 43 Средняя частота нейтральных и миролюбивых (1) и ритуализованных агонистических взаимодействий (2) в семейной паре монгольских песчанок перед началом (А) и в течение эксперимента с парнатом (Б). По вертикали — количество контактов за I час наблюдений.
Результат, полученный в ходе описанного эксперимента, в целом можно охарактеризовать положительно, хотя конечная цель опыта не была достигнута. Под влиянием агрессогенного психотропного средства у песчанок резко возросла частота территориальных конфликтов, вследствие чего одна семейная пара оккупировала часть территории другой семейной пары. Кроме того, увеличилась частота конфликтных взаимодействий между самцами и самками в семейных парах, что, однако, не привело к их распаду. Основную положительную роль в этом сыграла ритуализованная агрессия, которую с полным правом можно рассматривать в качестве стабилизирующего этологического механизма, блокирующего деструктивный эффект открытой агрессии. Результаты проведенного эксперимента демонстрируют ведущее значение открытой агрессии как этологического механизма монополизации пространства у территориальных видов и указывают на стабилизирующую, умиротворяющую функцию ритуализованного агонистического поведения не только в территориальных конфликтах, но и в системе внутригрупповых взаимоотношений.
[Обратите внимание, что эндогенное увеличение агрессивности не разрушило существующих социальных связей – ни между партнёрами, ни между социальными группами. При увеличении напряжённости отношений оба зверька ещё больше избегали открытых проявлений агрессии (преследования, схватки и пр.), в их репертуаре появляются полностью отсутствовавшие элементы ритуализованной агрессии, в основном боковые стойки. Во-вторых, частота миролюбивых взаимодействий в паре возросла пропорционально увеличению контактов агрессивной окраски, так что соотношений тех и других не претерпело существенных изменений.
Аналогичное увеличение ритуализованной агрессии с вытеснением неритуализованной сопровождало рост напряжённости территориальных отношений между парами. Перераспределив пространство, обе пары в общем сохранили структуру группировки, поскольку обе продолжали обитать на прежней территории. Отсюда вывод, что ритуализация взаимодействий при увеличении напряжённости контактов означает переход от обмена воздействиями между индивидами к процессам коммуникации внутри сообщества, от прямых действий к обмену демонстрациями. Последний подчиняется определённым (видоспецифическим) «правилам», управляется видовыми механизмами «социального контроля», почему и способен «снять» возникшее напряжение, реализовать «эластичность» видовой социальной организации.]
В заключение остается напомнить, что кроме описанных выше двух форм индивидуализации пространства, связанных с теми или иными проявлениями открытой и ритуализованной агрессии, у песчанок имеется и третья форма индивидуализации участков обитания в виде разнообразных способов запаховой маркировки территории. Этот механизм или, вернее, целая группа специфических популяционных механизмов, поддерживающих необходимую степень диффузности в распределении особей и предотвращающих возможность их чрезмерного скучивания и обострения внутривидовой конкуренции, требует тщательного анализа, которому и посвящается следующая глава».
«Этологические механизмы популяционного гомеостаза у песчанок (Mammalia, Rodentia). М.: Наука, 2000.С.103-113.

Показательно, что адекватность поведения системного целого происходит в условиях заторможенности и неадекватности поведения самих особей (опыты с галоперидолом) и поэтому демонстрирует эмерджентные свойства системы, нежели возможности индивидуальной активности.
В своё время yurvorспрашивал меня, будут ли агрессивными отношения в обществе, составленном из неагрессивных индивидов. Вышеописанные опыты показывают – безусловно да, если агрессивность находится «в типе» соответствующей социальной организации.
Обе серии исследований в области поведенческой фармакологии показали высокую устойчивость социальных (и популяционных) систем как способности воспроизводить специфические паттерны структуры, несмотря на постоянные изменения состава и собственных характеристик особей, образующих эти структуры отношений в разных частях видового ареала, несмотря на разные типы возмущений экологической и «напряжений» социальной среды. Последние особо интересны, так как отражают внутреннюю динамику социальных отношений в группировках, постоянные попытки индивидов «перестроить» существующую систему взаимосвязей в свою пользу.
Наиболее значимы следующие три типа «напряжений» внутри социальной системы: а) быстрые эндогенные изменения уровня агрессивности или иной мотивации (сексуальности, осторожности), определённый уровень которой поддерживает существующие отношения индивидов, — в том случае если изменения уровня не есть следствие побед и поражений индивидов в социальных контактах; б) неожиданное освобождение одной или более «вакансий» резидентов в сообществе, особенно «наверху» сложившейся «лестницы иерархии». Оно провоцирует остальных на попытки передела территорий, повышения собственного социального статуса при том, что для устойчивости сообщества как целого необходимо «перераспределить» часть аспектов социальной роли доминанта между оставшимися особями; в) изъятие всех или большей части особей, с полным разрушением пространственно-этологической структуры группировок. «Эластичность» социальной системы предполагает, что в процессе восстановления в новой сети группировок реализуется тот же «тип» отношений между отдельными особями в сообществе, между группировкой и внедряющимися вселенцами, наконец, между разными группировками, соединёнными потоком переселенцев нерезидентов. Даже если новая сеть поселений копирует прежнюю ни по плотности, ни по конкретному местоположению группировок (например, когда восстанавливается в достаточно однородном биотопе), она сохраняет тот же самый тип взаимного размещения друг относительно друга локальных поселений разной плотности и устойчивости.

Монгольская песчанка Meriones unguisculatus
Собственно опыты с нейролептиками, антидепрессантами и т.д. «психофармакологией» показали высокую «эластичность» системы в ситуациях а)-в) у «социальных» видов песчанок, и способность особей этих видов «собирать» видоспецифический паттерн социальных отношений даже в случаях, когда поведение особей неадекватно из-за сильного лекарственного воздействия или других причин.
Далее в книге В.М.Громов (2000) описывается реакция системы на появление «вакансий», изученная в экспериментах Д.П.Орленева (1990) в естественных поселениях монгольских песчанок. Наблюдения проводили на двух гектарных площадках, заложенных в центре поселения песчанок на залежи с высокой численностью. На одной экспериментальной площадке из 6 семейных групп удалили всех взрослых особей высокого ранга. Через двое суток после изъятия здесь начали отмечаться немеченые песчанки. Однако лишь некоторые из прибывших нерезидентов смогли задержаться и закрепиться; большинство не задерживалось на площадке более 2 дней подряд. Из 30 прибывших особей 9 были взрослыми перезимовавшими самцами, 6 вскоре покинули площадку, а оставшиеся 3 внедрились в те семейные группы, откуда изъяли взрослых, и быстро получили там доминантный статус. «Вакансии» взрослых самок всегда замещались молодыми самками из тех же групп: не отмечено случаев внедрения самок со стороны. Большинство вакансий взрослых особей заняли сеголетки из старших выводков. Как показывает вольерный эксперимент В.С.Громова (2000), в случае изъятия взрослых особей выводок оставшихся молодых зверьков не распадается и продолжает занимать прежнюю территорию. Заняв «вакансии» взрослых особей, молодые самцы демонстрируют свойственный им уровень охраны/маркировки территории, хотя обычно он существенно ниже.
Следовательно, устойчивость социальной организации к внутренним «напряжениям» обеспечивается существованием поведенческих компенсаторных механизмов. Они обеспечивают структурную и функциональную целостность группировки за счёт адекватных (и, главное, своевременных) перестроек структуры сообщества, в том числе перераспределение социальных ролей, формирование нового «социального ядра» группы, появление новых лидеров в охране и маркировке группового пространства.
Особенно у «социальных» видов, отличающихся от близких «несоциальных» в первую очередь тем, что реагируют компенсаторными изменениями психофизиологии и поведения не только на уплотнение, но и на разрежение населения вида, чтобы во втором случае можно было «собрать» специфическую структуру отношений между особями хотя бы в меньшем пространстве. А «несоциальным видам» это совершенно не важно или почти не важно. Они неспособны к такой «самосборке» системы.
Вместо резюме: популяционная биология после истребления у «социальных» и «несоциальных» видов грызунов
В понимании популяционной динамики конкурируют две концепции – стохастизм и регуляционизм. Первая предполагает, что популяция ведёт себя чисто мальтузиански – никаких внутренних регуляторов у неё нет, особи размножаются насколько позволяют ресурсы до тех пор, пока численность не превысит предел, за которым её снижают голод, хищники и болезни. Нижний предел численности, соответственно, определяется ресурсами.
Регуляционизм предполагает, что у популяции есть внутренние механизмы гомеостаза, которые заставляют её «тормозить» рост численности задолго до достижения верхнего предела и не позволяют численности «упасть» вследствие неблагоприятных средовых флуктуаций, болезней или истребительных мероприятий. Есть и промежуточный взгляд, согласно которому при колебаниях численности нижний предел определяется средовыми факторами, верхний – регуляторными способностями популяционной системы; все три изложены в книге Г.С.Розенберга, Д.П.Мозгового и Д.Б.Гелашвили «Экология. Элементы теоретических конструкций современной экологии» (2000, с.185-186).
Как это часто бывает, все три концепции оказываются применимыми, но в разных случаях или, точнее, для разных видов. Как я уже рассказывал, близкие виды птиц, млекопитающих, рептилий (близкие в экологическом плане; имеют сильно пересекающиеся, в ряде случаев вполне тождественные экологические ниши) могут быть ранжированы в ряд по «степени социальности».
На одном полюсе – «несоциальные» виды вроде норвежского лемминга, сибирского бурундука Eutamias sibiricus или полуденной песчанки, на другом наиболее «социальные» вроде монгольской песчанки с полёвкой Брандта Lasiopodomys brandti, есть и промежуточные (американские виды бурундуков, Громов, 2008). Из хорошо изученных песчанок к первому полюсу также близки тамарисковая песчанка Meriones tamariscinus (краснохвостая M.lybicus по степени «социальности» промежуточна). Из моих пёстрых дятлов – большой пёстрый и средний дятлы (сирийский посередине), белоспинный и малый пёстрый дятлы и т.д. Так или иначе, ранжирование экологически близких видов «по степени социальности» можно произвести в подавляющем большинстве случаев.
Именно тогда, когда экологические различия минимальны и данные виды как бы собираются нарушить правило Гаузе (особенно в периоды ухудшения погодных условий и другие моменты понижения доступности корма – видовые экониши конвергируют, и их перекрытие не падает, а растёт), они максимально различны между собой по «степени социальности». Это имеет существенное адаптивное значение, поскольку «социальные» и «несоциальные» виды существенно различаются между собой в характере регуляторной реакции на переуплотнение и разрежение в популяционной системе, что задаёт разные темы и формы восстановления численности.
У «несоциальных» видов она описывается на основании концепции стохастицизма, у «социальных» — регуляционизма, у промежуточных – смешанным вкладом первой и второй в соответствии в тем, какие контексты и типы взаимодействий в популяционной системе вида характеризуются высокой, какие – низкой социальностью.
Конечно, неверно заключение, что «несоциальные» виды всегда живут одиночно и мало общаются друг с другом; когда к тому вынуждает среда, они образуют плотные скопления и, столкнувшись друг с другом, взаимодействуют более чем интенсивно и агрессивно. См. ниже про кошарные поселения полуденных песчанок. Однако самые интенсивные взаимодействия здесь отнюдь не ведут ни к становлению устойчивых – асимметричных связей между двумя данными индивидами, ни к поддержанию характерной структуры связей, охватывающей всю группировку. Дело в том, что у них последействие агрессивных (и миролюбивых) контактов слабое, почти нулевое и не канализует поведение выигравшего и проигравшего в следующем контакте.
Увеличение интенсивности взаимодействий, особенно в агрессивном контексте здесь скорее дезорганизует отношения животных, и поэтому они стараются от таких взаимодействий уйти: дифференцируют время и место активности так, чтобы не встречаться друг с другом, и даже в плотном поселении вспыхнувшие было контакты быстро угасают.
Во взаимодействиях «несоциальных» видов много неритуализированной агрессии и много физических травм, вплоть до скальпирования самок, которые, что интересно, легко заживают и переносятся достаточно легко, например, не препятствуют рождению здоровых детёнышей (Гольцман и др, 1994). «Психологическое» же воздействие – специфический стресс, связанный с борьбой, ритуализированными демонстрациями, принимаемыми в этой борьбе и их релизерным воздействием на противника, которое и стрессирует, у «асоциальных» видов совсем незначительно.
У социальных же видов всё точно наоборот. Физическое воздействие на партнёра даже в агрессивных взаимодействиях невелико, в отличие от «психологического давления», которое существенно, тяжело переносится. Поэтому у соответствующих видов более развиты (и лучше дифференцированы) демонстрации ритуализированной угрозы, позволяющие «снимать» соответствующее «напряжение». Далее, ритуализированные демонстрации у «более социальных» видов выделены и «оформлены» лучше, чем у «несоциальных», они исполняются стереотипней и чётче дискретизируются во время взаимодействия и т.д. Поэтому «социальные» виды лучше способны выдерживать длительные и напряжённые конфликты, они же устанавливают более устойчивые и более однозначные связи с сородичами по сравнению с близкими «несоциальными» видами, особенно если устойчивость / однозначность связей относить к «количеству» / интенсивности агрессии или иного поведения, которым эта связь поддерживается и устанавливается (Попов, 1986).
Именно в силу этого демонстрации социальных видов – более эффективное средство коммуникации: в сообществе, основанном на агрессивном доминировании, обмен ими устанавливает более чёткую дифференциацию ролей, более устойчивую и с бόльшим последействием, чем обмен гомологичными демонстрациями у «несоциальных» видов. Функционирование однотипных демонстраций в коммуникативном процессе «социальных» видов к тому же и более помехоустойчиво: здесь возможно адекватное разрешение конфликтов и установление устойчивой асимметрии отношений особей в более напряжённых взаимодействиях, чем у видов «несоциальных» (Попов, Чабовский, 1995; Попов, Вощанова, 1996).
При переходе к более «социальным» видам также увеличивается сложность комбинирования ритуализованных демонстраций в структурированные последовательности действий во время стычек, возрастает средняя длина и устойчивость таких секвенций ритуализованной агрессии во время стычек. Устойчивость и направленность развёртывания поведенческих секвенций у участников взаимодействия начинает коррелировать со структурированностью последних, с выделенностью демонстраций и чёткостью переходов от одних демонстраций к другим. У «несоциальных» видов те же параметры коррелируют исключительно с уровнем мотивации особей (Попов, 1986; Гольцман и др., 1994; Попов, Чабовский, 1995).
Так, если агрессивное и брачное поведение вида слабо ритуализованы, больший уровень тестостерона обеспечивает больший риск для партнёра, поскольку интенсивность использования ритуализированных демонстраций во взаимодействиях положительно коррелирует с использованием открытой агрессии или неритуализированных попыток спаривания. Именно такой результат был получен для «несоциальных» видов песчанок рода Meriones (Попов, 2005)
Напротив, если степень ритуализации поведения высока, а соответствующие демонстрации сложны, разнообразны и хорошо «выделены» из канвы недемонстративной активности, больший уровень тестостерона должен обеспечивать меньший риск для партнёров животного. Здесь интенсивность использования ритуализированных демонстраций отрицательно коррелирует с использованием прямых действий, выражающих те же самые побуждения индивида, что естественным образом уменьшает риск травматических последствий. Это обнаружено у «социальных» видов песчанок, таких как монгольская Meriones unguisculatus (Попов, 2005).
С точки зрения популяционной динамики «социальные» виды птиц и млекопитающих отличаются от экологически близких «асоциальных» тем, что механизмы популяционной саморегуляции у них запускаются не только при переуплотнении, но и при разрежении популяции, особенно в случаях катастрофического падения численности от истребления, эпизоотий или бескормицы. Далее, у «асоциальных» видов регуляторный механизм, запускающийся при переуплотнении, в общем неспецифический: развивается стресс, вызывающий неизбирательную гибель зверьков.
У «социальных видов» не так: и гибель, связанная с переуплотнением, селективна, и воздействие стресса достаточно специфично – не приводя к существенной гибели зверьков, оно обеспечивает формирование более сложных систем иерархических отношений между особями в популяции. Соответственно, часть особей, которая при низкой степени социальности гибнет, здесь сохраняется на низкоранговых «социальных позициях» в семейных группах и других объединения особей, структурированных системой агрессивного доминирования, которые удерживают территорию (Мошкин, 2000).
В случае экстремального разрежения популяции – скажем, при истреблении связанном с дератизацией, между «социальными» и «асоциальными» видами наблюдаются того же рода отличия. Первые резко сокращают агрессию, увеличивают исследовательскую деятельность и обследование территории, и стремятся хотя бы на небольшом участке пространства воспроизвести те видоспецифические особенности социальной организации, которые были до истребления. Понятное дело, что когда численность вида была существенно выше, группировки с соответствующей социальной структурой могли покрывать куда большую территорию, но после дератизации единичные выжившие и рассеянные в пространстве зверьки должны найти друг друга, собраться вместе и воспроизвести прежнюю структуру отношений хотя бы в одной или в нескольких группировках.
Асоциальные виды после истребления почти не меняют ни поведения, ни конфигурации индивидуальных участков, несмотря на существенное падение плотности. Выжившие зверьки ведут себя так, как будто ничего не изменилось, а пустующие пространства если и заселяются, то животными со стороны.
Далее цитирую книгу С.А.Шиловой «Популяционная экология как основа контроля численности мелких млекопитающих» (М.: Наука, 1993), как раз посвящённую восстановлению популяций разных видов грызунов после дератизации:
«В Туве этот вид [монгольская песчанка – В.К.] образует поселения с высокой численностью (до 250 зверьков на 1 га). Песчанки жили на залежных землях, старых пахотах, возле кашар и на небольших понижениях рельефа в щебнистой степи, где мягкий грунт позволял устраивать норы. Поскольку монгольские песчанки участвуют в хранении возбудителя чумы, в районе наших работ проводились экспериментальные обработки территорий приманками, содержащими фосфид цинка, монофторин, глифтор и другие препараты. Площадь каждого опытного участка составила 80-100 га. Одномоментная гибель особей после обработок была не ниже 86-95%.
Маркировка территории (создание холмиков, помеченных мочой и помётом) относится к одной из распространённых форм территориального поведения монгольских песчанок и в нормальной обстановке постоянно регистрировалась у зверьков, находящихся под нашими наблюдениями (Громов, Попов, 1979). На обработанных участках, где после истребления сохранилось 5-15% выживших зверьков с первых же дней маркировочная активность возросла по сравнению с контролем более чем в 50 раз (Орленев, 1987, Кайдун, личн. сообщ.).
Важно, что на контрольной территории при существовании стабильных групп зверьки метили в основном те норы, которые находились в пределах их территории («свои» норы). В то же время, после обработки метки располагались преимущественно у нежилых норок (Р≥0,999; Орленев, 1987, Кайдун, личн.сообщ.).
Интересные соображения по поводу значения интенсивной маркировочной активности выживших зверьков высказывает Д.П.Орленев (1987). По мнению автора, помимо создания информационного поля для интеграции разрозненных особей метки могут выполнять другие разнообразные функции. Поскольку моча и секреторные выделения желёз несут информацию о половой принадлежности особи и её физиологическом состоянии, запаховые метки песчанок могут играть роль «праймер-феромонов» и стимулировать репродуктивную активность. Специфику размножения зверьков в условиях искусственной депрессии мы описываем ниже.
Показатели численности и маркировочной активности монгольских песчанок на опытном и контрольном участках после обработки родентицидами (по: Орленев, 1987)
| Время учёта | Через 2 сут. после обработки | Через 2 нед. после обработки | ||
| опыт | Контроль | опыт | контроль | |
| Площадь учёта м2 | 1720 | 3120 | 3000 | 2840 |
| Число норовых отверстий на 1 га | 1169 | 1314 | 1254 | 1384 |
| Число жилых норовых отверстий % | 0 | 71 | 19 | 82 |
| Число меток на 1 га | 0 | 83 | 410 | 106 |
| Число меток на 1 зверька | 0 | 0,8 | 50 | 1,0 |
Поскольку объединение особей, оставшихся после истребления, сочетается с массовым притоком мигрантов, очевидно, что восстановление нормальных популяционных структур может произойти только в случае падения общего уровня агрессии. Так и происходит в действительности. Среди примеров, подтверждающих снижение агрессивности в популяциях с низкой численностью, мы можем привести данные, полученные нашей экспедицией в Калмыкии (Шилов А., 1983). При изучении популяционной структуры поселений малого суслика было показано, что уровень игровых и агрессивных контактов молодняка зависит от плотности: при высокой плотности (до 40 зверьков на 1 га) частота контактов составляла в среднем 6,2 в час, а уровень игровых и агрессивных контактов, соответственно (в % %) – 20,7 и 63,3. В этот же период на участках с низкой численностью (менее 20 сусликов на 1 га) показатель игровых контактов значительно возрос (42,2), а агрессивных упал (34,2%).
Как мы уже говорили, монгольская песчанка ведёт семейно-групповой образ жизни, у этого вида социальные контакты определяют поддержание целостности внутрипопуляционных группировок (Громов, Попов, 1979). При сравнении поведенческих актов полуденной и монгольской песчанок нам удалось показать, что прямые социальные контакты регистрируются у монгольских песчанок в 6-7 раз чаще, чем у полуденных (Шилова, Громов, 1983).
В то же время полуденным песчанкам свойственна слабая социальная активность Изучение популяционных структур этого вида и форм социальных взаимодействий показало, что социальные контакты в этом случае не являются ведущим фактором в поддержании внутрипопуляционных группировок (Попов, 1981, Чабовский, 1986).
В соответствие с перечисленными особенностями социальных взаимоотношений у монгольских и полуденных песчанок процессы восстановления нарушенных популяционных структур происходили по-разному.
При обработке экспериментальных площадей, заселённых монгольской песчанкой (участки по 30-150 га, см. выше), проводилось индивидуальное мечение зверьков как на обработанных территориях, так и на их границах, вне пределов действия родентицидов. После гибели основной части аборигенного населения у песчанок, населяющих границы обработок и лишившихся соседей, значительно возросла исследовательская активность и преобладали поведенческие формы «поиска контактов». В результате стабильные группы, населяющие границы обработок, распались и входившие в них зверьки заселили освобождённую территорию. Здесь, совместно с выжившими аборигенами, они сразу же образовали «жилые островки» с высокой численностью (до 100-150 зверьков) и упорядоченными структурами.
Во всех сериях экспериментов поселения монгольских песчанок после обработки восстанавливались очень быстро, причём зверьки образовывали устойчивые стабильные группы. По справедливому предположению Д.П.Орленева (1981), формирование резерватов монгольской песчанки происходит не стохастически, а под контролем поведенческого регуляторного механизма.
Восстановительные процессы в группировках полуденных песчанок происходили по-другому. В Калмыкии под нашим наблюдением находились поселения полуденных песчанок двух типов: 1) расположенные в естественных участках с мягкими грунтами среди глинистой полупустыни и 2) антропогенные поселения, приуроченные к заброшенным кашарам, где рытьё нор возможно на старых валах антропогенного происхождения (Попов и др., 1989). В первом случае пространственная организация поселений носила характер, описанный для этого вида из других точек ареала (Попов, 1981; Верёвкин, 1981); индивидуальные участки взрослых самок были строго изолированы; в антропогенных поселениях они интенсивно перекрывались (Шилова и др., 1983). В обоих типах поселений среди взрослых зверьков значительно преобладало оседлое население. Песчанки проявляли исключительно высокий уровень привязанности к определённой территории.
Естественные поселения полуденных песчанок, площадью от 1 до 3 га, были обработаны родентицидами после индивидуального мечения всего населения, что давало возможность детально изучить механизмы выживания и формирования остаточных популяций. Хотя обработанная площадь была очень небольшой, процесс её заселения происходил крайне медленно, несмотря на обилие зверьков на окружающих участках. Песчанки, оставшиеся в живых, не пытались расширить свои индивидуальные участки и вступить в контакт с другими зверьками. Полное восстановление плотности произошло здесь лишь через 3 месяца. Территориальная структура восстановленного поселения соответствовала таковой до опыта: самки жили изолированно, а индивидуальные участки самцов перекрывались.
Аналогичная серия экспериментов была проведена в антропогенных поселениях. Аборигенное население было полностью изъято с трёх изолированных участков (земляных валов), площадь которых составляла 8-10 тыс. кв.м. Картина восстановления численности полностью повторила описанную выше для естественных поселений; оседлые зверьки, живущие на соседних участках, не предприняли никаких попыток немедленно расширить свою территорию. Поселения, освобождённые от аборигенов, осенью были заселены вновь только после выхода молодняка последующего сезона, то есть пустовали почти 10 мес. Зверьки не проявили тенденций к концентрации. Характерно, что при восстановлении численности сохранился принцип использования территории в антропогенных поселениях: самки, вселившиеся на валы, жили совместно, несмотря на обилие вокруг старых пустующих нор. Пространственная структура поселения полностью соответствовала таковой до опыта….
…Поскольку объединение особей, оставшихся после истребления, сочетается с массовым притоком мигрантов, очевидно, что восстановление нормальных популяционных структур может произойти только в случае падения общего уровня агрессии. Так и происходит в действительности. Среди примеров, подтверждающих снижение агрессивности в популяциях с низкой численностью, мы можем привести данные, полученные нашей экспедицией в Калмыкии (Шилов А., 1983). При изучении популяционной структуры поселений малого суслика было показано, что уровень игровых и агрессивных контактов молодняка зависит от плотности: при высокой плотности (до 40 зверьков на 1 га) частота контактов составляла в среднем 6,2 в час, а уровень игровых и агрессивных контактов, соответственно (в % %) – 20,7 и 63,3. В этот же период на участках с низкой численностью (менее 20 сусликов на 1 га) показатель игровых контактов значительно возрос (42,2), а агрессивных упал (34,2%)….
…Как мы уже говорили, монгольская песчанка ведёт семейно-групповой образ жизни, у этого вида социальные контакты определяют поддержание целостности внутрипопуляционных группировок (Громов, Попов, 1979). При сравнении поведенческих актов полуденной и монгольской песчанок нам удалось показать, что прямые социальные контакты регистрируются у монгольских песчанок в 6-7 раз чаще, чем у полуденных (Шилова, Громов, 1983).
В то же время полуденным песчанкам свойственна слабая социальная активность Изучение популяционных структур этого вида и форм социальных взаимодействий показало, что социальные контакты в этом случае не являются ведущим фактором в поддержании внутрипопуляционных группировок (Попов, 1981, Чабовский, 1986).
В соответствие с перечисленными особенностями социальных взаимоотношений у монгольских и полуденных песчанок процессы восстановления нарушенных популяционных структур происходили по-разному.
При обработке экспериментальных площадей, заселённых монгольской песчанкой (участки по 30-150 га, см. выше), проводилось индивидуальное мечение зверьков как на обработанных территориях, так и на их границах, вне пределов действия родентицидов. После гибели основной части аборигенного населения у песчанок, населяющих границы обработок и лишившихся соседей, значительно возросла исследовательская активность и преобладали поведенческие формы «поиска контактов». В результате стабильные группы, населяющие границы обработок, распались и входившие в них зверьки заселили освобождённую территорию. Здесь, совместно с выжившими аборигенами, они сразу же образовали «жилые островки» с высокой численностью (до 100-150 зверьков) и упорядоченными структурами.
Во всех сериях экспериментов поселения монгольских песчанок после обработки восстанавливались очень быстро, причём зверьки образовывали устойчивые стабильные группы. По справедливому предположению Д.П.Орленева (1981), формирование резерватов монгольской песчанки происходит не стохастически, а под контролем поведенческого регуляторного механизма.
Восстановительные процессы в группировках полуденных песчанок происходили по-другому. В Калмыкии под нашим наблюдением находились поселения полуденных песчанок двух типов: 1) расположенные в естественных участках с мягкими грунтами среди глинистой полупустыни и 2) антропогенные поселения, приуроченные к заброшенным кашарам, где рытьё нор возможно на старых валах антропогенного происхождения (Попов и др., 1989). В первом случае пространственная организация поселений носила характер, описанный для этого вида из других точек ареала (Попов, 1981; Верёвкин, 1981); индивидуальные участки взрослых самок были строго изолированы; в антропогенных поселениях они интенсивно перекрывались (Шилова и др., 1983). В обоих типах поселений среди взрослых зверьков значительно преобладало оседлое население. Песчанки проявляли исключительно высокий уровень привязанности к определённой территории.
Естественные поселения полуденных песчанок, площадью от 1 до 3 га, были обработаны родентицидами после индивидуального мечения всего населения, что давало возможность детально изучить механизмы выживания и формирования остаточных популяций. Хотя обработанная площадь была очень небольшой, процесс её заселения происходил крайне медленно, несмотря на обилие зверьков на окружающих участках. Песчанки, оставшиеся в живых, не пытались расширить свои индивидуальные участки и вступить в контакт с другими зверьками. Полное восстановление плотности произошло здесь лишь через 3 месяца. Территориальная структура восстановленного поселения соответствовала таковой до опыта: самки жили изолированно, а индивидуальные участки самцов перекрывались.
Аналогичная серия экспериментов была проведена в антропогенных поселениях. Аборигенное население было полностью изъято с трёх изолированных участков (земляных валов), площадь которых составляла 8-10 тыс. кв.м. Картина восстановления численности полностью повторила описанную выше для естественных поселений; оседлые зверьки, живущие на соседних участках, не предприняли никаких попыток немедленно расширить свою территорию. Поселения, освобождённые от аборигенов, осенью были заселены вновь только после выхода молодняка последующего сезона, то есть пустовали почти 10 мес. Зверьки не проявили тенденций к концентрации. Характерно, что при восстановлении численности сохранился принцип использования территории в антропогенных поселениях: самки, вселившиеся на валы, жили совместно, несмотря на обилие вокруг старых пустующих нор. Пространственная структура поселения полностью соответствовала таковой до опыта» (Шилова, ibid. С.110-18).
Источники
Гольцман М.Е., Попов С.В., Чабовский А.В. и др., 1994. Синдром социальности: сравнительное исследование поведения песчанок// Журн. общей биологии. Т.55, Вып.1, С.49-69.
Громов В.С., 2005. Ритуализованное агонистическое поведение грызунов// Успехи современной биологии. Т.125. №5. С.509-519.
Громов В.С., 2008. Пространственно-этологическая структура популяций грызунов. М.: Товарищество научных изданий КМК. 581 с.
Мошкин М.П., 2000. Популяционная физиология и этология млекопитающих и птиц в проектах РФФИ // Вестник РФФИ. № 3. С.5-17
Попов С.В., 1986. Социальные взаимодействия и социальная структура — возможные связи характеристик поведения и популяционных структур. Методы исследований в экологии и этологии. Пущино, С.121-140.
Попов С.В., 2005. Механизмы формирования устойчивых внутригрупповых отношений у песчанок (Rodentia, Cricetinae) // Бюлл. МОИП. Отд. Биол. Т.110. №4. С.98-100.
Попов С.В., Вощанова И.П., 1996. Динамика агрессивных взаимодействий у «социальных» и «несоциальных» видов песчанок (Rodentia, Gerbillidae). Зоол. ж. Т.75, №1, С.125-132.
Попов С.В., Чабовский А.В., 1995. Плотность популяции, социальная среда и поведение: возможные взаимосвязи// Экология популяций: структура и динамика. (Мат. совещ. Ч.I.). М., РосСХА. С.183-202.









{kind=link}