

Николай Колчанов, Валентин Суслов
В 2009 г. издательство Палеонтологического института РАН выпустило сборник «Проблемы происхождения жизни» по материалам одноименного коллоквиума, прошедшего годом ранее в Москве. Помещенные в нем доклады посвящены различным аспектам возникновения и ранней эволюции биосферы Земли, а также ее связям с миром органических молекул Вселенной.
Научный прогресс подтверждает необходимость специализации. Но есть и оборотная сторона медали: сужение кругозора, когда накопление фактов обгоняет их понимание, ведя к застою. Как известно, в науке всегда были «вечные» проблемы. Их решения порой приходится ожидать веками, они часто воспринимаются как «отвлеченные», но ученые разных поколений вновь возвращаются к ним. И дело не в стремлении «объять необъятное». «Вечные» проблемы междисциплинарны, требуют широкого кругозора, интенсивного обмена между весьма далекими друг от друга областями знаний. Словом, их значение не ограничивается только рамками конкретного содержания. Они объединяют отдельные направления в энциклопедическое знание там, где это не могут успешно сделать ни исследования по частным аспектам, ни заимствование экспериментальных методик. Происхождение жизни — один из таких «вечных» вопросов. Ему целиком посвящен рассматриваемый нами сборник.
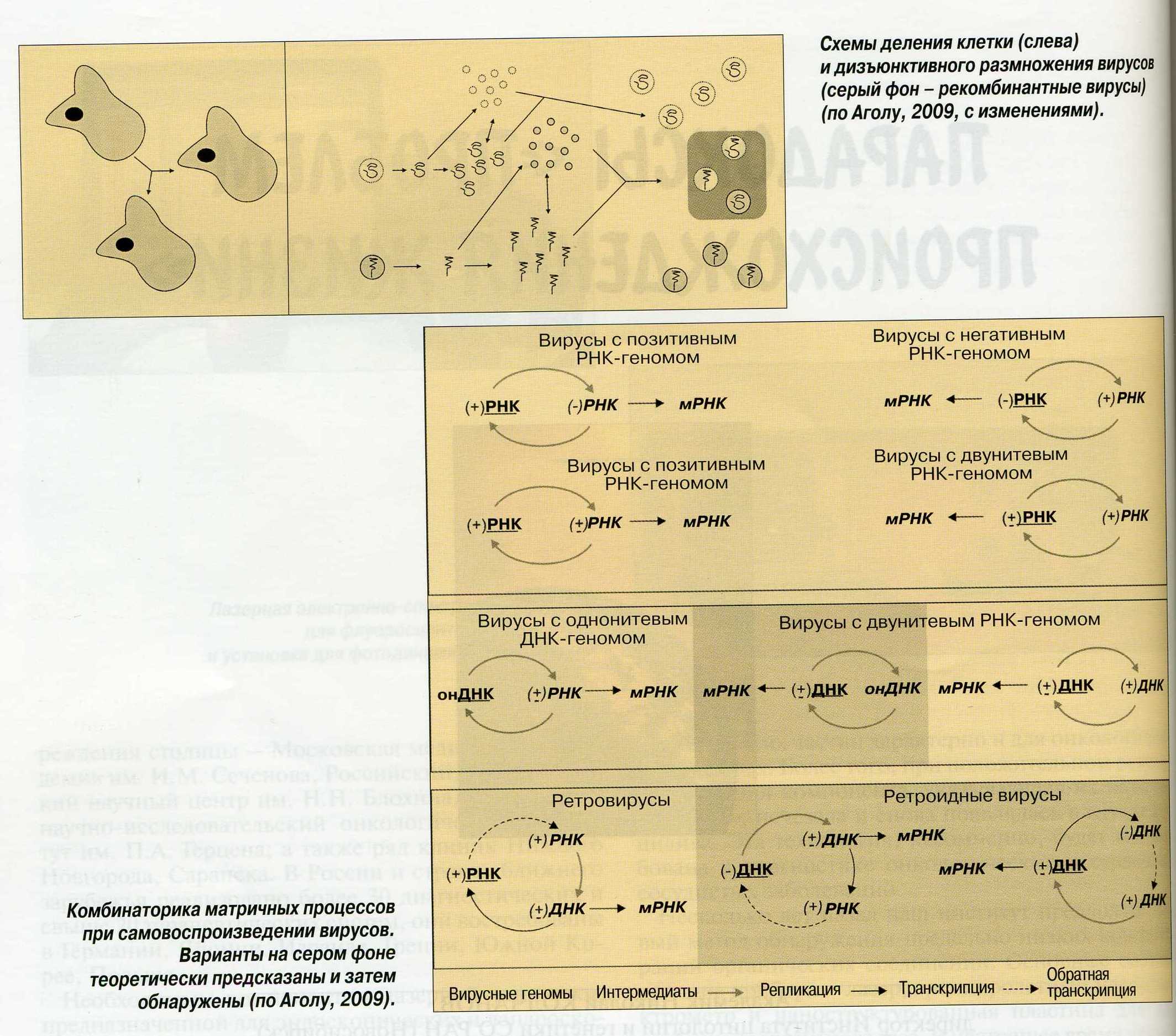
Рисунок 1 (вверху) и 2 (внизу). Схемы деления клетки (слева) и дизъюнктивного размножения вирусов (серый фон — рекомбинантные вирусы) (по Аголу, 2009, с изменениями).
Его авторы — представители разных дисциплин: астрономы (доктора физико-математических наук из Института космических исследований РАН Леонид Ксанфомалити и Лев Мухин, Николай Бочкарев — доктор физико-математических наук из Государственного астрономического института им. П. К. Штернберга МГУ им. М. В. Ломоносова), геологи (академик Николай Добрецов, доктора физико-математических наук Андрей Витязев и Галина Печерникова из Института динамики геосфер СО РАН), палеонтологи (академик Алексей Розанов), химики (кандидат физико-математических наук Валерий Снытников из Института катализа им. Г. К. Борескова СО РАН), биохимики (академик Александр Спирин, член-корреспондент РАН Александр Четверин), молекулярные биологи (член-корреспондент РАН и РАМН Вадим Агол), микробиологи (академик Георгий Заварзин, доктор биологических наук из Института микробиологии им. С. Н. Виноградского РАН Елизавета Бонч-Осмоловская) и физиологи (академик Юрий Наточин).
На самом деле авторов гораздо больше (в коллоквиуме участвовали 36 человек), поскольку к каждой главе сборника приложена стенограмма дискуссии. Последняя цементирует разнородность обсуждаемых тем, разъясняя моменты, специфичные для той или иной науки, тем самым придавая книге целостность и, что важно, делая ее интересной не только для конкретного специалиста.
Мероприятие проходило в рамках программы Президиума РАН «Происхождение и эволюция биосферы». Но в отличие от прежних конференций и изданных по их итогам коллективных монографий, в данном случае значительная часть материалов посвящена астробиологии.
Авторы сосредоточились на восьми проблемах, в решении которых за последние 20 лет появилось много нового. Речь идет о том,
- возможно ли собрать клетку из ее компонентов;
- о происхождении вирусов — до или после клетки;
- о значении для жизни органических веществ Вселенной;
- о мире РНК и возможности его существования на предземной стадии;
- условиях жизни на ранней Земле 4,6 — 4,0 млрд. лет назад и после, когда следы биосферы явно присутствуют;
- о глубинной биосфере нашей планеты как возможном аналоге первичных древних экосистем
- и, наконец, о происхождении мембран — липидной оболочки клетки, через которую идет обмен веществ между ней и внешней средой.
Соответственно первую, вторую, четвертую и восьмую из перечисленных проблем можно объединить как сценарий организмогенеза, т.е. как возник организм (в частности, простейший — клетка). Третья и пятая — в каких условиях внешней среды это происходило? Наконец, шестая — теория экогенеза: как организм взаимодействовал со средой и с себе подобными?
А теперь по порядку. В молекулярной биологии рутинными стали задачи по полной расшифровке геномов. За последние 20 лет особенно впечатляет прогресс в секвенировании1: широко используемые секвенаторы второго поколения снизили стоимость и время работ на 2-3 порядка. За рубежом заговорили о синтетической геномике2, цель которой — конструирование новых видов, либо комбинируя фрагменты геномов разных видов, либо их искусственным синтезом de novo (т.е. с самого начала).
В главе «Можно ли собрать клетку из ее компонентов?» известный ученый в области физико-химической биологии Александр Четверин, оценив перспективу лабораторного синтеза, перешел к возможности этого события в природе, указав на фундаментальное противоречие. С одной стороны, самосборка отдельных элементов клетки ныне хорошо изучена (липидные мембраны, белки, нуклеопротеиновые комплексы, включая такие сложные, как вирусы и рибосомы). Но стоит сформироваться мембране (ее самосборка наиболее проста), как поступление крупных мульти-молекулярных компонентов будет прервано, а низкомолекулярных — аминокислот, нуклеотидов, Сахаров и т.д., без чего невозможен метаболизм, — затруднено. Иными словами, для самосборки нужно беспрепятственное поступление компонентов, однако быстрое образование замкнутых мембранных полостей — компартментов — нарушит проток (все живые системы — проточные).
С другой стороны, без компартментализации эффективная эволюция затруднена. Ведь отбор оценивают не сами гены, а их продукты — фенотип. Следовательно, усовершенствование протоклеток тем эффективнее, чем сильнее связаны в пространстве и времени ген и продукт. Но проток, нужный для самосборки, не дает гарантии, что продукт не будет унесен от родителя и станет обслуживать в лучшем случае чужие гены, а в худшем вообще затеряется. Аналогичная ситуация наблюдается у вирусов с их дизъюнктивным (разделенным) размножением, когда ДНК(РНК) и белки синтезируются отдельно, и лишь затем при смешивании происходит самосборка, дающая как родительские, так и различные рекомбинантные вирусы. Выходит, с компартментализацией сформировать организм невозможно, а без нее — нельзя. Решение этого парадокса автор видит в открытом им явлении образования молекулярных наноколоний РНК на субстрате с нанометровыми порами. В качестве субстрата в лабораторных исследованиях он использовал гель, в природе же им мог быть монтмориллонит — широко распространенный глинистый минерал, формирующий поры подходящего размера.
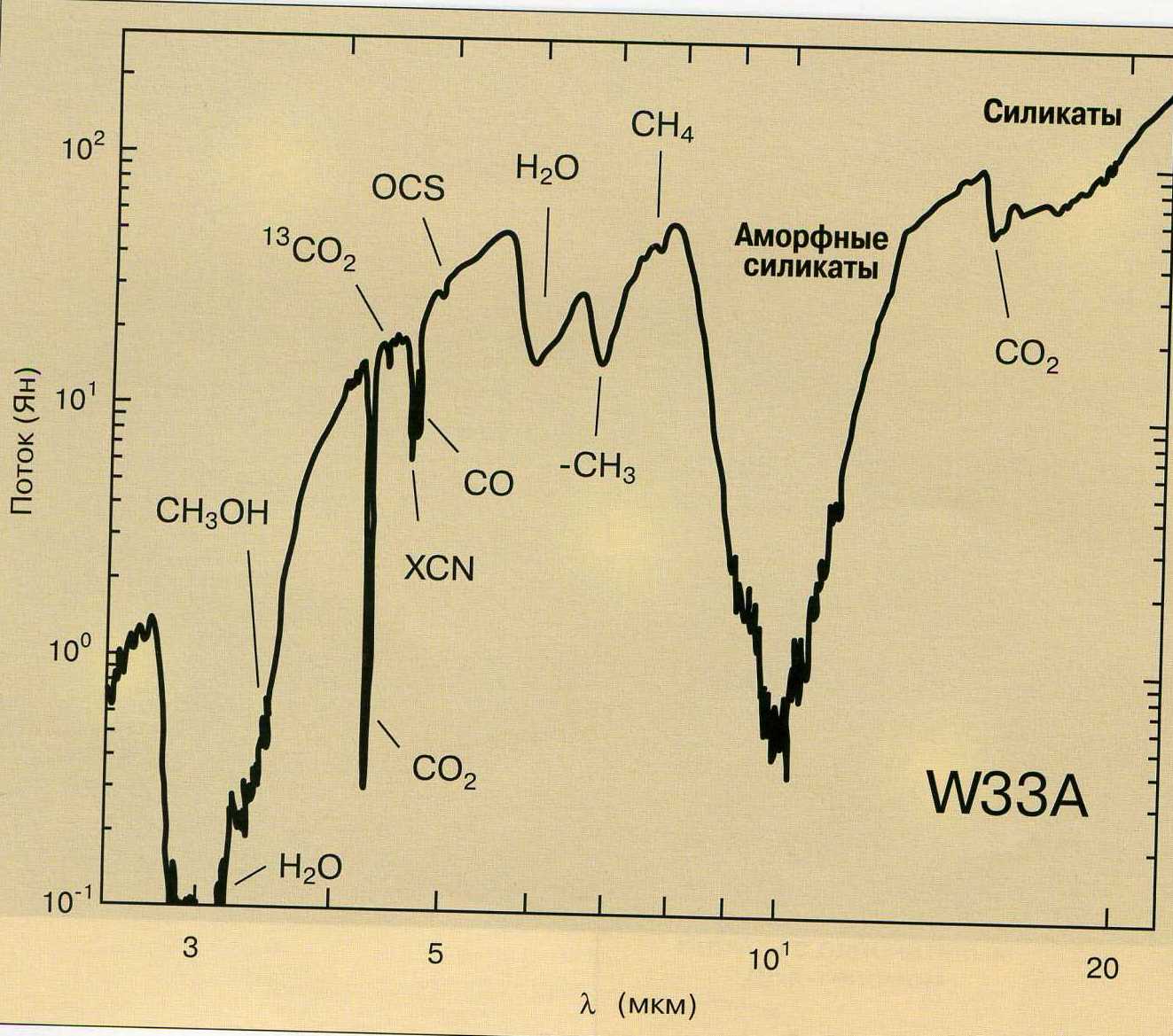
Рисунок 3. Инфракрасные полосы в спектрах пыли области звездообразования W33A, позволяющие судить о ее составе: по горизонтальной оси — длины волн, мкм; по вертикальной оси — спектральные плотности потоков излучения в янских Jy (1 Ян = 10-23 Вт/м2 Гц).
В опытах Четверина крупные молекулы на субстрате с нанопорами ведут себя как мухи в паутине — «заякорившись», они остаются на месте (немембранная компартментализация), но могут в широких пределах гнуться, сворачиваться, разворачиваться, меняя конфигурацию в пространстве — конформацию. Изменение последней лежит в основе репликации, транскрипции и других ферментативных реакций, обеспечивающих удвоение ДНК и метаболизм. В итоге потомки одной молекулы формируют наноколонию. Отбор среди них можно вести, меняя условия — увлажняя и подсушивая субстрат — и проток низкомолекулярных компонентов.
Изредка отдельные РНК покидают «нанопаутину» и, как показали опыты, переносятся по воздуху, инфицируя новый субстрат. Однако потенциально наилучший способ размножения — растрескивание последнего и перенос на его кусочках наноколониальных фрагментов. Итак, в отличие от точки зрения Крейга Вентера (США) — «отца» синтетической геномики — в природе, по мнению Четверина, прямая самосборка клеток невозможна. Возникновение жизни должно идти окольным путем — через промежуточную стадию молекулярных наноколоний.
Отметим: хотя восьмая проблема рассматриваемой книги — происхождение мембран — помещена в конце, в целом она продолжает первую, ведь они — неотъемлемые компоненты клетки. Например, в стенограмме дискуссии Валерий Снытников подчеркнул: алюмосиликатные глины типа монтмориллонита — хорошие катализаторы. А недавно было экспериментально показано: в порах данного минерала возможен безматричный синтез РНК. Но ведь катализатор истощится, когда РНК забьет поры. В результате наноколония будет расти краями, используя чистый монтмориллонит. Однако если ее окружить мембраной, то рост прекратится. Похоже, мембраны препятствуют и самосборке, и самому существованию наноколоний. В чем же тогда преимущество, если компартментализация возможна без них?
Как это бывает в «вечных» проблемах, ответ пришел из неожиданной области. Юрий Наточин (глава «Возникновение мембран») — крупный специалист в области физиологии почки — предположил: эволюционное преимущество должно быть энергетическим. Как известно, мембрана стабилизирует ионный состав внутри клетки и создает асимметрию концентраций ионов между внутренней и внешней средой. Эта асимметрия — основной аккумулятор энергии для синтеза аденозинтрифосфата (АТФ)3. Напротив, для молекулярных наноколоний нет различий между внутренней и внешней средой, ибо ионная концентрация легко меняется.
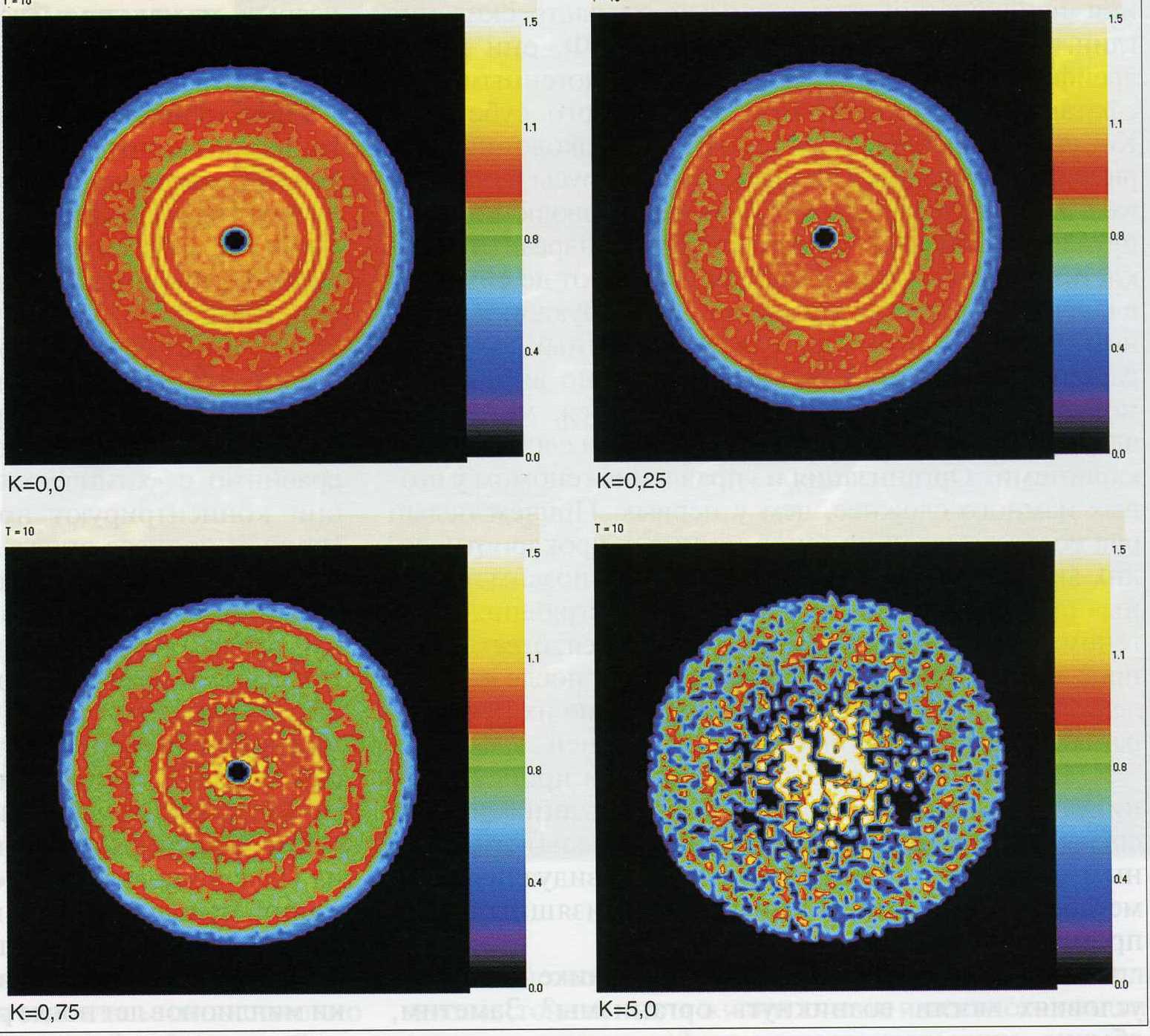
Рисунок 4. Графики логарифма плотности частиц для различных констант их коагуляции (К) в самогравитирующем диске, полученные компьютерным моделированием (по Снытникову, 2006).
Ферментативная активность макромолекул зависит от их конформации, а она, в свою очередь, от того, каких ионов и сколько в среде. Поэтому частые колебания их состава снижают эффективность метаболизма наноколоний и затрудняют их эволюцию. Можно сказать, что наноколония «не знает», куда эволюционировать — концентрация ионов может меняться быстрее, чем фиксируются мутации. Формируя внутреннюю среду, мембрана с трансмембранными белками-помпами стабилизирует ионный состав, канализируя4 эволюцию ферментов.
Это сразу сказывается на эволюции — отбираются лишь те ферменты, которые эффективны в данной среде. Такая эволюция намного эффективнее, чем неканализированная: во-первых, значительно меньше жертв выбраковывает отбор, во-вторых, приспосабливаясь к одним и тем же условиям, ферменты усложняются. Автоматически за счет избирательного транспорта ионов создается асимметрия их состава — аккумулятор энергии, впоследствии утилизованный эволюцией для синтеза АТФ, а у животных — и для передачи нервного импульса.
Вирусолог и молекулярный биолог Вадим Агол (глава «Вирус — до или после клетки») рассмотрел три гипотезы происхождения вирусов: или это потомки клеток, дегенерировавшие под влиянием паразитизма, или производные отдельных генов, или же доклеточные генетические элементы. Первые две теории доминировали в конце XX в., сейчас накапливаются факты в пользу третьей. Дело в том, что размножение вирусов принципиально отличается от клеточного деления и из него не выводится. Кроме того, по последним данным вирусы реализовали все мыслимые схемы комбинаторики матричных процессов самовоспроизведения нуклеиновых кислот (включая теоретически предсказанные начиная с 1970-х годов, в том числе и в лаборатории Вадима Агола).
А вот молекулярная генетика клеточных организмов свидетельствует о консерватизме схем деления и матричного самовоспроизведения. Значит, резонно предположить: будь вирусы производными от клеток, у них наблюдался бы аналогичный консерватизм.
Если принять в качестве протоорганизма наноколонию Александра Четверина, можно предположить: вирусы — первая попытка эволюции оторвать такую колонию от субстрата. Если аналогичных попыток было несколько, тогда происхождение вирусов полифилетично — они ведут начало от разных предков. Исходно протовирусы представляли собой механизм размножения наноколоний, независимый от растрескивания глин. Неспособные синтезировать АТФ, они могли дрейфовать в поисках обогащенного абиогенными фосфатами монтмориллонита или другого субстрата. Когда же более удачная попытка наноколоний оторваться дала первые клетки, то протовирусы использовали их как субстрат. С этого момента эволюционные пути и тех, и других тесно переплелись: паразитируя на клетках, вирусы участвовали (и участвуют до сих пор!) в формировании как прокариот, так и эукариот — одноклеточных протистов, а также животных, грибов и растений.
Более того, возможно именно вирусы помогли преодолеть эволюционный рубеж между доядерными прокариотами и организмами с ядром — эукариотами. Организация и управление геномом у вторых намного сложнее, чем у первых. Причем целый ряд соответствующих генов не имеет прокариотических аналогов и, возможно, они были «позаимствованы» предками эукариот у вирусов, «застрявших» в их геномах в результате мутаций. Получается, ответ на вопрос «происхождение вирусов — до или после клетки» лежит где-то посредине — формирование их началось раньше клетки, а закончилось вместе с ней.
Итак, между основными сценариями происхождения жизни — гомеостазом (способность динамически сохранять постоянство внутренней среды), матричным синтезом макромолекул и индивидуализацией метаболизма — авторы книги нашли изящный компромисс.
Следующая проблема в данном сборнике — в каких условиях могли возникнуть организмы? Заметим, обычно считается: происхождение жизни включает «подготовительную» стадию химической эволюции. Данные радиоастрономии и спектроскопической астрофизики (об этом речь идет в главе «Молекулы и их миграция во Вселенной» Николая Бочкарева) свидетельствуют: простые органические молекулы (размером до 13 атомов углерода) есть в атмосферах холодных звезд, планет, их спутников и в межзвездных газопылевых и молекулярных облаках. Но если сложная молекула не наблюдается, это еще не значит, что ее нет — возможно, у нас просто нет достаточно чувствительных приборов, чтобы ее идентифицировать. Да и различные галактики могут быть несхожими по составу органики.
Астрофизики предполагают: постоянно действующими химическими реакторами органического синтеза являются молекулярные облака и околозвездная среда. А ледяные оболочки малых тел — от пылинок до комет — основные «склады готовой продукции». Обмен же органическими молекулами, вероятнее всего, происходит в зонах столкновения галактик.
Словом, Вселенная постоянно синтезирует простую органику.
Парадокс, как отметил специалист в области химического катализа Валерий Снытников, заключается в несоизмеримости масштабов «астрономического» и «химического» времени. Так, химический период многостадийного синтеза сложных органических соединений с учетом восстановления катализаторов, перехода между промежуточными стадиями и т.д. длится от часов до десятков лет, тогда как астрономический начинается минимум с десятков тысяч лет. Подобный разрыв между промежуточными стадиями синтеза приводит к тому, что вся сложная органика успевает разложиться.
Итак, нужен компактный химический реактор и механизм концентрации в нем органики Вселенной. Валерий Снытников, проэкспериментировав с реальным метеоритным веществом с точки зрения химика-технолога и смоделировав астрофизические процессы в аккреционных дисках звезд, предложил на роль подобных реакторов области уплотнения газопылевой среды. Главную роль в формировании последних играет гравитационная нестабильность аккреционного диска5.
Согласно компьютерному моделированию, время жизни таких эфемерных астрономических объектов сравнимо с «химическим». Вращаясь вокруг звезды, они концентрируют простую рассеянную органику (причем процесс определенным образом взаимозависим) и каталитически синтезируют сложные вещества — аминокислоты, сахара и др. Если такое плотное облако станет зародышем планеты, то на ней может быстро сформироваться жизнь хотя бы уровня наноколоний. Так на стыке теорий катализа и астрофизики возник астрокатализ — оригинальная теория, объединяющая в единый процесс «подготовительную» стадию химической эволюции жизни и планетогенез.
Как указывают астрофизики Андрей Витязев и Галина Печерникова в главе «Ранняя Земля в тесном окружении молодых звезд», по современным данным последние возникают группами — скоплениями в очагах звездообразования. Стало быть, Солнце первые десятки миллионов лет после рождения провело в обогащенной межзвездной среде. Авторы, развивая стандартную модель происхождения Солнечной системы, показали: уже в первые миллионы лет ее существования было возможно образование планет и дифференциация их недр на ядро, мантию и кору. Вероятно, известная метеоритная бомбардировка, чьи следы хранит Луна (~3,7 — 3,9 млрд. лет назад), — следствие того, что Солнце покинуло звездное скопление.
А отсутствие значимого эксцентриситета (отклонения от окружности) орбиты Земли свидетельствует: она счастливо избежала губительных для жизни мегаимпактов, т.е. столкновения с космическим телом массой порядка Марса. Однако в дискуссии ученые отметили: при наличии на нашей планете Мирового океана вполне достаточно астероида диаметром порядка 400 — 500 км, чтобы, вскипятив его, практически уничтожить ее биосферу.
Далее. В разделе «Условия на поверхности Земли 4 — 4,6 млрд. лет назад» физик Лев Мухин показал: несмотря на тектоническую активность и метеоритную бомбардировку, даже на ранней планете вероятно сохранение участков земной коры с температурой, достаточной для существования жидкой воды. Основной проблемой возникновения жизни здесь должен быть недостаток фосфора. Этот элемент — один из самых редких во Вселенной. Выходит, нужно искать его «острова» в Галактике.
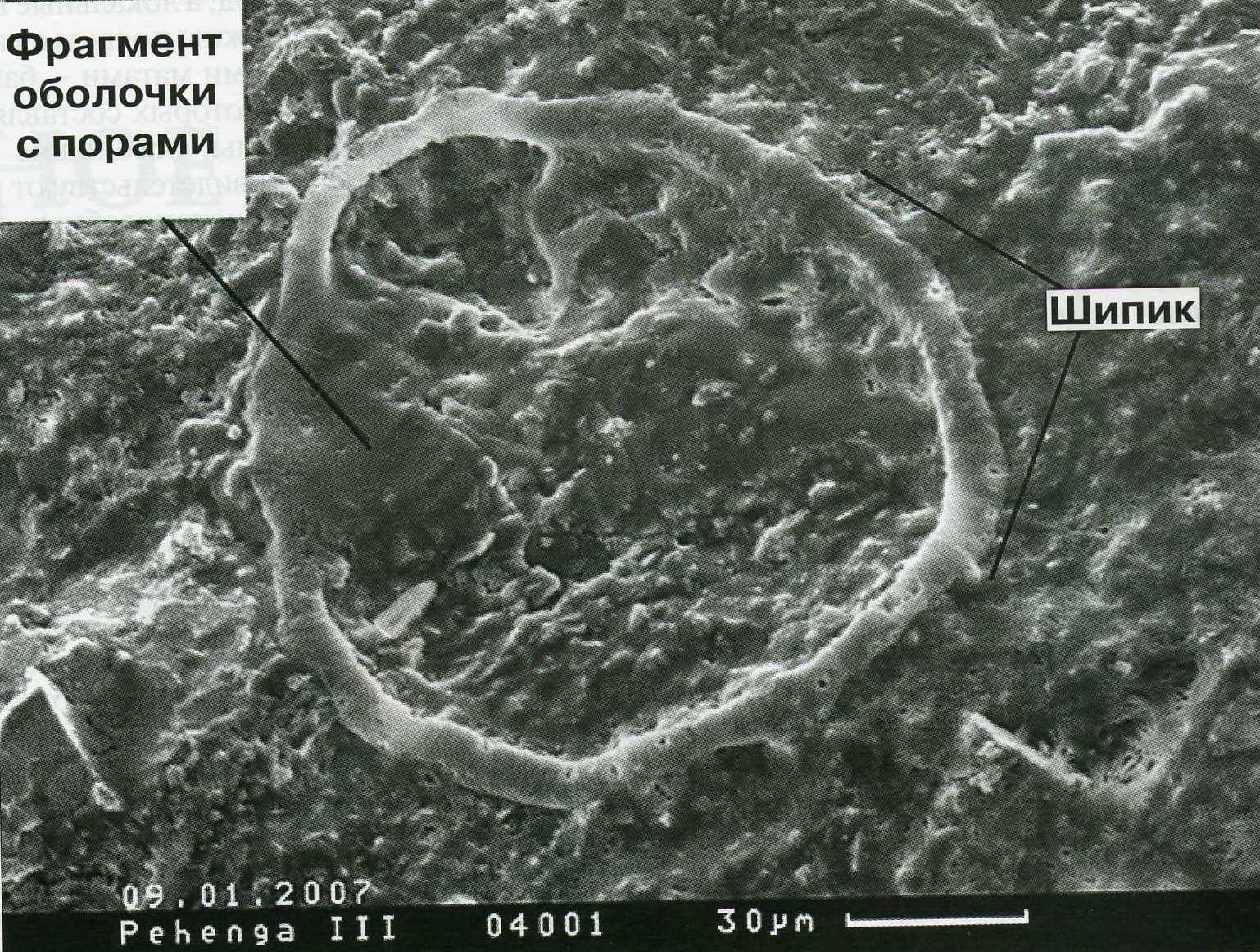
Рисунок 5. Окаменелость Pechengia melezhiki. Размер ~130 мкм и морфологические особенности не позволяют интерпретировать ее как прокариот (по Розанову, 2009).
Итак, для формирования жизни или заноса ее извне наша будущая Земля представлялась относительно комфортной уже на стадии аккреции либо сразу после ее окончания. Впрочем, то же самое можно сказать и о всех планетах земной группы. Значит, и там должны быть следы жизни.
Однако коллега Мухина, астроном Леонид Ксанфомалити разочаровал в этом участников коллоквиума (глава «От Марса к Европе: поиск биосферы на спутниках планет-гигантов»). Хотя на древнем Марсе был океан, а на современном еще существует жидкая вода, следов жизни здесь не найдено.
Магнитное поле там исчезло около 4 млрд. лет назад, большие постоянные водоемы существовали порядка 2 млрд. лет назад. Как уже говорилось, тогда на Земле уже была прокариотическая биосфера. А американские марсоходы в XX в. нашли несомненные метеориты на Красной планете, но не смогли установить их происхождение. Зато на нашей планете обнаружены вторичные метеориты с Марса — осколки его породы, выброшенные в космос сильным взрывом (вероятно, при ударах крупных метеоритов, оставивших свои следы-кратеры на поверхности). Марсианское происхождение впервые установлено методами тонкого химического и изотопного анализа при переизучении метеорита «Nakhla» в Египте. С тех пор за четверть века найдено около 30 вторичных метеоритов. Если предположить, что Марс и Земля обменивались породой, то земные прокариоты могли попасть на Красную планету в порах этой породы. Почему же они не прижились? Не обладает ли Земля особым физическим свойством или их сочетанием, необходимым для жизни (точнее — для жизни клеток известного нам химического состава)?
Крупный палеонтолог Алексей Розанов в главе «Псевдоморфозы по микробам в метеоритах» представил множество фотографий микроокаменелостей из углистых хондритов (они представляют подавляющее большинство каменных метеоритов. — Прим. ред.), не отличимых от окаменевших остатков земных прокариот, эукариот и экосистем — прокариотических матов. Изотопные данные не противоречат их биогенной природе. Эксперименты по выживанию бактерий и вирусов при кратковременном нагреве до 200°C показывают: у космических путешественников были шансы уцелеть даже при «жесткой посадке» (поскольку метеориты среднего размера внутри редко сильно разогреваются).
Если возраст углистых хондритов около 4,5 млрд. лет, то организмы, оставившие нам окаменелости, старше Земли и существовать могли только на других землеподобных планетах. Однако Леонид Ксанфомалити отметил уникальность нашей планеты — в ближнем космосе подобных ей нет.
Хотя авторы сборника договорились не давать определение жизни, подразумевалось, что она включает в себя минимум две взаимодополнительные сущности — организмы и экосистему. Первые путем матричных процессов воспроизводят генетическую информацию. Вторые, состоящие из функционально разнородных организмов, формирующих линейные и/или замкнутые трофические цепи, стабилизируют флуктуации протока первичных вещества и энергии.
Эти колебания связаны с геохимическими циклами, причем динамика их сильно отличается у разных планет. Сравнительная планетология свидетельствует, что Марс практически вышел из «коридора обитаемости» для биосферы земного типа, а Венера с самого начала своего существования в него не входила. Таким образом, заключает в главе «Первые экосистемы на Земле» ведущий эколог-микробиолог Георгий Заварзин, функционирование жизни в масштабе «астрономического» времени возможно, если экосистемы «впишутся» в геохимические циклы планеты, и чем богаче они видами и сбалансированнее, тем больше у них на это шансов. Отсюда два важных вывода. В случае возникновения жизни на планете, функциональное разнообразие должно появляться вместе с жизнью — экосистемы и организмы формируются не последовательно, а параллельно (пусть даже на уровне наноколоний).
В случае же занесения жизни из космоса наибольшие шансы имеет готовая экосистема. Поэтому крайне важна находка в метеоритах окаменевших остатков прокариотических экосистем, о которой сообщил Алексей Розанов.
Мысль Заварзина о единстве биосферы проиллюстрировала его коллега Елизавета Бонч-Осмоловская в обзоре «Высокотемпературные глубинные микробные сообщества — возможный аналог древнейших экосистем?». Глубоководные хемотрофные сообщества океанских гидротерм («черные курильщики») — часть фотоавтотрофной биосферы, ибо интенсивно обмениваются с ней химическими элементами и микробным населением. Экосистемы естественных нефтяных резервуаров, как оказалось, живут за счет захороненного органического вещества былых биосфер. Трофически автономными можно считать лишь глубинные гидролитосферные экосистемы интерстициальных вод (последние формируются в трещинах пород). Но и они изредка обмениваются с поверхностью микробным населением.
Насколько древними могут быть наземные и мелководные автотрофные экосистемы показал Алексей Розанов (глава «Условия жизни на ранней Земле после 4,0 млрд. лет назад»), проведя ревизию геологических и палеонтологических данных нижнего докембрия6. Сравнение фоссилизированных7 остатков современных прокариотических экосистем с докембрийскими не выявило принципиальных отличий. Это заставляет предположить: трофическая структура микробиальных сообществ и условия их фоссилизации, а значит, и существования не должны принципиально отличаться от современных.
Прокариотическая биосфера в промежутке между 4 и 2 млрд. лет назад приняла планетарный характер, заселив мелководные моря шельфа и выйдя из них на континенты. В этом с Алексеем Розановым солидарен и Георгий Заварзин, отметивший, что для защиты от ультрафиолета достаточно тонкого слоя восстановленных пород, а локальные высокие концентрации кислорода должны появиться на Земле вместе с цианобактериальными матами — бактериальными экосистемами, основу которых составляют цианобактерии — первые организмы, освоившие кислородный фотосинтез. О том же свидетельствуют ранние (2,04 млрд. лет) находки эукариотоподобных фоссилий (Pechengia melezhiki) Кольского полуострова.
Подробно историю взаимоотношений нашей планеты и существующей на ней биосферы проследил крупный геолог Николай Добрецов в главе «Корреляции между необратимыми и периодическими событиями глубинной геотектоники планеты Земля, глобальной тектоники плит и ключевыми моментами эволюции биосферы». По его мнению, эволюция Земли как планеты связана с необратимым понижением температуры ее недр и повышением вязкости мантии. Процесс теплопереноса нестабилен: на фоне постепенного остывания мантии отмечены циклические изменения ее температуры. Это соответственно определяет особенности тектоники плит (в частности, периодичность цикла Уилсона — от образования до распада суперконтинентов — пангей) и через вулканическую активность вносит вклад в глобальные геохимические циклы, характерную динамику оледенений, изменение состава изверженных пород, а в итоге — влияет на темпы эволюции биоты. В частности, периодичность глобальных вымираний, начиная с кембрий/ордовикского и заканчивая последним мел/палеогеновым, хорошо согласуется с рассчитанной в лаборатории Николая Добрецова периодичностью мантийных плюмов8, через которые сбрасывается излишек тепла в мантии.
Биосфера никак не может повлиять на циклику температуры мантии нашей планеты, но она в силах ускорить или замедлить вымирание либо преодолеть его последствия, реагируя на это в свою очередь эволюцией видов или перестройкой экосистем. По мере приближения к нашему времени биоразнообразие растет, а разрушительность кризисов падает.
Словом, из работ разных специалистов, собранных в рассматриваемом сборнике, вырисовывается довольно непротиворечивый сценарий. Конечно, не все вопросы решены (неясно, например, куда делись породы возрастом более 4 млрд. лет, если на древней Земле оставались холодные участки). Однако это означает только одно — существует новое направление научных исследований, нужны еще эксперименты, а соответственно, значительные открытия еще впереди.
Наука в России. 2010. №5.
Примечания
1Секвенирование — определение первичной аминокислотной или нуклеотидной последовательности белков и нуклеиновых кислот (прим. ред.).
2Геномика — раздел молекулярной генетики, посвященный расшифровке и сравнительному изучению геномов как целого (прим. авт.).
3Аденозинтрифосфат (АТФ) — нуклеотид, играющий роль универсального источника энергии для всех биохимических процессов, протекающих в живых системах (прим. ред.).
4Канализация эволюции — появление устойчивой тенденции изменения того или иного признака вследствие неизменного действия отбора в стабильных по тому или иному параметру условиях среды (в данном случае — по концентрации ионов и ионному составу). Канализацию эволюции не следует путать с ортогенезом, когда тенденция изменения признака ограничена его строением (прим. авт.).
5Аккреция — процесс выпадения вещества на космическое тело из окружающего пространства или концентрации вещества вокруг космического тела (прим. ред.).
6 На докембрий приходится большая часть геологической истории Земли — около 4.0 млрд. лет (прим. Ред.).
7Фоссилизация — полное или частичное замещение органики погибших организмов неорганическими соединениями окружающей среды (прим. Ред.).
8Плюм — горячий поток мантийного вещества, относительно независимый от общей картины конвективных течений в мантии. «Всплывая» по направлению от ядра к коре, он способен изменить очертания континента, пробудить или заново создать вулканы и т.д. (прим. авт.).
Рекомендуем прочесть
 День национального позора
День национального позора Оценка степени фрагментации местообитаний диких животных искусственными рубежами на примере Московского региона
Оценка степени фрагментации местообитаний диких животных искусственными рубежами на примере Московского региона «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Португалия: «Кто-то теряет, кто-то находит»
Португалия: «Кто-то теряет, кто-то находит» Трущобы Китая будут перестраивать
Трущобы Китая будут перестраивать Ирландия: Выборы «по-изумрудному»
Ирландия: Выборы «по-изумрудному»  Италия: Еврошестёрка
Италия: Еврошестёрка  «Мужской мозг», «женский мозг», и научный метод
«Мужской мозг», «женский мозг», и научный метод
