Дж. Хсйлмен (Jack Р. Hailman)
Под термином «инстинкт» применительно к повелению животных и человека обычно понимают достаточно сложные стереотипные формы активности, которые присущи всем особям данного вида, передаются по наследству и не требуют индивидуальною научения. Однако торможение при управлении автомобилем или удар бейсбольной битой-тоже сложные, стереотипные поведенческие акты, которые можно наблюдать у многих людей, но они, несомненно, не были бы возможны без обучения. Но, может быть, и для выработки стереотипных форм поведения у животных тоже требуется некоторое, не столь явное, научение? Иными словами, не содержат ли инстинкты и приобретенного компонента?
Для проверки этой гипотезы мы избрали в качестве экспериментальной модели один из типичных инстинктов пищевую реакцию птенцов чайки. Мои коллеги и я наблюдали повеление птенцов как в естественных условиях, так и в лаборатории, где мы провели ряд экспериментов с целью выяснить ход развития пищевой реакции. Мы пришли к выводу, что в нормальной выработке этой формы поведения значительную роль играет индивидуальный
опыт. Более того, наши исследования дают серьезные основания полагать, что и в развитии других форм инстинктивного поведения содержится элемент научения.
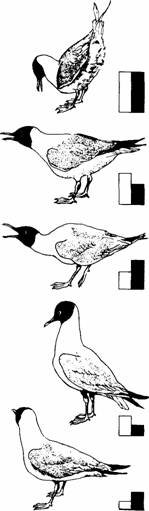
Смеющиеся чайки выводят птенцов на заболоченных прибрежных островах у восточного побережья Северной Америки. Тихо сидя в укрытии поблизости от гнезда, можно наблюдать за кормлением птенцов (рис. 112). Взрослая птица опускает голову с клювом, направленным вниз, перед семидневным птенцом. Если после предыдущего кормления прошло достаточно времени, птенец совершает сложное координированное движение-клюет клюв взрослой птицы и тянет его вниз. После нескольких таких клевков взрослая птица отрыгивает частично переваренную пищу. Таким образом, клевки птенца, по-видимому, являются формой выпрашивания корма. Потом птенец клюет рыбу, отрывая и заглатывая куски.
Следовательно, клевки — это также и способ поглощения пищи. Когда птенцы в гнезде уже наелись досыта, птица съедает остатки корма. Дальнейшие наблюдения позволили выявить ряд более сложных моментов во взаимодействии птенцов и взрослой птицы. Если обычное опускание клюва перед глазами птенцов не вызывает у них пищевой реакции, птица начинает слегка покачивать клювом из стороны в сторону. Такое движение обычно стимулирует клевки. Отрыгнув пищу в гнездо, птица дожидается, пока птенцы начнут кормиться.

Смеющаяся чайка Larus atricilla
Если этого не происходит, взрослая птица опять опускает клюв, как бы указывая на пищу. После этого птенцы обычно начинают клевать. Если же птенцы и теперь не реагируют на пищу, птица берет ее в клюв и держит перед ними. Как только птенцы начинают клевать корм, птица сразу отпускает его.
В связи с этими внешне простыми реакциями клевания возникает ряд вопросов относительно возможной роли индивидуальною опыта в развитии
пищевого поведения. Как может птенец касаться своим клювом клюва взрослой птицы при выпрашивании корма и отрывать куски от принесенной
пищи, если обе формы клевка в своей основе столь сходны? Почему птенец поворачивает голову в сторону при клевке во время выпрашивания, но не делает этого, когда клюет пищу? Нужна ли птенцу практика, чтобы добиться точности и координированности клевка? Почему голодный птенец клюет, а сытый-нет? Почему клевки птенца не бывают нацелены на красные лапы взрослой птицы или на другие окружающие предметы? Как птенец распознаёт пищу?
В поисках ответов на эти и многие другие вопросы наша группа занималась экспериментальным изучением поведения птенцов чайки с момента их вылупления и до достижения недельного возраста. К седьмому дню жизни формирование пищевых реакций у птенцов практически завершается. Кроме того, ограничивая наше исследование таким коротким периодом, мы могли более четко контролировать отдельные элементы онтогенеза и оценивать их вклад в поведение. Как это часто бывает, наше исследование породило больше вопросов, чем разрешило, однако мы получили много новых сведений.

Рис. 112. Нормальное пищевое поведение птенца смеющейся чайки включает два различных, но сходных типа клевков. На рисунках изображен птенец в возрасте около трех дней, у которого формирование пищевой реакции в основном завершено. Когда взрослая птица опускает голову (А), птенец точно нацеленным и хорошо скоординированным клевком хватает клюв родителя (Б) и тянет его вниз. После этого взрослая птица отрыгивает частично переваренную пищу на дно гнезда (В). и птенец начинает клевками поедать ее (Г).

Рис. 11З. Для проверки точности клевков птенцам предъявлялись карточки со схематическим изображением головы взрослой чайки. Клевки отмечены точками. Слева — результат эксперимента, проведенного с только что вылупившимся птенцом, справа — с тем же птенцом через два дня.
Рассмотрим сначала развитие точности клевков. Для изучения этой проблемы мы схематически изобразили голову взрослой птицы на маленьких карточках (рис. 113). Карточка, прикрепленная к подвижному стержню, могла перемещаться в горизонтальном направлении перед глазами птенца.
Собранные в гнездах яйца мы инкубировали в темноте, так что птенцы до начала эксперимента не получали никаких зрительных стимулов. После вылупления в первый же день каждому птенцу предъявляли подвижную двумерную модель головы взрослой птицы и позволяли ему клюнуть ее 10-15
раз. Место каждого клевка мы отмечали на модели точкой.
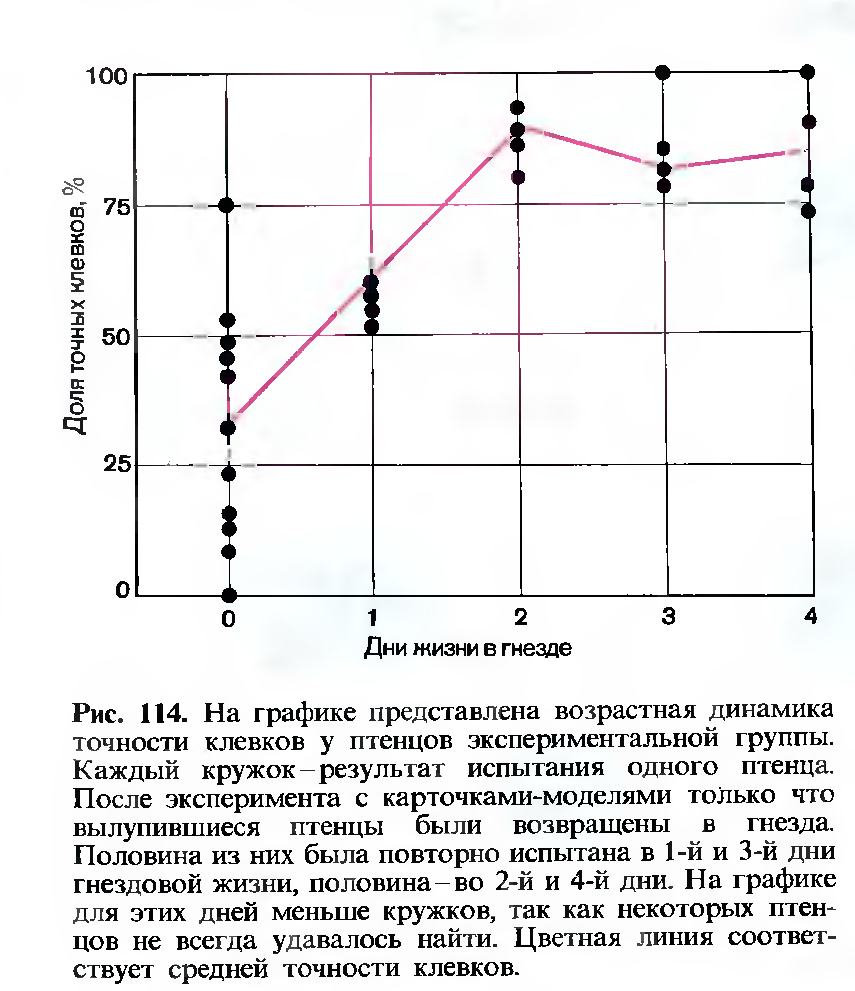
После эксперимента мы. предварительно пометив птенца, возвращали его в гнездо и брали взамен яйцо, из которого были слышны писки, т.е. яйцо с эмбрионом, близким к вылуплению. Меченый птенец, таким образом, воспитывался в естественных условиях приемными родителями. На 1-й, 3-й и 5-й дни после вылупления половину меченых птенцов тестировали вновь. Поведение остальных птенцов исследовали на 2-й, 4-й и 6-й дни. После каждого эксперимента птенцов опять возвращали в гнездо.
Опыты показали, что сразу после вылупления птенцов в среднем только треть их клевков достигает цели. На следующий день точность уже превышает 50%, а через два дня после вылупления достигает величины более 75% и в дальнейшем не меняется. Анализ данных для типичного птенца показывает, что разброс клевков, особенно горизонтальный, значительно уменьшается с возрастом.
Как же происходит столь быстрое повышение точности клевков? Для выяснения этого вопроса мы разработали более сложный эксперимент с использованием двух контрольных групп птенцов, выращенных в естественных условиях. Чтобы птенцы экспериментальных групп не могли приобрести опыта зрительной координации клевания, их содержали в темных брудерах. Птенцов одной из этих групп кормили искусственно. Птенцы второй группы не получали пищи в течение двух дней; они существовали за счет обильных жировых запасов. Для того чтобы выяснить, влияет ли на точность клевков нормальная активность, связанная с вылуплением, птенцов третьей группы извлекали из яиц сразу после появления эмбриональных писков и помещали в инкубатор.
В различные сроки после вылупления птенцам предъявляли модель головы взрослой птицы. Поведение птенцов регистрировалось на кинопленке, анализ которой позволял определять процент точных клевков. Во всех пяти группах точность с возрастом увеличивалась, но только в двух контрольных группах она достигала нормального уровня (свыше 75%). В отличие от зрительной депривации, оказывавшей заметное влияние на точносгь клевания, исключение нормальной активности, связанной с вылуплением, практически не влияло на результаты тестирования.
Наиболее осторожной интерпретацией этих результатов мог бы быть следующий вывод: зрительный опыт необходим для достижения максимальной точности клевания, но некоторое повышение точности происходит и в условиях зрительной депривации. В последнем случае улучшение может быть результатом совершенствования лозных реакций. Здесь тоже возможно влияние индивидуального опыта, так как весьма вероятно, что позные реакции эффективно отрабатываются у птенцов в условиях темного инкубатора.
Как научается птенец выбирать правильное расстояние для точного клевка? Наши наблюдения над только что вылупившимися птенцами позволяют предположить, что здесь действует механизм саморегуляции, основанный на восприятии расстояния. Если неопытный птенец встанет слишком близко к модели, то удар оказывается настолько сильным, что он может отбросить птенца назад на 2-3 см.
И наоборот, когда птенец находится слишком далеко от цели, он не достает до нее при клевке и падает вперед на 4 5 см. Старшие птенцы редко допускают такие серьезные ошибки; можно думать, что с возрастом они методом проб и ошибок научаются выбирать наилучшее расстояние.
Исследователи нередко рассматривали пищевую мотивацию (голод) как результат научения. Наши эксперименты позволяют предполагать, что по
крайней мере в основе голода лежит нечто врожденное. Ряд опытов показал, что у накормленных досыта птенцов, как и следовало ожидать, частота клевков возрастает с увеличением интервала времени после кормления. Однако такая же картина наблюдалась и у птенцов, которым мы ни разу не давали случая испытать чувство голода. Птенцов, вылупившихся в темном инкубаторе, искусственно кормили до насыщения на вторые сутки жизни, а затем тестировали на свету с моделями головы. Через час после кормления средняя частота клевков составляла у них 3,1 в минуту, а через два часа-5,1 (разница статистически достоверна).
Детали движений при клевке во время пищевой реакции мы анализировали с помощью высокоскоростной киносъемки. У птенца в возрасте нескольких дней пищевая реакция, к этому времени уже вполне сформированная, включает четыре основных компонента: 1) открывание и последующее закрывание клюва; 2) движение головы вверх и вперед по направлению к голове взрослой птицы, а затем вниз и назад; 3) поворот головы в сторону
в расчете на то. чтобы захватить клювом клюв взрослой птицы, и обратный поворот головы; 4) легкий толчок ногами вверх и вперед (см. рис. 114). Покадровый анализ фильма позволил выявить значительную изменчивость временных отношений между этими компонентами как у каждого отдельного птенца, так и у различных птенцов. С возрастом эта изменчивость несколько уменьшается, что, вероятно, связано с улучшением координации движений (подробно мы это явление не изучали).
Среди интересных деталей, выявленных при анализе кинолент, можно отметить тот факт, что у птенцов, выросших в естественных условиях, поворот головы в сторону наблюдался с возрастом все чаще. Чтобы выяснить, насколько это зависит от зрительного опыта, мы проанализировали
фильмы, заснятые во время тестов на точность клевания во всех пяти группах птенцов. Оказалось, что у птенцов, не имевших опыта клевания, сколько-нибудь заметный поворот головы наблюдался редко. У птенцов, выросших в гнезде, в первые сутки жизни поворотов головы не бывает, но затем эта реакция появляется и быстро совершенствуется.
Мы не знаем, какова конкретная роль индивидуального опыта в таком развитии поведения, но снятые фильмы дают основание для некоторых предположений. Иногда, когда птенец клюет с раскрытым клювом, клюв взрослой птицы, расположенный не строго вертикально, может оказаться между челюстями птенца. Тогда движение головы птенца по инерции вперед поневоле приводит к ее повороту в сторону. Возможно, именно так птенец
научается поворачивать голову, чтобы схватить клюв взрослой птицы.
Один из самых интересных вопросов, связанных с пищевым поведением птенцов,-это вопрос о том, как птенцы узнают своих родителей. Наблюдения
за развитием птенцов в естественных условиях показали, что птенцы иногда клюют не только клюв взрослой птицы, но и иные предметы, в том числе другие части тела родителя. Правда, большинство клевков все же нацелено на клюв, особенно в более позднем возрасте. Это позволяет предположить, что недавно вылупившиеся птенцы представляют себе родителей довольно смутно, но с возрастом, по мере накопления опыта, образ взрослой птицы становится у них все более четким.
Эту проблему мы исследовали с помощью набора различных моделей головы и клюва (рис. 115). Изменяя или устраняя отдельные части таких моделей, мы смогли выявить наиболее эффективные стимулы для пищевой реакции. Обычно модель закреплялась на стержне, который мог колебаться в такт с метрономом, так что скорость движения модели была всегда известна. В каждом опыте птенцу в случайном порядке предъявлялись пять моделей и при этом регистрировалось число клевков за определенное время (обычно за 30 секунд).
Наша первая задача состояла в том, чтобы выявить наиболее эффективные стимулы для птенцов, находившихся в темноте в течение суток с момента вылупления до начала эксперимента. В первых трех опытах птенцы одинаково реагировали на все модели, кроме той, у которой не было клюва. Таким образом мы показали, что новорожденные птенцы реагируют в основном на клюв родителей, а не на особенности головы (даже само наличие головы для птенцов этого возраста несущественно).
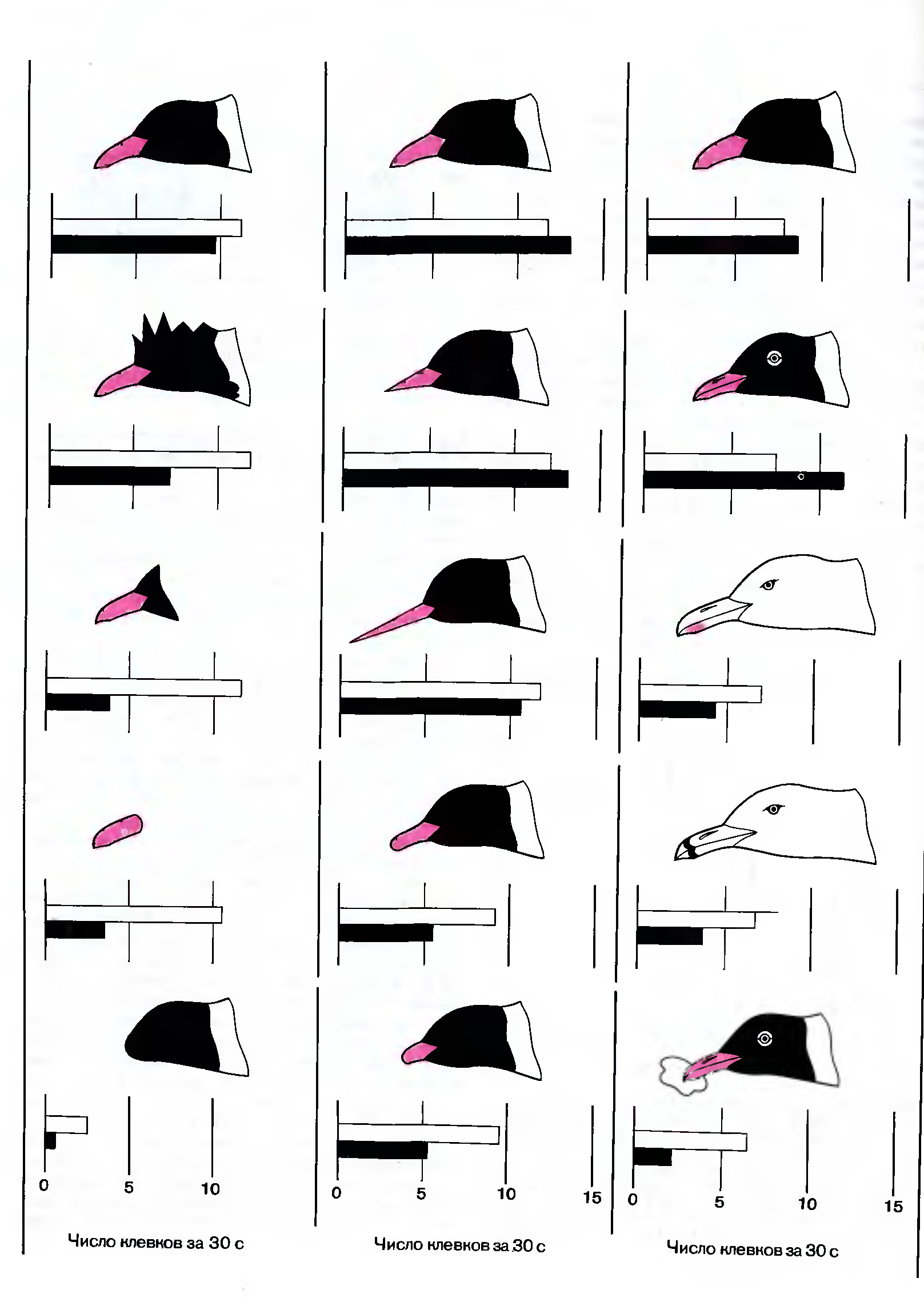
Рис. 115. Детали головы, наиболее эффективные как стимулы для пищевой реакции, исследовались в опытах с предъявлением наборов из пяти моделей только что вылупившимся (белые столбики) и более взрослым (черные столбики) птенцам смеющейся чайки. Набор, изображенный в правой колонке, включает модели головы серебристой чайки (третья сверху) и дэлавэрской чайки (четвертая сверху). Изображена также модель с пищей в клюве.
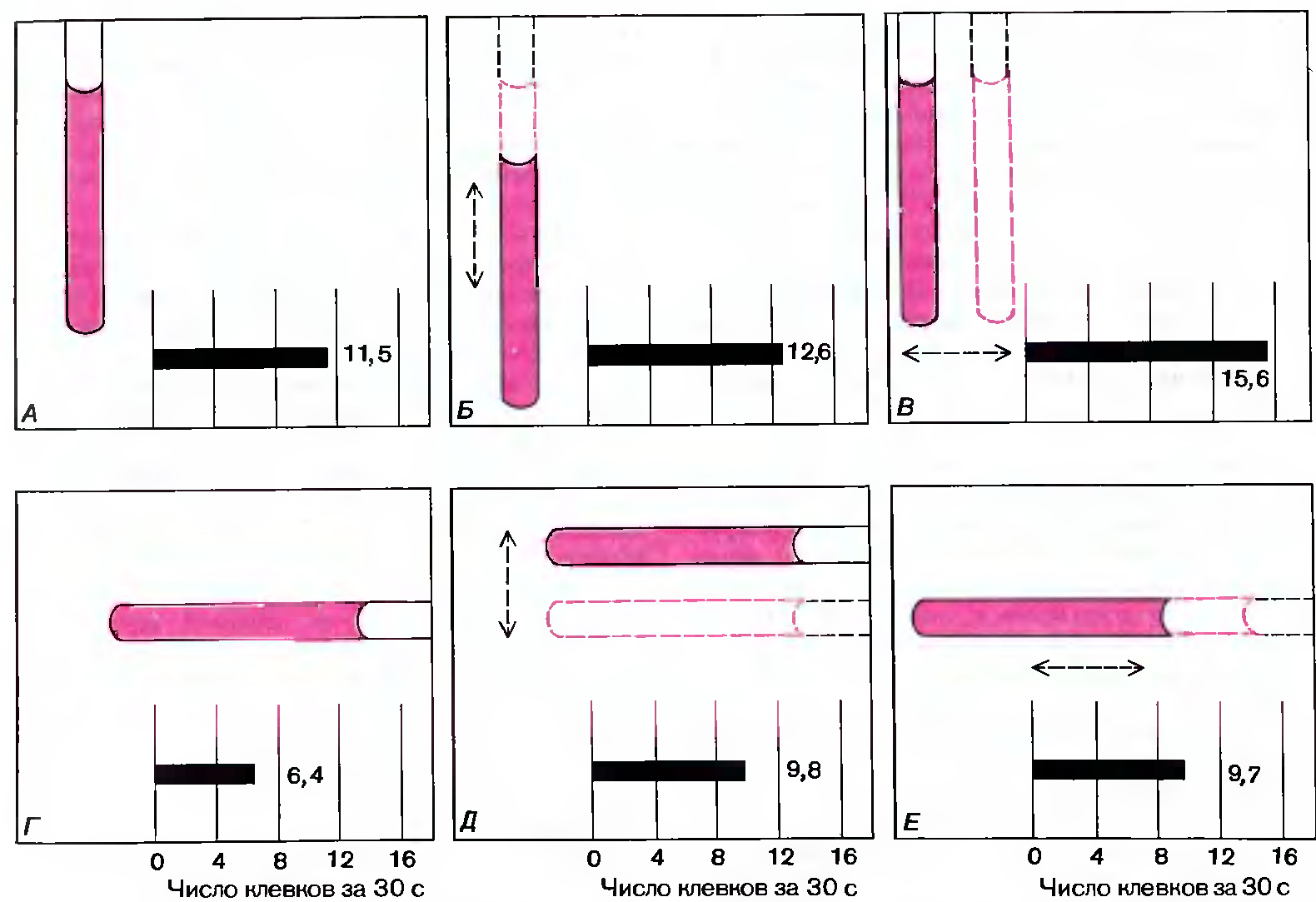
Рис. 116. Эффективность различных стимулов испытывали с помощью деревянных палочек, по толщине примерно равных клюву взрослой чайки. Подсчитывали среднее число клевков для группы из 25 птенцов за 30 секунд. Предъявляли: вертикальную неподвижную палочку (А); вертикальную палочку, движущуюся вертикально (Б) или горизонтально (В); неподвижную горизонтальную палочку (Г); горизонтальную палочку, движущуюся вертикально (Д) или горизонтально (Е).
В одном из первых трех опытов выяснился неожиданный момент: птенцы смеющейся чайки одинаково реагируют на модели, имитирующие их собственных родителей и взрослых особей серебристой чайки, несмотря на резкое различие между этими видами. У смеющейся чайки черная голова и красный клюв; у серебристой чайки оперение на голове белое, а клюв желтый с небольшим красным пятном на нижней челюсти. Птенцы смеющейся чайки реагировали именно на это пятно, воспроизведенное на модели.
Но если птенцы смеющейся чайки не отличают своих родителей от серебристых чаек, то интересно было бы знать: различают ли этих птиц птенцы серебристой чайки? Чтобы выяснить это, мы провели опыты с нашими моделями в большой колонии серебристых чаек на островах Гран-Манан в бухте Фанди. Оказалось, что птенцы серебристой чайки тоже не различают взрослых птиц обоих видов.
Это навело на мысль, что птенцы реагируют на какие-то элементарные особенности формы и движения, свойственные и красному клюву смеющейся
чайки, и красному пятну на клюве серебристой чайки. Хотя оптимальный стимул для реакции клевания, очевидно, несложен, он, видимо, должен обладать какими-то чертами, отличающими его от прочих окружающих объектов, таких как красные лапы родителей или травинки, поскольку клевки птенцов редко бывают нацелены на посторонние предметы.

Кольцеклювая чайка Larus delawarensis
Мы исследовали этот вопрос с помощью деревянных палочек, окрашенных в красный цвет. Палочку предъявляли птенцу в горизонтальном или вертикальном положении, причем в обоих случаях она либо была неподвижна, либо перемещалась в горизонтальном или вертикальном направлении (рис. 116).
Любая вертикальная палочка вызывала клевание с большей частотой, чем любая горизонтальная, а из вертикальных наиболее эффективной оказалась палочка, движущаяся в горизонтальном направлении. Это хорошо согласуется с естественной ситуацией, когда вертикально расположенный клюв взрослой птицы часто движется горизонтально перед глазами птенца.
Дальнейший анализ результатов показал, что вертикально движущийся вертикальный стимул вызывал не больше реакций, чем неподвижный вертикальный. Кроме тою, и горизонтальное, и вертикальное перемещение горизонтального стимула было одинаково действенно и, как эго ни странно, превосходило по эффективности неподвижный горизонтальный стимул. Наиболее осторожная интерпретация этих результатов может состоять в том,
что адекватными стимулами для пищевой реакции птенцов служат лва типа движения: 1) горизонтальное перемещение и 2) движение в направлении, перпендикулярном продольной оси стимула. Это позволяет объяснить, почему неподвижная вертикальная палочка оказывается не менее эффективным стимулом, чем вертикальная палочка, движущаяся вертикально. Дело в том, что в обоих случаях отсутствуют движения указанных типов. Однако в случае горизонтальной палочки вертикальное движение перпендикулярно продольной оси стимула и потому гак же эффективно, как горизонтальное, и реакция здесь при обоих движениях более интенсивна, чем при полной неподвижности стимула.
На следующем этапе мы испытывали вертикальные стимулы различных диаметров и пять скоростей движения. Палочка диаметром около 8 мм была наиболее эффективна вне зависимости от скорости движения. Скоростью, вызывающей наиболее интенсивную реакцию независимо от диаметра палочки, оказалась скорость 12 см/с. Эти результаты показывают, насколько точно реактивность в процессе эволюции подгоняется к параметрам стимулов. Толщина клюва взрослой птицы в вертикальном направлении 10,6 мм. а в горизонтальном-3,1 мм, так что средняя толщина составляет как раз около 8 мм. Кроме того, как показал анализ фильмов, снятых в естественных условиях, средняя скорость движения клюва при кормлении равна около 14,5 см/с.
Проведенный недавно эксперимент добавил еще одну особенность к образу идеального для птенцов стимула. Вертикальная палочка, находящаяся выше уровня глаз птенца, гораздо эффективнее, чем такой же стимул, предъявленный внизу. Кроме того, предпочтение отдается наклонным объектам. Такая избирательность уменьшает вероятность возникновения у птенца реакции на ноги родителей, расположенные, естественно, ниже уровня его глаз.
Теперь мы поняли, по крайней мере в общих чертах, каким образом только что вылупившийся птенец отличает клюв родителей от других внешних объектов. На следующем этапе нам предстояло выяснить, изменяется ли восприятие птенца в первые несколько дней жизни. Мы предъявляли те же три набора по пять моделей в каждом (см. рис. 115) семидневным птенцам и нашли, что их поведение замолю отличается от поведения только что вылупившихся птенцов.
Более взрослые птенцы оказались чувствительными даже к незначительным изменениям формы головы и клюва. Кроме того, они четко отличали модели собственных родителей — смеющихся чаек — от моделей серебристых чаек. Чтобы выяснить, развивается ли у птенцов серебристой чайки такое же предпочтение к своим родителям, мы пометили их и испытали примерно на
4-й и 7-й дни гнездовой жизни. Оказалось, что и у них реакция на модели птиц своего вида с возрастом увеличивалась, а на модели смеющихся чаек — уменьшалась.
Связано ли такое изменение в восприятии птенцов с научением, т.е. с опытом общения с родителями? Чтобы узнать это, мы провели серию экспериментов с тремя группами птенцов серебристой чайки, выведенными в инкубаторе. Птенцы первой группы получали небольшое количество пищи, после того как несколько раз клевали модель головы смеющейся чайки; у птенцов второй группы подкрепляли такую же реакцию на модель их собственного вида. Птенцы третьей, контрольной группы получали пищу без предварительного предъявления какой-либо модели. После двух дней такого обучения птенцы в тестах без пищевою подкрепления лучше реагировали на ту модель, с которой они имели дело раньше. Хотя это лишь предварительный эксперимент, полученные результаты позволяют думать, что изменения реакции у птенцов,
выросших в гнезде, могли быть результатом выработки условной связи.
Итак, наши данные указывают на то, что недавно вылупившийся птенец лучше всего реагирует на очень простые стимулы. Хотя экспериментатор может создать модель, в качестве стимула значительно более эффективную, чем настоящая птица, в естественных условиях признаки родителей точнее соответствуют «образу» стимула для пищевой реакции, чем особенности любого другого внешнего объекта. Кроме того, по мере выкармливания птенца родителями их образ в памяти птенца детализируется. Семидневные птенцы клюют только модели, достаточно сходные с взрослыми птицами того же вида.

Американская серебристая чайка Larus argentatus smithsonianus с птенцом
Наши результаты не вполне согласуются с данными этологических наблюдений Н.Тинбергена и А.Пердека, также изучавших поведение серебристой чайки. Они нашли, что перемещение красного пятна с клюва на лоб модели резко снижало ее эффективность как стимула. Так как в этом классическом опыте все основные элементы стимула как будто бы сохранялись, а менялось только их расположение, был сделан вывод, что только что вылупившиеся птенцы серебристой чайки имеют уже сформированный образ родителей.
Мы полагали, что вопрос о том, остаются ли все элементы стимула действительно идентичными, требует дополнительного изучения. В экспериментах Тинбергена и Пердека модель держали в руке, поэтому при ее маятникообразном движении вместе с кистью руки пятно на лбу двигалось медленнее и по более короткой дуге, чем на клюве. Кроме того, птенцу приходилось тянуться вверх, чтобы клюнуть пятно на лбу.
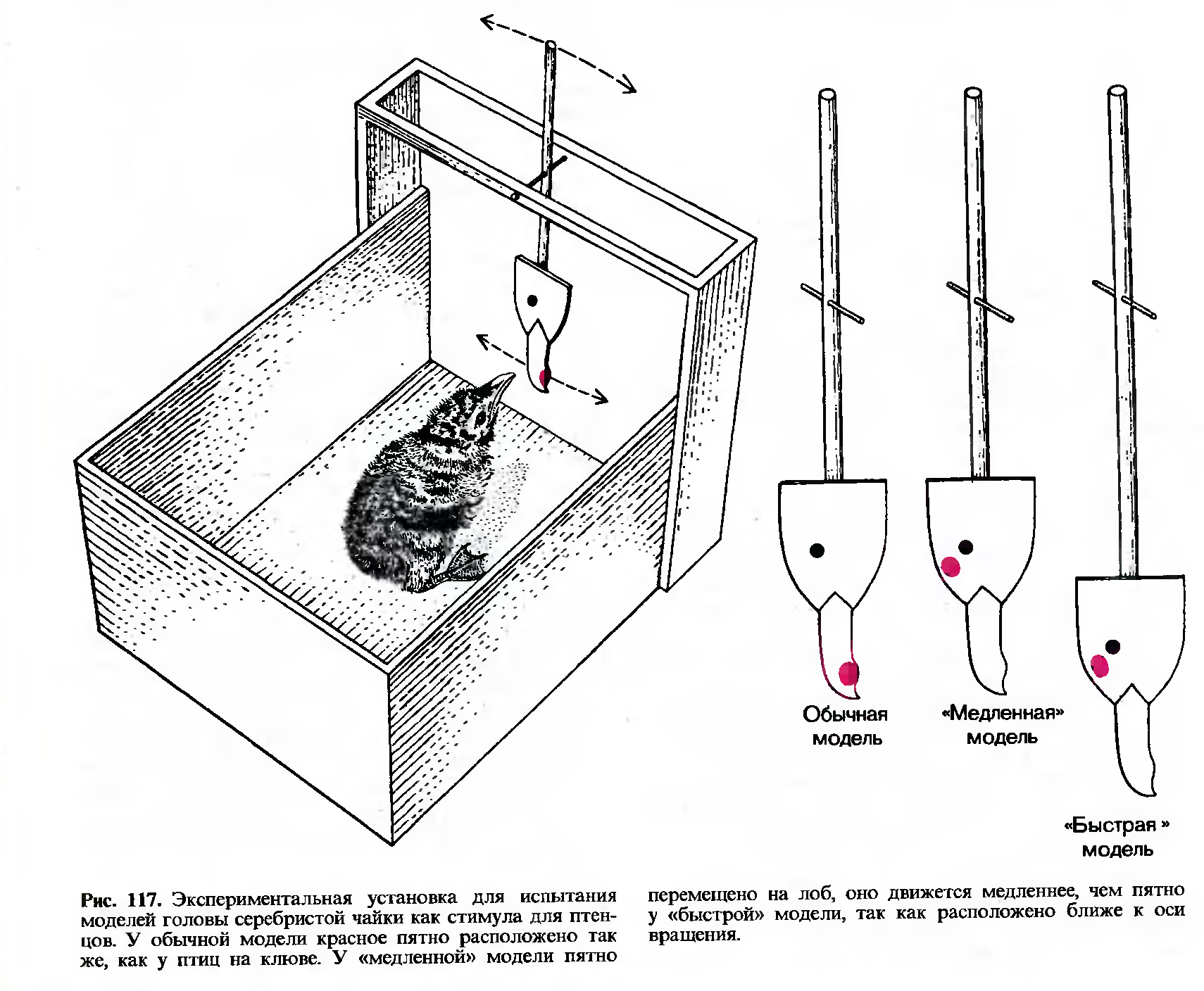 В связи с этим мы решили повторить опыты Тинбергена-Пердека, но добавили еще одну модель. Пятно у этой модели располагалось на лбу, но она закреплялась на стержне таким образом, что пятно оказывалось на таком же расстоянии от оси вращения, как и пятно на клюве у другой модели (рис. 117). Кроме того, наша экспериментальная установка была снабжена подвижным полом, высоту которого подбирали так, чтобы красное пятно на модели всегда оказывалось на уровне глаз птенца. Мы назвали нашу третью модель «быстрой», так как пятно у нее на лбу перемещалось быстрее, чем у «медленной» модели Тинбергена и Пердека.
В связи с этим мы решили повторить опыты Тинбергена-Пердека, но добавили еще одну модель. Пятно у этой модели располагалось на лбу, но она закреплялась на стержне таким образом, что пятно оказывалось на таком же расстоянии от оси вращения, как и пятно на клюве у другой модели (рис. 117). Кроме того, наша экспериментальная установка была снабжена подвижным полом, высоту которого подбирали так, чтобы красное пятно на модели всегда оказывалось на уровне глаз птенца. Мы назвали нашу третью модель «быстрой», так как пятно у нее на лбу перемещалось быстрее, чем у «медленной» модели Тинбергена и Пердека.
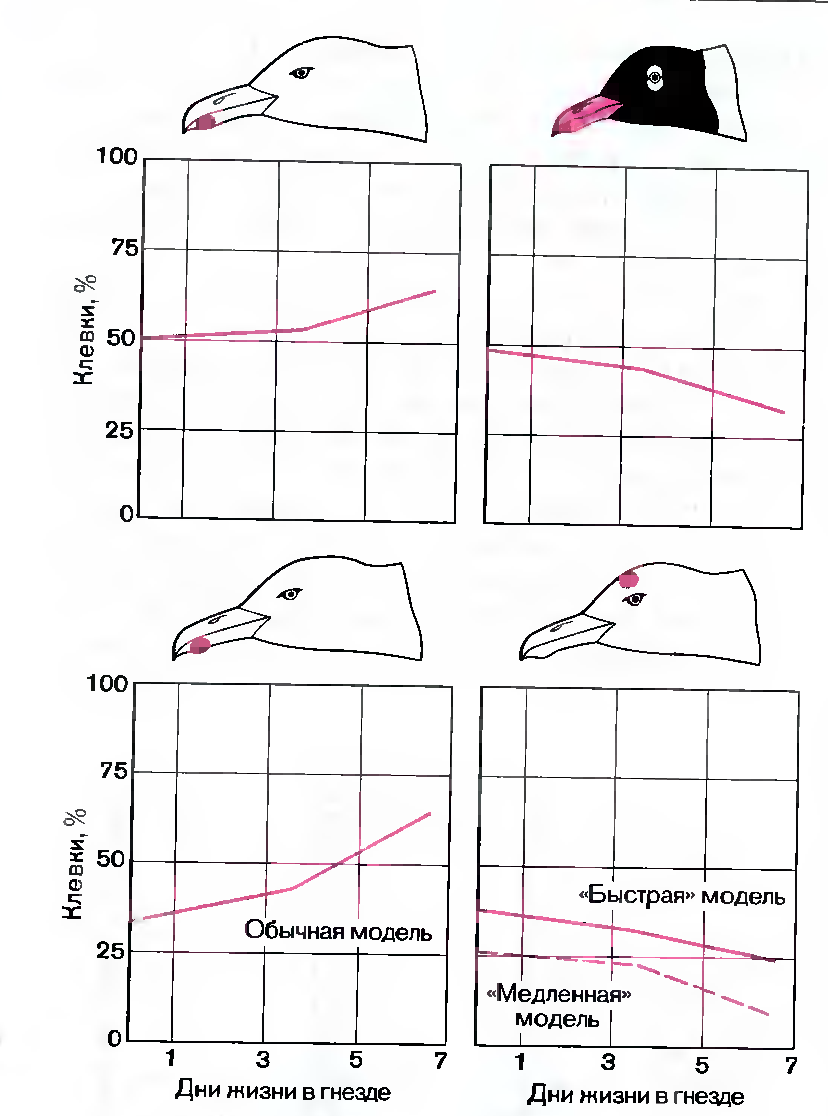
Рис. 118. Реакция птенцов серебристой чайки на различные модели изменяется с возрастом. Вверху слева-модель головы серебристой чайки, вверху справа-смеющейся чайки. Внизу-две модели головы серебристой чайки; у модели слева пятно расположено нормально (на клюве), у модели справа- на лбу.
Если наша гипотеза о роли скорости движения верна, то «быстрая» модель с пятом на лбу должна быть так же эффективна, как и модель с пятном на клюве.
Результаты оказались совершенно однозначными. Только что вылупившиеся птенцы одинаково охотно клевали и «быструю» модель с пятном на лбу, и обычную модель с пятном на клюве. Затем птенцы были возвращены в гнездо и подвергнуты повторным тестам на 3-й и 7-й дни жизни. Как мы и предполагали, реакция на модель с пятном на клюве постепенно улучшалась по сравнению с обоими вариантами модели с пятном на лбу.
Классическая трактовка экспериментов Тинбергена-Пердека сводилась к тому, что существует некий врожденный механизм, включающийся, как полагали, при восприятии определенных довольно сложных стимулов. Наши эксперименты показали, что пищевое поведение птенцов чайки вначале определяется сравнительно простыми стимулами, но позже в результате научения используются более сложные стимулы. Наши эксперименты указывают, видимо, на то, что необходимо заново исследовать и другие случаи, которые считались примерами действия врожденных «запускающих» механизмов.
Наши результаты подсказывают также новую интерпретацию еще одного представления, широко используемого исследователями поведения, — представления о классическом условном рефлексе. В общеизвестных экспериментах И.П.Павлова до начала или во время действия безусловного стимула, вызывающею биологически адекватную реакцию, животному предъявлялся новый для него условный стимул. После некоторого числа таких сочетаний животное начинало отвечать и на отдельно предъявляемый условный стимул. В классических экспериментах Павлова звук звонка подкреплялся пищей. Через некоторое время в ответ на воздействие одного лишь звонка у животного можно было наблюдать выделение слюны.
Психологов давно интересовало, насколько полезной в обычной жизни животного может быть такая способность устанавливать связь между стимулами различных модальностей. Как в эволюции могла выработаться способность к такой форме научения, которая в естественных условиях, по-видимому, так мало используется? Наши результаты наводят на мысль, которую стоило бы проверить. Вспомним о том, что вначале птенец реагирует на самые простые признаки клюва взрослой птицы (безусловные стимулы), но при этом одновременно воспринимает и множество других деталей головы (условные стимулы). Под влиянием пищевого подкрепления эффективным стимулом для клевания становится вес более детализированный зрительный образ.
Такого рода процесс, который я назвал «обострением восприятия», отличается от классического условного рефлекса, наблюдаемого в лаборатории, тем, что условный и безусловный стимулы здесь физически идентичны. Не исключено, что способность к выработке классических условных рефлексов сформировалась в эволюции главным образом как механизм, лежащий в основе феномена обострения восприятия. В этом случае исследователи классических условных рефлексов, по существу, изучают побочные эффекты этого механизма. Но сейчас эти соображения-не более чем одна из возможных гипотез.
Мы изучали и многие другие аспекты пищевого поведения, которые не можем здесь обсуждать из-за недостатка места. Но об одном вопросе необходимо упомянуть: речь идет о распознавании пиши. Узнают ли только что вылупившиеся птенцы пищу, когда сталкиваются с нею? Чтобы выяснить
это, мы поставили четыре чашечки с кормом в углах небольшого ящика и пронаблюдали, как выведенные в инкубаторе птенцы первый раз в жизни находят пищу. Время, затраченное птенцом на поиски пищи, оказалось образно пропорциональным частоте клевков. Этого и следовало ожидать, если исходить из того, что птенцы разыскивают пищу только методом проб и ошибок.
Если птенцам дать насытиться и пересадить их в другое место, пока они не проголодаются, то в повторном опыте они находят пищу гораздо быстрее. В третьем опыте необходимое время сокращается до минимума. Это нельзя объяснить увеличением частоты клевков, так как во втором и последующих опытах она лишь незначительно выше, чем в первом. Следовательно, птенцы очень быстро научаются распознавать пищу или по крайней мере ее местонахождение.
Если вылупившийся птенец вначале не умеет распознавать пищу, значит ли это, что он вынужден полагаться только на метод проб и ошибок, пока, случайно наткнувшись на корм, он не начнет быстро обучаться поискам? Наблюдения и эксперименты показывают, что есть несколько механизмов, помогающих птенцу ускорить первый поиск нищи.
Вспомним, что если птенец не клюет пищу, то взрослая птица обычно берет её в клюв. Я часто наблюдал, как при первом кормлении птенец продолжает клевать клюв родителя и после того, как отрыгнутая пища падает на дно гнезда. В конце концов взрослая птица подбирает пищу, и птенец натыкается на нее при клевке, нацеленном на клюв родителя. Это наводит на мысль об адаптивном значении малой точности клевков только что вылупившегося птенца, так как промахи повышают вероятность того, что он при этом случайно натолкнется на пищу.
Другой механизм, помогающий птенцу быстрее научиться распознавать пищу, связан с присутствием в гнезде других птенцов. Клевки одного птенца часто бывают нацелены на белый кончик клюва другого. Обычно птенцы вылупляются с интервалом около 12 часов, и к моменту вылупления следующего птенца старший уже получает корм. Если младший пытается во время такой кормежки клевать клюв старшего, то он тоже может наткнуться на пищу. Наблюдения за гнездами подтвердили, что иногда птенцы впервые находят корм именно таким способом.
Процесс распознавания корма мы изучали, используя три группы птенцов серебристой чайки, выведенных в инкубаторе. Птенцы контрольной группы помещались в коробки с пищей поодиночке. Птенцы двух экспериментальных групп размещались попарно, но в одном случае второй птенец тоже никогда не видел пищи, а во втором в качестве пары подбирался птенец, ранее кормившийся в коробке.
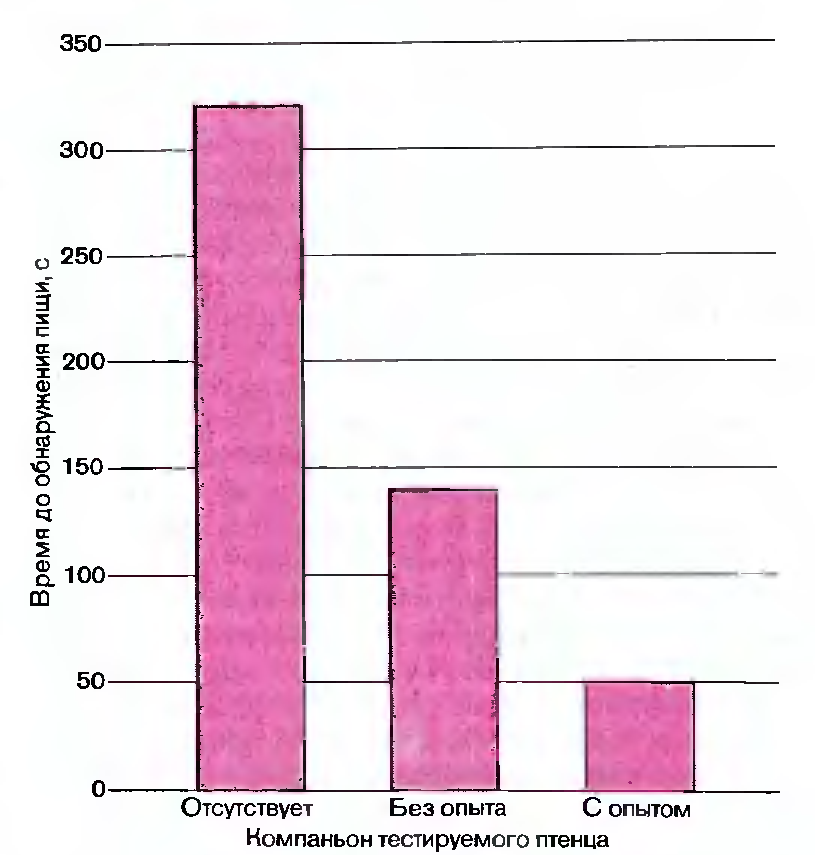
Рис. 119. Только что вылупившиеся птенцы должны были впервые найти пищу в одиночестве и в паре с другим птенцом. Больше всего времени эта задача отнимала
у одиночных птенцов, заметно меньше-у птенцов с неопытным компаньоном и меньше всего в паре с птенцом, уже имевшим опыт нахождения пищи. Результаты
эксперимента показали, что взаимодействие между птенцами помогает им обучаться распознавать пищу.
Оказалось, что птенцы, тестируемые поодиночке, дольше всех не находили пищи. Несколько меньше времени на поиск корма требовалось птенцу, компаньон которого был гак же неопытен, как и он сам. Быстрее всех с задачей справлялся птенец, находившийся в коробке вместе с птенцом, уже умеющим распознавать пищу (рис. 119).
Такие различия нельзя объяснить только более активным «исследовательским» клеванием по принципу проб и ошибок в присутствии компаньона, гак как клюв последнего, наоборот, отвлекал тестируемого птенца от таких проб. Присутствие неопытного птенца побуждало другого больше передвигаться в ящике, и это приводило к более быстрому, чем у одиночных птенцов, обнаружению пищи. Влияние обученного птенца этим не ограничивалось, так как все неопытные птенцы в его присутствии впервые находили пищу, целясь в клюв кормящегося компаньона.
Теперь мы можем резюмировать выявленную нами общую картину развития поведения. Птенец чайки вступает в жизнь, обладая недостаточно координированной, неточной реакцией клевания, определяемой такими простыми признаками стимула, как форма и характер движения (мотивацией
служит голод, источниками стимула — родители и другие птенцы в гнезде). Птенец вначале не способен распознавать пищу, но, промахиваясь при клевках, нацеленных на клюв другой особи, он натыкается на корм и быстро научается узнавать его.
Благодаря пищевому подкреплению птенец запоминает облик родителей. В результате практики повышается точность клевков и совершенствуется оценка расстояния до цели. Птенец научается также поворачивать голову при выпрашивании пиши, что ведет к появлению отличия этой реакции от клевков, направленных непосредственно на пищу.
Обнаруженная картина позволяет думать, что и развитие других инстинктов включает элемент научения. Для формирования стереотипной видоспецифической формы поведения важно только, чтобы процесс научения у всех особей данного вида был очень сходным. На описанном примере с чайками ясно видно, что поведение нельзя осмысленно расчленить на врожденные и приобретенные формы, так же как невозможно выделить какую-либо часть поведенческого репертуара, связанную только с научением. Развитие поведения-это мозаика, обусловленная непрерывным взаимодействием организма и среды.
ЛИТЕРАТУРА
Hailman J.P. The ontogeny of an instinct: the pecking response in chicks of the laughing gull (Larus atricilla L.) and related species, Behaviour, suppl. 15, 1967.
Klopfer P.H., Hailman J. P. An Introduction to Animal Behavior; Ethology’s First Century, Prentice-Hall, Inc., 1967.
Lehrman D. S. A critique of the Konrad Lorenz’s theory of instinctive behavior, The quarterly Review of Biology, Vol. 28, No. 4, 337-363, December 1953.
Tinbergen N.. Perdeck A. C. On the stimulus situation releasing the begging response in the newly hatched herring gull chick (Larus argentatns argentatus Pont), Behaviour, VoL 3, 1-39, 1950.
Источник К.Уэлти, Дж.Сторер, К.Пенникуик и др. Птицы. М.: Мир, 1983. С.136-146.
P.S. Иными словами, J.P.Hailman показывает, что врождённа здесь лишь «болванка инстинкта», его стрежневое движение к верному объекту и в верном направлении. А точность распознавания ключевого стимула, в ответ на который надо это делать, и точность реализации инстинктивного акта «оттачивается» во взаимодействиях птенца и родителя (что Лоренц называл «кругом [социальных] компаньонов», Kumpan).
То же самое происходит при становлении в онтогенезе ритуализированных демонстраций видового репертуара, скажем, ухаживания и угрозы, которые суть инстинктивные акты, запускаемые в ответ на распознавание образа ключевого раздражителя — другого сигнала ряда, т.ч. демонстрации по ходу взаимодействия выступают попеременно релизером и реализацией ответа на него.
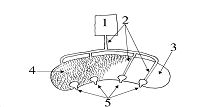
Как показывают тщательнейшие исследования онтогенеза агонистических демонстраций озёрной чайки, выполненные Tom Groothuis с соавт., и здесь дефинитивные формы демонстраций не «созревают» сами собой, но направленно «доводятся» до таковых взаимодействиями в группах чайчат. будущие агонистические демонстрации развиваются ритуализацией движений силовой борьбы, постепенно оказывающиеся всё более «задержанными» и «акцентированными». Но не экспрессивных реакций, отражающих общее возбуждение в процессе борьбы, исходно «демонстративных» и «ярких», но с крайне неустойчивой формой.
Позы угрозы взрослых озёрных чаек. Разные позы связаны с разным «соотношением сил» тенденций к атаке (темные столбцы) и избеганию (светлые столбцы).
Источник: M. Moynihan. Some Aspects of Reproductive Behavior in the Black-Headed Gull (Larus ridibundus L.) and Related Species// Behaviour Suppl., 1955.ведение
На начальных этапах онтогенеза агонистических и брачных демонстраций птиц основным внутренним фактором, определяющим их появление в потоке действий животного, является уровень тестостерона. У 10-дневных птенцов озёрной чайки Larus ridibundus инъекции тестостерона ускоряют проявление ритуализованных демонстраций и одновременно ведут к сильным респираторным проблемам (более чем существенным при интенсивных столкновениях между птенцами), а также к задержкам роста, нарушениям защитной окраски. Временное введение тестостерона вызывало продолжительное, но затем исчезающее проявление демонстративного поведения. При повторных инъекциях тестостерон стимулировал появление в первую очередь именно тех демонстраций, которые при первой стимуляции появлялись чаще всего (Ros et al., 1994).
По мере роста (с возрастом) уровня тестостерона в крови птенцов, подкрепляемого учащением конфликтов в группах, эффект от дополнительной стимуляции гормоном падает, оставаясь значимым лишь для наиболее частых и продолжительных демонстраций. Остальные появляются под воздействием чисто социальных факторов, тем более что чайчата и так достаточно мотивированы к постоянным взаимодействиям друг с другом. При последующем взрослении предъявление демонстраций у молодых чаек полностью освобождается от опосредования тестостероном (Ros et al., 1994).
Вообще первое появление в онтогенезе специфических структур поведения — ритуализованных демонстраций детерминировано эндогенно – гормональными изменениями или сдвигом мотивационного состояния «внутри» особи, а демонстрации первоначально служат лишь выражением внутренних изменений вовне. Далее с накоплением опыта эффективного употребления соответствующего набора демонстраций акт демонстрирования теряет связь с исходной детерминацией «изнутри» (скажем, со сдвигами уровня агрессивности или страха) и делается маркёром некоторой проблемной ситуации вовне, в социальных взаимоотношениях между индивидами.
То есть разные демонстрации «агрессивного» ряда сигналов в процессе индивидуального развития вместо внутренней детерминации оказываются детерминированы социально. Появление каждой из них теперь детерминируется наступлением именно той проблемной ситуации, которую данная демонстрация «маркирует» как сигнал и орудием разрешения которой она служит как деятельность в следующие моменты взаимодействия.
Онтогенетический процесс превращения действий в демонстрации исследовали у озёрных чаек. Использовали метод каспар-гаузер эксперимента: поскольку развитие демонстраций ускоряется в результате контактов с родителями и чужими взрослыми птицами (вторгающимися на гнездовой участок), птенцов выращивали группами по 2-4 до возраста 9 недель.
Затем чайчат из разных групп объединяли и они совместно росли в авиарии (далее они там и размножались). Все три главные агонистических демонстрации («кашлянье», «распластанная» и «наклонная» позы) развиваются непосредственно из движений страха и агрессии путём ритуализации отдельных действий, осуществляющих эти намерения индивида. Агрессивные действия появляются в возрасте около недели, достигают пика около двух недель и снова делаются редкими в 6 недель (примерное время подъёма на крыло). Основные проявления страха – затаивание и бегство; в период пребывания в гнезде частота обоих действий медленно, но постоянно растёт (Groothuis, 1989а-b).
Во внутригрупповых конфликтах молодые чайки защищаются от агрессора эффективней взрослых. Они затаиваются и вблизи него неожиданно подпрыгивают, резко и сильно клюются, после чего отбегают назад. Это типичный конфликт стремлений к нападению и к бегству (присутствующих у животного одновременно), ещё не начавший ритуализироваться. Если агрессор не уходит, но продолжает двигаться вблизи затаившегося птенца, то агрессия пересиливает, и часто неожиданно для него самого (Groothuis, 1989а-b).
По мере превращения агрессора из врага в оппонента и социального компаньона по агрессивным взаимодействиям в данной группе у чайчат происходит ритуализация прямых действий страха и агрессии. Молодые чайки всё чаще и чаще останавливаются после атаки пришельца вместо завершающего клевка, сами действия нападения всё более ингибируются, а завершающий отскок просто исчезает. Отдельные движения становятся всё более «гротескными» и «демонстративными», как будет демонстративным любое резкое движение, если оно вдруг остановится и удерживается «остановленным», не «смазываясь» (без тремора, перерывов и пр.).
Открытая (неритуализованная) агрессия сменяется ритуализованной, из начальных движений нападения развиваются агонистические демонстрации. Первые 5 недель они очень изменчивы по форме. Это именно экспрессивная реакция (выразительное движение), а не демонстрация «нужного» сигнала в «нужной» ситуации, тем более что их появление в потоке действий птицы определяется именно внутренними факторами. Затем форма всех четырёх демонстраций фиксируется и стереотипно воспроизводится в телодвижениях чаек, а их употребление полностью детерминируется социальными факторами (коммуникативным «значением» каждого сигнала в данной группе, Groothuis, 1989a-b).
По мере накопления истории побед и поражений каждой особи у всех чаек в авиарии постоянно увеличивается синхронность и согласованность употребления демонстраций в агонистических взаимодействиях – даже у птиц, контактирующих друг с другом редко или вообще никогда. Это маркирует превращение демонстрации из пантомимы в знак и тем самым завершение процесса эмансипации.
Благодаря эмансипации участники взаимодействия эффективно сигнализируют партнёру о специфических ситуациях ухаживания, угрозы, предупреждения об опасности и пр., даже если не находятся в соответствующем мотивационном состоянии (скажем, недостаточно возбуждены). А партнёр, соответственно, может эффективно реагировать на сигнал независимо от мотивационного состояния и непосредственной «втянутости» во взаимодействие, сильно влияющей на данное состояние. Последнее наиболее важно для «зрителей» процесса – позволяет им эффективно использовать сигналы непосредственных участников взаимодействия (или исходы последнего) для корректировки собственных программ поведения.
Ритуализуясь, движения постепенно становятся всё более «вычурными» и, постепенно обретая типологически определённую форму и отделяясь от действий, а не «кристаллизуются» из «ярких», но крайне «текучих» экспрессивных реакций.
Во-вторых, ключевую роль здесь играют агрессивные взаимодействия, нападения и угрозы в группах птенцов. Направленное «движение» прямых действий агрессии к дефинитивным формам демонстраций происходит лишь при достаточной частоте и интенсивности конфликтов чайчат. Если она ниже, демонстрации оказываются аберрантными или недоразвитыми. Причём «запуск» ритуализации вызван побуждением каждого птенца максимизировать воздействие на других более, чем все они воздействуют на него, реализующимся через выделение общих форм (комплексов действий) в индивидуальных телодвижениях, выступающих предикторами последующего поведения птенца (Groothuis, 1993).
В третьих, здесь образ демонстраций, репрезентируемый партнёру, оказывается всё более специфичным, типологически определённым и точно воспроизводимым в телодвижениях особи именно в связи с усилением их предикторной функции, не только полезной для всех, но и снижающей риски агрессии для демонстратора (Groothuis, 1993; Groothuis, Meeuwissen, 1992).
В четвёртых, действующая причина происходящего – процессы, компенсирующие негативный эффект роста уровня тестостерона при интенсификации взаимодействия в группах чайчат. Этим сохраняется гомеостаз на уровне организма, побочным же следствием – дифференциация форм демонстраций с использованием как предикторов в соответствующих взаимодействиях. См. статьи в приложении:
https://vk.com/doc187880884_445188255?hash=ddced5fbe9..
https://vk.com/doc187880884_445188258?hash=2ba9dc477f..
https://vk.com/doc187880884_445188259?hash=43fc5082e4..
https://vk.com/doc187880884_445188261?hash=0cb4ecb1b7..
Действительно, развитие аберрантных демонстраций у большинства чайчат в малых группах (у некоторых они всё же созревают нормально) – следствие меньшего уровня тестостерона, недостаточно стимулируемого слишком редкими столкновениями и агрессией. Иными словами, тестостерон и множественные физиологические эффекты его действия внутри организма оказываются не самостоятельным фактором формообразования для демонстраций, но лишь передаточным механизмом социальных, формообразующей роли социальных функций. Если их не хватает, демонстрации дифференцируются замедленно или с искажением. Собственно, через подобные «трансмиссии» социальное управляет биологическим везде и всегда.
Как раз этот момент и не понимают биологизаторы, почему biological reductionism — «плохая теория», упускающая из виду важную часть реальности. Врождённость (в смысле генетическую детерминированность) инстинктов не следует преувеличивать. Существенна сама способность животного создавать сложные формы видоспецифического поведения в «нужные» моменты взаимодействия, которые [формы] развиваются без обучения и вообще без какого-либо контакта с конспецификами.
Другой важный момент: при сравнении поведения разных видов «врождённость» означает типологическую определённость форм демонстраций и других комплексов фиксированных действий животного, их видоспецифичность, а не чисто генетическую детерминацию онтогенетического развития таких комплексов фиксированных действий, вроде приводимой последовательности ухаживательных демонстраций селезня кряквы.

В анализе онтогенеза инстинктивного поведения «врождённость» означает устойчивую реализацию специфических форм соответствующего поведения в индивидуальном развитии при исчезающее малой роли социальных влияний, которые в раннем опыте животного могут «дошлифовывать» форму инстинктивного действия до видовой нормы.
Ещё в 1950-е гг., в дискуссии со сравнительными психологами Лоренц утверждал, что все инстинкты врождённы в первом смысле, но многие не врождённы во втором.
«В теле или в поведении любого организма нет абсолютно ничего, что не зависит от внешней среды и в определенной степени не подвержено модификации благодаря окружению… То, что наследуется, — не сам признак, но пределы его модифицируемости» (Lorenz, 1956: 53)».
И далее
«конечно, мы не знаем, насколько то, что мы называем «врожденное поведение» прямо генетически детерминировано и насколько обязано эпигенетическим процессам… Для того, чтобы оценить модифицируемость посредством обучения, несущественно, каким образом определяется «ригидность поведения». Для этой цели нам не надо знать, как много действительно прямо зависит от генов и как много является результатом эпигенеза» (Lorenz, 1956: 67).
Например, если волчонок в подсосном периоде не будет упираться в живот матери (или вообще во что-то), и не может массировать лапками молочную железу при сосании, у него не формируются адекватные формы взрослого пищевого поведения, и в первую очередь запасание пищи (рис., А-Б).
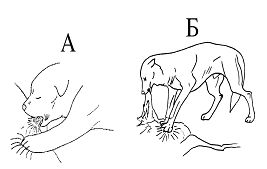 У всех этих животных заметна эмоциональная неуравновешенность, что проявлялось в сравнительно низком пороге оборонительного поведения и частых фрустрациях. Эти индивиды часто создавали в группе (с т.н. нормальными) конфликтные ситуации»
У всех этих животных заметна эмоциональная неуравновешенность, что проявлялось в сравнительно низком пороге оборонительного поведения и частых фрустрациях. Эти индивиды часто создавали в группе (с т.н. нормальными) конфликтные ситуации»
Тут существенно, что для нормального формирования пищевого поведения волчат достаточно надеть на соску диск, чтобы детеныш в него упирался лапками, сделав прокол в соске таким, чтобы молоко не вытекало без существенных усилий детёныша. Тот же эффект даёт использование приспособления, имитирующего тело матери»
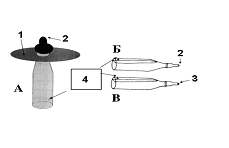
(Я.К.Бадридзе. Волк. М., 2003).
То есть в отличие от внутренних факторов (по которым наблюдается полиморфизм) социальные влияния можно смоделировать искусственно, и развившееся таким образом поведение не отличается от «естественного» по форме или по эффективности, если модель верна. Поэтому главный критерий оценки «врождённости» инстинктов – исчезающее малая роль социальных влияний и/или прошлого опыта животного на формирование соответствующих паттернов поведения после завершения онтогенеза, формирующего те же агонистические или брачные демонстрации.
В отличие от генетиков, этологов не интересует, каковая относительная роль генетических и средовых факторов в самом процессе онтогенеза, формирующем этот инстинкт как «временный орган животного» (а она может очень сильно варьировать). То есть требования к «врождённости» поведения у этологов в норме существенно мягче «генетических»: обладал бы соответствующий паттерн поведения типологически определённой формой сразу по завершении онтогенеза, до систематических социальных влияний и какого-либо личного опыта.
Всё это было чётко проговорено Лоренцем в дискуссии со сравнительными психологами по проблеме «врождённое – приобретённое в поведении» в 1950-е годы (Lorenz, 1955, 1956), но изрядно подзабылось в 1960-е, когда большинство этологов «врождённость» видоспецифического поведения стало понимать просто как «генетическую детерминированность». Это внесло изрядную путаницу, достигшую максимума в книге Р.Хайнда «Поведение животных» (см. Е.А.Гороховская «Этология. Рождение научной дисциплины», 2001).