

Резюме. Ещё одна «работа над ошибками»: почему урбанизацию «диких» видов птиц и млекопитающих нельзя интерпретировать по лекалам доместикации домашних (увы, я сперва данному подходу доверял). Обсуждаются системные различия поведенческих изменений при урбанизации «диких» видов птиц и млекопитающих, с одной стороны, и доместикации, с другой. Показана, что распространённое сегодня использование второй как модели и лекала в анализе первой — ошибка, основанная на ложной аналогии. Анализ агрессивности, оборонительного исследовательского поведения, отношения к новизне у «городских» птиц показывает, что сравнительно с особями исходных популяций они не делаются ни «добрей», ни «доверчивей» (как это случилось с домашними животными).
Даже наоборот: агрессивность «городских» птиц возрастает, вместе со смелостью и лучшим отреагированием потенциальной опасности, более точной дифференциацией её от «просто беспокойства» (к нему «городские» особи делаются всё индифферентней). Крайне популярный сейчас показатель дистанции взлета отражает не первое, а второе, почему его уменьшение (в среднем) в городах — признак большей устойчивости популяционной системы к средовому стрессу «вообще», а не рост доверчивости особей к конкретным проблемам или опасностям. По мере урбанизации вида повторяемость поведения птиц падает, агрессия, смелость, бегство от потенциальной опасности или, напротив, принятие риска с исследованием новых мест и объектов из индивидуальной характеристики делаются ситуативными, поведение в целом — всё более гибким и контекстуальным, с лучшим распознаванием специфики ситуации, выбором способа действия, более точным «дозированием» реакций по цели (в чём состоит развивающая роль урбосреды).
Урбанизация разрушает т. н. поведенческий синдром (англ. behavioural syndrome): существующие в исходных популяциях коррелятивные связи между разными поведенческими характеристиками, прежде всего, исследованием,смелостью, агрессией и принятием риска.
Запуски связанных с ними форм поведения у «городских» птиц взаимно независимы, в противоположность «сельским» или «лесным» особям, что максимизирует точность выбора поведения в проблемной ситуации и его переключения на другое сообразно обстановке.
Средневековая фраза «городской воздух делает свободным» распространяется на организацию поведения «городских» птиц и млекопитающих, тем сильней, чем больше времени прошло от начала урбанизации вида. Реже урбанизация вида перестраивает поведенческий синдром в качественно иной. Напротив, доместикация создаёт свой синдром поведенческих и морфологических признаков, отсутствующий в природных популяциях. Показано, что образование городской популяции, становление её устойчивости на территории города идут при такой же осторожности к человеку и технике, вообще «городскому метаболизму», что и в исходных. Она понижается только по завершении урбанизации, когда вид растёт численно на территории города, иногда с запозданием на десятки лет.
1. Урбанизация не меняет отношения к реальной опасности
Содержание
- 1 1. Урбанизация не меняет отношения к реальной опасности
- 2 2. Урбанизация усиливает агрессию
- 3 3. Разрушение поведенческого синдрома при урбанизации
- 4 4. Дистанция взлёта — маркёр резистентности к беспокойству, а не ответа на опасность
- 5 5. Осторожность городских птиц не снижается, но делается контекстуальней
Объяснение неизменного сокращения дистанции взлёта «городских» птиц отбором на доброжелательность к людям — ключевое звено отождествлений урбанизации и доместикации (Møller, 2010, 2014; Møller et al., 2008, 2012, 2015). Если вынуть его, распадётся вся цепь, и сделать это необходимо — за внешним сходством здесь скрыты разные причины.
Главное в одомашнивании — отбор на позитивное восприятие людей, вольера и т. д. искусственной обстановке, организующей жизнь животных, прочие изменения происходят по корреляциям (Трут, 2007). Для «диких» предков это опасность, «включающая» оборонительное поведение — сперва настороженность, потом бегство, при невозможности избежать — агрессию, и всегда сильный стресс. Напротив, одомашненные потомки всё названное воспринимают положительно (меньше страха, меньше агрессии) и активно используют в поведении, от побуждения к «социальным подсказкам» до постановки лап на вольерную сетку беляевских лис (Hare et al., 2012; Хейр, Вудс, 2014; Nawroth et al., 2016). Психофизиологические изменения, созданные отбором, «превратили Савла в Павла»: животные стали «добрей» и «доверчивей» к объектам, ранее вызывавшим страх и агрессию.
Сходным образом объясняется неизменное сокращение средней дистанции взлёта1 при урбанизации разных видов птиц. Различия в экологии здесь нивелируются по мере освоения «рукотворных ландшафтов», а особенно городов: перекрывание кормовых и пространственных ниш растёт пропорционально степени трансформации полной неразличимости (Фридман, Суслов, 2018). Напротив, различия в психике (в степени неофобии, смелости, исследовательском поведении, инновативности, в прочих качествах, значимых в освоении новой и экстремальной среды) вполне сохранны, хотя изменяемы урбанизацией. Что особенно видно в реакции на новизну — скорость начала кормёжки на новых кормушках (данные по разным польским городам, Tryjanowski et al., 2015, 2016) и на знакомых кормушках с новым, пугающим птиц объектом (изучено в связи с инновативностью у 9 видов птиц о.Барбадос: трёх голубиных — Zenaida aurita, Columbina passerina и Patagioenas squamosus, 6 воробьиных — танагр L.barbadensis и Tiaris bicolor, трупиалов Quiscalus lugubris и Molothrus bonariensis, тиранна Tyrannus dominicensis, цветочницы С.flaveola, Ducatez et al., 2017).
Мол, в «сельских» или «лесных» популяциях вида появление человека (собак, техники) было опасностью, реакция на которую — настороженность, тревожные крики, при большем сближении — бегство. Там особи приспособлены к своевременному обнаружению и избеганию этих контактов, что отражает дистанция бегства, оцениваемая в подходах наблюдателя к птицам2 (Møller, 2008, 2014; Blumstein, 2014). И вот, то же самое давление отбора, что «сделало» ручными беляевских лис, по ходу урбанизации меняет психофизиологию «городских» птиц так, что прежняя опасность им делается безразличной, а потом и вызывает позитивное отношение: люди, их постройки, коммуникации, техника толкают к исследованию и использованию «в интересах» животного. Согласно этому объяснению, в межпопуляционных сравнениях сокращение дистанции взлёта у «городских» птиц сравнительно с «сельскими» и «лесными» есть мера урбанизованности популяции, т. к. отражает степень «претворения» прежнего страха и стресса в позитивное отношение под влиянием того же отбора, что действовал при доместикации (Møller, 2009, 2010; Carrete, Tella, 2011, 2017).
Так, сравнением изменений дистанции взлёта в 714 «сельских» и «городских» популяциях 159 видов птиц на широтном градиенте с севера на юг Европы (n=12495) показано, что у тех и других она сокращается, корреллируя с меньшим обилием хищных птиц. Однако они тоже урбанизируются, и многие виды в городах многочисленней, чем вовне (Фридман, Ерёмкин, 2009: 25-26; Фридман и др., 2016). Дистанция взлёта при подходе человека у «городских» птиц везде ниже, чем у «сельских», но разность между ними при большем обилии хищников в южной Европе оказывается больше, чем в Центральной и Северной, а не меньше, как следует из рабочей гипотезы авторов (не заметивших это расхождение данных с сделанными предположениями, Diaz et al., 2013).
Они же в межвидовых сравнениях рассматривают различия дистанций взлёта (в одной и той же ситуации «подхода» и вспугивания) как предикторы разной предрасположенности видов к урбанизации (чем она меньше — тем более, Møller, 2010, 2014). Увы, другие их данные показывают поверхностность этого сходства. Анализ зависимости оборонительного поведения при отлове от уязвимости «городских» vs «сельских» птиц к нападениям домашних кошек Felis catus или перепелятников Accipiter nisus подтверждает снижение страха у первых3 (данные по 15 видам, 1132 особям в Гранаде (Испания) и Brønderslev (Дания), Møller, Ibáñez-Álamo, 2012). В руках они реже стараются вырваться, реже клюются (для авторов — признак сниженной агрессивности, хотя это сомнительно), чаще остаются неподвижно лежать на открытой ладони. Однако они чаще издают тревожные крики (т. е. лучше квалифицируют случившееся как «поимку хищником») и «крики ужаса», чаще теряют перья, т. е. сильнее стрессированы (Møller, Ibáñez-Álamo, 2012, table 1-2).
Попытки вывести эти различия из уязвимости к нападениям кошек или перепелятников неудачны. Для большинства поведенческих переменных (табл.1-2 статьи) влияние данных факторов не обнаруживается, за исключением частоты потери перьев и клевков при поимке. В первом случае значимо влияние риска нападения кошек, во втором — перепелятников, причём уязвимости к этим опасностям отрицательно коррелируют между собой, а изменения поведения между «городскими» и «сельскими» особями однонаправленны. Наконец, различия в частоте потери перьев между ними коррелируют с риском атаки перепелятника, но отрицательно: он ниже именно там, где перья теряют чаще (Møller, Ibáñez-Álamo, 2012, fig.1). Межгрупповые различия в частоте клеваний руки лучше объяснимы временем с момента урбанизации, чем опасностью «здесь и сейчас» (Møller, Ibáñez-Álamo, 2012, fig.1, 2).
Действительно, наследуемость укороченной дистанции взлёта «городских» кроличьих сычей Athene cunicularia значимо ниже (в 2-3 раза, в зависимости от метода установления), чем вне города (Carrete et al., 2016). Этот вид-специалист открытых травянистых ландшафтов стал успешным синурбанистом по всему ареалу, в США и в Южной Америке. У «городских» птиц выше и выживаемость, и репродуктивный успех, их популяции растут численно несмотря на застройку и т. д. (Rebolo-Ifrán et al., 2017; Baladron et al., 2020).

Семья кроличьих сычей в урболандшафте
Индивидуальная повторяемость дистанции взлёта кроличьего сыча выше, чем у других видов, т. е. именно здесь она максимально характеризует особь, а не ситуацию. Падение наследуемости при урбанизации, однако, показывает рост ситуативности данного показателя также и здесь (Carrete, Tella, 2013, 2017); тем более это верно при однотипном сокращении дистанции взлёта у прочих видов. Эти данные ярче всего показывают фиктивность «отбора на доброжелательность» к людям и новизне при урбанизации или, как минимум, неадекватность попыток его исследовать, используя уменьшение дистанции взлёта (Møller, Díaz, 2018; Geffroy et al., 2020). Так загрязнение воздуха не определить по запаху, хотя в первую очередь обращаешь внимание именно на него.
 2. Урбанизация усиливает агрессию
2. Урбанизация усиливает агрессию
Доместицированные виды в сравнении с «дикими» предками действительно делаются «добрей» и «доверчивей» к людям и обстановке, ими созданной и совершенствуемой. Птицы в урбанизированных популяциях — скорее наоборот: (I). C ростом «смелости» агрессивность4 «городских» птиц и млекопитающих чаще всего увеличивается, в ответ на специфические стимулы и просто на беспокойство (т. н. urban wildlife syndrome, Parker, Nylon, 2008; Evans et al., 2010; Lowry et al., 2013)5.
Единственный аспект, в котором она безусловно снижается у птиц и млекопитающих — образование скоплений высокой плотности при кормлении, на гнездовании и т.д. ситуациях, особенно вызванных «точечностью» подходящих городских территорий. В сравнении с сельскими популяциями, здесь выше терпимость к близости конспецификов, агрессия более или менее замещена персонализованными взаимоотношениями или (в анонимных скоплениях) безразличием. Это т. н. поведенческий островной эффект (англ. island effect), описанный у «городских» млекопитающих, рептилий (Baxter-Gilbert, Whiting, 2019) и особенно птиц. Сопряжение ситуативного роста терпимости в одних ситуациях взаимодействия с конспецификами с общим подъёмом агрессии в других отражает лучшую переключаемость агрессивности «городских» особей сообразно ситуации (Garroway, Sheldon, 2013; Lowry et al., 2013; Hardman, Dalesman, 2018). Его проявление — как и подъёма агрессивности, сопряжённой со смелостью, у «городских» особей — сильнее всего зависит от конфигурации «архипелага» видовых биотопов, размеров, связности «островов», степени изменённости их растительности, а также плотности населения, но почти не зависит от видовой специфики (Parker, Nylon, 2008, 2012).

У «городских» водяных легуанов Intellagama lesueurii встречаемость следов от укусов растёт сравнительно с их числом в полуприродных и природных биотопах, но лишь у самцов. У самок растёт в полуприродных биотопах сравнительно с природными, но значимо не меняется при переходе в городские. Сила укуса «городских» особей, однако, не увеличивалась, как это бывает у ящериц c ростом агрессии, вызванным переуплотнением. Учитывая подъём плотности населения по градиенту урбанизации «вверх» и специфику социальной организации вида (самцы охраняют территории, перекрывающиеся с неохраняемыми участками обитания самок, образующих группы дружественных особей, либо используют стратегию «рейнджеров», в поисках контактов с самками вторгающихся на участки территориальных особей и отступающих при отпоре; в обоих случаях социальные отношения самцов структурированы конкуренцией за самок) урбанизация усиливает агрессию не «вообще», но в ситуациях, и ранее значимых для социальной организации вида (Baxter-Gilbert, Whiting, 2019).

Большая агрессивность при охране гнезда и смелость по отношению к людям во время кормёжки показаны для городских лысух Fulica atra. Выраженность обоих различий пропорциональна времени с начала урбанизации; степень стрессированности и использование антропогенных кормов, напротив, не изменялись. Лысухи направленно выбирали территории, позволяющие вырастить больше птенцов, «оценивая» структуру растительности по качеству укрытия будущего гнезда и подходов к нему. Лучшие они занимали дольше, там держались устойчивей и защищали агрессивней. Не отмечено сколько-нибудь сходной оценки источников антропогенных кормов (Minias, Janiszewski, 2016; Minias et al., 2017, 2018).

Выводок лысух на Мартыновском пруду в Санкт-Петербурге. 5 августа 2020 г. Источник Фонтанка.ру
У чёрного коршуна Milvus migrans в разных префектурах вокруг залива Сагами (Япония) интенсивность атак на человека и территориальная агрессия увеличиваются при большей урбанизации территории, т. е. при меньшей площади лесных и с/хландшафтов, приходящейся на птицу (кроме рисовых чеков, Galbreath et al., 2014). Рост агрессивности вместе со смелостью отмечен у «городских» певчих воробьёв Melospiza melodia сравнительно с «сельскими» (Scales et al., 2011), сопряжённо с большим уровнем кортикостерона, общим и увеличенным в ответ на стрессор — но не тестостерона (Davies et al., 2017). Его уровень тестостерона в «городских» популяциях не увеличен или (в меньшем числе случаев — у чёрных дроздов, певчих воробьёв) даже снижается (Fokidis et al., 2011; Bonier, 2012, table 2; Partecke, 2014; Davies, Sewall, 2016; Davies et al., 2017). Видимо, это общее правило.
Т.е. гормональные изменения при урбанизации чаще всего не сопряжены с поведенческими или морфологическими, что представляет собой часть разрушения (реже — перестройки) поведенческого синдрома, формирующего индивидуальность птиц в исходных популяциях. Подробный анализ ниже, здесь отметим, что так поведение «городских» птиц не просто разнообразится, но, главное, делается много менее инерционным. Выбор поведения в следующий момент (в т.ч. и агрессии) меньше канализирован предыдущей активностью и её мотивацией, облегчается переключение к другим её видам, не «продолжающим» данную, но альтернативным (противоположно разрешающим конфликт «приближение — бегство», наиболее важный в урбосреде, вроде неофобии и неофилии, агрессии и бегства, отступления и смелости, бегства и приближения с исследованием и пр.). Во внегородских популяциях это затруднено ввиду отрицательных корреляций между поведенческими альтернативами в рамках «синдрома», и положительных — поведения со своей мотивационной подосновой. У «городских» птиц первое более или менее эмансипируется от второго — как это присуще ритуализованным демонстрациям всех видов птиц независимо от урбанизации (Фридман, 2013).

Агрессивный ответ на проигрывание песни также усилен у «городских» больших синиц сравнительно с «лесными», однако сильнее варьирует в разные дни, в разной обстановке, в разных районах города и пр. (Hardman, Dalesman, 2018). Что независимо подтверждено данными о большей интенсивности у них 1) тревожной вокализации (обусловленной готовностью окрикивать хищника, не боясь его атаки) и 2) клевков при взятии в руки (Senar et al., 2017).
«Городские» кроличьи сычи агрессивней по отношению к хищнику, чью пластиковую модель выставляли у входа в нору: меньше времени проходит до появления птиц у модели и начала агрессивных демонстраций/атак. Они также менее неофобны (быстрее исследуют новый корм) и терпимей по отношению к человеку (подпускают ближе), у них сильней корреляция между принятием риска (отражённым в падении дистанции взлёта) и исследованием, а связь между дистанцией взлёта и атакой на хищника, напротив, разорвана. Птицы с меньшей дистанцией взлёта быстрее используют новый корм, активней исследующие активней и в атаке на хищника. Здесь урбанизация ликвидирует поведенческий синдром, бывший во внегородских популяциях, и создаёт качественно иной (Carrete, Tella, 2017, table.2-3).

Сходный рост территориальной агрессии в ответ на проигрывание песни/позывов конспецификов показан у видов, исходно связанных с растительностью пустыни, пятнистого кривоклювого пересмешника Toxostoma curvirostre и черногорлого тауи Melozone aberti. освоивших г.Феникс в штате Аризона. Здесь у «городских» птиц ниже латентный период сближения с «нарушителем», длительней выдача специфических ответов на «вторжение» (пяти разных типов), чем в исходных популяциях; неспецифические ответы, в т.ч. признаки общего возбуждения значимо не отличались (Fokidis et al., 2011, table 2-3, fig.1).

Черногорлый тауи в г.Финикс, Аризона. Источник stonechat
Интенсивность ответа на проигрывание у пересмешников значимо выше там, где остались большие площади пустынной растительности, ниже — где она в основном экзотическая, у тауи — в местах с большей плотностью населения. Интересно, что рост ритуализованной агрессии при урбанизации произошёл без значимых изменений уровней тестостерона и кортикостерона. Ситуативные изменения территориальности обоих видов также не связаны с уровнями данных гормонов. Однако поимка и взятие в руки «городских» пересмешников снижает их общий уровень тестостерона значимо сильней, чем «пустынных». Она же повышает уровень общего и свободного кортикостерона значительней у «городских» тауи, т.е. они больше стрессируются реальной опасностью (Fokidis et al., 2011).

Пятнистый кривоклювый пересмешник в г.Финикс, Аризона, Источник stonechat
Подъём агрессии отмечен в урбанизированных популяциях двух австралийских видов6: вороны-свистуна Gymnorhina tibicen и черношапочной манорины Manorina melanocephala. 71% первых атакуют кого-то из трёх видов нарушителей их гнездового участка — прохожих, велосипедистов или почтальонов, треть непременно гоняет пешеходов. Распознавая людей индивидуально, вороны особенно агрессивны к некоторым из них, ими ассоциируемым с реальной опасностью. Сходная дифференциация нарушителей, с более интенсивной реакцией в дальнейшем на «особо опасных», отмечена у «городских» многоголосых пересмешников. Птицы таковыми «сочли» наблюдателей, в течение 4-х дней подходивших к гнездовому дереву, стоявших там 30 с и 15 с касавшихся рукой гнезда. Их сразу запомнили и потом реагировали всё агрессивней, обнаруживая издалека (с 14 м.) при равнодушии к обычным пешеходам, в т.ч. шедшим прямо под деревом (Levey et al., 2009; Lowry et al., 2013).
Второй вид при урбанизации меняет реагирование на беспокоящий стимул — громкий глухой звук, сходный с работой индустриального пресса (80 дБ, на 15 дБ больше обычно используемых в изучении антропогенного шума). «Городских» или «сельских» птиц в Мельбурне (или окрест) привлекали криком «на хищника», включённым в агрессивный ответ на опасность вместе с некоторыми демонстрациями (Lowry et al., 2011, table 1). Наблюдатель отмечал число прибывших птиц, выбирал каждую вторую слева в группе, и дальше прослеживал их её поведение в ответ на шум (проигрываемый с перерывом в 3 с после 40 с криков) или в контроле (ничего не делали, лишь наблюдали). Хронометраж вёлся издалека и неброско одетыми, т.ч. на птиц влияли лишь звуки (Lowry et al., 2011).

Предупреждающий сигнал черношапочной манорины на кормушке лорикету
Реакция черношапочных манорин на оба стимула задана известным конфликтом «нападение-бегство». «Соотношение сил» обоих у данной конкретной птицы хорошо видно по поведению (Lowry et al., 2011, table 1). Максимум «бегства» проявляется в отлёте на >5 м, его уменьшение — в отлёте на дистанцию покороче; ещё меньший уровень — когда птица, взлетев, не отлетает прочь, а делает полёт-прыжок, приземляясь на то же место или другое, но не далее метра. Примерное равновесие побуждений выражено интенсификацией осматриваний: птица остаётся сидеть и усиленно вертит головой. Превалирование агрессии видно уже по специфическим демонстрациям и крикам: «заострённой» позе с прижатым оперением, тело, шея и клюв наклонены вниз и вытянуты вперёд, к источнику проблем. Часто птицы издают тот самый крик «на хищника», которым их привлекали. При дальнейшем подъёме агрессии птица, оставаясь такой же вытянутой, раскрывает крылья, сгибает в суставах запястья и машет ими вверх-вниз, с поднятым вверх хвостом, открытым клювом и непрерывными криками. Максимум агрессивности выражен в движении к источнику проблем в «сгорбленной» позе с прижатым оперением (тело обращено к нему, но голова и клюв — вниз), прыжками или полётом-прыжками. Дальше следуют активный подлёт и атака (Lowry et al., 2011).
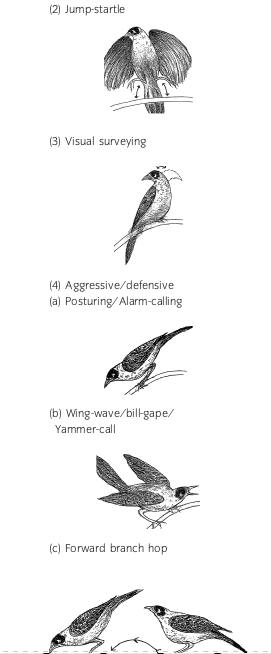
Внешние выражения конфликта нападение-бегство у черношапочной манорины. Источник Lowry et al., 2011.
Lowry et al. (2011) отметили повышенную реактивность «городских» манорин при проигрывании криков «на хищника». Первые реагируют более коллективно: чаще всего приближаются группами >10 особей (35% случаев против 5%) и наоборот, группами <5 и 5-10 птиц приближаются реже (22,5% и 0% у первых, по 47,5% у вторых). В реакции на беспокоящий звук «лесные» манорины демонстрируют в основном избегание (слетает прочь половина, среди «городских» — 22,5%), «городские» — интерес, приближение и спаянную с ними агрессию (треть птиц против 5% у «лесных»). Большая интенсивность испуга «лесных» манорин также выражена в большей дистанции слёта: далее 5 м улетает 1 из 9 «городских» птиц и 13 из 19 «лесных». Примерное равенство конкурирующих побуждений у тех и других встречено одинаково часто (Lowry et al., 2011, table 2).
«Городские» манорины были вообще возбудимее «лесных»: 15% первых и лишь 7,5% вторых сменило поведение в контрольной ситуации, т. е. спонтанно, просто в новом месте, хотя большинство было индифферентно. Из фокальных птиц лишь один «городской» медосос (но большинство «лесных») в опытной ситуации слетел с дерева, на которое они прибыли. Далее первые значимо чаще интересовались источником звука и исследовали его (помимо агрессии). Рост смелости и исследовательской активности «городских» медососов могут быть следствием больших плотности населения и размера колоний. Для больших синиц показано увеличение того и другого в присутствии компаньона (Lowry et al., 2011).
Агрессивный ответ на угрозу гнезду отмечен и в других урбанизированных популяциях, по контрасту с тревогой и бегством в исходных (Jones, 2008; Levey et al. 2009). В случае медососов стимул, использованный для её вызывания, был им неизвестен, не связан прямо с людьми и сам по себе не давал информации, опасен он птицам/гнезду или нет, в отличие от видимого движения угроз, использованных в прочих работах. Поэтому привыкание к нему в предыдущие годы жизни в городе маловероятно, что ведёт к выводу о качественном изменении мотивационной подосновы поведения при урбанизации вида (Lowry et al., 2011).
Увеличение агрессивности «городских» птиц не всегда заметно, т. к. они лучше дозируют «выплеск» агрессии по цели, последний сильнее зависит от контекста. Уменьшение повторяемости агрессии отражает её превращение из индивидуальной характеристики в преимущественно ситуативную7 (Hardman, Dalesman, 2018). В эту же сторону изменяются прочие поведенческие характеристики «городских» птиц, прежде всего дистанция взлёта, неофобия/неофилия, смелость и готовность исследовать новые территории, объекты и т.д.
Так, на юго-западе Аргентины сравнивали привыкание «городских» vs «сельских» кроличьих сычей к движению наблюдателя в сторону птиц, сидящих у гнездовой норы. Он вызывает страх (оцененный дистанцией взлёта) и/или агрессию, оцененную баллами по пяти видам реакции: вокализациям, поклонам, распушению перьев с раскрыванием крыльев (как бы пытаясь стать больше), угрожающему полёту и демонстрации «ныряющей атаки». Этот ряд соответствует росту угрозы гнезду и отражает уровень агрессии владельца. Опыт с подходом ставили в двух вариантах: 1) в течение 9 дней подряд (по утрам) и 2) по 5 раз в один и тот же день с 50минутным перерывом, фиксируя, будет реакция усиливаться или, наоборот, ослабляться после того, как выяснена безопасность объекта (Cavalli et al., 2018).
Дистанция взлёта «сельских» сычей была выше, однако укорачивалась в первом варианте опыта (однотипно у самок и самцов); у «городских» же она много ниже и неизменна. Во втором варианте она укорачивалась в обоих популяциях, но более выражено у «сельских» сычей. Однако «городские» сычи значимо агрессивнее к хищникам, связанным именно с урбосредой, вроде собак; правда, в обоих популяциях агрессия сходно растёт на протяжении жизненного цикла. Везде самки A.cunicularia агрессивней самцов реагируют на потенциальную опасность у гнёзд, но между популяциями и повторностями опыта не нашли значимых различий в агрессивности в обоих его вариантах (Cavalli et al., 2018).
Результат отражает общее правило: видовые сигналы в урбосреде, вопреки её зашумлению и обилию разнотипных «помех» коммуникации (машины, прохожие, громкие звуки etc.), в полной мере сохраняют семантику и эффект (Phillips, Derryberry, 2018; Akçay et al., 2020а), а если меняются в частоте, интенсивности, других особенностях выдачи, то чтобы сохранить инвариант формы вопреки «шуму» в канале связи (Akçay, Beecher, 2019). В одной и той же ситуации «городские» птицы чаще используют ритуализованные демонстрации, позывы, «тихие» песни, т. е. сигналы, действующие на короткой дистанции, в сравнении с дальнодействующими, прежде всего рекламным пением. Способность обнаружить захватчика, самку и других значимых адресатов демонстративного поведения при этом не изменяется (Ríos-Chelén et al., 2015; Narango, Rodewald, 2018; Grabarczyk, Gill, 2019). Зашумлённость урбосреды также увеличивает агрессивность, ведёт к большей интенсивности специфических ответов на проигрывание пения конспецификов (Akçay et al., 2020b).
Следовательно, у «городских» сычей укорочено последействие привыкания к потенциальной опасности. Появившись, оно действует в разные моменты одного дня, но не в разные дни. Что вполне адаптивно — ситуация вокруг «городской» птицы за сутки может резко меняться: скажем, появятся подросток или кот, реально опасные для них и гнезда. Вокруг «сельских» птиц это менее вероятно: и меньше людей/кошек/собак оказывается на участке в единицу времени, бедней и разнообразие их намерений. Авторы делают вывод, что «городские» сычи пластичней, и быстрей «перестраивают» активность сообразно ситуации, чем «сельские» (Cavalli et al., 2018).
Описан неслучайный «переток» A.cunicularia между обоими популяциями, показывающий, что особи именно с данными качествами предпочтительно заселяют город из региона и/или остаются в нём, будучи там рождёнными. Поэтому большая филопатрия «сельских» сычей коррелирует с их индивидуальностью (большими неофобией и страхом перед людьми); напротив, у «городских» она независима от неё, но пропорциональна плотности поселений (Rebolo-Ifrán et al., 2015; Luna et al., 2019).
3. Разрушение поведенческого синдрома при урбанизации
Большая гибкость/контекстуальность «управления выдачей» этой и других форм поведения «городских» птиц неизменно сопряжены с направленным изменением при урбанизации «среднего уровня» связанных с ними качеств особи. С одной стороны, поведенческие изменения в ответ на проблемную ситуацию (вроде прикрепления к привычной кормушке с двух сторон цветных пластиковых стержней, или шара, или трёх шаров с разных сторон) в урбосреде сильно зависимы от особенностей обстановки, таких как как расстояние от источников антропогенного корма, вроде отелей и пикниковых точек в цитируемом исследовании 9 барбадосских видов (Ducatez et al., 2017). С другой, сами поведенческие изменения при урбанизации устойчивы и однонаправлены: дистанция взлёта/неофобия падают, смелость/неофилия растут, как и инновативность, готовность исследовать новое пространство или объект, причём «городские» особи обследуют интенсивней, и движутся быстрее, но менее тщательно, и т. д. см.разд.2.
Мы видим (II): урбанизация меняет не только выраженность разных форм поведения (агрессии, беспокойства, окрикивания хищника, и пр.), но, главное, их распространённость между особями разного «качества» и/или связанность разных форм в индивидуальной активности. Последнее — т. н. поведенческий синдром, связанный с индивидуальностью особи, иногда — у большой синицы и ряда других видов — во многом её формирующий (Sih et al., 2004; Sih, de Guidance, 2012).
Урбанизация его разрушает или (реже) перестраивает. В исходных популяциях характеристики, связанные с альтернативами в реакции на новизну, она же потенциальная опасность (принимать риск, действуя сообразно ситуации и/или избегать его, отступая прочь и т. д.) чаще всего коррелируют между собой. Большая смелость (принятие риска, проактивное разрешение проблемных ситуаций, с антитезой — пассивным переносом) здесь положительно связаны с большей агрессией, часто также и с большей активностью, и отрицательно — с неофобией в отношении корма или объекта.

Искусственный отбор, проведённый на больших синицах, данные антитезы закрепил и усилил. Птицы, селектированные на смелость и скорость исследования, при предъявлении новых объектов чаще сближались с ними, активнее изучали. Отобранные на робость/медленное исследование, напротив, замирали и были сильно стрессированы. Однако патологическая стереотипия в присутствии новизны в обоих вариантах одинаково часта, линии сравнимы по средней величине выброса кортикостерона в присутствии новых объектов, но только у смелых птиц он был в целом сильнее; только у робких развившийся при этом стресс коррелировал с величиной выброса (Baugh et al., 2017).
Иными словами, большая дивергенция в типах индивидуальности, достигнутая за счёт отбора, отнюдь не снижает страх перед потенциальной опасностью на физиологическом уровне. Также воздействовала на больших синиц и урбанизация. В урбосреде дихотомия «быстрых» vs «медленных» исследователей выражена чётче, устойчивей, связана с отбором по «генам индивидуальности», SERT и DRD4. Каждая из альтернатив индивидуальности «нашла» себе наиболее подходящий участок урболандшафта с преобладанием именно тех потенциально опасных объектов, к которым устойчивей именно он. Для первых это машины, вторых — прохожие, их те или другие синицы подпускают ближе всего; сообразно этому оба варианта неслучайно перераспределяются между районами города (Sprau, Dingemanse, 2017).
В естественной среде обитания данные корреляции свойственны всем позвоночным (Sih et al., 2004; Bókony et al., 2012; Miranda, 2017), но «рассыпаются» при урбанизации вида. Вариации выраженности разных характеристик оказываются несвязанными у одних и тех же индивидов, хотя в «городской» популяции их средние значениям меняются в одну сторону относительно исходных. Так, у «сельских», но не «городских» чиманго Milvago chimango уровни неофобии и исследовательской активности скоррелированы. Страх перед приближением к новому объекту здесь положительно связан с временем, проходящим до первого контакта с ним, и отрицательно — с длительностью последующего исследования (Biondi et al., 2020, fig.4-5).

Меченый чиманго ищет корм в выброшенном мусоре в г.Санта-Роза, Аргентина (Solare, Sarasola, 2019)
.«Городские» птицы смелей, менее неофобны, быстрей/интенсивней исследуют новый объект, помещаемый близ кормушки, однако разные характеристики при переходе от «сельских» птиц к «городским» меняются независимо друг от друга. Не выявлено различий между популяциями в общих затратах на исследование новых объектов, также как связи между неофобией и длительностью исследования после того, как птица таки решилась приблизиться. Чиманго из пригородов по всем характеристикам ближе к «городским», показывая, среди прочего, что «старт» урбанизации дан именно там (Biondi et al., 2020).
Лодзинские лысухи8 меньше укрывают гнездо (располагают его ближе к берегу и открытой воде, часто на открытых местах) и агрессивней в его защите: ближе подпускают людей к гнезду, чаще атакуют их. У них также снижен уровень стресса (оценка пропорцией гетерофилов/лимфоцитов в крови). Уменьшение страха перед человеком и хищниками при размещении гнезда, однако, не коррелируют с агрессией в его защите от людей, других лысух и водоплавающих птиц. Её уровень пропорционален числу гнездовых пар лысух на водоёме, т.е.зависит от контекста (Minias, 2015; Minias et al., 2018).
У 9 видов барбадосских птиц, значительно различающимся по инновативности (взято из базы данных Overington et al., 2009) последняя отрицательно связана с дистанцией взлёта, но только вне города, в урбанизированных популяциях связь отсутствует. «Городские» L.barbadensis смелей и лучше решают проблемные задачи, чем «сельские», но добавление новых объектов не меняет их времени возвращения на кормушку (Ducatez et al., 2017, fig.1).
Отрицательная связь между острым стрессом (поимка и взятие крови) и территориальностью выявлена у «сельских», но не «городских» J.hyemalis. Закартировав территории, дальше проигрывали записи песен владельцам, фиксируя поведение в ответ на «захватчика». У «городских» юнко оно интенсивней по всем контролируемым переменным, показывая рост агрессии при урбанизации. У «сельских», но не «городских» юнко последняя долговременно подавляется острым (но не контрольным) стрессором; по такому параметру, как время, проведённое на минимальной дистанции от «захватчика», у стрессированных «городских» птиц она даже усилена. Интересно, что сила ответа на стрессор не связана с телесными (ширина груди) и поведенческими (дистанция взлёта) характеристиками, отражающими индивидуальную чувствительность к нему, т. е. отмеченные изменения отражают преобразования на уровне всей популяции (Abolins-Abols et al., 2016, сравни Фридман и др., 2016).
Сходным образом исчезли корреляции между качествами, составляющими индивидуальность, при урбанизации других видов птиц: серых юнко Junco hyemalis (Atwell et al., 2012), чёрных дроздов Turdus merula (Miranda et al., 2013), больших синиц (Riyachi et al., 2017); певчих воробьёв (Evans et al., 2010; Scales et al., 2011) и др. Сходные изменения фиксируются при урбанизации млекопитающих: щёткохвостые поссумы Trichosurus vulpecula на границе урболандшафтов с лесными в Сиднее (Wat et al., 2020); обыкновенные полёвки Microtus arvalis и полевые мыши Apodemus agrarius из городских (Берлин), пригородных и сельских местностей в северо-западном Барнденбурге9 (Massa et al., 2020; Dammhahn et al., 2020), а также рептилии (водяной легуан, Baxter-Gilbert et al., 2019).

«Городские» юнко в Сан-Диего
Иногда поведенческий синдром при урбанизации лишь перестраивается в качественно иной (кроличий сыч, Carrete, Tella, 2017; северный кардинал, Huang et al., 2020). Наиболее сохранен поведенческий синдром у домового воробья Passer domesticus — одного из двух видов птиц, у которых не описаны популяции вне населённых пунктов (второй — домовая ворона Corvus splendens). Он включает принятие риска, страх перед новым объектом и/или видом корма (фиксировали латентный период до приближения к чучелу кольчатой горлицы, чучелу хищника и новому корму, и число перелётов по приближению) и общую активность. Синдром равно присущ всем изученным птицам из 4 населённых пунктов Венгрии, взятых на градиенте урбанизации от деревень (Nemesvámos и Dóramajor, птицы пойманы на животноводческом комплексе и в сыроварне) до среднего (Veszprém, пойманы у ресторана фастфуда) и крупного города (Будапешт, пойманы у автобусных остановок).
«Сельские» особи значимо отличаются от «городских» структурой поведенческого синдрома, а количественно — степенью интегрированности разных черт индивидуальности в осуществление поведения, по которому их тестировали. У первых он составляется неофобией в отношении объекта (0,8) и корма (0,71), принятием риска (0,51) и общей активностью. У вторых он связал только 3 качества из 4: общую активность (вклад -0,95), неофобию в отношении объектов (0,5) и принятие риска (0,35). Средние значения этих качеств и сила связей между ними значимо отличаются у птиц из разных поселений, однако нет какой-то очевидной зависимости этих различий от степени урбанизации (Bokony et al., 2012).
Именно разрушение и/или перестройка корреляционных плеяд, структурирующих поведение «сельских» или «лесных» популяций, максимизируют проявление а) ситуационной вариативности поведения «городских» птиц, б) его зависимости от контекста и в) лёгкости переключения между поведенческими альтернативами. Примеры а)-в) анализируем далее; здесь отметим, что все три трансформации растут пропорционально времени от начала урбанизации, независимо от конкретных подробностей биологии данного вида в данном городе, экологических факторов, особенно действующих на него и пр. В первые годы существования «городские» популяции почти не отличаются по этим признакам от исходных, кроме большей лабильности жизненной стратегии и следующей отсюда большей вариативности значимых черт экологии/поведения. Направленные изменения — поведенческие, морфологические, генетические, — появляются и усиливаются позже, через первые десятки лет после освоения города (Miranda, 2017; Minias et al., 2018; Sepp et al., 2020, также см.выше).
В средние века говорили «городской воздух делает свободным»: отчасти это верно для урбанизации птиц и млекопитающих10. Лабилизация жизненных стратегий «городских» птиц и нескоррелированность составляющих её форм поведения (они же черты индивидуальности или «качества» особи, оцениваемые отбором) дают им максимум 1) свободы выбора действий в проблемной ситуации, в первую очередь в плане «следования за средой» и прогноза её изменений, при 2) ослаблении видовых автоматизмов, часто гибельных в урбосреде, полной «экологических ловушек» (Фридман, Ерёмкин, 2009: 90-100).
В результате урбанизации большие смелость, неофилия, интенсивность исследования нового из отличия «лучших», выигрывающих конкуренцию, делаются характеристикой всей популяции. Лишь первые особи в «сельских» и «лесных» популяциях вида отвечают исследованием новых и необычных объектов (либо территорий), все другие «автоматически» беспокоятся и слетают, в «городских» первый способ ответа проявлен у всех. Его выраженность сильно варьирует между видами и усиливается со временем от начала урбанизации, однако все «городские» популяции по этому признаку много однородней исходных. Всеобщая смелость «городских» птиц ещё сильней увеличивает вариативность ответа на встречу с новым и необычным объектом и/или переключаемость особей между двумя вариантами реагирования, объясняя значительное — и пороговое — уменьшение средней дистанции взлёта в городах независимо от изменений реальной опасности (Tättle et al., 2018, 2019; Gotanda et al., 2020).
Неслучайно освоение урболандшафта и других экстремальных сред обитания — более северных частей ареала и океанических островов — увеличивает относительную массу мозга сравнительно с близкими формами и исходными популяциями. Их экстремальность во многом связана с повышенной переменностью условий существования в пространстве (гетерогенность) и времени (нестабильность, изменчивость «кружева» биотопов, прочих условий существования), абсолютной или относительно оптимума видового ареала. Последняя «движет» эволюцию в сторону относительно большего мозга у птиц (Sayol et al., 2016, 2018, 2020), а также обеспечивает усложнение социального поведения и «культурных традиций» у шимпанзе: оно пропорционально разнородности ландшафта, сезонности климата, в первую очередь дождей и дальности группировок от плейстоценовых лесных рефугиумов (Kalan et al., 2020).
В самих «городских» популяциях распределение мозгов по размеру обладает значимо меньшим эксцессом, чем в исходных — при общем его увеличении у части видов11 (Dale et al., 2015, цит.по: Фридман и др., 2016). С другой стороны, при урбанизации растёт разнообразие как индивидуальностей, так и форм их реагирования на события ближайшего окружения, будь то новые, потенциально опасные, или видотипичные — корм, хищники, сигналы от партнёров и конкурентов (часто также в нетипичном контексте, скажем сильней искажённые шумом, см.данные по двум типам песен, используемых в дальней и ближней коммуникации домовым крапивником, Troglodytes aedon, Grabarczyk, Gill, 2020). Следовательно, в «городских» популяциях «больше равенства» между особями как в выраженности поведения, усиливаемого отбором при урбанизации, прежде всего смелости, исследования нового и двигательной активности, так и в интегральных характеристиках («качество» особи, оцениваемое отбором).
4. Дистанция взлёта — маркёр резистентности к беспокойству, а не ответа на опасность
В третьих, уменьшение дистанции взлёта в урбанизированных популяциях вызвано отнюдь не ростом «доверчивости», но лучшей дифференциацией объектов и ситуаций. Чем больше времени прошло от начала урбанизации, тем лучше «городские» особи отделяют реальную опасность от «просто беспокойства». В т.ч. пропорционально ему растёт дисперсия дистанции взлёта, уменьшившаяся в начале процесса сравнительно с исходной популяцией (Møller, 2010a). При полной сохранности реакции бегства в первом случае (иногда она даже усилена), растёт безразличие во втором12, с лучшим переключением между ними при следовании друг за другом в потоке событий.

Мексиканские чечевичники в Нью-Йорке (где они интродуценты, как везде на востоке США)
См. данные о реакции «городских» больших синиц на три разных вида потенциальной опасности: знакомых и незнакомых людей у гнезда, а также перепелятника (Vincze et al., 2019); то же показано для обыкновенных белок Sciurus vulgaris (Uchida et al., 2018); в ответ на незнакомый шум (крик кита или гудок корабля) «городские» мексиканские чечевичники Haemorhous mexicanus быстрей подходили к кормушке и интенсивно питались, чем «сельские» и «пригородные». Гудок корабля также активизировал перемещения у «городских» и «пригородных», но не «сельских» птиц. Такой разницы не было в ответ на прочие типы новизны: новые структура среды, корм и объект (Weaver et al., 2019). В исходных популяциях, наоборот, оба аспекта нового и неизвестного неразличимы и равно вызывают тревожно-оборонительную реакцию.
Так, у разных видов оказывается, что меньшая дистанция взлёта не увеличивает успех бегства от потенциальной опасности, отчасти и наоборот. Настороженность (поднятие головы при кормлении и осмотр вокруг) считается эффективным средством обнаружения хищника и снижения риска гибели у тех, кто чаще осматривается, почему частота этого поведения должна отражать уровень страха перед ним. Тем более что эти реакции «не бесплатны», приходится прерывать кормление. По этой причине уменьшение дистанции взлёта «городских» птиц разных видов для отождествляющих урбанизацию и доместикацию — маркёр перехода в восприятии людей, собак, пешеходов, велосипедистов и пр. объектов от «страха» к «доверчивости».
Однако исследование Kunter Tätte et al. (2019) показывает, что это неверно. Более бдительные особи позже замечают опасность, хуже оценивают риск и позже спасаются, т.е. более уязвимы для хищника. Авторы предварительно оценивали бдительность у разных особей по % времени, проведённому с поднятой головой, а затем тестировали их реакцию на опасность, изображавшуюся подходом человека к птицам. Кроме неё, оценивали 3 последовательных стадии поведения бегства от хищника: дальность детекции (новый объект замечен, его начинают отслеживать), момент оценки его как опасного и момент слёта. Им соответствуют 3 дистанции: детекции (и настороженности), тревоги и бегства. Авторы определяли их у разных видов птиц в 3-х странах Европы с января по март 2017 г., каждый раз в двух разных ландшафтах, «сельском» и «городском», расположенных неподалёку. Общий объём данных включал 1408 наблюдений за отдельными птицами 50 разных видов, находившихся в данных местах; 733 птицы 42 видов в «городских» опытах и 675 из 52 видов в «сельской»; 35 видов наблюдали в обоих (Tätte et al., 2019).
В городах птицы более насторожены, чем вовне. Независимо от размера группы, настороженность увеличивалась с ростом дистанции обнаружения и массы тела при размещении птиц на высоких, но не низких присадах — т.е. где ожидалась меньшая значимость бдительности (Tätte et al., 2019, fig.3). Размер группы на этот параметр не влиял. Филогенетический сигнал (т.е. влияют ли родственные связи некоторых видов на близость их поведенческих характеристик) не влиял на дальность детекции, но влиял на массу тела и дистанцию бегства.
А вот бегство в городах в целом запаздывает. Больше осматривающиеся птицы раньше замечают подходящего, дольше следят за ним, но задерживаются со взлётом. Большая настороженность в обоих типах ландшафтов ведёт к меньшей дистанции взлёта, т.е. потенциально более рискованному поведению, причём в «городских» местах наблюдений коэффициент регрессии выше, чем в «сельских» (независимо от размера группы и высоты присады). Масса тела и размер стаи здесь незначимы (Tätte et al., 2019, fig.2, 4).
Поэтому бдительность «городских», отчасти и «сельских» птиц не обусловлены полностью задачами оборонительного поведения, но в значительной мере суть побочные следствия отвлечения внимания птиц «просто» беспокойством. Вклад последнего в городе значительно выше: больше вызывающих его объектов, в исходной популяции априори воспринимающихся как опасность (пешеходы, велосипедисты, машины, кормушка, могущая оказаться ловушкой и пр.), а частью и ею являющиеся (собаки, кошки и пр.). Каждый раз птица оценивает: возникший предмет беспокойства на деле опасен или им можно пренебречь? На это уходит лишнее время, тем более что оценка зависима от контекста.
Чем выше устойчивость птиц к беспокойству, тем ниже дистанция взлёта (также независимо от конкретной опасности). Они также тратят на настороженность тем меньше времени, чем лучше дифференцируют ситуацию, отделяя опасность для жизни от беспокойства. Оба параметра увеличиваются по ходу урбанизации, объясняя снижение настороженности «городских» птиц тех 35 видов, что представлены в обоих типах местностей (Tättle et al., 2019).
Большая настороженность здесь — компенсаторная реакция на более затруднённую дифференцировку, в природе это делать существенно легче, хотя и не вполне получается без «означивающих» опасность криков (т. н. сигналов-символов, англ.referential signals, Фридман, 2013), «силами» мозга и анализаторов самой птицы. Отсюда связи урбанизации с настороженностью неоднозначны: в части работ показано её уменьшение в урболандшафтах, в другой отсутствие различий. Выводы Tätte et al. (2019) разрешают противоречие: поза настороженности не связана с оценкой угрозы жизни, связываемой с подходом исследователя или иных существ, но отражает уровень беспокойства (назойливого, но неопасного), присущий данному участку среды обитания, «сельской» или «городской». Дистанция взлёта — комплексный показатель, равно зависящий от обоих причин (в первом случае взлёт — просто перемещение, максимум — зряшная трата времени, во втором — начало бегства в укрытие для спасения жизни).
Пропорция тех и других закономерно меняется на градиенте урбанизации. В городах гибель сильно снижена даже с учётом кошек, проводов, стёкол и т.д. факторов риска, почему среднеожидаемая продолжительность жизни в «городских» популяциях разных видов обычно выше, чем в «сельских» или «лесных» (Sepp et al., 2018, 2020). И наоборот, беспокоящее воздействие людей, техники, других факторов «городского метаболизма», растёт по «городскому градиенту» вверх быстрее, чем падает первое. Скорей всего, этот разрыв пропорционален размеру города, интенсивности уличной жизни и пр. Масштабы строительства, интенсивность движения людей и транспорта и пр. также коррелируют с размерами города (Фридман и др., 2016; Oueslati et al., 2015). Вне города наоборот — хищники значимая проблема, а общее беспокойство невелико (Møller, Ibáñez-Álamo, 2012; Samia et al., 2016). Оценивая эти два стрессора одним показателем — изменением дистанции взлёта в «городских»» популяциях сравнительно с исходными, важно разделить вклады того и другого в происходящее.
Kyoko M. Gotanda (2020) отделил изменения дистанции взлёта в ответ на исчезновение реальной опасности от вызванных урбанизацией и показал различия между причинами, определяющими то и другое. Он сравнил дистанции взлёта малого земляного вьюрка Geospiza fuliginosa
а) на необитаемых островах Santa Fe, Española: без завезённых хищников (домашних кошек и крыс), но с естественными — змеями, разоряющими гнёзда, болотной совой Asio flammeus и галапагосским канюком B.galapagoensis, ловящими взрослых. Последний населяет 4 острова из 8 вошедших в исследование (Santa Fe, Española, Isabela, Santa Cruz;
б) на необитаемых островах Baltra и North Seymour, успешно очищенных от кошек и крыс 13 и 8 лет назад;
в) у «сельских» vs «городских» вьюрков на островах о. Floreana, Isabela, San Cristobal, Santa Cruz, населённых обоими типами хищников. Там же возможно исследовать изменения дистанции взлёта в ряду городов разного размера (успешно освоенных). «Городские» биотопы для тестирования выделяли в собственно городе, «сельские» — на участках за несколько км от него, не посещаемых туристами.

Малые земляные вьюрки из центральной части острова Санта-Круз, самка и самец (ниже). Источник bluerock
 На двух островах вообще без завезённых хищников, но с аборигенными, дистанция взлёта значимо ниже, чем на на двух других, где завезённых хищников ликвидировали. Контринтуитивный факт: она отнюдь не упала через примерно 10 лет после их ликвидации, лишь её вариабельность выросла (Gotanda et al., 2020). Это контрастирует с данными по «возвращению» хищников через первые десятки лет их отсутствия. Там настороженность жертв падала почти до 0 (обзор см. Reznick et al., 2008), однако это первое исследование её изменения за столь короткий срок после устранения хищника. Видимо, запуск реакции бегства, хотя и прерывает кормление, не настолько времязатратен, чтобы отметаться отбором сразу, как станет ненужен.
На двух островах вообще без завезённых хищников, но с аборигенными, дистанция взлёта значимо ниже, чем на на двух других, где завезённых хищников ликвидировали. Контринтуитивный факт: она отнюдь не упала через примерно 10 лет после их ликвидации, лишь её вариабельность выросла (Gotanda et al., 2020). Это контрастирует с данными по «возвращению» хищников через первые десятки лет их отсутствия. Там настороженность жертв падала почти до 0 (обзор см. Reznick et al., 2008), однако это первое исследование её изменения за столь короткий срок после устранения хищника. Видимо, запуск реакции бегства, хотя и прерывает кормление, не настолько времязатратен, чтобы отметаться отбором сразу, как станет ненужен.
Большинство видов в «обычных» обстоятельствах существования, вне экстремальных погодных условий, кормятся достаточно эффективно, чтобы располагать избытком времени и энергии, растрачиваемыми «впустую», на разные формы активного и пассивного бездельничанья (Дольник, 1995: 170-176). В форме ложных взлётов и пр. реакций оно фиксируется даже на на миграционных остановках, где птицы кормятся наиболее интенсивно, с минимальным отвлечением на другие активности. Больше того, именно в это время настороженность птиц возрастает — как и в более плотных скоплениях (т. е. при большей близости потенциальных конкурентов), при максимальной дистанции взлёта, т. е. максимуме чувствительности к беспокойству (Beauchamp, 2016; Mikula et al., 2018). Постоянство дистанции взлёта через 8 или 13 лет после устранения хищников говорит о её передаче через социальное обучение, рост вариативности — о накоплении «ошибок» реагирования (в обе стороны), уже не корректируемых отбором (Gotanda et al., 2020).
Дистанция взлёта в «сельских» биотопах 4-х населённых островов (хотя хищники там имеются!) сравнима с её нижним пределом на о. Baltra и North Seymour. Урбанизация опускает её до ещё меньшего уровня. Снижение пропорционально людности города, в малых городах дистанция взлёта не отличается от таковой в «сельских» биотопах, в крупнейшем — значительно ниже. В том же ряду возрастает её коэффициент вариации, т. е. на дистанцию всё сильнее влияет контекст, в котором осуществляется контролируемое беспокойство (чем птица занимается и как течёт уличная жизнь вокруг, Gotanda et al., 2020, fig.1).
Что это значит? И угроза для жизни, и «просто» беспокойство птицей распознаются сначала, как новый и необычный, внезапно появившийся и резко движущийся объект (особенно если к ней или в её сторону), фиксируются взглядом и вызывают стресс. Лишь потом, на гораздо меньшей дистанции, птица дифференцирует их друг от друга (с т.ч. информирования о реальной опасности беспокойство — просто шум в канале связи). Их реакция и дистанция, с которой она вызывается, зависят от соотношения вероятностей встретить то и другое в общей выборке «стрессирующих встреч».
В сельской местности явно превалируют первые, т.ч. большая часть потенциальных опасностей «на всякий случай» отреагируются птицами как реальные. Насторожившись, они не успокаиваются, а взлетают сразу или после короткого слежения, и отлетают подальше (сравни с Kitchen et al., 2010; Lowry et al., 2013; Tättle et al., 2018). Всё же в сравнении с «дикой природой» пропорция «беспокойство vs угроза для жизни» здесь несколько увеличена в пользу первой, почему дистанция взлёта соответствует нижнему пределу таковой для островов, где хищники «были недавно». В «сельских» биотопах к ним добавляются кошки, но общая частота беспокоящих встреч с людьми, скотом, техникой и пр. в общем потоке жизни птиц возрастает гораздо сильнее. В «городских» популяциях вьюрков дистанция бегства падает ещё ниже, причём при сравнимом риске хищничества. Разрыв, вызванный урбанизацией вида, тем выше, чем крупнее город, и не связан с опасностью для птиц (Gotanda et al., 2020, fig.1, 2).
Максимум дистанции взлёта отмечен на о.Флореана в самом маленьком городке Puerto Velasco Ibarra (111 жителей): она значимо больше, чем в других городах, и не отличается от таковой вовне. У птиц города, следующего по величине, Puerto Villamil на о.Изабела (2,164 жителями) дистанция взлёта значимо уменьшается, что продолжается в более крупных городах. Т.е. её сокращение при урбанизации пороговое, начинается в городах, больших некоторого размера (Gotanda et al., 2020, tabl.3).
По мере урбанизации вьюрки осваивают среду, где беспокойство экспоненциально растёт до максимума в крупнейшем городе, а риск хищничества прежний или падает. В большинстве случаев данный конкретный город хищники осваивают позже жертв на годы-десятки лет (дербник, перепелятник — мелкие певчие птицы, тетеревятник — дрозды, вяхири Columba palumbus, пустельги, ушастые совы Asio otus и пр. Фридман, Ерёмкин, 2009, табл.2; Фридман, 2010, 2018; Фридман и др., 2016). Однако ни в одном из этих случаев не наблюдается обычного для природы катастрофического подъёма смертности при «возвращении» хищника, видимо, вследствие очень высокой гетерогенности урбосреды. Исследования систем хищник-жертва в поле и в микрокосмах показывают, что при прочих равных гибель от хищничества растёт с увеличением однородности их общей среды обитания и снижается гетерогенизацией (Фридман, Суслов, 2018).
Поэтому — как и вследствие ресурсного изобилия у видов, успешно урбанизирующихся — риск хищничества не может рассматриваться всерьёз как существенная причина в объяснениях всего спектра «городских» изменений, от поведения до элементарных эволюционных явлений, при безусловной важности интенсификации «помех» во всех каналах связи, используемых особями. Естественно ждать приспособления «городских» птиц к этому возрастанию их «зашумлённости», когда вдруг появившийся быстро движущийся объект «по умолчанию» отреагируется как общее беспокойство, на которое не обращаешь внимание в тех местах, где оно обычно случается (но не, например, при вылетах для кормления за город или посещении природных территорий).
Для «городских» птиц риск таких изменений восприятия минимален: если объект реально опасен, это обнаружится при прослеживании. Поэтому в более крупных городах у них ожидаемы
а) более долгое слежение,
б) более поздний взлёт,
в) лучшая переключаемость между «дольше не обращать внимание» и «немедля спасаться» сообразно обстоятельствам каждой встречи, состоянию особи, обстановке вокруг.
Т.е. у «городских» птиц в противоположность «лесным» ответ на «встречу» с новизной и/или потенциальной опасностью из «гарантированного» делается зависимым от контекста, что увеличивает коэффициент вариации дистанции взлёта по мере того, как она уменьшается.
Универсальность а)-б) общеизвестна, была показана на разных видах птиц (Samia et al., 2016; del Barco-Trillo, 2018; Tätte et al., 2018, 2019). В т.ч. дистанция взлёта G.fuliginosa в городах падает до уровня, многажды меньшего, чем отмеченная при «естественном» риске хищничества, или чем в «сельских» биотопах того же острова, где риск хищничества сравним с «городским». По той же причине в ряду городов она наибольшая в самом малом городе: там психологический эффект общего беспокойства «суммируется» с восприятием риска хищничества, а не «вычитается» из него (Gotanda et al., 2020, рис.2).
Действительно, беспокоящее воздействие шума никуда не девается: не представляя опасности, он отвлекает внимание «городских» птиц, определяя рост настороженности вдоль градиента урбанизации. Дистанция взлёта «городских» домовых воробьёв снижена сравнительно с «сельскими», но те и другие после отлова одинаково быстро прячутся, будучи побеспокоенные людьми на новом месте жительства. Время пребывания в убежище после беспокойства у тех и других падало в течение 8 последовательных испытаний, однако «городские» воробьи восстанавливались быстрее (Vincze et al., 2016).
Дополнительное проигрывание шума 6 видам птиц (лазоревке P.caeruleus, зарянке Erithacus rubecula, большой синице, зяблику Fringilla coelebs, московке Parus ater и поползню Sitta europaea) в 46 разных точках последнего снизило частоту посещений кормушек, интенсивность кормления там и повысило уровень настороженности всех видов. Большая урбанизованность территории снижала частоту посещения кормушек и интенсивность кормления «в ту же сторону», что и проигрывание шума, но значимо не влияла на уровень настороженности. Одинаковость поведенческого ответа на шум данных 6 видов контрастирует с разной степенью их урбанизации в Англии, опровергая предположение, что большая чувствительность к шуму и/или большая настороженность, более дальнее обнаружение реальной опасности блокируют урбанизацию (Merrall, Evans, 2020).

Урбанизировавшиеся зарянки в Великобритании
Действительно, более крупные и более «робкие» виды, где настороженность исходно повышена, урбанизируются не хуже прочих. При этом их дистанция взлёта в относительном выражении сокращается намного сильней. Парадоксально, но именно у них, кажущихся урбофобами, изменения, созданные урбанизацией, масштабней и разносторонней сравнительно с урбофилами. В отличие от вторых при освоении урбосреды их риск гибели не снижается, а скорее растёт (с учётом большей вероятности ударов о провода, стекла и пр.), т. е. «необходимая» настороженность также увеличивается (данные метаанализа, del Barco-Trillo, 2018). Что с неожиданной стороны подтверждает вывод об отсутствии принципиальных урбофобов и возможности урбанизации для всех видов региональной авифауны, пусть с разной скоростью и в разных экологических обстоятельствах (Фридман и др., 2016; Фридман, 2018).
Другой вывод — уменьшение дистанции взлёта связано не с оценкой опасности и своевременным бегством от неё, а с другими процессами, косвенно воздействующими и на данный показатель. Aнализ Tättle et al. (2018) показал, что реакция на опасность выражена другим показателем — дальностью слёта (англ. distance fled) в ответ на воздействие. Чем она выше, тем сильнее последнее воспринимается как угроза жизни и выше страх перед ней. Способ бегства и дистанция до ближайших убежищ — главные предикторы дистанции слёта (оценка для 699 птиц 17 разных видов в «городских» vs «сельских» биотопах Эстонии). Дистанция взлёта, часто используемая как мера опасности, от них не зависит вовсе, но значимо возрастает с размером стаи (Tättle et al., 2018).

Дистанция взлёта и дальность слёта при вспугивании положительно коррелируют для крупнейших видов, но не связаны у видов поменьше (отражая многажды большую «цену» самых энергозатратных реакций — полёта, Дольник, 1995). Урбанизация уменьшает как дальность слёта, так и его частоту (в ответ на одно и то же воздействие), отражая большую точность оценки риска «городскими» птицами. Что подтверждается положительной корреляцией между дистанциями начала отслеживания и взлёта лишь в «сельских», но не «городских» биотопах. Уменьшение медианы дальности слёта, вызванное урбанизацией, сравнимо с таковым для медианы дистанции взлёта (Tättle et al., 2018).
Изменения поведения, вызванные шумом, в урбосреде менее выражены, а иногда обращаются в противоположность. У черноголовых гаичек P.atricapillus проигрывание шума также снижает готовность посещать кормушки, однако последняя «вверх» по городскому градиенту растёт вместе с «базовым» уровнем шума. Городские особи лучше дифференцируют последний от «острых» подъёмов зашумлённости, и реагируют только на них (Van Donselaar et al., 2018, цит.по Merrall, Evans, 2020).
Сходным образом действуют пешеходы, машины и т. д. раздражители. По завершению урбанизации вида данные изменения скорей «во благо»: большая сложность урбосреды в сочетании с более интенсивным слежением за динамикой обстановки вокруг толкает больше исследовать окружение и при появлении нового, неизвестного использовать в своих интересах — вместо страха и бегства. И наоборот, меньшая эффективность кормления в этом случае не критична, из-за большей ресурсообеспеченности используемых «городских» биотопов. Антропогенный шум «маскирует» видовые сигналы тревоги, из-за чего северные кардиналы и прекрасные расписные малюры Malurus cyaneus в ответ на них не проявляют реакций избегания хищника, однако это не ведёт к значимому снижению выживаемости (Grade, Sieving, 2016; Zhou et al., 2020; цит.по: Merrall, Evans, 2020).

Горная гаичка
Подтверждаются выводы Tattle et al. (2019), что обвальное уменьшение дистанции взлёта в процессе урбанизации — следствие одновременного роста резистентности к общему беспокойству и способности отделять его от реальной опасности. Реакция на первое у «городских» птиц прежняя, а то и усиленная, что обсуждается ниже; на второе урбанизация вырабатывает безразличие или даже позитивное отношение (приближение, активное исследование и т. д.). Так, сорочий жаворонок Grallina cyanoleuca13 успешно освоил Мельбурн (Австралия), дистанция взлёта «городских» птиц упала в 2,5 раза (12 ± 1 vs 35 ± 3 m). «Сельские» птицы чувствительней к беспокойству: вспугнутые людьми, собаками, машинами, птицами, громкими звуками, они чаще взлетают (100% vs 57,6%) и отлетают на большее расстояние. Падение дистанции взлёта сугубо контекстуально: вблизи а/д она в 1,9 раза ниже, чем вдали от них (7 ± 0,6 м vs 13 ± 2 м). Однако не найдено значимого влияния на неё присутствия людей, собак или конспецификов, а также дистанции, с которой стартует приближающийся человек, т.е. факторов важных именно для оценки опасности (Kitchen et al., 2010, table 1).
Люди чаще вспугивали птиц вблизи а/д и пешеходных тропинок, чем вдали от них (55% и 20%), что отражает способность птиц дифференцировать разные участки урбосреды по уровню беспокойства и вести себя соответственно. Однако затраты времени на настороженность (24,1%) и кормление (65.5–69.4%) значимо не отличались на обоих участках, как и процент вспугнутых птиц (86% и 75%), отражая стрессированность «городской» популяции. На долю обоих активностей в бюджете времени значимо влияют пол птиц и присутствие конспецификов (увеличивает настороженность), но не собственно беспокоящие воздействия (Kitchen et al., 2010, table 2).

Дальше увидим, что этот стресс средовой, существующий долговременно, а не вызванный конкретным объектом и/или ситуацией, за которыми идут наблюдения в подобных работах. Все эти данные опровергают расхожее мнение, что урбанизация «диких» видов птиц рождает «доверчивость» к тому, что прежде было опасностью.
У G.fuliginosa дистанции начала отслеживания [потенциальной опасности] и взлёта в общем случае не коррелируют между собой (если взлёт — реакция лишь на опасность, корреляция должна быть положительной). На о.Сан-Кристобаль она даже отрицательна. Другой интересный момент — независимо от величин дистанции взлёта, на всех островах она увеличивается пропорционально размеру группы, а не падает (Gotanda et al., 2020), как можно было бы ожидать, будь она ответом на реальную опасность (в больших скоплениях риск для его членов в целом снижается, Фридман, 2016). Это верно также для разных видов птиц Европы, у которых оценивали изменения дистанции взлёта в парках и на кладбищах: она падает пропорционально плотности пешеходов, но растёт пропорционально скорости их движения (почему везде ниже на кладбищах, чем в парках), размеру «зелёного острова» внутри города и величине группы особей, к которым происходит подход (Mikula, 2014; Morelli et al., 2018), но почти не зависит от величины парка, «естественности» растительности и пр.
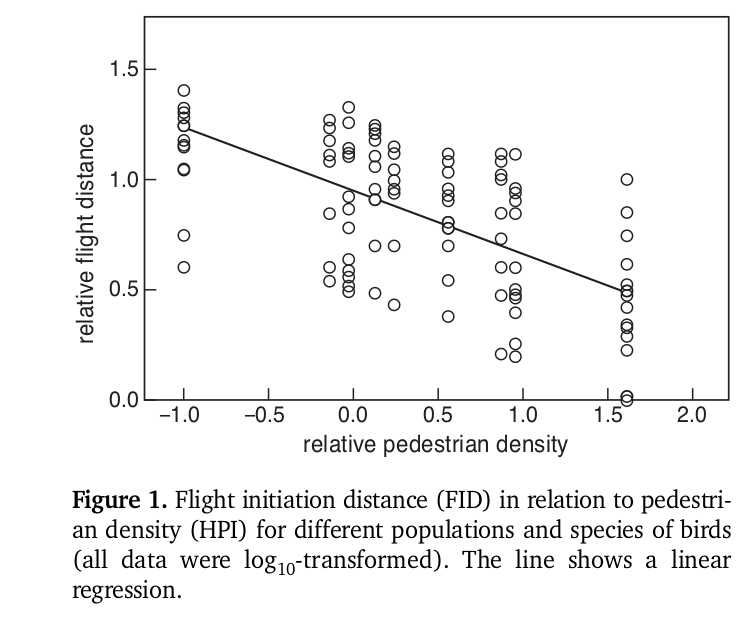
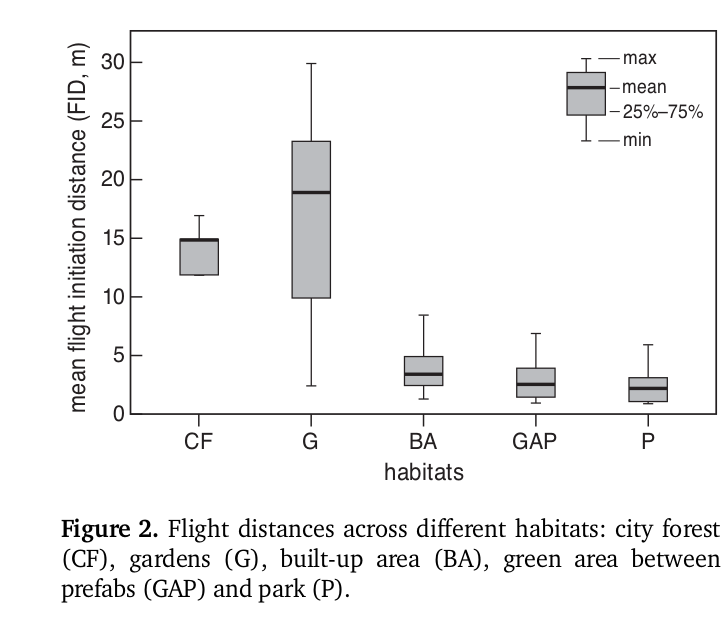
Дистанция взлёта снижается пропорционально интенсивности пешеходного потока (данные по 20 видам птиц из 10 пражский парков). Второй рисунок — изменения дистанции взлёта (показаны среднее, первый и третий квартиль) в разных типов городских местообитаний: слева направо — городские леса, сады, застроенные территории, участки озеленения между домами каркасно-модульного типа (т. н. prefab house), парки. Источник: Mikula, 2014.
 Т.е. эта зависимость — простое следствие взаимного возбуждения птиц в стайке: настороженность одной снижает порог реагирования другой и т. д., почему они вместе взлетают раньше. По этому механизму в скоплениях распространяются волны самовозбуждения, ведущие к «ложным паникам», вроде обычных для кормящихся на отмели куликов, лебедей и других птиц; регулярное появление реальных причин для взлёта просто запускает эту реакцию раньше (Фридман, 2016).
Т.е. эта зависимость — простое следствие взаимного возбуждения птиц в стайке: настороженность одной снижает порог реагирования другой и т. д., почему они вместе взлетают раньше. По этому механизму в скоплениях распространяются волны самовозбуждения, ведущие к «ложным паникам», вроде обычных для кормящихся на отмели куликов, лебедей и других птиц; регулярное появление реальных причин для взлёта просто запускает эту реакцию раньше (Фридман, 2016).
Отсюда понятно, каким образом урбосреда является «развивающей», в смысле прогресса когнитивного поведения «городских» особей сравнительно с «лесными», кроме прямого позитивного влияния на них и на увеличение мозга более гетерогенной, изменчивой, и (главное) более экстремальной среды обитания14 в городе. Видам, которые уже освоили город, он предоставляет много больше ресурсов, чем в исходных сообществах; а это оставляет птицам много больше свободного времени, чем в исходных местообитаниях, заполняемого «активным бездельничаньем». Этот термин введён В.Р.Дольником (1995) для обозначения деятельности, единственная задача которой — «сбросить» энергию индивида через разные формы активности, в случайном порядке «перебираемые» животным в игровой, нефункциональной манере. Внешне они мало отличаются от «настоящей» кормёжки, запасания, агрессии, патрулирования территории и пр. действий, но употребляются «не ради результата». Так саморегулируется суточный бюджет энергии, «приход» совмещается с «расходом» (Дольник, 1995: 173).
«Активное бездельничанье» — ещё и «пересмотр» отношения к новым и непривычным объектам15, параллельный тому, что уже происходит в «городских» популяциях (интерес, приближение и исследование вместо настороженности и «автоматического» бегства либо отслеживания в напряжении, преобладающих до урбанизации). Т.е. урбосреда в силу сложности, гетерогенности и изменчивости её компонентов предоставляет возможность для прогресса исследовательской и рассудочной деятельности, и одновременно «подталкивает» «городских» птиц к использованию составляющих её объектов, ситуаций и процессов именно в этом качестве. Поэтому она развивающая «даже дважды».
Наконец, «городские» кроличьи сычи при значительно меньшей дистанции взлёта имеют такой же уровень кортикостерона, как сельские. У отдельных индивидов также нет связи между его содержанием (в перьях, т. е. долговременным) и величинами дистанции взлёта в конкретный момент. Не связана с ним и выживаемость отдельных птиц в городе, повышенная вдвое сравнительно с «сельскими» сычами (Rebolo-Ifrán et al., 2015), т. е. по завершению урбанизации конкретные беспокоящие объекты или события в городе всё же не действуют как дополнительный16 стрессор — ни в каждый отдельный момент, ни долговременно, почему урбосреда и может быть развивающей.
На мексиканских чечевичниках также показано, что присутствие человека стрессирует «сельских» птиц значимо сильнее, чем «городских». Реакция тех и других на хищника была одинаковой, также как разница в реагировании на модель яcтреба vs модель горлицы (Weaver et al., 2018), т. е. урбосреда не меняет реакцию «городских» птиц на опасность. Больше того, регулярное беспокойство человеком снижало способность к решению задач на новый способ кормодобывания у «сельских», но не городских чечевичников, выращенных в лаборатории. Птицы должны были сдвинуть клювом скользящую крышку с приоткрытой кормушки слева направо, чтобы получить доступ к семенам подсолнечника. Лучший предиктор решения задачи — долгий подробный осмотр птицей проблемного поля перед собственно действием. Он подавляется беспокойством у «сельских», но не «городских» птиц (Cook et al., 2017).
5. Осторожность городских птиц не снижается, но делается контекстуальней
В большей или меньшей степени, но рост переключаемости и зависимости от контекста при урбанизации захватывает все черты индивидуального поведения, изученные в сравнении с исходными популяциями. У одних и тех же видов на более урбанизированных территориях дистанция взлёта значимо ниже, но больше зависит от ситуации: типа растительности, интенсивности посещения, способа передвижения — пешеходы, велосипедисты и пр., а главное, активности интереса к птицам (Møller, 2008, 2010а, 2014; Mikula, 2014; Blumstein, 2014). При прочих равных больший уровень шума укорачивает дистанцию взлёта, т. к. в этих условиях люди могут подойти ближе (Petrelli et al., 2017). У «городских» (но не сельских) кроличьих сычей дистанция взлёта значимо укорачивается на протяжении репродуктивного периода; будучи потревоженными, они быстрей возвращаются к гнезду, хотя и не агрессивней в его защите, т. е. быстрей восстанавливаются после стресса (Cavalli et al., 2016).
Другой пример: по сравнению с сиэтльцами берлинцы значимо больше заботятся о птицах (ставя кормушки, развешивая гнездовья) и меньше их беспокоят (активно отпугивая и преследуя). Это различие сохраняется на всём протяжении городского градиента (Clucas, Marzluff, 2012, fig.2-3), определяя большую дистанцию взлёта в «сельских» популяциях обыкновенного скворца Sturnus vulgaris и американской вороны C.brachyrhynchos вокруг Сиэтла, чем скворца и серой вороны вокруг Берлина. При движении по городскому градиенту «вверх» это различие уменьшается (как и абсолютные величины дистанций взлёта), и делается незначимым в собственно городе у ворон, но не у скворцов. Домовый воробей в Берлине и странствующий дрозд Turdus migratorius в Сиэтле устойчивы к беспокойству со стороны человека, дистанция взлёта у них минимальна, и почти не меняется в зависимости от степени урбанизации или беспокойства птиц человеком. При одних и тех же уровнях последнего средняя дистанция взлёта скворцов и врановых вокруг Сиэтла выше, чем вокруг Берлина (Clucas, Marzluff, 2012). Видно, что падение дистанции взлёта «городских» птиц отражает отнюдь не опасность, следующую из появления данного человека здесь и сейчас, но отношение к птицам, сложившееся в культуре горожан. Можно предположить, что при изменениях этой последней дистанция взлёта (и оборонительное поведение в целом) также будут меняться, и это предположение оправдывается, см.ниже.
То же видим в контактах с человеком и техникой у видов, освоивших «островные» биотопы в других антропогенно изменённых ландшафтах. Так, черный коршун, тетеревятник, перепелятник, канюк B.buteo, осоед Pernis apivorus, орел-карлик Hieraaetus pennatus освоили лесные микрофрагменты на Верхнем Дону. Там они достигают значительно большей плотности, чем в крупных лесных массивах (Фридман и др., 2016, рис.8), одновременно меняя ответ на направленный интерес наблюдателя. В лесах хищники обнаруживали его за 50-100 м от гнезда и беспокоились, с криками летая над гнездом. В «островных» биотопах реакция противоположна: даже при прохождении в 10-20 м от гнезда они затаиваются, вжимаясь в леток и внимательно наблюдая за людьми, чтобы немедля и бесшумно исчезнуть при их остановке. Возможен также отлёт заранее, но не поведение, видимое издалека. Это реакция именно на направленный интерес к птицам и их гнездам. Гнездящимся парам вполне безразличны едущие мимо машины, люди и техника на с/хработах, их голоса, сходное с выстрелом щёлканье кнута. Настороженность возникала немедля с появлением тихих людей с биноклями, их направление на гнездо заставляло птиц вжаться в лоток и пр. (Галушин и др., 2015).
Другой пример — гуси Anser spp. считаются одними из самых осторожных птиц, а охота на них — одной из самых сложных. В пойме р.Унжи напротив г.Кологрива (Костромская обл.) усилиями охотоведа А.А.Васечкина с 1989 г. она запрещена. В последующие годы сформировалась многотысячная остановка гусей на весеннем пролёте, исключительно толерантных к приближению наблюдателя, (если оно постепенно, с остановками при появлении признаков беспокойства в стае). Они быстро успокаиваются даже при направленном беспокойстве (чтобы прочитать шейные кольца, или когда пойманные птицы бьются в сети). Однако они не стали «доверчивей»: легко отделяют наблюдателей от смертельно опасных охотников, и сверхосторожны по отношению к ним, как и к машинам того типа, из которых раздавались выстрелы — при безразличии к тракторам и т. д. с/хтехнике (Галушин и др., 2015).
Это верно и для оборонительного поведения. Как минимум до успешного завершения урбанизации (включая третий этап экспоненциального роста городской популяции, Evans et al., 2010) «городские» птицы весьма чувствительны даже к простому беспокойству людьми, техникой и другими агентами городской жизни, хотя позже выработают резистентность. Часто наоборот, в это время они более насторожены и избегают беспокойства не менее чем реальных опасностей, стремление оставаться «невидимыми» определяет пространство и время их активности в городе. Общее снижение настороженности (включая резкое сокращение дистанций взлёта) сильно запаздывает, отмечаясь тогда, когда в экологии «городских» особей уже мало что изменяется.
Успешное освоение урболандшафта совершается при повышенной осторожности17: птицы, быстро заселяющие природные территории города и даже небольшие «зелёные острова» среди застройки, ведут себя максимально незаметно, так что почти не попадаются на глаза. Это хорошо согласуется с подъёмом уровня гормона стресса — кортизола в начале урбанизации «диких» видов птиц, как на другие сдвиги видотипичного «кружева» биотопов — пятна нарушений, фрагментацию извне, инсуляризацию изнутри и пр. (Bonier, 2012, table 2). Лишь постепенно всё большая часть объектов и ситуаций, «возникающих» перед особью в урбосреде, вызывает всё более равнодушное отношение, не имеющее ничего общего с позитивным отношением к прежним опасностям. Именно в этот период падает средняя дистанция взлёта, увеличивается её вариация, но падает повторяемость у одних и тех же птиц.
Так ведут себя многие успешно урбанизирующиеся виды (точнее, их динамичные популяции) — европейский тювик А.brevipes на Ставрополье, орёл-могильник Aquila heliaca18 в Волгоградской обл., белоспинный дятел в лесопарках Москвы (Фридман и др., 2016). Успешная урбанизация кустарниковых большеногов Alectura lathami в пригородах Сиднея не снизила их дистанции взлёта на «островах» городских ООПТ, с естественной растительностью. Она понижается (несильно, на уровне тенденции) при поселении в более гемеробных ландшафтах (сады, улицы, лужайки) — лучше просматриваемых, с большим уровнем беспокойства, но сравнимых по реальной опасности. Коэффициент вариации у дистанций взлёта «городских» птиц также растёт, показывая сугубую ситуативность их реагирования. Дистанция настораживания при переходе в урболандшафт снижается много медленней, т. е. они не «доверчивей» к потенциальной опасности (Hall et al., 2020).
Так, чёрные вороны C. corone orientalis при быстром освоении г.Иркутска (1967-1979 гг.) оставались предельно осторожными. Как при сохранении G.fuliginosa прежней дистанции бегства на необитаемых островах, очищенных от хищника ~10 лет назад (Gotanda, 2020), здесь сохранялась «инерция» реальной опасности. В 1950-е годы окрестности Иркутска бесконтрольно использовались для охоты. Вороны строго выдерживали дистанцию выстрела и слыли крайне осторожными птицами. Хотя они и гнездились в пригородах Иркутска, но чётко тяготели к окрестным сёлам. Большое количество ворон ежегодно зимовало вокруг Иркутска. Они кормились на городской свалке, но на ночёвку улетали в пойменные ельники в 5-10 км вверх по р.Иркут. В центре города птицы не останавливались, преодолевая его только по воздуху. Весной в предгнездовое время летящие вороны даже окрикивали прохожих на городских улицах — как и других хищников.

Чёрные вороны в Иркутске, Источник: «Иркутск глазами птиц, птицы глазами Иркутска»
Даже поселившись в городе, вороны освоили лишь крыши многоэтажных домов и в днём не рисковали спускаться вниз. Они добывали корм в ранние утренние часы, успевая за столь ограниченное время даже запасти его впрок на крышах. Лишь в 1979 году, когда популяция превысила 100 гнездящихся пар, некоторые вороны спускались и днём. Уже через год они появились в скверах, на пешеходных дорожках парков, во дворах и т.п. Дистанция вспугивания упала до 5-10 м. К началу 1980-х гг. вороны в поисках корма совершали патрульные полёты в скоплениях людей, хватали добычу среди публики. Тогда же они утеряли осторожность на гнёздах и гнездовых участках: открыто таскали строительный материал, демонстрировали брачное поведение, окрикивали хищников, изгоняли территориальных конкурентов, не обращая внимания на людей и машины (Липин и др., 1983, цит. по: Фридман и др., 2009).
Помимо урбанизации, «инерционное» сохранение прежней настороженности всё время освоения новых местообитаний (часто и далее) фиксируется и в других «рукотворных» ландшафтах. Так, остались сторожкими клинтухи C.oenas, в 2010-х гг.19 начавшие гнездиться на ЛЭП и держащиеся в с/хландшафте: быстрый рост численности птиц этого экотипа, независимо появившегося в самых разных районах юга России и Украины, ведёт ко всё более частому появлению в сёлах, потом и гнездованию там. Всё это время дистанция взлёта оставалась высокой: её снижение, рост безразличия к людям отмечены лишь с 2016 г., после быстрого роста численности в новом ландшафте (Соколов, 2017).

Клинтухи, гнездящиеся в открытом ландшафте на ЛЭП. Источник Соколов, 2017.
Белолобые ласточки Petrochelidon pyrrhonota, селясь на мостах и эстакадах, сталкиваются с реальной опасностью — несущимися машинами, но эти места весьма привлекательны. Противоречие разрешилось большей резистентностью птиц к транспортному потоку (т.е. уменьшением дистанции взлёта) и одновременно микроэволюционными изменениями: укорочением крыльев, делающим их маневренней. Т.е. к опасности они не стали доверчивей, но лучше «уворачиваются» от неё, почему могут подпустить ближе. Действительно, гибель ласточек значительно сократилась (Brown, Brown, 2013). Сокращение гибели через несколько лет после освоения микроурбанотерриторий отмечено и у скалистой ласточки Ptyonoprogne rupestris, в последние 30 лет начавшей гнездиться во Франции и Швейцарии на зданиях, в Албании – в брошенных бомбоубежищах, на северном Кавказе – на балконах многоэтажных зданий, в недостроенных или заброшенных домах, в заброшенных зданиях рудника Эльбрусского, на сводах гротов в парках г.Кисловодска, на высокогорной станции канатной дороги на хребте Мусса-Ачитара и пр. (Glutz von Blotzgeim u.a., 2012; Тельпов и др., 2012).

Белолобые ласточки собирают грязь для гнезда близ лужи
Следовательно, повышенная толерантность к людям, способность гнездиться рядом с человеческими постройками, даже «не выходя» из внутригородских «островов» лесных, луговых, заболоченных и т.д. биотопов отражает успех урбанизации, причём начавшейся довольно давно, а не его предпосылки (Møller, Diaz, 2018b). Сравнение 50 пар близких видов показывает, что способные устойчиво гнездиться ближе к человеку, при повышенном беспокойстве, а то и опасностей, успешней урбанизируются и чаще вытесняют близкие формы, а не наоборот. Они же в городе начинают гнездиться раньше, и размножаются дольше, чем в сельских местообитаниях (Møller, Diaz, 2018a). Всё это показывает их главное конкурентное преимущество в урбосреде – большую устойчивость «городских» вариантов популяционной системы сравнительно с исходным и её опережающий рост при освоении города сравнительно с подъёмом всех видов «возмущений» (Фридман, Ерёмкин, 2009, табл.3; Фридман и др., 2016; Фридман, Суслов, 2018).
Чёрные точки – места, где скалистые ласточки стали гнездиться на зданиях, малиновый – в 1993-1996 гг., оранжевый – 1993-96 &2013-2016 гг., зелёный – 2013-2016 гг. Больше всего таких случаев в основном ареале вида в альпийском регионе, вне его это случается гораздо реже. Источник vogelwarte.ch

Гнездо скальной ласточки Ptyonoprogne rupestris на административном здании санатория «Красные камни» в Кисловодске (а, б – в 2011 году; в, г – в 2012 году). Источник Тельпов и др., 2012.
Ещё более это верно для млекопитающих (McCleery, 2010). В «городских» колониях чернохвостых луговых собачек Cynomys ludovicianus, изолированнных застройкой г.Денвера (штат Колорадо), настороженность повышена. Только со временем она снижается до уровня, меньшего, чем описан у вида в открытых травянистых ландшафтах (Magle, Angeloni, 2011). «Городские» зверьки лучше дифференцируют потенциальную опасность от беспокойства к которому привыкли. Анализ влияния разных видов человеческой деятельности на оленей вапити Cervus elaphus и вилорогов Antilocapra americana вдоль транспортного коридора через национальный парк Grand Teton (США) показал устойчивое уменьшение настороженности, вероятностей бегства вглубь массива, перемещения вдоль трассы и затаивания при усилении потока а/т и, соответственно, шума.
И наоборот, возрастала чувствительность к объектам, звуки от которых контрастировали с «фоном» шумов а/т (пешеходам и мотоциклистам), интенсифицировался ответ на них оборонительным поведением (Brown et al., 2012). Искусственное воспроизведение шума а/д увеличило бдительность C.ludovicianus и дистанцию, при которой они убегают от приближающегося человека. Эти изменения усиливались в следующих повторностях опыта, однако так луговые собачки регировали только на данного наблюдателя, а не всех прохожих (Shannon et al., 2016).
Подобные данные ещё раз показывают, что урбанизация вида не означает его синантропности. Первое может следовать из второго, но у меньшинства видов, находящего в городах аналоги исходных биотопов (каменка Oenanthe oenanthe, горихвостка-чернушка Phoenicuros ochruros, канареечный вьюрок Serinus serinus, домовый воробей, касатка Hirundo rustica и воронок, сирийский дятел D.syriacus, хохлатый жаворонок Galerida cristata, коноплянка Linaria cannabina, щегол Carduelis carduelis и пр., см. Фридман, Ерёмкин, 2009, табл.2, Фридман, Суслов, 2018, табл.2, и выше). Это прежде всего здания и сооружения, стройки, пустыри с луговыми участками разной степени вытоптанности, мозаичные участки полей, садов, строений сельского типа и т.д. элементы скал или «традиционного с/х ландшафта», сохраняющиеся в городе. И наоборот, виды-синантропы могут не быть урбанистами, обитать преимущественно или только в с/хландшафтах и сельских населённых пунктах: таковы многие воробьи, ткачики, овсянки, канареечные вьюрки и пр.
Урбанизация может вести к синантропности и когда вид из природных территорий города переходит к гнездованию прямо в застройке (с переходным этапом обитания на «зелёных островах» всё меньшей площадью и сильней изолированных селитебной зоной). Это также присуще сугубому меньшинству видов, успешно урбанизировавшихся в смысле устойчивого существования на «архипелаге» природных территорий, большей численности там, чем в местообитаниях региона и пр. Если такое случается, то не сразу по появлению жизнеспособной городской популяции, но через первые десятки лет, как у чёрного дрозда, вяхиря, сойки Garrulus glandarius и т.д. (Фридман, Ерёмкин, 2009; Evans et al., 2010).
Сущностные различия урбанизации и синантропии как приспособлений птиц к разным (лишь отчасти пересекающимся) «рукотворным ландшафтам» хорошо видно из того, что т.н. «традиционные синантропы» в современных городах сделались весьма уязвимы, а недавно урбанизировавшиеся виды – процветают, в т.ч. в «третьем» мире, где ускоренное развитие городских систем началось относительно недавно, с 1990-х гг. Из трёх крупных родов – Corvus, Turdus и Columba (каждый из >40 видов, распространённых по всему миру, и сильно различных по «позиции» на оси специалист-генералист) за этот период урбанизируются все, испытывающие «давление» новых агломераций (Evans et al., 2010).
Большая ситуативность реакции «городских» птиц проявляется в том, что одни и те же особи меняют дистанцию взлёта в зависимости от местонахождения, в одних районах города или местобитаниях внутри участка она значимо больше, чем в других, при одном и том же способе беспокойства. Исходя из собственного опыта, они «маркируют» разные элементы урболандшафта как более или менее безопасные районы, и держатся там более или менее настороженно в отношении одних и тех же объектов. Так, калининградские вяхири Columba palumbus, в городе подпускающие вплотную, как сизари, за городом на кормёжке также осторожны, как и дикие (Е.Л.Лыков, личн.сообщ.). Другой пример — чёрные дрозды в городских парках Мадрида более насторожены, меньше кормят птенцов и избегают тропинок, когда по ним движется поток отдыхающих. Они давно приспособились к фактору беспокойства, в целом став более резистентными, и именно потому «делают поправку» на его изменения внутри участка (Fernández-Juricic, Tellería 2000, цит.по: Kitchen et al., 2011).
Отсутствие общего снижения страха при урбанизации также показывают ряд контринтуитивных данных о поведении «городских» птиц. С одной стороны, более инновативные виды и более неофобны, позже показываются на кормушках при добавлении новых объектов. Однако они же быстрей показываются после беспокойства на контрольных кормушках, т. е. не «более боязливы», а лучше дифференцируют ситуацию и точней реагируют на контекст (переменная «дистанция до антропогенных источников корма», Dukatez et al., 2017). Действительно, время задержки для прилёта на кормушку с новым объектом сильно зависит от типа последнего, и варьирует между видами (опыты с 9 видами о.Барбадос, 2 из которых, различающиеся по инновативности — C.passerina и L.barbadensis — вообще к ним нечувствительны). Оно значимо различалось в зависимости от контекста (близость источника антропогенного корма: как надо полагать, признака относительной безопасности этой кормушки), для всех 3-х типов новых объектов, опять же, с учётом видовой специфики (у L.barbadensis — нет). Меньшая неофобность в данной ситуации, присущая другими видам, не зависела от инновативности и ли «сельской» vs «городской» среды в местах постановки опытов (Dukatez et al., 2017). Видим, что
а) адекватная оценка контекста присуща всем видам, это значимый фактор реагирования везде, независимо от инновативности вида (т. е. опосредованно его интеллекта), урбанизация лишь усиливает способность к этому,
б) урбанизация не меняет страх перед потенциальной опасностью — это скорей видовая характеристика, причём он усилен у видов а) более «сообразительных» — более инновативных, с большей относительной массой мозга (Greggor et al., 2016; Dukatez et al., 2017) и б) давно синантропных, особенно приспособившихся к постоянному контакту с человеком ещё в сельских поселениях, т. е. до урбанизации (домовый воробей, врановые, трупиалы). В ряде случаев урбанизация неофобию даже увеличивает.
Страх перед новым объектом у врановых (как и у домовых воробьёв, мексиканских чечевичников или блестящих воловьих птиц Molothrus bohariensis) гораздо выше, чем у других птиц; у них же сильней индивидуальные вариации в его проявлении, особенно в отношении пищи (Brown, Jones, 2016; Stanback, Burke, 2020). Что сопрягается с лучшей дифференцировкой подобных объектов и ситуаций, в которых к ним приближаются, именно у врановых птиц в сравнении с «не-врановыми» (а не наоборот, как можно думать).
Опыты A.L.Greggor et al. (2016) с подстановкой кормовых столиков с арахисом, в угол которых крепили разные типы объектов
а) явно искусственный,
б) подобный куску городского мусора,
в) подобный деревяшке или иной природной вещи
показывают, что не-врановые их не дифференцируют, вероятность избегания, интенсивность посещений и частота кормления в зависимости от а)-в) у них не меняются. Тестировали 12 урбанизированных видов птиц, 5 из них — врановые (галка, сойка, грач, серая ворона, сорока, прочие — лазоревка, большая синица, зарянка, чёрный дрозд, вяхирь и зяблик), в их среде обитания: сельских и городских местностях востока и юго-запада Англии. Врановые более неофобны в отношении всех трёх: значимо дольше избегают, реже подходят к столикам, много менее интенсивно питаются, чем не-врановые (Greggor et al., 2016, fig.4-5). В противоположность последним, врановые дифференциально реагируют на объекты: сокращение посещения и питания менее всего выражено в случае б), несколько больше — в), максимально в а) (Greggor et al., 2016, fig.2). Наконец, именно у «городских», но не «сельских» птиц (равным образом врановых или нет) избегание столиков с объектом, подобным городскому мусору, значимо быстрей сокращается во времени, чем с другими объектами, т. е. урбанизация делает относительным страх перед теми из них, что скорее всего попадались птицам ранее (Greggor et al., 2016, fig.3).
Ещё важней эти корректировки для урбанизированных видов млекопитающих, больше боящихся человека, чем собак и других хищников, в т.ч. потому что охотники убивают свою добычу в 14 раз чаще, чем естественные враги тех же видов за сравнимое число попыток. Люди также убивают самых здоровых и взрослых особей, своего рода «репродуктивный капитал» популяции, а не ослабленных и стрессированных, как естественные враги и даже домашние собаки. Среди сайгаков Saiga tatarica, пойманных борзыми, все особи имели те или иные патологии, среди отстрелянных охотоведами именно с целью выбраковки больных — лишь 33%. То же касается пойманных ими зайцев в сравнении с отстрелянными охотниками (Darimont et al., 2015; Северцов, Шубкина, 2019).
Урбанизируясь, разные виды млекопитающих, независимо от размеров и величин территорий, меняют использование пространства и/или временные режимы активности так, чтобы минимизировать встречи с людьми, в т.ч. реже пересекать дорожки и другие места, где возможен контакт с отдыхающими и их собаками, выходить на поляны, к домам и т.д. Они посещаются ночью или в другие часы с наименьшей вероятностью контакта. Пандемия коронавируса 2020 г. стала естественным экспериментом, показавшим много большую важность «ландшафтов страха», созданных человеческой активностью (о них — Zanette, Clinchy, 2019), для «городских» популяций млекопитающих сравнительно с сельскими.
Карантин и другие противоэпидемические мероприятия сильно снизили посещаемость всех мозаик урболандшафта, от природных территорий до кварталов застройки. Обитающие здесь виды стали активны в местах, которых при наличии там людей избегали, и в часы, когда раньше прохожие их там не видели. Причём это не недавно вселившиеся, а местные особи, их поведение отражает прекрасное знание территории, просто в обычном режиме «городского метаболизма» они были «невидимы» обывателям (Manenti et al., 2020) — как европейский тювик в Предкавказье и другие успешные синурбанисты.
Наконец, большая толерантность «городских» птиц к потенциальным опасностям отнюдь не снижает способность оценивать риск, связанный с приближением к ним и использованием. Она сохраняется в полной мере, иногда даже усиливается — как и способность дифференцировать реальные угрозы жизни от общего беспокойства. Скажем, при посещении новых кормушек учитывать наличие рядом с ними, с одной стороны, кошек, с другой — людей и собак, задерживаясь в первом случае и не реагируя во втором. Так, Tryjanowki et al. (2015) в 8 разных польских городах (и соответствующих пунктах округи) исследовали время, проходящее до посещения разными видами птиц новых кормушек в зависимости от ряда параметров окружения.
Всего выполнено 138 опытов по их постановке (80 в урбосреде, 58 вне города), на них отслеживали появление 43 видов птиц, из них наиболее обычны 9 — большая синица, лазоревка, болотная гаичка Parus palustris, сойка, зарянка, домовый воробей, зеленушка Chloris chloris, поползень, чёрный дрозд (в порядке падения вероятности посещения новой кормушки первыми, Tryjanowski et al., 2015, table 1, fig.1). Общая частота посещения ими новых кормушек значимо отличается, и больше отражает численность в месте опыта, хотя также и неофобию как оценку ситуации в плане возможной опасности. Меньшее время задержки показывает большую готовность принимать риск (следующий из появления новых объектов) и исследовать их вместо бегства, т. е. степень неофилии, большее — соответственно, неофобии. Межвидовые различия по последней вполне сохраняются — домовые воробьи и чёрные дрозды появляются на новых кормушках позже не только больших синиц и лазоревок, но и соек с болотными гаичками. Как и межполовые различия — самцы больших синиц отличаются большей готовностью к принятию риска (а не избеганию) вместо самок, и именно они значимо чаще бывают на новых кормушках, особенно первыми (Tryjanowski et al., 2015).
С другой стороны, урбанизация значима: все виды быстрей появляются на новых кормушках в городе, чем вовне. Однако не менее значимо окружение: наличие других кормушек вокруг сокращает время задержки до посещения новых, присутствие кошек — наоборот, удлиняет, а вот пешеходов или собак — вовсе незначимо. Отмечена тенденция к сокращению латентного периода в присутствии других птиц, на грани значимости (Р=0,057, table 1 работы) (Tryjanowski et al., 2015).
Появление на знакомых кормушках также зависит от окружения и меняется при добавлении к ним новых объектов (ярко-зелёный шар из резины с пучком волос, данные по 14 польским городам, Tryjanowski et al., 2016). В городских биотопах посещаемость их разными видами (особенно самым частым, большой синицей) не зависит от их численности и температуры. Т.е. даже в условиях, способствующих проявлению страха, кормление там — следствие не случайного обнаружения в общем поиске, но знание расположения «местных» кормушек и направленное их использование. Присутствие новых объектов сильней сокращало число «городских», нежели «сельских» больших синиц; тот же эффект выявлен для других видов-посетителей, но слабее. Наличие других видов птиц вокруг увеличивало число посетителей, также как и число кошек и (намного слабей) пешеходов; напротив, число собак негативно влияло на посещаемость. Эти зависимости выражены сильней в городах, чем вовне (Tryjanowski et al., 2016).
В присутствии реальной опасности (кошек) кормушка, даже с новым объектом, активней используется как безопасный источник корма (вместо ожидаемого испуга): напротив, собаки, беспокоящие птиц, заметно снижают её ценность. Урбанизация, по всей видимости, почти не влияет на уровень неофобии (сравни с данными по 9 барбадосским видам, Dukatez et al., 2017), но явно улучшает оценку риска и одновременно повышает готовность рисковать, когда «есть смысл». Обе работы снова показывают, что «городские» особи (в отличие от одомашненных) не делаются «доверчивей» к новым объектам.
Заключение
Резюмируя, в изменениях при одомашнивании восприятие опасности и беспокойства амальгамированы и вместе меняются в сторону сперва уменьшения, потом — обращения на 1800. При урбанизации, наоборот, восприятие реальной опасности и «просто беспокойства» отделены друг от друга, т.к. в урбосреде они требуют разной реакции. Настороженность к первым остаётся такой же (иногда повышается), ко вторым — радикально снижается, при значимо большей способности их дифференцировать между собой и легче переключаться от полного покоя, без каких-либо признаков тревоги к осматриванию и бегству. Всё это мотивирует птиц к исследованию и использованию соответствующих объектов и ситуаций, ранее подавлявшихся страхом и бегством. В общем потоке событий городской жизни второе встречается намного чаще (примерно тем чаще, чем урбанизированней территория, на которой селятся птицы, или часть участка, в которой она действует «здесь и сейчас»). Следовательно, общее уменьшение дистанции взлёта при урбанизации отражает именно и только его.
«Городские» особи лишь лучше фильтруют поток стимулов, всё более становясь безразличными к тему, к которым лучше привыкнуть и не обращать внимания, поскольку они преобладают в потоке на входе. Однако те немногие, что надо отреагировать, они реагируют сильней, чем в исходных популяциях: характер реакции в сравнении с ними здесь не изменён, тогда как при доместикации он меняется на противоположный. Эти поведенческие изменения наряду с «рассыпанием» корреляций, обязательных для птиц из исходных популяций, суть адаптации широкого значения, повышающие приспособленность особи сразу в нескольких (часто многих) полярных друг другу проблемных ситуациях, чьё разрешение требует активных действий животного, меняющих прежнюю модель поведения, и одновременно — резистентность к средовым «возмущениям» в ситуациях, требующих устойчивой реализации прежней активности им вопреки. Лучшая дифференциация того и другого, более точное разделение времени и усилий животного между ними дополняет это приспособление, хорошо соответствующее максимальной контрастности урбосреды как в пространстве (гетерогенность) так и во времени (нестабильность каждой ландшафтной мозаики «здесь и сейчас» при быстрой направленной эволюции их общего «кружева» на более длинном временном интервале).
Дистанция взлёта как показатель скрывает два разных процесса оценивания нового и неизвестного — как опасности и как безразличного и/или отчасти интересного, первый ведёт к бегству, второй — привыканию, отчасти приближению и исследованию. Урбанизация их разделяет, и делает обе оценки более точными, в чём «развивающее» воздействие урбосреды. При доместикации, наоборот, животные вообще отказываются от оценивания и своё окружении воспринимают всё более дружелюбно. Поэтому уменьшение дистанции взлёта при урбанизации — негодный аргумент в отождествлении её с доместикацией. Оно не отражает «уменьшение страха» перед бывшей реальной опасностью, а без этого предположения не гомологизируешь оба процесса. Данная гомологизация ложной аналогии, когда обобщение разделение явлений происходит по основанию, важному для исследователя, без специального доказательства его существенности для самих сравниваемых феноменов.
Примечания
1Причём у тех видов, где это специально исследовали, масштаб уменьшения сильно зависит от величины города, освоенного птицами (Møller, 2008, 2014; Møller, Ibáñez-Álamo, 2012; Morelli et al., 2018), и плотности пешеходов в местах пребывания птиц, но не зависит от величины парка, сохранности местообитаний и пр. характеристик самих экосистем (6 видов в оригинальной работе D.Blumstein, 2014; Mikula, 2014; 32 вида в мета-анализе Samia et al., 2017). Оно пропорционально времени, прошедшему с начала урбанизации, но не зависит от величины мозга самих птиц, хотя эта последняя и/или величина оптической доли — хороший предиктор дистанции взлёта в исходных популяциях (42 в мета-анализе Symonds et al., 2016, fig.3-5).
2Наблюдатель здесь идёт к птице спокойным шагом без остановок, с постоянной скоростью. Дистанция регистрируется в момент взлёта в шагах, затем переводят в метры. Учитываются лишь те случаи, когда птицы видят наблюдателя издали, и расстояние до них значительно. Перебегание, перепархивание и пр. не учитывается. В случае с дятлами мы учитывали дистанцию, на которой часть птиц, прервав прежнее поведение, вместо взлёта уходит на другую сторону ствола (она устойчиво меньше дистанции взлёта), и долю птиц, ушедших за ствол. Не учитывали дистанцию, при которой взлетают эти птицы (при продолжении подхода наблюдателя). Неточность оценивания компенсировали большим их количеством. Для репрезентативности данных дистанции оценили в максимально разнообразных условиях — для разных особей, разных поселений вида, в разные годы и месяцы, при разной погоде, для птиц разного статуса, пола и возраста и пр. (это т. н. максимально гетерогенная выборка).
3Эти различия не универсальны, см. исследование P.major выше (Senar et al., 2017)
4Большой минус анализа в работах, цитируемых ниже — в них повсеместно речь идёт об агрессивности «вообще», авторы игнорируют качественные различия агрессии к конспецификам (территориальной, или связанной с доминированием-подчинением в группах) и к хищникам, гнездовым конкурентам etc, хотя они многажды подтверждены этологическими экспериментами (Ryabitsev, 1995).
5Не всегда: она снижена у «городских» северных кардиналов Сardinalis cardinalis (оценка по интенсивности атак и агонистических демонстраций при подстановке чучела конспецифика того же пола). У них же значимо увеличены смелость, готовности исследовать новое помещение (оценка частотой прыжков и перелётов), но снижены: страх приближения к новым объектам в контексте кормления и в знакомом пространстве, уровень кортикостерона при поимке (он и слабей поднимается при предъявлении стрессора). Однако «сельские» особи, более агрессивные к конспецификам, медленней восстанавливают активность (т. е. хуже преодолевают страх) после предъявления модели перепелятника Купера вместе с криком тревоги острохохлых синиц Baeolophus bicolor, к которому кардиналы чувствительны (Huang et al., 2020).
6Австралийские города вообще отличаются повышенной агрессивностью видов, освоивших их успешней прочих. К названным двум надо добавить серёжчатого медососа Anthochaera carunculata, молуккского ибиса, солдатского чибиса, сорочьего жаворонка Grallina cyanoleuca и обыкновенную майну Acridotheres tristis (Lowry et al., 2013).
7За некоторым исключением, вроде реакции гнездовых пар кроличьих сычей на возможного хищника у норы (Cavalli et al., 2016).
8Заселение Лодзи началось в 1990-х, учёты 1994-2002 гг. показывают гнездование лишь на окраинах города. В годы наблюдений, 2009-2014 гг., лысуха расселилась до центра (Minias, 2015, fig.1).
9Отличия индивидуальности «городских» серых полёвок (повышенные смелость, принятие риска, исследовательская активность), однако, исчезают при тестировании после 9-14 нед. содержания в лаборатории. Поведение «сельских» особей, наоборот, не меняется; его «городские» особенности здесь созданы фенотипической пластичностью и пока что не закреплены собственно микроэволюцией (Massa et al., 2020).
10В первую очередь в экологическом плане: комплекс сосуществующих близких видов не ограничивает вселение новых на данную территорию, рост перекрывания ниш на разных территориях пропорционален трансформации их природных сообществ, гемеробности ландшафта, но не ведёт к интенсификации конкуренции (Фридман, Ерёмкин, 2009: 3; Фридман, Суслов, 2018: 85-86, 117-120), сосуществование хищников и жертв на «островах» городских или пригородных лесов не ведёт к циклике, характерной для природных местообитаний (Марченко, 2008), значительная численность хищников отнюдь не мешает образованию и росту численности популяций жертв (серые вороны Corvus cornix с тетеревятниками A.gentilis vs рябинник T.pilaris в Москве, Морозов и др., 2015). Это т.н. парадоксы городских экосистем.
11Ещё важный момент — сигналы птиц, прежде всего песня, в урбосреде полностью сохранили видоспецифичность формы и определённость значений, несмотря на зашумленность (остались «честными», по модной сейчас социобиологической терминологии) и даже сделались более значимыми. Больше того, в ответ на рост зашумлённости «городские» особи интенсифицируют подачу сигнала и меняют структуру вокализаций, т.ч. вероятность детекции и точность распознавания возрастают несмотря на помехи (Akçay, Beecher, 2019; Akçay et al., 2020ab). Так, самцы горных гаичек Parus gambeli, обитающие в «сельской» части градиента урбанизации, с низким уровнем антропогенного шума, при дополнительном его проигрывании интенсифицировали выдачу позывов за счёт песен. Обитавшие на более урбанизированных территориях, с большим уровнем шума, напротив, интенсифицировали пение. Везде обработка шумом меняла частотные характеристики позывов, но не песен (La Zerte et al., 2017).
12При этом потенциальная опасность продолжает стрессировать птиц. Например, частота дыхания — одно из прокси уровня стресса, у «городских» больших синиц Parus major устойчиво повышена в сравнении с «сельскими», также как уровень сердечных сокращений (Charmantier et al., 2017). То же отмечено у мексиканских чечевичников (Weaver et al., 2018). Это независимое подтверждение, что урбанизация отнюдь не обращает страха в «доверчивость», стресс сохраняется, в т.ч. в виде большей дистанции настораживания (англ.alert distance) у особей с меньшей дистанцией взлёта. «Городские» птицы с ним просто лучше справляются: они в целом раньше обнаруживают подходящего человека, но позже взлетают сравнительно с «сельскими», т. е. дольше остаются в настороженном состоянии и подпускают ближе (Samia et al., 2017, fig.2). Ряд данных показывает, что рост безразличия к беспокойству обусловлен воздействием антропогенного шума, с накапливающимся эффектом (Petrelli et al., 2017), что объясняет зависимость от времени с начала урбанизации вида (Symonds et al., 2016).
13Вид территориален, с круглогодично постоянными парами, члены которых совместно кормятся на земле (в основном насекомыми) в городских парках, во дворах и на улицах.
14в смысле не суровости условий существования — они как раз мягче, а их исторической нетипичности для подавляющего большинства видов птиц и млекопитающих, осваивающих урбосреду, и выхода за пределы фундаментальной ниши последних.
1521.09.2010 в «Лосином острове» наблюдал как ворон С.corax дразнил пару собак – из той стаи бездомных собак, что шляется между визит-центром в Мытищинском лесопарке и Нехлюдовым рукавом. Сидя на сухом дереве над ними, он крукал так, что в его круканьях был акустический компонент, очень сходный с собачьим лаем (как минимум на мой слух). Собаки злились и лаяли в ответ, заходясь всё больше и больше; в том числе потому, что ворон перелетал по ветвям кроны всё ниже и ниже к псам, и вставлял свои круканья точно между их лаями.
16К «базовому» стрессирующему воздействию урбосреды, отражённому в учащении дыхания, большей интенсивности криков бедствия и пр. реактивности «городских» птиц сравнительно с «сельскими».
17Как исключение, оно сопрягается с ростом толерантности к человеку у некоторых синантропов, ставших таковыми вне города, в с/х ландшафте и сельских населённых пунктах (Møller, 2009).
18Повышенная осторожность к людям и технике сохраняется несмотря на благожелательность людей к орлам, примелькавшимся у селений и не вызывающим интереса. Численность вида растёт, вопреки сокращению популяции малого суслика (Фридман и др., 2016).
19Просмотр дневниковых записей позволил А.Ю.Соколову (2017) отнести начало освоения новой ниши к более ранним срокам — с 2005-х гг. или даже с 1999 г., когда в ряде районов Воронежской обл. клинтухи стали регулярно встречаться в агроландшафтах.
Рекомендуем прочесть
 «Самодоместикация»: модель, которая не везде работает
«Самодоместикация»: модель, которая не везде работает Смелость города берёт
Смелость города берёт «Город как заказник»: (пока?) упускаемая возможность
«Город как заказник»: (пока?) упускаемая возможность Модели урбанизации «диких» видов птиц
Модели урбанизации «диких» видов птиц Урбанизация — не доместикация
Урбанизация — не доместикация Урбанизация «диких» видов птиц/млекопитающих как доместикация домашних
Урбанизация «диких» видов птиц/млекопитающих как доместикация домашних Разбор популярных мифов про охрану природы
Разбор популярных мифов про охрану природы Что не так с социобиологией?
Что не так с социобиологией?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}