

Взрослый анубис грумингует молодого, парк Нгоронгоро, Танзания
В продолжение темы воспроизводства социальных инноваций в череде поколений у животных
«Robert M. Sapolsky1,2*, Lisa J. Share1
1 Department of Biological Sciences and Department of Neurology and Neurological Sciences, Stanford University, Stanford, California, United States of America, 2 Institute of Primate Research, National Museums of Kenya Karen, Nairobi, Kenya
Введение
Содержание
- 1 Введение
- 2 Результаты и обсуждение
- 3 Материалы и методы
- 4 References
Одна из целей приматологии — научиться понимать огромную вариабельность в социальном поведении приматов. Раньше учёные исследовали различия между видами, например, тот факт, что образование пар более распространено среди лесных, чем наземных приматов (Crook and Gartlan 1966). Также уделялось внимание географическим различиям в поведении внутри вида (Whiten et al. 1999). Часто такие различия отражают факторы окружающей среды (например, корреляция между количеством осадков и временем питания) или, теоретически, могут отражать генетический дрейф. Однако всё больше свидетельств в пользу того, что повадки, специфические для разных групп, могут также представлять собой «традиции» или «культуры» (мы будем использовать второй термин, в соответствии с почти установившимся среди приматологов консенсусом, что этот термин можно применять не только к людям, а и к другим приматам).
Как и в случае людей, такую «культуру» можно определить как особенности поведения, общие для какой-то популяции, но необязательно для других представителей вида, которые не зависят от генетики или экологических факторов и которые переживают своих изобретателей (Kroeber and Kluckhohn 1966; Cavalli-Sforza 2000; de Waal 2000; de Waal 2001). Определённая таким образом передача культуры существует у человекообразных обезьян (McGrew 1998; Whiten et al. 1999; van Schaik et al. 2003), других обезьян (Kawai 1965; Cambefort 1981; Perry et al. 2003), китообразных (Noad et al. 2000; Rendell and Whitehead 2001), а также у рыб и птиц (Laland and Reader 1999; Laland and Hoppitt 2003). Особо впечатляющий пример — шимпанзе (Pan troglodytes) по всей Африке демонстрируют вариабельность в 39 видах поведения, касающихся использования орудий, груминга и ухаживания (Whiten et al. 1999), и есть сообщения о раскопках орудий шимпанзе возрастом почти тысячу лет (Mercader et al. 2002).
Почти все такие случаи не-человеческой культуры имеют отношение либо к технологии (например, использование шимпанзе «молотков» для колки орехов), либо к добыче пищи, либо к коммуникации. В этой работе мы задокументировали появление уникальной культуры у павианов анубисов (Papio anubis), относящейся к общей структуре и социальной атмосфере стаи. Мы также описываем физиологические корреляты социальной атмосферы стаи, передачу соответсвующих особенностей поведения после своих изобретателей, и возможные механизмы передачи.
Результаты и обсуждение
Обстоятельства, приведшие к возникновению уникальной культуры
В начале 1980-х Лесная стая ночевала на деревьях в 1 км от туристского лагеря. В этот период сильно выросла открытая мусорная яма возле лагеря. Это привлекло соседнюю стаю павианов, Мусорную стаю, которая ночевала возле ямы и питалась почти исключительно там.
К 1982 году многие самцы из Лесной стаи стали ходить на рассвете за пищей к мусорной яме. Эти поедатели отбросов не отличались от тех, кто не питался отбросами, ни по возрасту (данные не приведены), ни по среднему рангу доминирования, но они были более агрессивными (табл. 1); такую агрессивность можно рассматривать как условие для конкуренции за доступ к отбросам с самцами Мусорной стаи. Поедатели отбросов также участвовали в большем числе столкновений за доминирование внутри Лесной стаи, чем те, кто не питался отбросами (заметим, что частота столкновенй за доминирование независима от исхода, и таким образом, от ранга).
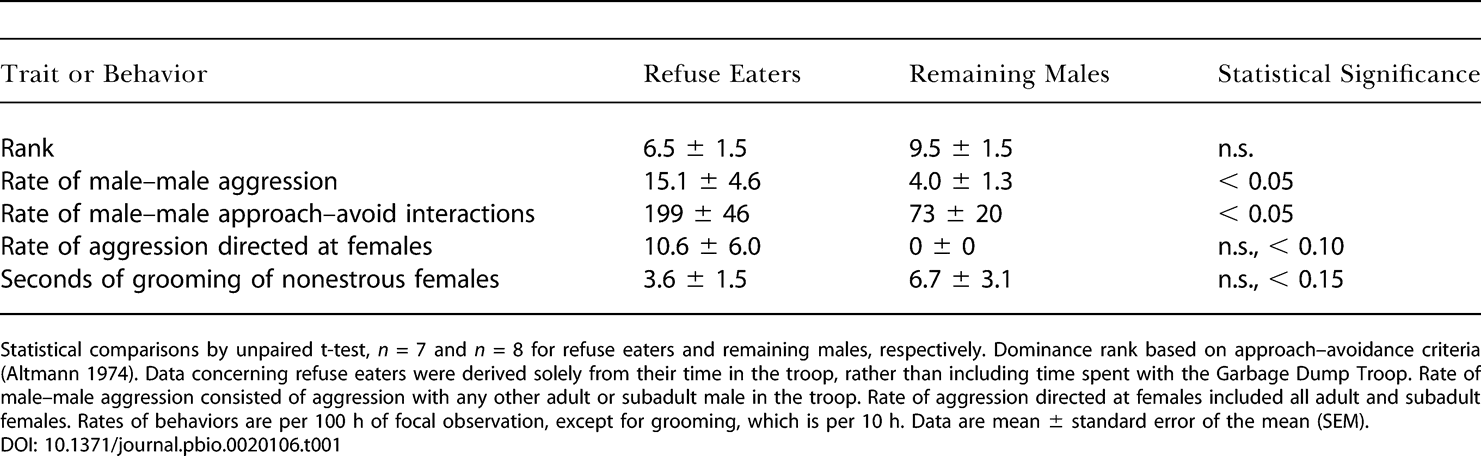
Характеристики самцов Лесной стаи как функция того, конкурировали ли они с Мусорной стаей за отбросы
В 1983 году случилась вспышка бычьего туберкулёза из-за появления на свалке заражённого мяса. С 1983 по 1986 год умерло большинство членов Мусорной стаи, а также все те члены Лесной стаи, которые питались отбросами (46% всех взрослых самцов); никто другой из Лесной стаи не умер (Tarara et al. 1985; Sapolsky and Else 1987).
В результате этих смертей сильно изменился состав Лесной стаи, так что там стало меньше взрослых самцов и больше взрослых самок; это увеличило отношение самки:самцы более чем вдвое (табл. 2). К 1986 году поведение стаи заметно изменилось, так как выжили только менее агрессивные самцы.

Состав стаи перед и после вспышки туберкулёза
Из-за этих событий наблюдения за стаей были прекращены, и до 1993 года велась только перепись. Вместо этого начались исследования стаи Талек, примерно в 50 км отсюда.
В 1993 году неформальные наблюдения за Лесной группой показали, что поведенческие особенности, наблюдаемые к 1986 году, сохранились. Что особенно важно, к 1993 году в стае не оставалось ни одного самца с периода 1983–1986 гг; все тогдашние взрослые самцы присоединились к стае после 1986 г. Таким образом, отличительные особенности поведения, появившиеся в середине 1980-х из-за избирательных смертей, поддерживались следующей когортой взрослых самцов, которые появились в группе. Чтобы запечатлеть этот феномен, в 1993 г. фокусные замеры (focal sampling) среди Лесной стаи возобновились. Данные по Лесной стае 1993–1996 гг (далее Л93–96) сравнивались с двумя другими наборами данных, которые служили контролем: наблюдениями в 1993–1998 гг за стаей Талек (далее Т93–98) и наблюдениями за самой Лесной стаей перед эпидемией (1979, 1980, 1982; далее Л79–82). Эти два контрольных набора данных не отличались существенно один от другого и были объединены, далее Т93–98/Л79–82.
Нетипичные черты поведения самцов Лесной стаи
Взаимодействия доминирования (dominance interactions) между самцами.
Самцы Л93–96 и Т93–98/Л79–82 одинаково часто участвовали во взаимодействиях типа «приближение-избегание» (approach–avoidance dominance interactions) (данные не приведены). Более того, стабильность доминирования (измеренная в процентном отношении тех ВПИ, которые отражали смену доминирования между двумя самцами смежного ранга), не отличалась (16% В± 5% и 20% В± 5% соответственно для Л93–96 и Т93–98/Л79–82, n.s. [статистически не значимо — not significant]). Также не было разницы в средней продолжительности правления самца наивысшего ранга (примерно год).
Несмотря на эти сходства, поведение, касающееся доминирования, отличалось у Л93–96 от двух контрольных случаев такими особенностями, которые, вероятно, способствовали меньшему стрессу для низкоранговых самцов. Первый пример касается ВПИ между самцами с разницей в иерархии более чем два ранга. В подавляющем большинстве таких взаимодействий побеждали более высокоранговые особи. Поскольку кто-то, находящийся на две и более ступеньки ниже в иерархии, редко представляет для самца серьёзную угрозу, взаимодействия между так далеко отстоящими особями обычно представляют собой не настоящую конкуренцию, а притеснение или смещение высокоранговым самцом подчинённого [в смысле, стукнул или согнал с места]. В Т93–98 и Л79–82 примерно 80% ВПИ были между самцами более двух рангов разницы в иерархии. В Л93–96, напротив, такие ВПИ составляли существенно меньший процент (рис. 1A). Вместо этого непропорционально большую часть составляли ВПИ между самцами соседних рангов (причем, как отмечалось, без разницы в стабильности доминирования) (рис. 1B). Более того, высокоранговые самцы в Л93–96 были более «толерантны» к самцам намного низшего ранга, так как было непропорционально много случаев изменения ранга среди самцов с разницей более чем в два ранга (рис. 1C). Таким образом, в Л93–96, при обычном уровне стабильности доминирования, ВПИ просходили в основном среди животных соседних рангов, а к низкоранговым самцам высокоранговые относились более толерантно и меньше их третировали.
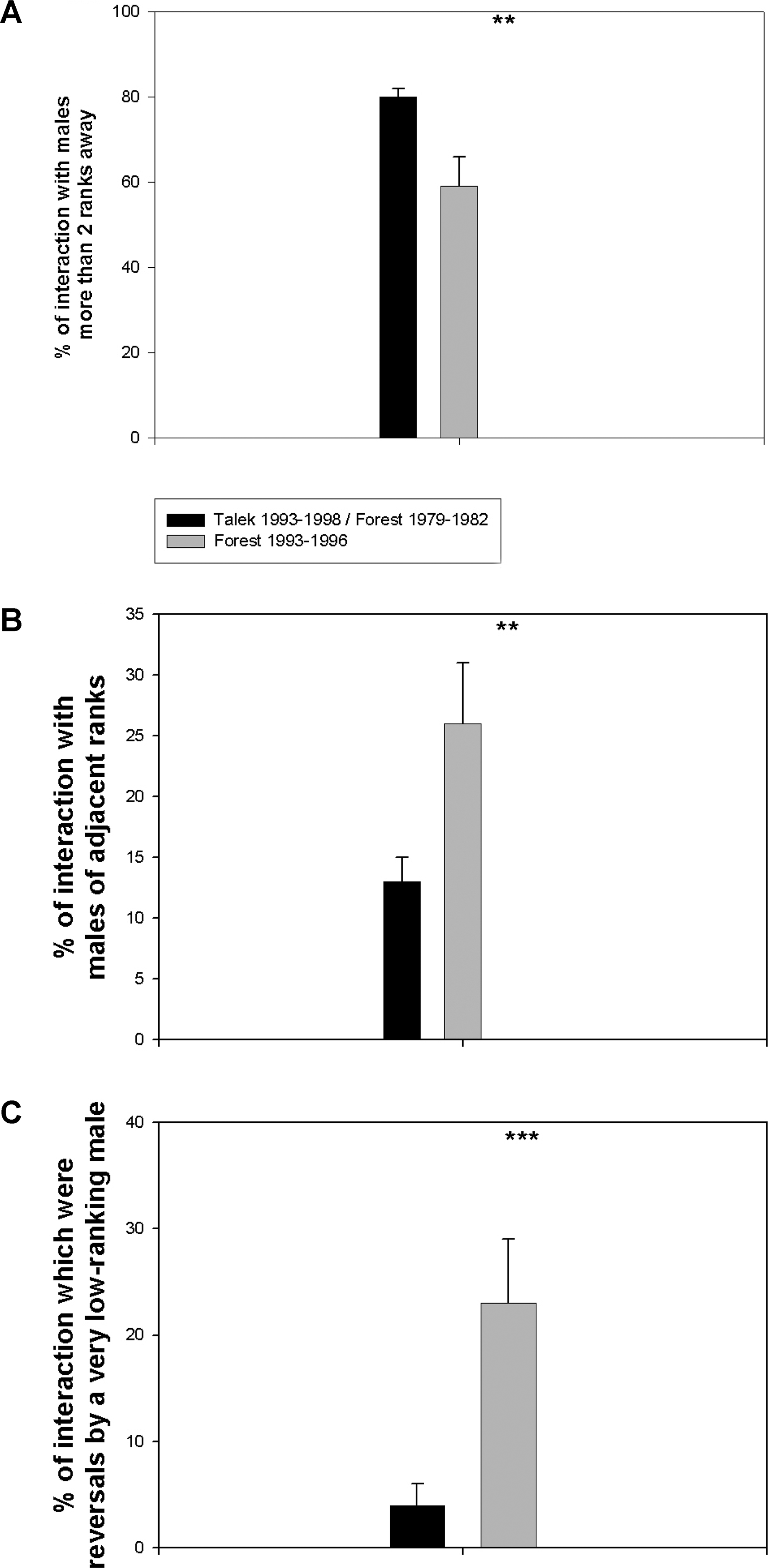
Характер взаимодействий доминирования между самцами
(A) Процент ВПИ между самцами более чем двух рангов разницы.
(B) Процент ВПИ между самцам смежных рангов.
(C) Процент ВПИ, демонстрирующих изменение доминирования внутри пары, где один из самцов стоит ниже более чем на два ранга. Среднее В± SEM, ** и *** означают p < 0.02 и p < 0.01 соответственно, по t-test, считая за точку данных каждого самца/год. Всего участвовали 10 разных самцов из Л93–96, 31 разных самцов из Т93–98, и 19 разных самцов из Л79–82. В принципе, результат (B) мог объясняться меньшим количеством самцов в Л93–96 по сравнению с другими стаями (меньший размер группы не меняет количество соседних особей для конкретного субъекта, но уменьшает количество несоседних). Однако такие же результаты получились, когда размеры стай были искусственно уравнены путём исключения самых высоко- и низкоранговых (данные не приведены).
Агрессия
Характер агрессивных отношений тоже отличался у Л93–96 и Т93–98/Л79–82 в сторону, предположительно, менее стрессового окружения для подчиненных в Л93–96. Во всех стаях было одинаковое общее количество агрессивных взаимодействий (табл. 3). Однако агрессия в Л93–96 чаще, чем в контроле, случалась между близкоранговыми животными (то есть ближе двух ступенек по рангу), а не представляла прямую агрессию высокоранговых самцов в отношении сильно низкоранговых; второй тип взаимодействия для подчинённного является особенно стрессовым из-за своей обычной непредсказуемости. Кроме того, самцы в Л93–96 реже, чем в Т93–98/Л79–82, проявляли прямую агрессию в отношении самок.
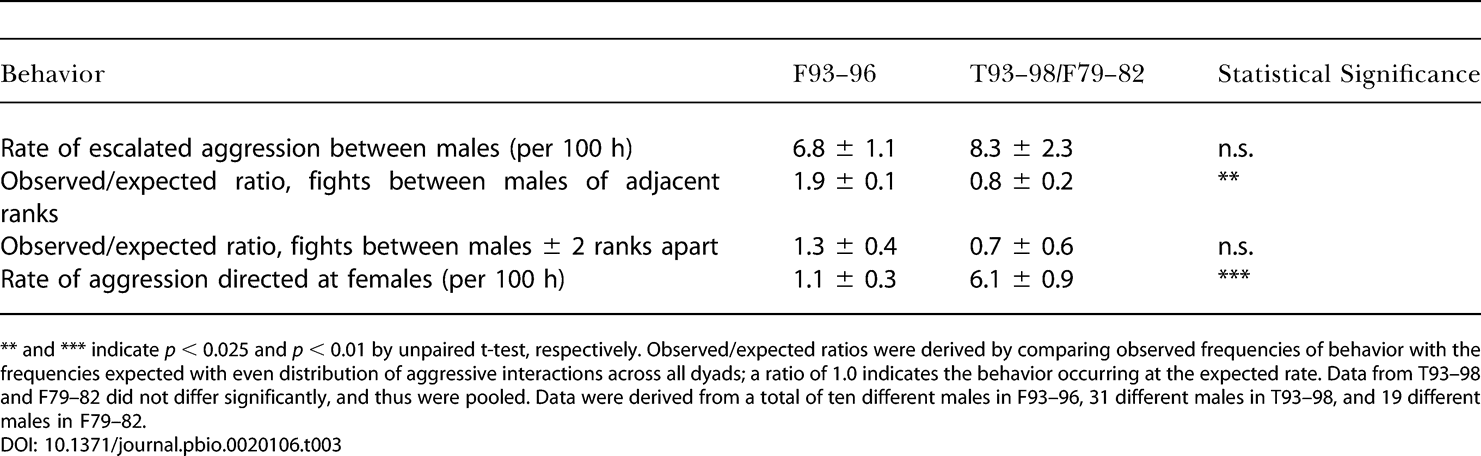
Характер агрессии в Лесной стае 1993–1996 гг по сравнению со стаей Талек 1993–1998 гг и Лесной стаей 1979–1982 гг
Мы рассмотрели данные по примирительному поведению (т.е. актам дружеского поведения между парами, следующими после агрессивных взаимодействий (de Waal and van Roosmalen 1979)) в Л93–96 и Т93–98/Л79–82. Однако никаких проявлений примирения между самцами ни в какой стае мы не обнаружили, что согласуется с прежними сообщениями (Cheney et al. 1995).
Дружеское поведение (affiliative behavior)
Количественных данных по актам дружеского поведения для Л79–82 не было. Но самцы Л93–96 занимались грумингом чаще, чем контрольные самцы в Т93–98 (рис. 2A), эта разница определялась более частым грумингом между самками и самцами. Самцы Л93–96 также чаще находились на близком расстоянии от других животных, чем самцы Т93–98 (рис. 2B). По количеству взрослых самцов-соседов (т.е. до 3 м) самцы во всех стаях не различались, но самцы Л93–96 чаще, чем самцы Т93–98, соседствовали со взрослыми самками, детьми, подростками и молодёжью.
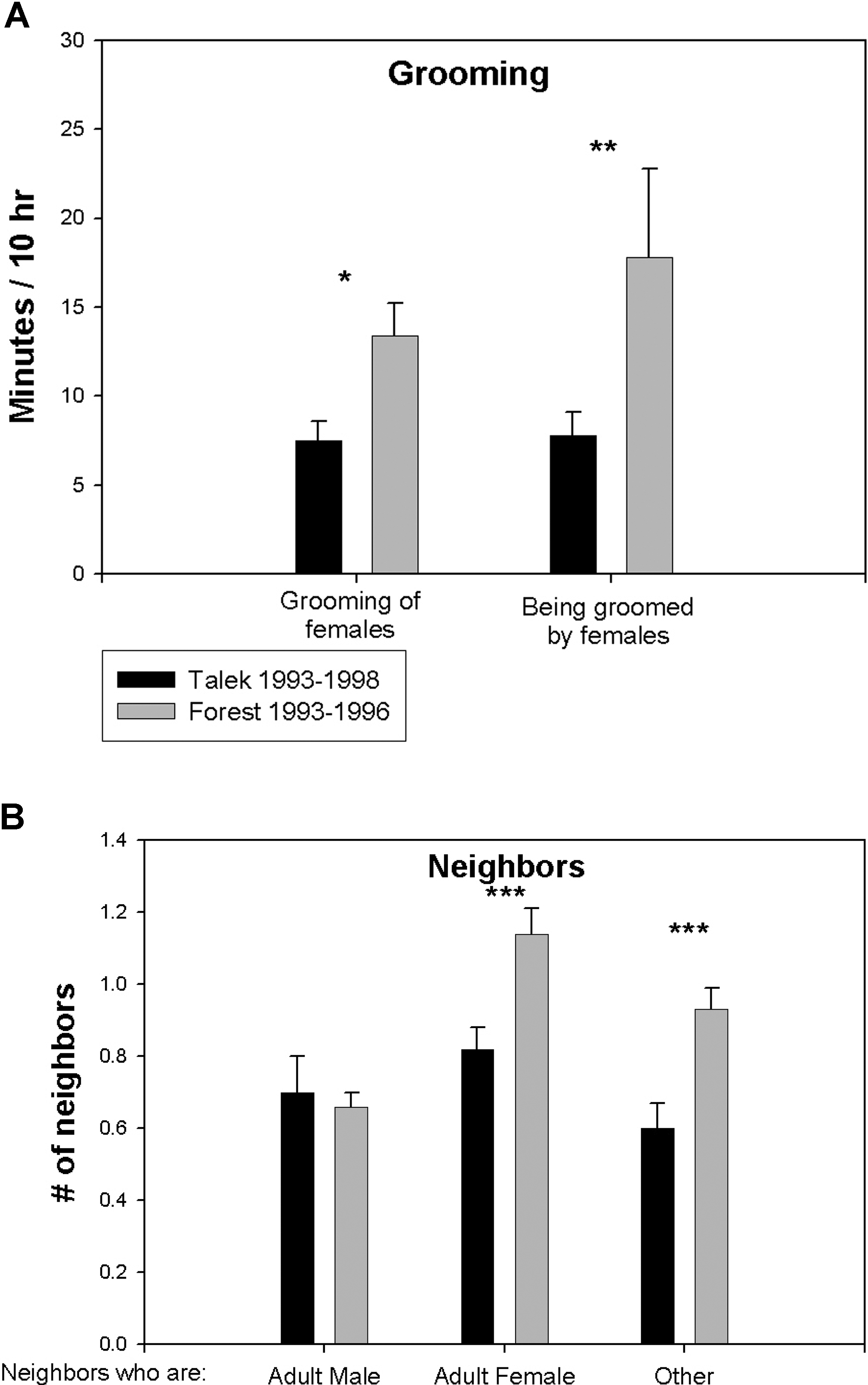
Характер актов дружеского поведения
(A) Количество груминга с участием взрослых самцов в Лесной стае 1993–1996 и стае Талек 1993–1996. Первая пара столбиков представляет среднее время, которое взрослые самцы грумили взрослых самок; вторая пара — среднее время, которое самки грумили самцов.
(B) Сравнение среднего количества соседей (т.е. в окрестностях 3 м) взрослых самцов в двух стаях. Mean В± SEM. *, **, and *** indicate p < 0.05, 0.02, and 0.01, respectively, by unpaired t-test. Всего участвовали 10 разных самцов и 17 разных самок из Л93–96, 31 разных самцов и 21 разная самка из Т93–98, и 19 разных самцов и 23 разные самки из Л79–82.
Сексуальное поведение
Сексуальное поведение в Л93–96 и Т93–98/Л79–82 не различалось. Процент не беременных и не кормящих самок в эструсе в день не отличался (27% В± 7% и 30% В± 4% соответственно, n.s.). Более того, отношение между рангом самца и его репродуктивным успехом не отличалось (R2 корреляции между рангом и репродуктивным успехом: 0.25 В± 0.25 and 0.54 В± 0.10 соответственно, n.s.).
Физиологические корреляты поведенческих особенностей Лесной стаи
Таким образом, самцы Л93–96 относительно часто участвовали в дружеском поведении, а низкоранговые самцы относительно редко подвергались агрессивным и субординационным нападкам от высокоранговых. В стабильной иерархии низкоранговые самцы павианов проявляют физиологические признаки стресса, такие как повышенный базальный уровень глюкокортикоидов (гормонов надпочечной железы, вырабатываемых в ответ на стресс), гипертензию, а также пониженный уровень холестерина липопротеинов высокой плотности (high density lipoprotein cholesterol), факторов роста и лимфоцитов (circulating lymphocytes) (Sapolsky 1993; Sapolsky and Share 1994; Sapolsky and Spencer 1997). Мы проверяли, действительно ли у подчинённых самцов из Л93–96 физиологические признаки стресса, вызванного субординацией, проявлялись меньше, чем в других группах.
Так и оказалось (рис. 3A). В Л79–82, то есть перед вспышкой туберкулёза, подчинённое положение сочеталось с повышенным базальным уровнем глюкокортикоидов, как и у других видов, у которых подчинённое положение влечёт за собой сильные стресс-факторы и низкую частоту разрядки напряжения (coping outlets) (Sapolsky 2001). Хотя глюкокортикоиды помогают пережить сильный стресс, хроническое повышенное их содержание повышает риск предрасположенности к диабету (glucose intolerance), гипертензии, разных видов язвы, а также подавления репродуктивной и иммунной систем (Sapolsky et al. 2000). В противоположность картине в Л79–82, где подчинённое положение было связано с физиологией, характерной для хронического стресса, у подчинённых самцов в Л93–96 базальный уровень глюкокортикоидов повышен не был (для Т93–98 доступной информации не было).
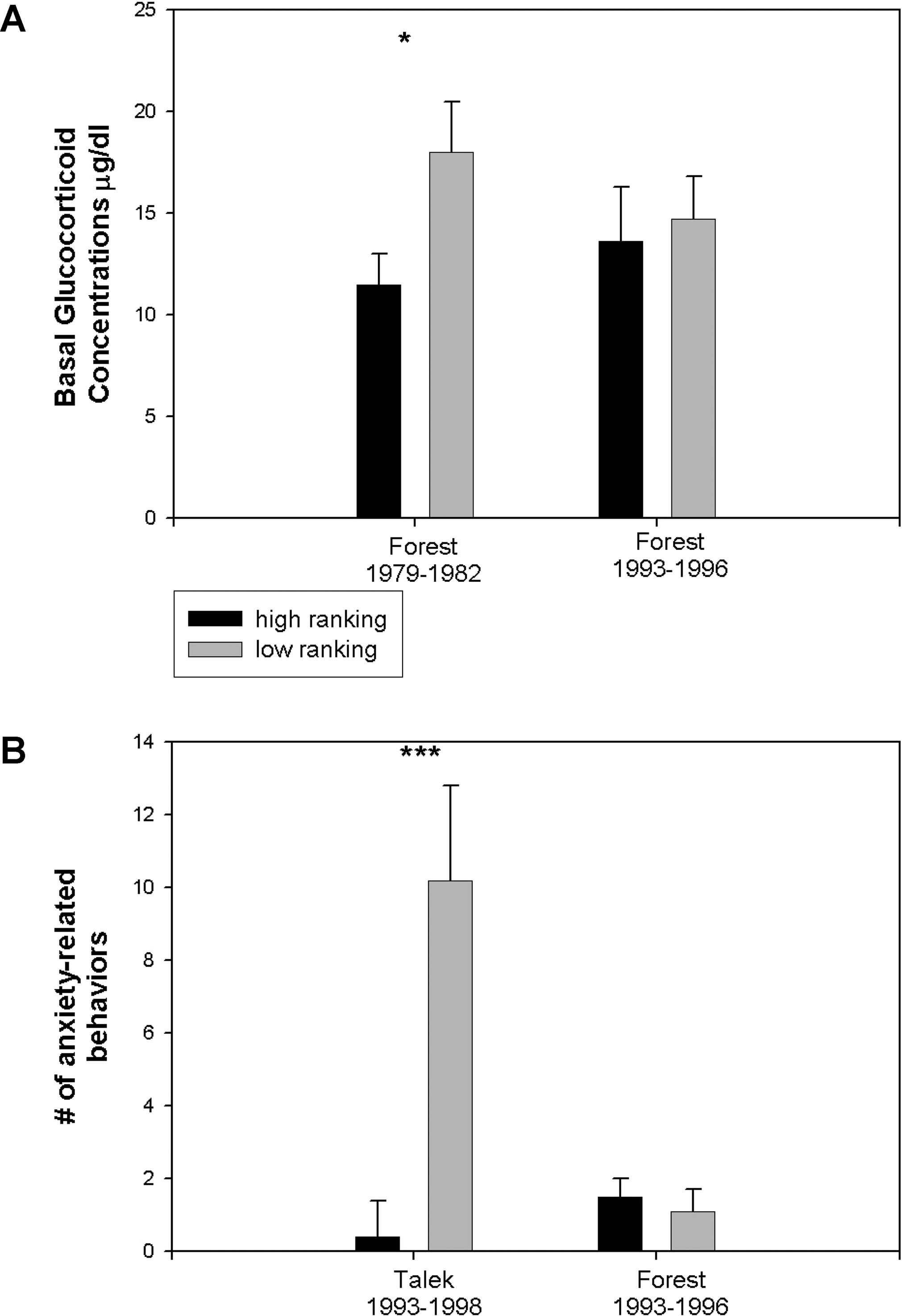
Физиологические профили, относящиеся к стрессу
(A) Базальный уровень глюкокортикоидов (Ојg/100 ml). Самцов поделили на высоко- и низкоранговую половины, по критерию «приближения-избегания». Уровень основного глюкокортикоида — кортизола — измерялся методом радиоиммуноанализа (radioimmunoassay).
(B) Количество связанных с тревожностью поведенческих актов, наблюдаемых в течение 10–20 мин после введения ОІ-carboline-3-carboxylic кислоты (M-156, Research Biochemicals International, Natick, Massachusetts, United States), за вычетом аналогичного количества после введения только наполнителя (dextrin in 1 ml saline); 0.5 г вещества в 1 мл раствора вводилось внутримышечно метательным шприцом (dart syringe) (Pneu-Dart, Inc., Williamsport, Pennsylvania, United States), выпущенным из духового ружья с расстояния 5 м. Среднее В± SEM. * и *** означают p < 0.05 и p < 0.01 соответственно по unpaired t-test. Данные получены в общей сложности от 10 разных самцов из Л93–96, 31 разных самцов из Т93–98 и 18 разных самцов из Л79–82.
У подчинённых самцов Л93–96 отсутствовал ещё один физиологический маркер стресса. Мы искусственно вызывали у самцов состояние тревоги впрыскиванием внутримышечно ОІ-carboline-3-carboxylic кислоты — антагониста бензодиазепинового рецептора, который вызывает у приматов поведенческие и физиологические признаки тревоги (бензодиазепиновые рецепторы связывают транквилизаторы, такие как валиум и либриум, и служат медиатором их успокаивающего эффекта) (Ninan et al. 1982). Самцам вводили вещество в те дни, когда у них не было драк, травм или совокуплений. В качестве контроля в отдельные дни им вводили только наполнитель (в случайном порядке чередования). Потом за самцами наблюдали исследователи, не знавшие о впрыскиваниях.
ОІ-carboline-3-carboxylic кислота не оказывала влияния на поведение высокоранговых самцов ни в Т93–98, ни в Л93–96 (рис. 3B). У низкоранговых самцов в Т93–98 вещество усиливало тревожное поведение, но в Л93–96 — нет (отмеченными признаками тревоги являлись почёсывание, ритмическая тряска головой в бодрствующем состоянии, повторяющееся вытирание носа и жевательные движения у одиноко расположенных самцов (Ninan et al. 1982; Aureli and van Schaik 1991; Castles et al. 1999)).
Таким образом, в более типичных стаях Л79–82 и Т93–98 подчинённое положение имело вполне определённые физиологические корреляты. В противоположность этому, у самцов Л93–96 отсутствовали эти зависящие от ранга отличия.
Потенциальные механизмы, лежащие в основе передачи культуры
Через десять лет после смерти более агрессивных самцов Лесная стая сохраняла свою отличительную социальную обстановку вместе с её физиологическими коррелятами. Особо важно, как отмечалось, что никто из взрослых самцов в Л93–96 не был членом стаи в конце зпидемии туберкулёза. Напротив, эти самцы присоединились позже в молодом возрасте, приняв местный стиль социальных отношений. Некоторые исследователи отмечают, что толерантные и коммуникабельные социальные устои способствуют социальной передаче (e.g., van Schaik et al. 1999), именно такие условия были в Л93–96.
Этот случай социальной передачи напоминает некоторые случаи в прошлом. Например, молодые макаки резус (Macaca mulatta), которых содержали вместе с короткохвостыми макаками (M. artoides), перенимали более мирный стиль вторых (de Waal and Johanowicz 1993). Кроме того, павианы анубисы (Papio anubis) и гамадрилы (P. hamadryas) различаются по социальной структуре, но самки каждого из видов, пересаженные в качестве эксперимента в группу другого вида, перенимают новую социальную структуру в течение нескольких часов (Kummer 1971).
Было предложено несколько моделей для объяснения передачи культуры (Whiten et al. 1999; de Waal 2001; Galef 1990). Для большей ясности полезно сначала рассмотреть их применимость для передачи «технологии», а уже потом применять их для случая передачи социальной атмосферы в Л93–96. Примером первой может служить раскалывание орехов каменными молотками западноафриканскими шимпанзе (Boesch and Boesch 1983; Boesch 2003) — повадка, передающаяся в течение поколений.
В моделях «инструктирования» в использовании шимпанзе орудий молодых активно учат применению молотков. Для случая Л93–96 модели инструктирования предполагали бы, что новоприбывшие самцы участвуют в социально вознаграждающих (например, груминг) или неприятных взаимодействиях (сгон с места или атака) в зависимости от усвоения ими традиций стаи. Ключевым вопросом в этих моделях является, кто именно производит «инструктаж». Точно так же, как и термин «культура» применительно к поведению животных, употребление термина «инструктирование» тоже вызвало споры: некоторые предпочитали концепцию «активной модификации поведения» другими особями, в результате которой происходят изменения. Впечатляющим примером этого является то, что когда молодые воловьи птицы Molothrus ater учатся исполнять свою песню, характерную для данной популяции, они сначала выдают недифференцированный репертуар песен, а самки реагируют на соответствующий диалект демонстрацией просьбы к совокуплению, таким образом положительно подкрепляя и оттачивая нужное поведение (Smith et al. 2000).
В «наблюдательной» модели, применительно к использованию шимпанзе орудий, молодые учатся разбивать орехи, наблюдая и копируя старших. В приложении к Л93–96 прибывшие самцы моделировали бы своё поведение на основании поведения резидентных самцов.
В модели «способствования» в примере шимпанзе нахождение поблизости со взрослым с их молотками увеличивает вероятность того, что молодые будут экпериментировать с молотками и сами изобретут такой навык. Применительно к павианам, поведение самцов Л93–96 было бы неявным начальным (default) состоянием, в котором в отсутствие обычного уровня агрессии со стороны самцов (как в сторону других самцов, так и с сторону самок) самки становятся более дружественными, а при более дружеском поведении самок прибывшие самцы склонны становиться менее агрессивными. Вероятно, то же самое можно выразить, сказав, что начальное состояние могло возникнуть в стае из-за высокого соотношения самки:самцы (так что самцам нужно было меньше конкурировать за доступ с самкам в эструсе).
Наконец, к павианам можно применить модель «самоотбора», в которой определённые типы самцов могли попадать в такую стаю с большей вероятностью, чем другие (заметим, что тот факт, что самцы переходили в стаю из совокупности окружающих стай, исключает возможность ещё одной модели, по которой культура передаётся генетически).
Мы оценивали эти модели, анализируя случаи, когда молодые самцы попадали в стаю в известные дни и наблюдались потом в течение по крайней мере 2–6 месяцев. Таким образом мы искали такие поведенческие признаки среди новоприбывших самцов, которые могли отличаться между Л93–96 (пять таких случаев перехода) и Т93–98/Л79–82 (12 случаев перехода).
Многие взаимодействия с участием новоприбывших самцов нe различались (табл. 4). Все прибывшие самцы в Л93–96, Т93–98 и Л79–82 атаковали и сгоняли самок с мест кормёжки или отдыха одинаково часто. Кроме того, несмотря на отличающуюся структуру доминирования в Л93–96, резидентные самцы в Л93–96, Т93–98 и Л79–82 относились к новоприбывшим самцам одинаково. Проходил одинаковый промежуток времени перед тем, как новоприбывший самец впервые подвергался атаке со стороны резидента, и во всех трёх стаях прибывшие самцы одинаково часто участвовали в агрессивных взаимодействиях и взаимодействиях доминирования (отметим, что из-за того, что в Л93–96 самцов было вдвое меньше, чем в Т93–98 или Л79–82 частота таких взаимодействий для любой конкретной пары самцов отличалась). Мы исследовали случаи, когда резидентные самцы вели себя агрессивно по отношению к прибывшим самцам, определяя, случалось ли такое чаще в течение 20 минут после того, как прибывший проявлял агрессию, чем в другие, случайно выбранные, промежутки времени (de Waal and Yoshihara 1983; de Waal and Johanowicz 1993). Мы не обнаружили никаких свидетельств такой обусловленности поведения (данные не приведены).
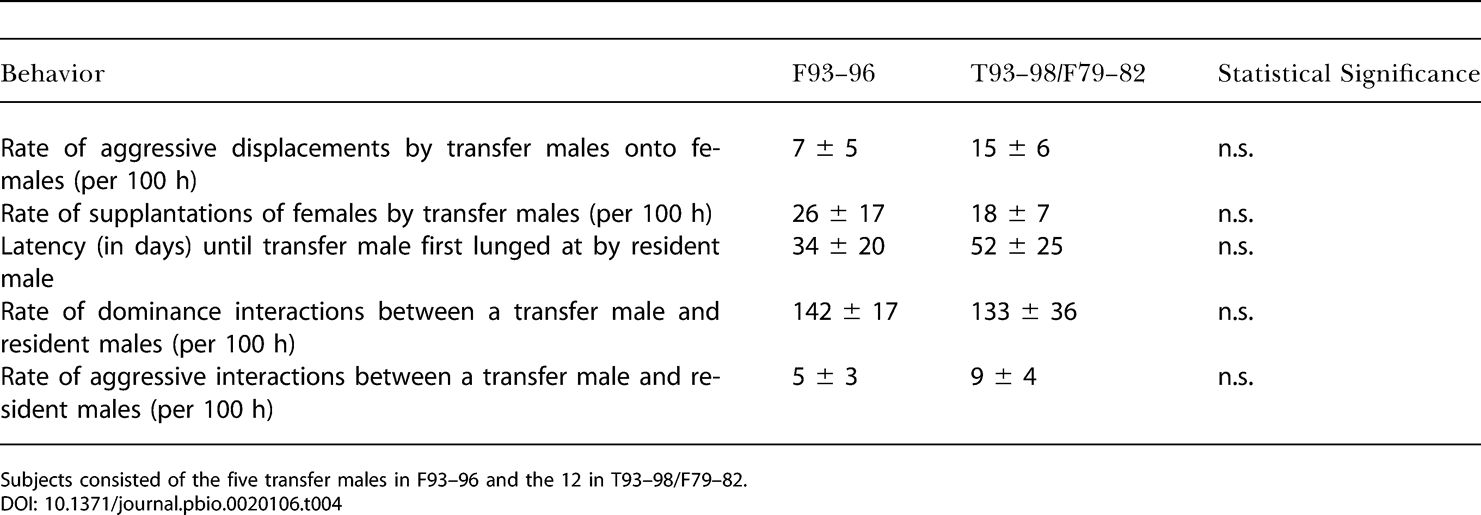
Поведение новоприбывших самцов
Далее мы исследовали дружеские взаимодействия между самками и новоприбывшими самцами и обнаружили впечатляющие отличия между Л93–96 и Т93–98/Л79–82, состоящие в том, что самки Л93–96 вели себя с новоприбывшими самцами так же дружественно, как и с резидентами. У новоприбывших в Л93–96 самцов проходило меньшее время до того, как самки впервые грумили их или предcтавляли себя им (being presented to by a female), чем у прибывших самцов в Т93–98/Л79–82 (рис. 4A). (Эту разницу между Л93–96 и Т93–98, состоящую в более коротком промежутке времени до представления и груминга, нельзя отнести на счёт какой-то одной самки: в первых взаимодействиях с пятью прибывшими самцами в Л93–96 участвовали три разные самки). Более того, прибывшие в Л93–96 сидели ближе к самкам и чаще участвовали в груминге, чем прибывшие в Т93–98/Л79–82 (рис. 4B). Хотя самки в эструсе чаще взаимодействуют с самцами, чем самки, не находящиеся в эструсе (Smuts 1999), процент самок в эструсе во всех стаях не отличался (см. выше). Кроме того, непохоже, чтобы отношение самок Л93–96 к прибывшим самцам зависело от их поведения (de Waal and Yoshihara 1983; de Waal and Johanowicz 1993). Чтобы это проверить, сначала мы рассмотрели случаи, когда резидентные самки вели себя дружески в отношении прибывших самцов, определив, было ли это более вероятным в течение 20 минут после проявления дружеского поведения со стороны прибывшего, чем в другое, случайным образом выбраное, время. Потом мы проверили, уменьшалась ли вероятность дружеского поведения со стороны самки для прибывшего самца в течение 20 минут после проявления агрессии. Мы не обнаружили свидетельств ни того, ни другого (данные не приведены).
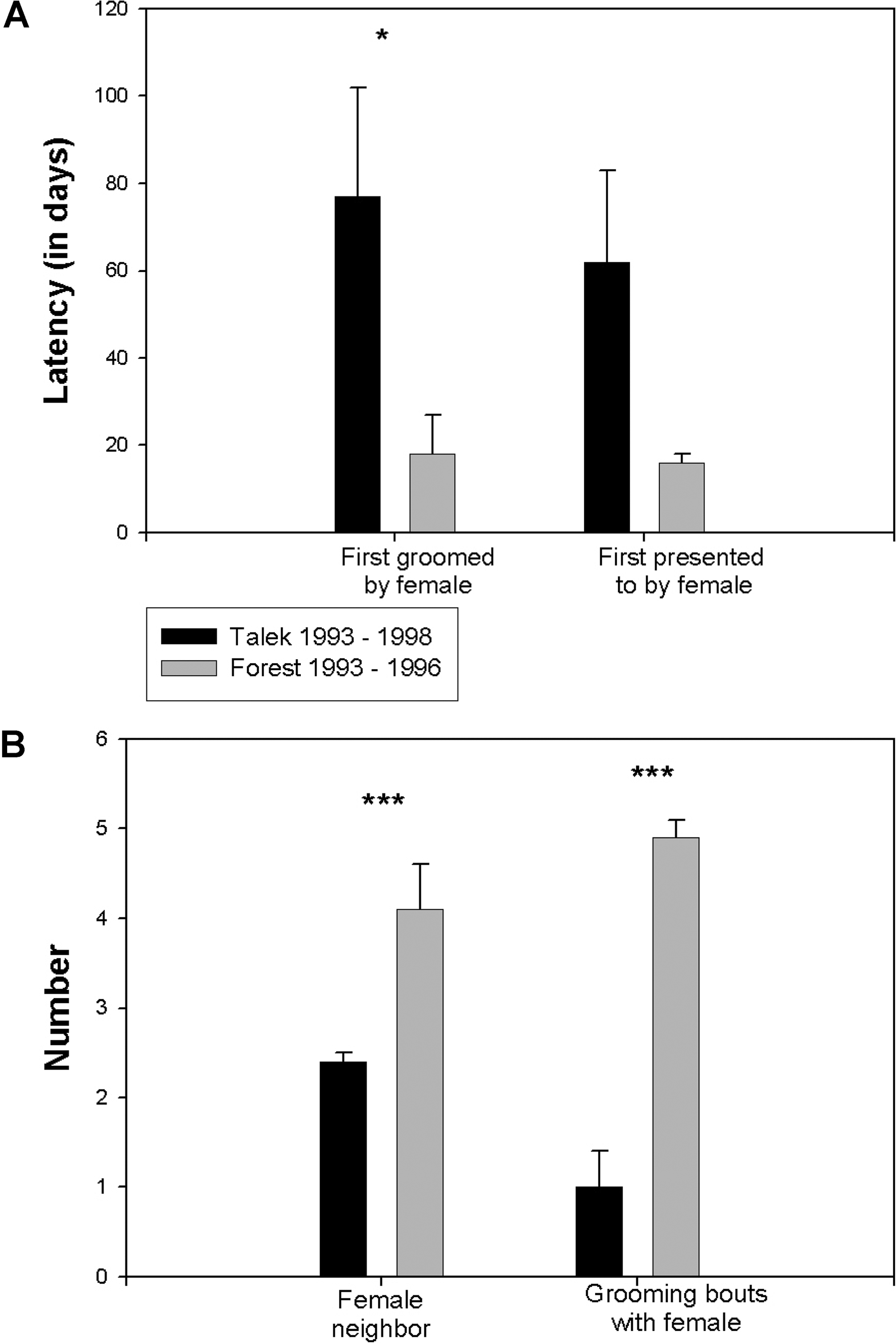
Характер взаимодействий между резидентными самками и прибывшими самцами
(A) Промежуток времени (в днях) до того как новоприбывший самец впервые был представлен самке (справа) или подвергался грумингу (слева).
(B) Среднее количество взрослых самок, находящихся поблизости, за один обзор (т.е. в пределах 3 м; слева) и среднее количество участий в груминге с самками за 100 часов наблюдения (справа) для прибывших самцов. Среднее В± SEM. * и *** означают p < 0.05 и p < 0.01 соответственно, по unpaired t-test. Время до первой презентации себя самкой приближается к достоверности (p < 0.08). Данные получены в общей сложности от 10 разных самцов и 17 разных самок из Л93–96, и 31 разных самцов и 21 разных самок из Т93–98.
Эти данные проливают некоторый свет на механизмы социальной передачи в Л93–96 (хотя их далеко не достаточно для подробного анализа того, в равной ли степени эти механизмы отвечают за передачу всех компонентов культуры Л93–96, а именно меньшей агрессивности самцов, большей дружественности самок и менее жесткой структуры доминирования). То, что отношение резидентов к прибывшим не зависело от их поведения, свидетельствует против модели интструктирования; это согласуется с тем, что вообще существует относительно мало свидетельств «инструктажа» в передаче культуры приматами (кроме людей) (de Waal 2001; исключение см. в Boesch 1991). Одинаковая частота сгона (displacement) прибывшими самцами самок во всех трёх стаях свидетельствует против самоотбора (то есть возможности того, что прибывшие в Л93–96 самцы уже заранее вели себя иначе, чем самцы, попадающие в другие стаи). Это не удивительно. Хотя молодые самцы могут несколько раз переходить из стаи с стаю, прежде чем остаться в какой-то одной (Pusey and Packer 1986), так же как и переходить из неё впоследствии (Sapolsky 1996), мы не видели среди этих животных достаточно свидетельств систематического выбора различных стай, как этого требовала бы модель самоотбора.
Напротив, данные поддерживают модели наблюдения и способствования/начального состояния. Поскольку резидентые самцы вели себя по отношению к прибывшим во всех стаях одинаково, передача путём наблюдения в Л93–96 могла происходить только если это были наблюдения того, как резидентные самцы вели себя по отношению к самкам или друг к другу [с усвоением «типического» в их поведении, а не индивидуальных вариаций]. Некоторые, но не все, исследования говорят в пользу наблюдательных моделей социальной передачи у других приматов (Visalberghi and Fragaszy 1990; Whiten 1998; Boesch 2003; Whiten et al. 2003); о павианах в настоящее время по этом вопросу мало данных. Как было показано, новоприбывшие самцы в Л93–96 относительно часто участвовали в дружеских взаимодействиях с самками. Преобладание самок в Л93–96 — правдоподобное объяснение их необусловленного (unconditial) (или по крайней мере менее обусловленного) роста толерантности и дружественности к самцам (включая новоприбывших), поскольку у самцов стаи было количественно меньше возможностей для проявления агрессии к самкам. (Заметим, что такое искажённое соотношение полов по непонятной причине сохраняется в этой стае и по сей день.) Таким образом, данные по дружескому поведению говорят в пользу модели способствования/начального состояния, только если существует чувствительность в предпочтениях (preferential sensitivity) относительно характера взаимодействий с самками.
В результате анализа возникает предположение, что никакой социальной передачи не было, а особенности Л93–96 — простое следствие возникшего соотношения самок и самцов в 2:1. Чтобы это проверить, мы проанализировали пять других доступных исследований стай павианов с отношением взрослых самок к самцам 2:1 или больше, в которых были количественные данные, сравнимые с нашими данными (Seyfarth 1976, 1978; Strum 1982; Bercovitch 1985; Noe 1994). Ключевой вопрос — больше соответствовали эти предыдущие данные стае Л93–96 или контрольным стаям. Предыдущие данные были больше похожи на контрольные стаи (и существенно от них не отличались) по таким параметрам как относительное время, в течение которого самцы грумили самок (по Seyfarth 1978), относительное время, в течение которого самки грумили самцов (Seyfarth 1978), частота межполовой агрессии (Seyfarth 1976, 1978), структура доминирования самцов (Noe 1995) и характер агрессии между самцами (Strum 1982; Bercovitch 1985). В противоположность этому, никакие количественные параметры не напоминали стаю Л93–96. Это указывает на то, что особенности Л93–96 уникальны и особым образом поддерживаются, а не являются типом социальной структуры, которая автоматически возникает при преобладающем количестве самок. Таким образом, в рамках модели способствования/начального состояния, в этой стае начальное состояние не могло быть вызвано недостатком самцов вообще, а более вероятно, недостатком агрессивных самцов.
Необусловленный (или менее обусловленный) характер модели начального состояния представляет собой загадку, так как эта модель требует, чтобы самки вели себя относительно дружественно к прибывшим самцам, которые, тем не менее, первоначально ведут себя агрессивно по отношению к ним. Это вроде бы противоречит существующему долгое время в приматологии упору на индивидуальные отношения (то есть вряд ли самки не различали относительно неагрессивных резидентных самцов и агрессивных новоприбывших). У этого неожиданного вывода есть прецедент в литературе по социальной эпидемиологии, где упоминается т.н. «социальная столица», в которой увеличение здоровья и продолжительности жизни в обществе является функцией признаков, относящихся ко всему обществу, уровень которых выше уровня отдельных личностей или индивидуальных социальных сетей (Kawachi et al 1997).
Резюмируя сказанное, можно сказать, что мы наблюдали обстоятельства, в результате которых в стае павианов возникли отличительные поведенческие особенности вместе с физиологическими коррелятами. Более того, это поведение принималось новыми членами стаи; наши данные, хоть и, конечно, не убеждают полностью, дают основание предположить, что это, скорее всего, происходило по наблюдательной модели или модели способствования/начального состояния. И наконец, что до некоторой степени уникально для исследований приматов (кроме людей), наши находки касаются передачи через поколения не материальной, а социальной культуры.
В связи с этими находками возникают некоторые вопросы. Стрессовый физиологический профиль, обычно связанный с подчинённым положением в стаях павианов, влечёт отрицательные последствия для здоровья (Sapolsky 1993; Sapolsky and Share 1994; Sapolsky and Spencer 1997). Отличительные особенности связанной с рангом физиологии в Л93–96 позволяют предположить, что у подчинённых самцов из этой стаи такие патологии могут быть выражены меньше. Другой вопрос касается последствий того, что культура в Л93–96 какое-то время оставалась стабильной. Отличительной особенночтью человеческой культуры является её кумулятивность (т.е. нововведения базируются друг на друге), а для других приматов есть только в лучшем случае слабые свидетельства этого (Boesch 2003). Таким образом было бы интересно увидеть, не возникнут ли со временем у социальной традиции Л93–96 дополнительные черты.
Противоположный вопрос относится к обстоятельствам, которые могли бы разрушить культуру Л93–96. Эта культура может быть разрушена, если в стаю перейдёт много самцов одновременно, или если какой-нибудь прибывший самец вместо того, чтобы перенять местную культуру, воспользуется ею в своих целях. Теория игр предполагает, что Л93–96 уязвима для таких «мошенников». Ещё один вопрос касается судьбы самцов, рождённых в Л93–96, когда они переходят в другие стаи. Согласно моделям реципрокного альтуизма (Axelrod and Hamilton, 1981), если какой-то один самец из Л93–96, перейдя в другую стаю, продолжит вести себя в стиле, усвоенном с рождения, он окажется в конкурентном проигрыше. Однако если двое самцов из Л93–96 присоединятся к другой группе одновременно и будут между собой поддерживать принятый в Л93–96 стиль взаимодействий, они могут иметь конкурентное преимущество. Это может быть способом передачи этого социального стиля в другие стаи.
И наконец, наши находки поднимают вопрос их применимости к пониманию социального поведения человека и его передаче. В истории человечества полно примеров избирательной потери некоторых демографических слоёв общества (например, относительный недостаток взрослых мужчин после Гражданской войны в Америке или относительный недостаток девочек в современном Китае из-за практики репродуктивных технологий с уклоном в сторону мужчин и инфантицида с уклоном в сторону женщин). Настоящие данные свидетельствуют, что демографические искажения могут имет долговременные, даже затрагивающие много поколений, последствия, в том числе существенное изменение качества жизни какой-либо социальной группы.
Материалы и методы
Объектом исследования была стая (Лесная стая) павианов анубисов (Papio anubis), живущих в заповеднике Масаи Мара в Кении. Анубисы живут в стаях из 30–150 животных, включая многих самцов, с полигамией и значительной агрессией между самцами. В пубертантном периоде самцы меняют стаи, и став взрослыми, достигают некоторого ранга в несколько текучей иерархии доминирования. В противоположность этому, самки остаются в стае, где они родились, наследуя ранг, на ступеньку меньший ранга своих матерей.
Наблюдение велось в течение 1978–1986 гг. каждое лето, а с 1993 г. — непрерывно. Кроме этого, непрерывно, начиная с 1984 г., велось наблюдение над другой стаей (стаей Талек). Данные по поведению снимались как 20-минутные фокусные замеры (focal samples) (Altmann 1974). В годы, когда велось только летнее наблюдение, было собрано 45 замеров на особь в сезон; в другие годы собиралось в среднем по три замера на особь в неделю в течение всего года. Замеры распределялись в течение дня одинаковым образом для каждого индивида. В течение замера фиксировалось социальное поведение, питание и груминг. Ранги определяли по взаимодействиям «приближения-избегания», включая акты избегания (avoidances), сгоняния (supplants) и представления (presentations) при отсутствии агрессии. Агрессия в случае обострения конфликта включала выпады с открытой пастью, преследования и кусания. Перед каждым замером и после него производился обзор ближайших соседей.
Репродуктивный успех оценивали неявно на основании частоты совокуплений и сопровождений (сохранения на протяжении по крайней мере одного замера эксклюзивного совокупления и близости к самке, находящейся в эструсе). Числовой вес каждого такого сопровождения или совокупления уточнялся вероятностью зачатия в этот день (Hendrickx and Kraemer 1969).
Эндокринологические данные собирались при условиях, позволяющих измерения базального уровня стероидных гормонов (Sapolsky and Share 1997). Объектам без их ведома впрыскивали анестетик из духового шприца (blowgun syringe) между 7 и 10 утра, только в те дни, когда они не были больны, травмированы, не участвовали в сопровождении и не участвовали недавно в драке. Образцы кровы собирались в течении 3 минут после анестезии.
References
- Altmann J (1974) Observational study of behavior: Sampling methods. Behaviour 48: 1–33. Find this article online
- Aureli F, van Schaik C (1991) Post-conflict behaviour in long-tailed macaques (Macaca fascicularis). I. The social events. Ethology 89: 89–100. Find this article online
- Axelrod R, Hamilton W (1981) The evolution of cooperation. Science 211: 1390–1396. Find this article online
- Bercovitch F (1985) Reproductive tactics in adult female and adult male olive baboons [dissertation]. Los Angeles: University of California, Los Angeles. 235 p.
- Boesch C (1991) Teaching in wild chimpanzees. Anim Behav 41: 530–532. Find this article online
- Boesch C (2003) Is culture a golden barrier between human and chimpanzee? Evol Anthropol 12: 82–91. Find this article online
- Boesch C, Boesch H (1983) Optimization of nut-cracking with natural hammers by wild chimpanzees [Pan troglodytes verus]. Behaviour 83: 265–286. Find this article online
- Cambefort J (1981) A comparative study of culturally transmitted patterns of feeding habits in the chacma baboon Papio ursinus and the vervet monkey Cercopithecus aethiops. Folia Primatol 36: 243–263. Find this article online
- Castles D, Whiten A, Aureli F (1999) Social anxiety, relationships and self-directed behaviour among wild female olive baboons. Anim Behav 58: 1207–1215. Find this article online
- Cavalli-Sforza L (2000) Genes, peoples and languages. New York: North Point Press. 224 p.
- Cheney D, Seyfarth R, Silk J (1995) The role of grunts in reconciling opponents and facilitating interactions among adult female baboons. Anim Behav 50: 249–257. Find this article online
- Crook J, Gartlan J (1966) Evolution of primate societies. Nature 210: 1200–1203. Find this article online
- de Waal F (1983) Chimpanzee politics: Power and sex among apes. Baltimore (Maryland): Johns Hopkins University Press. 223 p.
- de Waal F (2000) Primates: A natural heritage of conflict resolution. Science 289: 586–590. Find this article online
- de Waal F (2001) The ape and the sushi master: Cultural reflections of a primatologist. New York: Basic Books. 256 p.
- de Waal F, Johanowicz D (1993) Modification of reconciliation behavior through social experience: An experiment with two macaque species. Child Dev 64: 897–908. Find this article online
- de Waal F, van Roosmalen A (1979) Reconciliation and consolidation among chimpanzees. Behav Ecol Sociobiol 5: 55–71. Find this article online
- de Waal F, Yoshihara D (1983) Reconciliation and redirected affection in rhesus monkeys. Behaviour 85: 224–241. Find this article online
- Galef B (1990) The question of animal culture. Hum Nature 3: 157–178. Find this article online
- Hendrickx I, Kraemer D (1969) Observations on the menstrual cycle, optimal mating time and pre-implantation embryos of the baboon, Papio anubis and Papio cynocephalus. J Reprod Fertil (Suppl.) 6: 119–131. Find this article online
- Kawachi I, Kennedy B, Lochner K, Prothrow-Stith D (1997) Social capital, income inequality, and mortality. Am J Public Health 87: 1491–1498. Find this article online
- Kawai M (1965) Newly acquired precultural behavior of the natural troop of Japanese monkeys on Kishima Islet. Primates 6: 1–30. Find this article online
- Kroeber A, Kluckhohn C (1966) Culture: A critical review of concepts and definitions. New York: Random House. 223 p.
- Kummer H (1971) Primate societies: Group techniques of ecological adaptation. Wheeling (Illinois): Harlan Davidson, Inc. 160 p.
- Laland K, Hoppitt W (2003) Do animals have culture? Evol Anthropol 12: 150–159. Find this article online
- Laland K, Reader S (1999) Foraging innovation in the guppy. Anim Behav 52: 331–340. Find this article online
- McGrew W (1998) Culture in non-human primates? Annu Rev Anthropol 27: 301–328. Find this article online
- Mercader J, Panger M, Boesch C (2002) Excavation of a chimpanzee stone tool site in the African rainforest. Science 296: 1452–1455. Find this article online
- Ninan P, Insel T, Cohen R, Cook J, Skolnick P, et al. (1982) Benzodiazepine receptor-mediated experimental “anxiety” in primates. Science 218: 1332–1334. Find this article online
- Noad M, Cato D, Bryden M, Jenner M, Jenner K (2000) Cultural revolution in whale songs. Nature 408: 537–538. Find this article online
- Noe R (1994) A model of coalition formation among male baboons with fighting ability as the crucial parameter. Anim Behav 47: 211–224. Find this article online
- Perry S, Baker M, Fedigan L, Gros-Louis J, Jack K, et al. (2003) Social conventions in wild white-faced capuchin monkeys: Evidence for traditions in a neotropical primate. Curr Anthropol 44: 241–268. Find this article online
- Pusey A, Packer C (1986) Dispersal and philopatry. In: Smuts B, Cheney D, Seyfarth R, Wrangham R, Struhsaker T, editors. Primate societies. Chicago: University of Chicago Press. pp. 250–266.
- Rendell L, Whitehead H (2001) Culture in whales and dolphins. Brain Behav Sci 24: 309–382. Find this article online
- Sapolsky R (1993) Endocrinology alfresco: Psychoendocrine studies of wild baboons. Recent Prog Horm Res 48: 437. Find this article online
- Sapolsky R (1996) Why should an aged male baboon transfer troops? Am J Primatol 39: 149–155. Find this article online
- Sapolsky R (2001) The physiological and pathophysiological implications of social stress in mammals. In: McEwen B, editor. Coping with the environment: Neural and endocrine mechanisms. Volume 4, Section 7, Handbook of physiology. Oxford: Oxford University Press. 517 p.
- Sapolsky R, Else J (1987) Bovine tuberculosis in a wild baboon population: Epidemiological aspects. J Med Primatol 16: 229–234. Find this article online
- Sapolsky R, Share L (1994) Rank-related differences in cardiovascular function among wild baboons: Role of sensitivity to glucocorticoids. Am J Primatol 32: 261–270. Find this article online
- Sapolsky R, Share L (1997) Darting terrestrial primates in the wild: A primer. Am J Primatol 44: 155–163. Find this article online
- Sapolsky R, Spencer E (1997) Social subordinance is associated with suppression of insulin-like growth factor I (IGF-I) in a population of wild primates. Am J Physiol 273: R1346. Find this article online
- Sapolsky R, Romero L, Munck A (2000) How do glucocorticoids influence the stress-response? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev 21: 55–78. Find this article online
- Seyfarth R (1976) Social relationships among adult female baboons. Anim Behav 24: 917–938. Find this article online
- Seyfarth R (1978) Social relationships among adult male and female baboons. II. Behaviour throughout the female reproductive cycle. Behaviour 64: 227–247. Find this article online
- Smith V, King A, West J (2000) A role of her own: Female cowbirds, Molothrus ater influence the development and outcome of song learning. Anim Behav 60: 599–609. Find this article online
- Smuts B (1999) Sex and friendship in baboons, 2nd ed. Cambridge (Massachusetts): Harvard University Press. 336 p.
- Strum S (1982) Agonistic dominance in male baboons: An alternative view. Int J Primatol 3: 175–188. Find this article online
- Tarara R, Suleman M, Sapolsky R, Wabomba M, Else J (1985) Tuberculosis in wild olive baboons, Papio cynocephalus anubis (Lesson), in Kenya. J Wildlife Dis 21: 137–144. Find this article online
- van Schaik C, Deaner R, Merrill M (1999) The conditions for tool use in primates: Implications for the evolution of material culture. J Hum Evol 36: 719–741. Find this article online
- van Schaik C, Ancrenaz M, Borgen G, Galdikas B, Knott C, et al. (2003) Orangutan cultures and the evolution of material culture. Science 299: 102–105. Find this article online
- Visalberghi E, Fragaszy D (1990) Food-washing behaviour in tufted capuchin monkeys, Cebus apella and crab-eating macaques, Macaca fascicularis. Anim Behav 40: 829–836. Find this article online
- Whiten A (1998) Imitation of the sequential structure of actions by chimpanzees (Pan troglodytes). J Comp Psychol 112: 270–281. Find this article online
- Whiten A, Goodall J, McGrew W, Nishida T, Reynolds V, et al. (1999) Culture in chimpanzees. Nature 399: 682–685. Find this article online
- Whiten A, Horner V, Marshall-Pescini S (2003) Cultural panthropology. Evol Anthropol 12: 92–105. Find this article online
Оригинал в Plos Biology
Жаль, переводчик остался неизвестен
Рекомендуем прочесть
 Угнетение бедностью
Угнетение бедностью #Книги // Рабочая (работающая) Америка, 2012 (The State of Working America, 12th edition)
#Книги // Рабочая (работающая) Америка, 2012 (The State of Working America, 12th edition) Анализ научного текста и новые мировые тенденции
Анализ научного текста и новые мировые тенденции Роль демократии в сокращении конфликтов преувеличена
Роль демократии в сокращении конфликтов преувеличена Мимозка и индийский кулан: саморазрушительный симбиоз
Мимозка и индийский кулан: саморазрушительный симбиоз «Мужские права» и выгребная яма истории
«Мужские права» и выгребная яма истории Палеогенетики обнаружили «двойное гражданство» у домашних кошек.
Палеогенетики обнаружили «двойное гражданство» у домашних кошек. Кто и против кого выступал в Таиланде в минувшем году. Портрет противников
Кто и против кого выступал в Таиланде в минувшем году. Портрет противников
