

Добытое мясо диких животных (так называемый bushmeat) в тропиках часто коптят, чтобы сохранить
Человек как суперхищник
Из-за длительных коэволюционных отношений (обычно они ограничивают уровень эксплуатации жертв, особенно взрослых особей) природных пресс хищников редко ведёт к столь экстремальным последствиям редко навязывают нечеловеческие хищники.
Между тем, вопрос о том, можно ли считать нынешнюю и будущую эксплуатацию опромышляемых видов устойчивой, вызывает жаркие споры, особенно в рыболовстве [первую — точно нет, второе возможно, если от слов перейти к делу, преодолев сопротивление гг.бизнесменов, в первую очень окоротив им руки, т. е. исключив существующее сейчас непропорционально большое влияние на демократию — СМИ, выборы, суд и правительство, сравняв его с таковым прочих граждан, для чего средства социального влияния — СМИ, помещения для собраний и митингов, пр. площадки общественной активности должны быть в общественной же собственности. Отчасти это возможно, если общины охотников ради сохранения добычи смогут защитить тропические леса и другие биомы тропической области от немедленной и главной угрозы — сведения ради пастбищ для скота и других с/хугодий. Однако долговременные последствия этого успеха будут негативны и в случае роста численности таких общин, и в случае продажи добытого на сторону: оба фактора интенсифицируют промысел, и рано или поздно популяции жертв будут подорваны. Прим.публикатора].
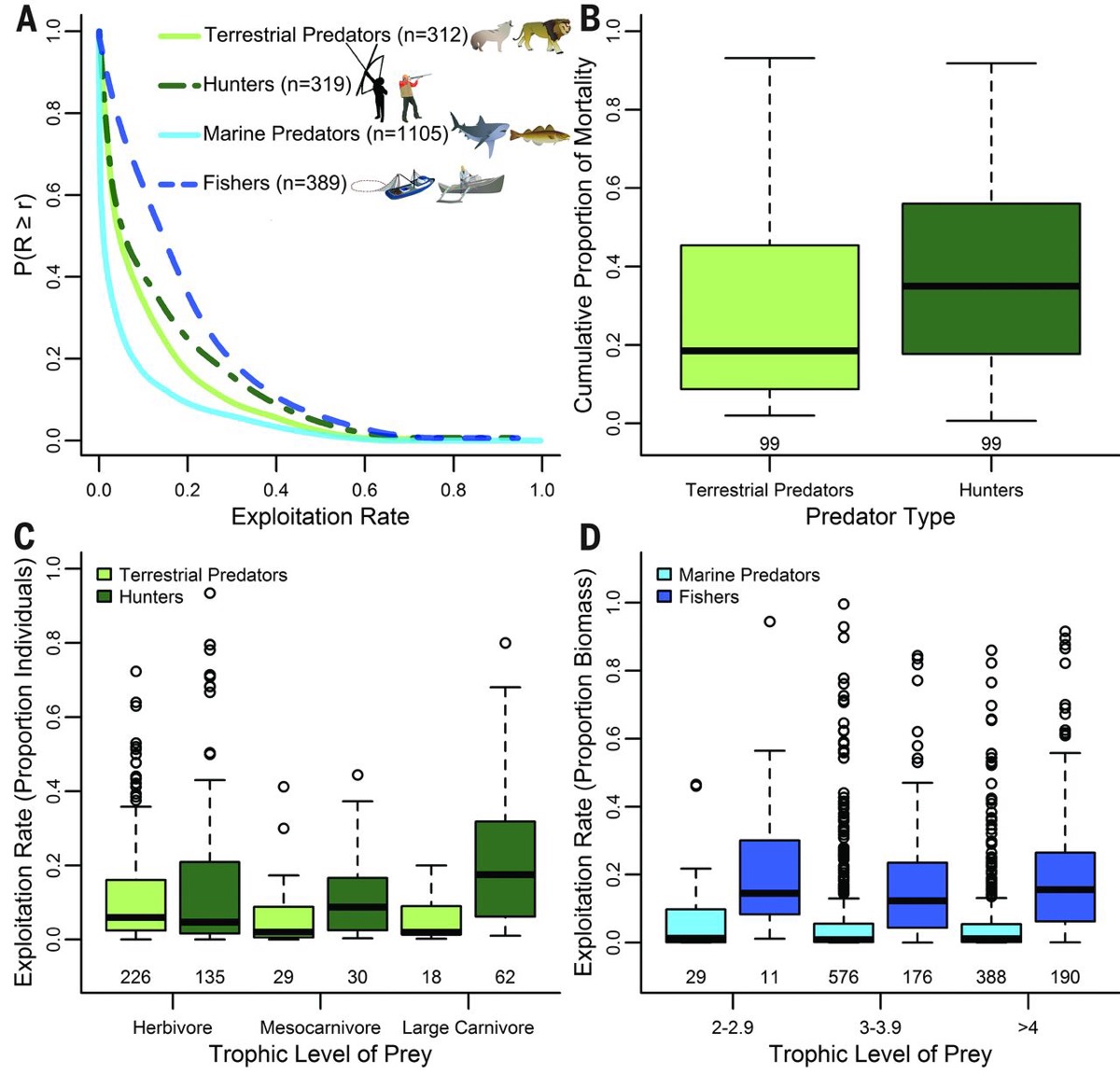
Рис. 1. Модели эксплуатации запасов добычи (взрослые особи) людьми и хищниками в природе. (A) Добавочные кумулятивные функции распределения, показывающие вероятность того, что нагрузка хищников или промысла R будет больше или равна заданной годовой конечной скорости эксплуатации r, рассчитанной по численности особей в популяциях (наземные млекопитающие) или их биомассе (морские рыбы). (Б) Доля ежегодной смертности в популяциях добычи, вызванной охотниками vs всеми прочими потребляющими их наземными хищниками. (C и D) Интенсивность эксплуатации видов-жертв промысла и природных хищников на разных трофических уровнях в (C) сухопутных и (D) морских сообществах. Усы диаграммы — расстояние от верхних и нижних квартилей до наибольших и наименьших не выпадающих значений.
Сходным образом, независимо полученные попарные сравнения уровней смертности жертв от изъятия охотниками (0.35) были в 1.9 раз выше, чем от изъятия их же всей совокупностью местных хищников (0.19) (парный текст Вилкоксона V = 1605, P = 0.004) (Рис. 1B).
[Т.е. в Африке крупные хищники сохранились лучше, и в плане разнообразия видов, и в плане численности каждого из них, и больше мест, где они могут свободно охотиться, не конкурируя с современными охотниками, хорошо вооружёнными и экипированными].
[Хотя травоядные пострадали от людей сильнее хищников, сокращение местообитаний и вытеснение домашним скотом тут перевесило эффект охоты. При прочих равных больше страдают самые крупные и самые мелкие виды зверей. Прим.публикатора ]
[тут я не согласен, это вариант европейского мифа о благородном дикаре, в вариации «природа знает лучше». Хищники тоже не умеют; точнее, устойчиво они используют жертву или нет, зависит от множества привходящих, прежде всего средовых, обстоятельств, которые они не могут контролировать, скажем, возможность переключиться на другие корма, как у лис и песцов в немышиные годы, и численность этих заместительных жертв, связанная, скажем, с урожаем зелёной массы, ягод или беспозвоночных — если речь идёт о гнездах и птенцах наземногнедящихся видов птиц — зависящих уже совсем от других факторов. В отличие от диких видов, человек это может просчитывать и такими вещами управлять. Прим.публикатора]
Про влияние промысла на популяции рыб и прочих биоресурсов
Ю.П.Алтухов
«При изучении природных рыбных популяций (стад) обнаруживается ярко выраженная гетерогенность, их дифференцированность на более мелкие, генетически отличающиеся субпопуляции. Это, например, было показано около 40 лет назад для американского морского окуня Sebastes mentella, чьи стада обитают на больших глубинах в районах п-ва Лабрадор и о-ва Ньюфаундленд (Алтухов, 2003).

Sebastes mentella
Обнаружение такой системной организации популяций имеет принципиальное практическое значение. Очевидно, что если мы хотим осуществлять рациональный промысел, имея дело с системой, то должны подходить к ней как к целому с учетом ее внутренней структуры. Вместе с тем рыбаки обычно игнорируют эту организацию стад, вследствие чего происходит разрушение популяционных систем. 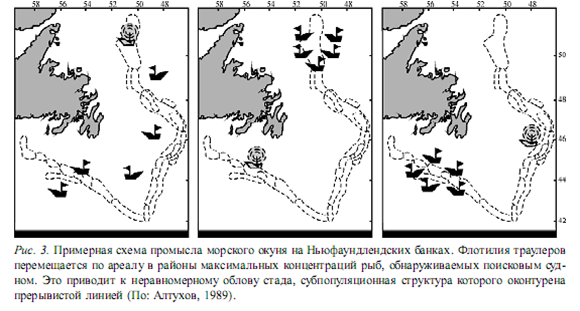 Для иллюстрации сказанного обратимся к типичной картине морского рыбного промысла, стратегия которого, как известно, включает две главные акции — разведку достаточно плотных скоплений рыб поисковым судном и, после их обнаружения, вылов флотилией промысловых судов. Чтобы показать эту картину в динамике, можно изобразить ее в виде серии следующих друг за другом «кадров» (рисунок), которые демонстрируют вскрытую нами цепь генетически отличающихся субпопуляций морского окуня. Понятно, что при подобном типе промысла, когда суда всякий раз устремляются в тот участок ареала, где скопление рыб характеризуется максимальной плотностью, одни субпопуляции перелавливаются, другие недолавливаются.
Для иллюстрации сказанного обратимся к типичной картине морского рыбного промысла, стратегия которого, как известно, включает две главные акции — разведку достаточно плотных скоплений рыб поисковым судном и, после их обнаружения, вылов флотилией промысловых судов. Чтобы показать эту картину в динамике, можно изобразить ее в виде серии следующих друг за другом «кадров» (рисунок), которые демонстрируют вскрытую нами цепь генетически отличающихся субпопуляций морского окуня. Понятно, что при подобном типе промысла, когда суда всякий раз устремляются в тот участок ареала, где скопление рыб характеризуется максимальной плотностью, одни субпопуляции перелавливаются, другие недолавливаются.
В конечном счете происходит нарушение естественно сложившихся каналов миграционной связи между элементами системы, разрушается генетическая структура популяции. Этого можно было бы избежать, равномерно облавливая стадо как целое, с непременным учетом пространственной субпопуляционной организации.
Столь на первый взгляд абстрактная схема получила прямые доказательства при изучении последствий промыслового воздействия на подразделенные популяции тихоокеанского лосося — нерки, размножающейся в озерах Камчатки. Важная биологическая особенность нерестовых популяций нерки — уникальная картина изменчивости производителей по длине тела: самки характеризуются одновершинным распределением, тогда как для самцов прослеживается четко выраженная двувершинность (Алтухов, 1983). В это время в водоемах обнаруживаются три легко распознаваемые группы рыб: мелкие самцы, крупные самцы и самки, занимающие между двумя группами самцов промежуточное положение по признаку «длина тела».
Мало того, в процессе промысла можно видеть, как со временем в исследуемой популяции существенно возрастает доля мелких половозрелых самцов. Такие, как правило, трехлетние самцы (их местное название на Камчатке — «каюрки», канадцы называют их «джек», американцы — «грилз») лишь с небольшой частотой встречаются в нативных, мало облавливаемых стадах.
Напротив, в популяциях, испытывающих систематическое промысловое воздействие, количество мелких, рано созревающих самцов резко возрастает. Этот процесс в настоящее время в большей или меньшей мере характерен практически для всех популяций тихоокеанских лососей, размножающихся в разнообразных речных системах по обе стороны Северной Пацифики и интенсивно облавливаемых промыслом с начала нашего столетия.
Ярким примером, иллюстрирующим это правило, может служить стадо нерки оз. Дальнего (п-ов Камчатка), биология которого детально изучена начиная с 1930-х гг. благодаря работам Ф.В. Крогиус (цит. по Алтухов, 1989). Если в 1930-х гг . численность нерестовой части дальнеозерского стада составляла около 100 тыс. производителей, а доля каюрок среди половозрелых самцов не превышала 0,2%, то в 1960-1970-е гг. . численность производителей сократилась до 2-5 тыс. и доля каюрок увеличилась до 38 % .
В чем же причина столь драматических изменений? Исследования показали, что главный фактор — селективный морской промысел, из поколения в поколение нарушающий генетическую структуру стад нерки из-за непропорционального изъятия жаберными сетями крупных, более гомозиготных старых самцов. Другие рыбы, идущие на нерест и отличающиеся генетически от крупных самцов, облавливаются промыслом или равномерно (самки), или недолавливаются (мелкие самцы), что и приводит к резкому изменению исторически сложившейся популяционно-генетической структуры стада (Алтухов, 1989).
Дело в том, что в нерестовых стадах нерки существует весьма консервативная система так называемых селективных скрещиваний. При формировании брачных пар на нерестилищах самки отдают предпочтение старым, медленно растущим, более гомогизотным крупным самцам и лишь в маловодные годы и на мелководных нерестилищах, куда крупные самцы не могут проникнуть, репродуктивный успех сопутствует быстрорастущим молодым, более гетерозиготным самцам.
Промысел нарушает естественную систему воспроизводства, и более гетерозиготные (гетерозисные) мелкие самцы во все большей мере передают свои гены последующим поколениям (хотя обычно гетерозисные животные характеризуются увеличенными размерами, у нерки это не так, поскольку рыба размножается только раз в жизни и после нереста производители погибают). Доля крупных рыб в стаде уменьшается, нарушается равновесное соотношение полов, увеличивается скорость полового созревания, сокращается средняя продолжительность жизни и, как следствие, возрастает темп смены поколений.
Одновременно падает численность стада, так как мелкие самки имеют более низкую плодовитость. Таким образом, в условиях снижения воспроизводительной способности стада даже постоянный по интенсивности промысел, вполне совместимый с изначальными продукционными возможностями популяции, приводит к сокращению ее численности в поколениях только из-за непропорционального изъятия рыб определенных генотипов (Алтухов и др., 1997; Altukhov et al., 2000).
Обнаруженные процессы свойственны не только стадам тихоокеанских лососей, но и другим видам рыб — объектам промысла. Во всех до сих пор детально исследованных случаях картина была однотипной – мониторинг промысловых стад вскрывает их измельчание, омоложение, возрастание доли рано созревающих мелких самцов. Поскольку направление отбора оказывается неизменным (в пользу гетерозигот), внутрипопуляционная компонента генного разнообразия возрастает, тогда как межпопуляционная — падает, приводя к снижению локальной генетической дифференциации….
Искусственное воспроизводство. Генетические последствия искусственного воспроизводства лучше всего рассмотреть на примере лососей. Как уже подчеркивалось, их стада — сложноструктурированные популяционные системы, состоящие из множества дискретных субпопуляций. Если мы воспроизводим такие системы искусственно на рыбоводных заводах, то должны осуществлять сбор половых продуктов на всем протяжении нерестового хода, а не ограничиваться использованием лишь части дифференцированного генофонда. Чем более рельефна субпопуляционная структура популяции, тем меньше шансов воссоздать целое по его отдельной части. К сожалению, это обстоятельство на рыбоводных заводах нередко игнорируется и, как следствие, генетическое разнообразие популяций сокращается, что показано, например, для Salmo clarki и S. salar.
Нами выполнен мониторинг трех соседствующих нерестовых популяций горбуши Южного Сахалина — двух нативных (реки Фирсовка и Бахура) и одной искусственно воспроизводимой (р. Найба) (расстояние между реками — несколько десятков километров) (Алтухов и др., 1997). Популяция Найбы поддерживается рыбоводным заводом, благодаря деятельности которого численность местного стада увеличилась, судя по уловам, в несколько раз. Вместе с тем за последние годы в биологической структуре рыбоводного стада произошли изменения: рыба стала заметно крупнее, увеличилась частота самцов, стала сокращаться численность.
Для выяснения механизма процесса, приведшего к такого рода сдвигам, мы проделали следующее:
1) по совокупности аллозимных локусов, идентифицируемых методами электрофореза, сравнили генетические характеристики самцов-производителей, используемых и отбраковываемых в рыбоводном процессе;
2) проследили динамику соотношения полов и длины тела рыб в ряду поколений искусственно воспроизводимой найбинской популяции;
3) сопоставили ее генетические и биологические параметры с аналогичными параметрами двух нативных стад, размножающихся в соседних реках.
Кроме того, для выяснения связей индивидуальной гетерозиготности с биологически важными признаками у самцов, помимо длины тела, исследовали частоту аномалий (искривлений, расщеплений, срастаний) жаберных тычинок как показатель стабильности онтогенеза. По степени выраженности эти нарушения можно подразделить на две группы: слабые (затронута только одна жаберная дуга) и сильные (затронуты две и более дуг); в сравнительном анализе нами принимались во внимание только сильные аномалии.
Динамика длины тела и соотношения полов у найбинской горбуши оценивалась по рыбоводным материалам начиная с 1973 г . за весь период ее регулярного разведения. Аналогичные данные для самовоспроизводящейся популяции горбуши соседней р. Фирсовки собраны в процессе наших собственных работ. Идентификация генотипов осуществлялась по шести аллозимным локусам.
На рыбоводном заводе при искусственном оплодотворении отдельных порций икры, каждую из которых получают от 50 самок, используются молоки 20-30 самцов. Кроме того, сбор половых продуктов, как правило, осуществляется в начале нерестового хода, когда преобладают самцы, и в середине его, когда соотношение полов близко к равновесному . Сравнение самцов, использованных в рыбоводном процессе (n = 300, длина тела 52,6 ± 0,2 см), с отбракованными («контрольная» группа, n = 293, длина тела 47,8 ± 0,1 см) свидетельствует о предпочтении рыбоводами крупных рыб. Для отбираемых самок такой селективности не выявлено. Особенно важен тот факт, что между контрольной группой самцов и самцами, использованными в скрещиваниях, различия наблюдаются также и по уровням аллозимной гетерозиготности, и по частоте рыб с аномалиями жаберных тычинок: крупные самцы, взятые для рыбоводных целей, оказались более гомозиготными по сравнению с контрольной группой (доля гомозиготных рыб 0,443 ± 0,029 и 0,369 ± 0,028 соответственно; различие достоверно при Р < 0,05); у них же выше частота рыб с аномалиями жаберных тычинок (Р < 0,05).
Если предположить, что такой отбор был в той или иной мере систематическим на протяжении более 16 лет искусственного воспроизводства горбуши р. Найбы и, как правило, в воспроизводство не вовлекалась «арьергардная» часть стада, характеризующаяся избытком самок, следовало ожидать вполне определенных сдвигов в биологической структуре популяции: снижения гетерозиготности по аллозимным локусам, увеличения длины тела, нарушения оптимального соотношения полов за счет нарастания доли самцов (так как сбор икры осуществляется главным образом на раннемигрирующих группах рыб, среди которых высока частота самцов).
Из статьи «Динамика генофондов при антропогенных воздействиях»
Рекомендуем прочесть
 Рыба наперечёт
Рыба наперечёт Рыба или мясо? а есть ли у нас выбор?
Рыба или мясо? а есть ли у нас выбор? Экологичное сельское хозяйство: «Рис и рыба: что обеспечивает успешность комплексных рисоводческих агроценозов?»
Экологичное сельское хозяйство: «Рис и рыба: что обеспечивает успешность комплексных рисоводческих агроценозов?» Что было у берегов Камчатки незадолго до экологического ЧП
Что было у берегов Камчатки незадолго до экологического ЧП IPBES: Сеть жизни планеты под угрозой
IPBES: Сеть жизни планеты под угрозой Стресс в раннем онтогенезе как адаптивное явление
Стресс в раннем онтогенезе как адаптивное явление Ещё деконструкция мифов об охране природы
Ещё деконструкция мифов об охране природы «В тисках у холоднокровных»
«В тисках у холоднокровных»
