

В.В. Cуслов, Н.А. Колчанов
В истории естествознания теории эволюции и теории неизменяемости видов одинаково популярны, однако камнем преткновения для всех додарвиновских эволюционистов был поиск движущих сил эволюции. Историческая заслуга Ч. Дарвина – не столько создание эволюционной теории вообще, сколько признание за неопределенной изменчивостью статуса самостоятельного биологического феномена. Эволюция по Дарвину – постоянное преобразование в ряду поколений индивидуальной неопределенной изменчивости, «мало интересной для систематика», в изменчивость дискретных групп – таксонов. Отбор – логическое следствие изменчивости. Таким образом, Дарвин впервые создал теорию эволюции, не требующую особых внешних сил, которые пришлось постулировать его предшественникам.
Однако, каким образом такая непрерывная эволюция приводит к дискретному результату? В организации регуляторных районов генов, активных сайтов белков и в генных сетях присутствуют эволюционно-консервативные модули, задающие пространства возможностей, ограничивающие изменчивость и канализирующие эволюцию. Это позволяет по-новому взглянуть на закон гомологических рядов Н.И. Вавилова и другие случаи параллельной эволюции. Модули самоподдерживаются за счет дупликаций и конвергентной молекулярной эволюции. Выводя из-под отбора часть мутаций, модули способствуют их накоплению в квазинейтральном режиме, подготавливая дальнейшую эволюцию и обеспечивая ее неповторимость.
Введение
Содержание
- 1 Введение
- 2 Эволюционные концепции до и после Дарвина
- 3 Закон гомологических рядов Н.И. Вавилова
- 4 Гомологические ряды и генные сети
- 5 Регулоги – паттерны коэкспрессирующихся генов
- 6 Устойчивость регуляторных модулей в эволюции
- 7 Регуляторные модули и блочно-модульная эволюция
- 8 Эволюция в пространствах возможностей или порядок за счет разнообразия
В 1972 г. палеонтологи Н. Элдридж и С. Гулд сформулировали теорию прерывистого равновесия (пунктуализм), согласно которой в ходе филогенеза большую часть времени таксоны неизменны. Интенсивные морфофункциональные перестройки характерны лишь для относительно кратковременных периодов кладогенеза (эволюционный взрыв), после чего таксон вновь впадает в стазис (Eldredge, Gould, 1972). Большой интерес к этой теории был вызван не столько ее новизной (сходные взгляды высказывались и ранее (Берг, 1922; Симпсон, 1948; Давиташвили, 1970; Грант, 1991)), сколько тем, что авторы вновь привлекли внимание к проблеме дарвиновской эволюционной теории: каким образом непрерывная эволюция приводит к дискретному результату – новым таксонам?
В статье рассмотрены особенности структурно-функциональной организации различных биологических единиц, от макромолекул до генных сетей, и выдвинута гипотеза о существовании в них эволюционно консервативных модулей различной природы.
Мутации и дупликации генов обеспечивают комбинаторику модулей. Эволюция, идущая за счет комбинаторики и экспансии модулей, в локальном масштабе обеспечивает адаптацию, а в долговременном – препятствует утрате модулей. При этом мутации обеспечивают унификацию фенотипа, самоподдерживая модули. Существование модулей позволяет объяснить параллелизм, выявляемый на фенотипическом (гомологические ряды) и молекулярно-генетическом материале.
Обратные связи в модулях уменьшают последствия генетического груза, обнейтраливая изменчивость. Ослабление или разрушение обратных связей проявляется в стабилизации фенотипа – всплеске дарвиновской неопределенной изменчивости. Восстановление обратных связей вновь унифицирует фенотип, переводя таксон из режима эволюционного взрыва в режим стазиса.
Эволюционные концепции до и после Дарвина
С момента зарождения естественных наук вообще и биологии в частности понятия «эволюция» и «отбор» использовались не менее широко, чем представления о неизменности видов (элементов). Термин «эволюция» дословно означает развертывание (от лат. evolutio), иными словами – процесс изменения системы, детерминированный ее исходным состоянием и внешними воздействиями (Dobretsov et al., 2008).
Последние лишь ускоряют или замедляют развертывание. А.А. Любищев (1982) полагал, что в предельном случае (если известны исходное состояние системы и параметры воздействий) изменения системы хорошо предсказуемы – процесс детерминистически воспроизводим. Такая интерпретация эволюции служила и служит неотъемлемым компонентом большинства космогонических систем, начиная от Анаксимандра (ок. 610–540 до н. э.) и кончая современными теориями эволюции Вселенной, звезд, планет, климата, ландшафта и т. д. (Воронцов, 1999). Более того, эволюционизм в естественных науках доминировал, как ни странно, на протяжении большей части европейского Средневековья, по крайней мере, с того момента, когда А. фон Больштедт (1193/1207–1280) оформил основные положения алхимии[1], связав ее с физикой Аристотеля (Джуа, 1966). Опровержение аристотелевой физики Галилеем, Ньютоном и Торричелли (Льоцци, 1970) одновременно привело и к отказу от эволюции[2], после чего в естественных науках надолго утвердились взгляды о неизменности элементов (видов)[3].
Концепция отбора также широко использовалась. В частности, исторический оппонент Ч. Дарвина Ж.-Б. Ламарк считал отбор (хотя и не использовал этот термин) неотъемлемым фактором эволюции. Основные факторы эволюции по Ламарку:
1) стремление живых существ к совершенству, понимаемому как увеличение разнообразия способностей взаимодействовать с внешней средой;
2) это стремление реализуется благодаря наследованию благоприобретенных признаков (адаптивных модификаций – в современной терминологии), в результате упражнения различных органов;
3) результаты такой тренировки передаются потомству у раздельнополых видов при эндогамии лишь при условии подбора пар, тренирующих признаки в одном и том же направлении длительное время[4] (Ламарк, 1911). Именно невыполнение подбора пар у человека объясняет, по Ламарку, отсутствие эволюции современного Homo sapiens (Ламарк, 1911)[5].
Показательно, что упоминание Ламарком отбора (в форме подбора пар) как фактора эволюции совершенно выпало из поля зрения как его последователей, так и критиков[6], поскольку и те и другие рассматривали индивидуальную изменчивость не как важный биологический феномен, а скорее как помеху, мешающую описать некую идеальную особь – диагноз таксона[7]. Оперирование идеальными объектами, позволившее создать экспериментальную биологию, было заимствовано биологами из физики Галилея–Ньютона. Усредненная идеальная особь – носитель диагноза таксона – оставалась центральным объектом биологии от К. Линнея до Ж. Кювье (Кашев, 1963). С точки зрения Ламарка, изменчивость благоприобретенного признака в природных популяциях – лишь следствие разной степени его тренировки (Ламарк, 1911).
Но поскольку общее направление тренировки одинаково для всех, потому что задается внешней средой, одинаковой для всех, признак должен возникать многократно и конвергентно у разных особей. В такой ситуации большинство вариантов подбора пар равнозначны, отсекаются лишь крайние, нежизнеспособные случаи. Сходных взглядов придерживался Л.С. Берг, дополнив общую внешнюю среду (хорономия) общей внутренней средой (белки с их общим набором пространственных конформаций) (Берг, 1922, 1977).
Между тем характерным свойством живого является именно феноменальная изменчивость на всех уровнях его организации. В мире нет двух одинаковых особей, не говоря уже о таксонах. Исторической заслугой Ч. Дарвина является объяснение эволюции именно через феномен биоразнообразия. Отметив на материале домашних животных, что сходные изменения могут наблюдаться при разных внешних условиях так же, как различные изменения – при сходных условиях, Дарвин (1941) впервые рассмотрел изменчивость как самостоятельный важный биологический феномен.
Таким образом, Дарвина можно считать предтечей генетики – в научно-исследовательской программе биологии он подготовил генетике одно из центральных мест. Хотя исследование изменчивости – одно из самых древних направлений[8] биологии (Гайсинович, 1935; Вавилов, 1967а; Воронцов, 1999; Фандо, Музрукова, 2008), отношение к изменчивости как к побочному явлению – неправильности развития или помехе, наведенной средой, привело к вытеснению таких работ на периферию научного знания или вообще в область искусства. Именно поэтому работа Г. Менделя, вышедшая в разгар дискуссий вокруг дарвиновского «Происхождения видов …», осталась неоцененной современниками, хотя и не раз цитировалась (Голубовский, 2001). Также были забыты А. Келликер и К. Негели (Филипченко, 1977), впервые указавшие на роль нейтральных изменений в эволюции[9].
В ламарковской триаде «стремление к совершенству» приписывается таксону наподобие механической силы. В зависимости от того, действует сила или нет, таксон, мыслимый как нечто монолитное, может эволюционировать. По Дарвину, таксон состоит из множества различающихся элементов (особи или семьи, как у общественных насекомых (Дарвин, 1991)), элементарные акты взаимодействия которых («борьба за существование») определяют эволюцию[10], которую можно определить как преобразование в череде поколений индивидуальной неопределенной изменчивости, «мало интересной для систематика» (Дарвин, 1991), в изменчивость групповую и далее в константные таксономические признаки (Левонтин, 1978).
Это преобразование идет под действием отбора – дифференциального переживания, которое само есть следствие биоразнообразия. На место идеализированной особи Дарвин поставил реально существующие популяции – группы особей (иначе невозможно выявить изменчивость), связанные родством (иначе невозможно выявить наследственность). Эволюция, таким образом, является неотъемлемым сущностным свойством таксона: в триаде «изменчивость–отбор–наследственность» все три члена логически связаны. Такая логическая стройность и предопределила как успех теории, так и бурную дискуссию вокруг нее. В ходе дискуссии, которую можно определить как «эволюция: да или нет», данные Менделя, касающиеся константных 11 нейтральных признаков[11] (Мендель, 1935), не прибавляли аргументов ни противникам, ни сторонникам дарвинизма, поэтому остались невостребованными.
Дарвиновская постановка проблемы потребовала пересмотреть определение эволюции. В его теории эволюция является детерминистически невоспроизводимым и потому преимущественно дивергентным процессом (Дарвин, 1991). В ее ходе одна закономерная траектория развития спонтанно и случайно заменяется на другую – одну из множества возможных. Вероятностный характер замены следует из определения неопределенной изменчивости, под которой Дарвин понимал признаки с неустойчивым, варьирующим в череде поколений и от особи к особи проявлением (с широкой нормой реакции в современных терминах) (Дарвин, 1991) и отличал их от внезапного, случайного изменения признака, константно наследуемого в череде поколений – «спорта»[12].
Однако введение такой случайной категории, как неопределенная изменчивость, в качестве базового элемента научной теории, более того, картины развития мира, настолько шло в разрез с общепринятыми научными концепциями, что вызвало интуитивное отторжение значительной части научного сообщества[13]. Вся последующая драма идей вокруг дарвинизма, как это отмечали Филипченко (1977) и Воронцов (1999), по сути, связана с попытками либо ограничить, либо вовсе исключить базовый элемент случайности. Таким образом, как только дискуссия «эволюция: да или нет» сменилась на дискуссию «неопределенная изменчивость: да или нет», законы Менделя обрели актуальность и вскоре были переоткрыты.
Сам Дарвин первоначально рассматривал неопределенную изменчивость лишь как изменчивость, механизмы и закономерности возникновения которой предстоит изучить, а затем был вынужден отступить на позиции ламаркизма (теория пангенезиса). Результаты поисков закономерностей неопределенной изменчивости Дарвин (1941) обстоятельно изложил в работе «Изменение животных и растений в домашнем состоянии». В частности, в ней был систематизирован накопленный на тот момент в науке материал по межпородным и межвидовым скрещиваниям и по плодовитости гибридов. На качественном уровне Дарвин описал доминирование, наследование, связанное с полом, и расщепление гибридов в F2, но при этом остался равнодушен к числовым соотношениям[14]. Оставаясь верным себе, Дарвин искал законы именно варьирования признаков (экспрессивности и пенетрантности в современной терминологии), а не комбинаторики константных признаков (Darwin, 1877; Дарвин, 1941). Теория пангенезиса предлагала такой, пусть и умозрительный[15], механизм.
Справедливости ради надо заметить, что и в генетике вопрос о закономерностях экспрессивности и пенетрантности признаков был поставлен более чем через 20 лет после переоткрытия законов Менделя (Timofeeff-Ressovsky, 1927; Вавилов, 1967б). Первые генетики, напротив, отказались не только от неопределенной изменчивости, но и от отбора вообще (мутационная теория Коржинского –де Фриза (Коржинский, 1899)). Впоследствии отбор был восстановлен, прежде всего в рамках балансовой теории С.С. Четверикова (Левонтин, 1978). В синтетическую теорию эволюции отбор вошел как сито, фильтрующее мутации (по сути – дарвиновские «cпорты») и рекомбинации (Грант, 1991). Таким образом, константная изменчивость, которая исходно, по Дарвину, являлась результатом взаимодействия неопределенной изменчивости и отбора, в рамках СТЭ служит пассивным материалом для отбора и должна накапливаться пропорционально дивергенции таксонов.
Закон гомологических рядов Н.И. Вавилова
Между тем сравнение уровней молекулярной, морфофизиологической и биохимической дивергенции говорит об отсутствии такой корреляции. Так, несмотря на большие морфологические и поведенческие различия между человеком и шимпанзе, полногеномные сравнения дают оценки межвидовых различий в пределах 1–2 %, т. е. не выше, чем между шимпанзе и другими человекообразными обезьянами (Wildman et al., 2003; Glazko et al., 2005).
В то же время сравнение генов-ортологов позволяет выявить консервативные и быстро эволюционирующие функциональные группы генов. Так, у млекопитающих быстро и адаптивно эволюционируют гены иммунитета, гены, связанные с репродукцией, гены ольфакторных рецепторов и гены модификации хроматина. У человекообразных к ним добавляются гены транспорта (ионный транспорт, липидный транспорт, транспорт О2), гены нервной системы (в том числе гены допаминовой и серотонинэргической систем), транскрипционных факторов и раннего эмбриогенеза, в частности, ответственные за развитие антериорной части тела (Wildman et al., 2003; CSAC, 2005; Arbiza et al., 2006) (рис. 1).

Объяснение см. в тексте
Полногеномные исследования заставили пересмотреть монофилию таксонов, выстроенную на эмбриональном, морфологическом и биохимическом сходстве. Например, для эукариот оспорено единство ракообразных и их родство с насекомыми (Шаталкин, 2003; Cook et al., 2005; Regier et al., 2005). У прокариот поставлена под сомнение монофилия такой консервативной группы, как цианобактерии (Захаров-Гезехус, 2008). Аналогичные примеры есть в частной систематике любого крупного таксона (млекопитающие (Asher, 2007), птицы[16] (Edwards et al., 2005) и т. д.).
Сходные результаты дал многолетний эксперимент по доместикации. Высокий уровень изменчивости у экспериментально доместицируемых видов (лисицы, норки) демонстрировал параллелизм с домашними животными и даже птицами, тогда как генетический анализ не выявил соответствующего генетического разнообразия.
Напротив, разные новации имели сходную генетическую основу (Беляев, 1987; Трапезов, 2008; Трут, 2008). Во всех этих группах дестабилизация фенотипа объясняется общим вектором отбора, направленным на слом нейрогуморальных регуляторных систем (Беляев, 1987; Науменко, 1988; Шульгина, 2007). Однако дестабилизация затронула сравнительно небольшую фракцию генов, включающую функциональные группы формирования скелета, габитуса и онтогенеза (Маркель, Бородин, 1990; Шумилина, 2007; Трут, 2008), транспорта, систем рецепции сигналов внешней среды (нервная система, сезонный ритм, поведение) (Беляев, Трут, 1983; Попова, 2002), метаболизма (Маркель, 2008) и размножения (Науменко и др., 1983). Таким образом, под отбор во многом попали те же группы генов (рис. 1, столбец 10, подчеркнуты), что и при эволюции гоминидной линии приматов..
Параллельная и независимая эволюция в тех же функциональных группах генов отмечена и в линии низших узконосых обезьян (рис. 1, ветка 1, столбец 9). Наконец, аналогичные быстро эволюционирующие функциональные группы генов можно выявить и у доместицированных растений[17] (Paterson et al., 1995; Гончаров, 2007): гены архитектоники, плодовитости, транспорта, регуляции обмена между внутренней и внешней средой (гены рецепции сигналов, метаболизма экзогенных субстратов (Glazko, 2003), гены устойчивости к неблагоприятным факторам (Cooper et al., 2003)[18]. На рис. 1 в столбце 10 эти функциональные группы отмечены курсивом.
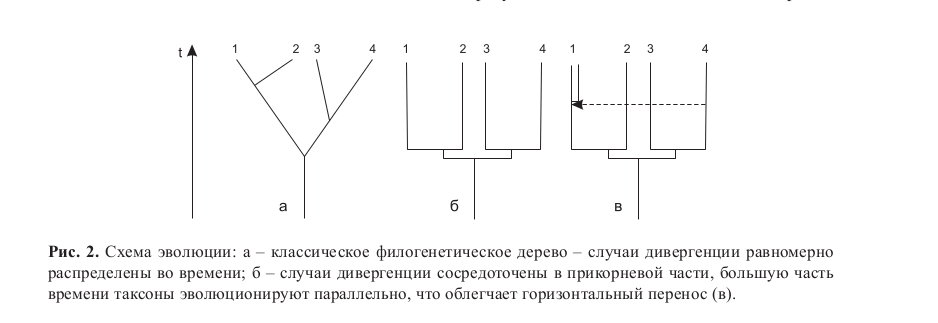
Таким образом, налицо не только общий вектор отбора, но и параллельные ряды функционально общих модулей, на которые он периодически действует. С учетом этих данных картина эволюции больше соответствует не классическому дереву с длинным стволом и ветвями разной длины (рис. 2, а), а «кусту», разветвляющемуся у самого корня. Большинство веток «куста» имеют сходную длину и эволюционируют параллельно (рис. 2, б). Такая параллельная эволюция, в частности, позволяет успешно обмениваться генами различным видам прокариот (Ochman et al., 2000; Шестаков, 2003), в компактном геноме которых чужой ген может дать адаптивное преимущество, лишь если он не разрушит уже сложившуюся корегуляцию генов (Tamames, 2001; Шестаков, 2007; Захаров-Гезехус, 2008) (рис. 2, в).
Наличие общих функциональных групп генов препятствует чрезмерной дивергенции таксонов. Время от времени интенсивная эволюция затрагивает определенные группы генов, порой повторно у одного и того же таксона. Часто у разных таксонов это одни и те же функциональные группы, обогащенные генамиортологами (Kumar et al., 1996; Wildman et al., 2003; Evans et al., 2004; Katoh Y., Katoh M., 2005;Shi et al., 2006; Гунбин и др., 2007).
В статье мы попытаемся ответить на два взаимосвязанных вопроса:
1) чем обусловлена устойчивость функциональных групп генов в филогенезе, несмотря на различные локальные векторы отбора каждого отдельного таксона;
2) почему при постоянном накоплении мутаций и сохранении этой устойчивости эволюция идет скачками.
Обнаруженная картина эволюции заставляет обратиться к закону гомологических рядов Н.И. Вавилова (Вавилов, 1920, 1935):
1) виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предсказать параллельные формы других видов и родов;
2) семейства характеризуются определенным циклом изменчивости, проходящим через все роды и виды.
Н.И. Вавилов структурировал признаки, входящие в гомологические ряды, в форме радикала – набора фенотипических признаков, записанных последовательно по степени константности их проявления (Вавилов, 1920, 1935).
Таким образом, признаки, фенотипическое проявление которых константно у одного вида, часто константны и у родственных видов. Напротив, варьирующие признаки вида, вероятно, будут варьировать и у его родственников (закон Уолша (Walsh, 1864; Любищев, 1957; Дарвин, 1991)). Согласно Вавилову, морфотип любой популяции можно описать радикалом. Его левая часть – константные признаки – будет у всех особей практически одинаковой, характеризуя таксон высокого ранга – род, семейство, отряд. Правая часть – признаки с широкой нормой реакции – распадется по степени варьирования на группы, соответствующие низшим таксонам (от подвида до жорданона – монотипической популяции) (рис. 3). Так, например, в радикале злаков, такие качественные признаки, как «остистость/безостость», «голозерность» должны войти в левую часть радикала, а широко варьирующие признаки «окраска зерна», «высота соломины», «кустистость», «засухоустойчивость» – в правую. Позже Вавилов отметил, что порядок признаков в радикале близких таксонов может меняться.

Так, «стекловидность зерна», обычно широко варьирующая, оказалась константной для персидской пшеницы. Для одних рас пшеницы окраска колоса и стебля может быть константной, для других – варьирующей. Часто такая изменчивость носит хорошо выраженный клинальный характер (так, высота соломины параллельно увеличивается у разных рас к северу и по мере поднятия в горы) (Вавилов, 1967б).
Ценность этого взгляда в том, что уникальное сочетание константных и варьирующих признаков «физиономически» характеризует вид, причем в отличие от современных концепций вида константные и варьирующие (интересовавшие Дарвина) признаки одинаково важны, а репродуктивная изоляция, если она есть, – лишь один из признаков в радикале (Вавилов, 1931; Vasilyeva, 1999).
Формирование радикала вида в эволюции Н.И. Вавилов объяснял через взаимодействие генетической составляющей дарвиновской неопределенной изменчивости предковой популяции с разнообразием контрастных физико-географических факторов внешней среды в пределах небольшого ареала (Вавилов, 1931). Этому удовлетворяли центры происхождения культурных растений (Вавилов, 1965), приуроченные к гористым районам с пересеченной местностью и разнообразными, часто контрастными условиями. В таких обстоятельствах вся наличная изменчивость небольших редко взаимодействующих популяций-полуизолятов имела наилучший шанс проявиться и быть закрепленной отбором[19]: из первоначального полиморфизма выделялась левая, константная часть радикала.
Тогда его правая часть – либо незавершенный эволюционный процесс, либо признаки, полиморфизм которых выгоден (или безразличен) виду (системе популяций – линнеону) в целом (Вавилов, 1931; Мейен, 1978). Н.И. Вавилов не успел закончить разработку классификации признаков злаков по степени константности проявления (Вавилов, 1967б), поэтому определение радикала в его работах (Вавилов 1920, 1935) осталось схематичным. Позже С.В. Мейен, базируясь на законе гомологических рядов, предложил мероно-таксономический метод классификации. Признаки объединяются функционально и/или по общности положения в классы – мероны (Мейен, 1977). Примеры меронов на уровне генов – в столбце «Биологическая функция» таблицы рис. 1. Каждый мерон может входить составной частью в мерон более высокого ранга и делиться на мероны низкого ранга. Ряд меронов, объединенных в различных таксонах общим правилом преобразования, образует рефрен (Мейен, 1977, 1978). Строки таблицы на рис. 1 можно рассматривать как рефрены меронов генов, подчиняющиеся кимурианскому критерию адаптивной эволюции.
Гомологические ряды и генные сети
Генетической основой радикала, по Н.И. Вавилову, в равной мере служили общий пул гомологичных генов и общее сходство онтогенетического пути от гена до признака. Позднее Г.Ф. Гаузе (2002), сформулировав обратную зависимость между приспособляемостью (адаптивной модификацией) и приспособленностью, показал, что обе характеристики могут взаимно компенсировать друг друга, обеспечивая константность фенотипа. И.И.Шмальгаузен (1968) предположил, что уменьшение степени варьирования (переход признаков из правой части радикала в левую) может происходить именно за счет стабилизации онтогенетического пути, ведущего к одной из возможных модификаций.
Отбор на устойчивость ее фенотипического проявления должен привести к фиксации различных, необязательно гомологичных, мутаций. М.М. Камшилов на дрозофиле смоделировал этот процесс, а также показал влияние межгенных взаимодействий на амплитуду модификаций (при введении мутации eyeless в различные генотипы) (Камшилов, 1935, 1939, 1940а, б, 1974). Таким образом, вавиловский радикал должен опираться не только на совокупность генов, но и на граф связей между ними, а также – для варьирующих признаков – на состояние критических параметров внешней и внутренней среды[20]. Параллелизм может затрагивать любой из этих уровней (Мейен, 1978). С игнорированием этого системного момента связаны неудачи многих попыток сопоставить гомологическим рядам морфологических признаков молекулярно-генетические морфологические ряды.
В современной терминологии сходство морфологических признаков должно отражать сходство не отдельных генов, а генных сетей – функциональных групп координированно экспрессирующихся генов. Любая генная сеть (ГС) имеет:
1) группу генов («ядро»), обеспечивающую выполнение ее функций;
2) центральные регуляторы – транскрипционные факторы, организующие гены «ядра» в координированно экспрессирующиеся кассеты генов при взаимодействии с общими сайтами связывания их регуляторных районов;
3) рецепторы, запускающие работу ГС в ответ на внешние сигналы;
4) пути передачи сигналов с рецепторов ГС на ее центральные регуляторы.
Часть продуктов генов «ядра» ГС также играют роль сигналов, взаимодействуя с рецепторами как собственной ГС, так и других генных сетей и формируя регуляторные контуры с положительными и отрицательными обратными связями (Колчанов и др., 2000).
Режим функционирования ГС определяют:
(а) наборы молекул-регуляторов и
(б) набор генов «ядра» сети,
(в) граф сети – структура связей между ее компонентами,
(г) состояние критических параметров внешней и внутренней среды и
(д) начальное состояние переменных генной сети.
Таким образом, в генных сетях можно выделить три иерархических информационных уровня: функциональный (а и б), чьи закономерности связаны с конкретными молекулярными механизмами взаимодействия молекул; структурный (в); и параметрический (г и д) (Колчанов и др., 2000; Лихошвай, Колчанов, 2008).
Морфологические признаки, представляя законченный результат онтогенетического процесса, хорошо визуализуются. Визуализация генных сетей менее очевидна – в разное время один и тот же ген может участвовать в разных кассетах или в разных генных сетях, изменения начальных параметров меняют режим функционирования генных сетей. Таким образом, фенотипическим гомологическим рядам логичнее сопоставлять не сами генные сети, а их состояния в тот или иной момент онтогенеза.
Регулоги – паттерны коэкспрессирующихся генов
Состояние генной сети – своего рода ее моментальный снимок – можно оценить по микрочиповым профилям коэкспрессии. Паттерны коэкспрессии, консервативные в пределах цветковых растений, образуют, например, гены, входящие в генную сеть формирования цветка (рис. 4, а). В пределах таксона Metazoa консервативно коэкспрессируются гены, входящие в генные сети циркадного ритма (рис. 4, б) или Hh-каскада (рис. 4, в). У прокариот консервативно коэкспресссируются гены базовых генетических процессов и основных метаболических путей (Teichmann, Babu, 2002, 2004).
Для консервативных модулей коэкспрессии был предложен термин «регулог» (regulog). Регулоги были обнаружены как у про-, так и у эукариот. У прокариот гены, входящие в регулог, часто сохраняют свое взаиморасположение и положение на кольцевой хромосоме, что связано с оперонной структурой генома прокариот (Teichmann, Babu, 2002; Захаров-Гезехус, 2008). У эукариот в ряде случаев отмечена связь регулогов и определенных районов хромосом (Lindsay et al., 1999; Paterson et al., 2000; Taddei et al., 2001; Teichmann, Babu, 2002; Pauli et al., 2006).
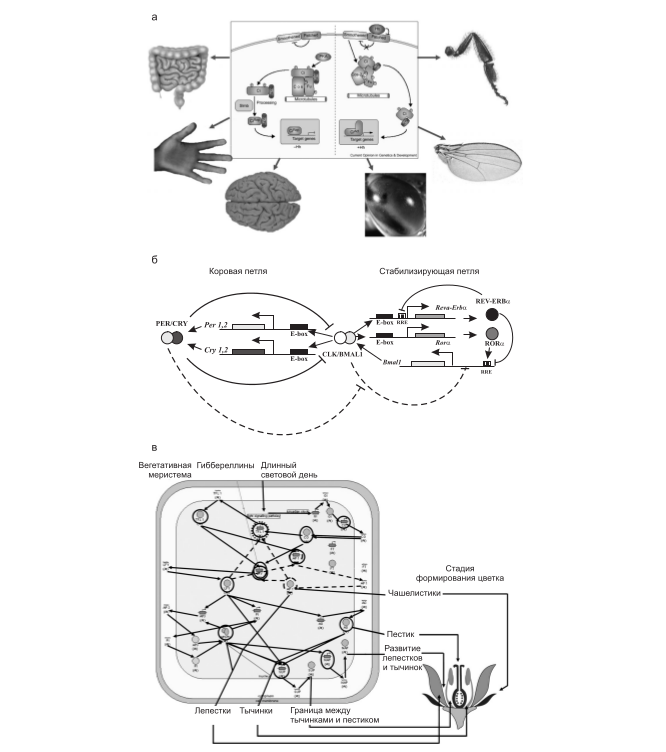
Рис. 4. Примеры трех генных сетей, гены которых формируют консервативные паттерны коэкспрессии у разных таксонов – регулоги: а – генная сеть Hh-каскада (по: Nybakken, Perrimon, 2002); б – генная сеть циркадного ритма (по: Badiu, 2003); в – генная сеть морфогенеза цветка (по: Honma, Goto, 2001; Theissen, Saedler, 2001).
Для выявления регулогов между удаленными таксонами используют несколько стандартных процедур. При использовании только микрочиповых данных сначала проводят поиск генов-ортологов в сравниваемых таксонах, затем отобранные гены в каждом отдельно взятом таксоне тестируют на коэкспрессию в микрочипах при контрастных условиях (разные условия среды, разные стадии онтогенеза и т. п.). Полученные выборки генов для каждого отдельного таксона сравнивают со случайной выборкой, а затем проводят стандартный тест на робастность микрочипового метода, удаляя часть выборки и повторяя всю процедуру заново.
Выборки коэкспрессирующихся генов, прошедшие всю процедуру, считают регулогами, принадлежащими одной генной сети (Stuart et al., 2003). Такое сравнение 3182 микрочиповых экспериментов ортологов Homo sapiens (6591 ген), Drosophila melanogaster (5802 гена), Caenorhabditis elegans (5180 генов) и Saccharomyces cerevisiae (2434) выявило 22163 консервативных случая коэкспрессии. Они распределились по 12 консервативным паттернам, состав генов в которых коррелировал с хорошо изученными генными сетями (Stuart et al., 2003), как древними (сети «домашнего хозяйства» и базовых процессов – клеточного цикла, трансляции, транскрипции, протеосомной деградации белков, липидного метаболизма), так и молодыми (например, сети нейрогенеза).
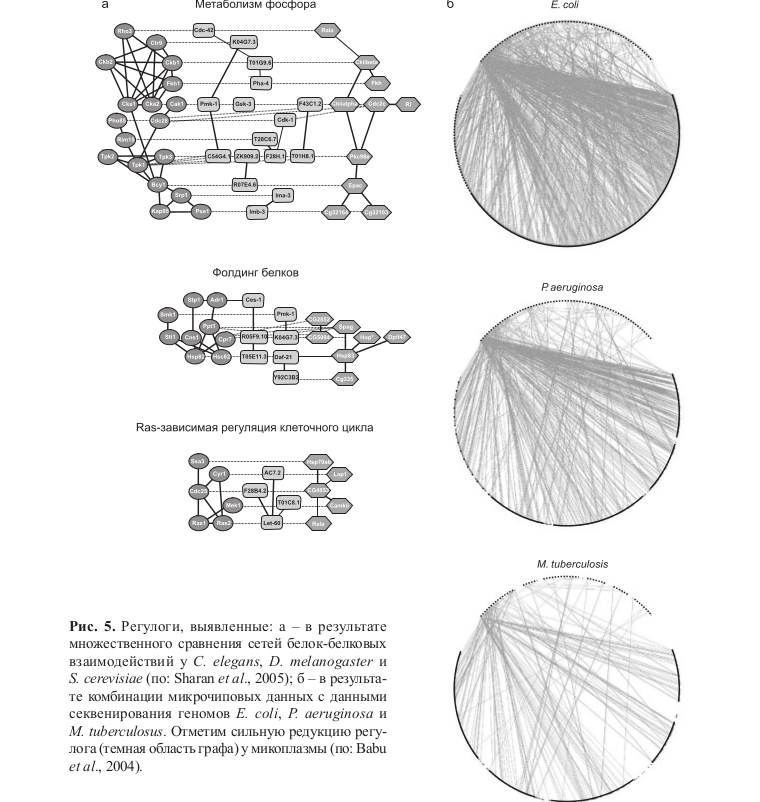
Стандартной процедурой выявления регулогов также является сравнение микрочиповых данных, полученных в контрастных условиях, с данными белок-белковых взаимодействий (Sharan et al., 2005) (рис. 5, а) или с данными секвенирования генов: гены, демонстрирующие устойчивую коэкспрессию и регулируемые общим специфичным транскрипционным фактором, считаются принадлежащими к общему регулогу (Snelet al., 2004; Teichmann et al., 2004) (рис. 5, б).
Паттерны коэкспрессии можно разделилить на три типа:
- Генным сетям базовых процессов «домашнего хозяйства» соответствуют паттерны с высококонсервативным составом и коэкспрессией ортологов, а также с высокосвязным графом ГС;
- Паттерны, в которых состав ортологов варьировал при переходе от дрожжей к многоклеточным (сети регуляции транскрипции, межклеточной коммуникации, а также сети сложных белковых комплексов – рибосомы, протеасомы), связаны с появлением новых функций;
- Паттерны, в которых консервативность состава ортологов не соответствует консервативности коэкспрессии (нейрогенез), связаны с дивергенцией функций в эволюции (Chervitz et al., 1998; Peregrin-Alvarez et al., 2003; Stuart et al., 2003; Bergmann et al., 2004).
Три типа паттернов коэкспрессии заставляют вспомнить вавиловский радикал: тип 1 соответствует левой части, тип 3 – правой части с неустойчивым (варьирующим) фенотипическим проявлением, тип 2 – признакам, которые невозможно включить в радикал на данном таксономическом уровне. Замечательно, что для генов с неизвестной функцией ассоциация с такими паттернами позволяет предсказывать функцию (Stuart et al., 2003; Huynen et al., 2004), что взаимодополнительно к закону гомологических рядов.
Для многоклеточных эукариот функциональный паттерн коэкспрессии распадается на регулоги отдельных клеточных типов и стадий дифференцировки. Так, в общем регулоге, ответственном за сперматогенез, регулог, объединяющий гены, экспрессирующиеся в клетках Сертолли и митотических сперматогониях, выявляется также в различных соматических тканях (в частности, в него входят выше упоминавшиеся гены липидного транспорта).
Отдельные частично перекрывающиеся регулоги можно выявить для разных мейотических и постмейотических стадий созревания сперматозойда (Eddy, 2002; Сhalmel et al., 2007).
Нужно отметить, что у мейотических и постмейотических регулогов также отмечено минорное перекрывание с соматическими регулогами (в частности мозг) и более широкое – с малигнизированными клетками (Schlecht, Primig, 2003; Сhalmel et al., 2007; Jacobs et al., 2007; Jacobs et al., 2008).
В регулогах можно выделить «ядро», содержащее гомологичные гены (как ортологи, так и паралоги) и «шлейф» негомологичных видоспецифичных генов. Размеры «ядра» могут сильно варьировать по составу генов и/или по степени коэкспрессии (Bergmann et al., 2004).
Так, «ядро» небольшой генной сети циркадного ритма у Metazoa (рис. 4, б) консервативно (Badiu, 2003) – мутации в гене казеинкиназы эпсилон I укорачивают циркадный ритм у дрозофилы, грызунов и человека, снижая уровень фосфорилирования белка PER (Vanselow et al., 2006). Мутации по другим казеинкиназам также сохраняют гомологичность, хотя направленность нарушений (удлинение или укорочение циркадного ритма) в разных видах может не совпадать (Xu et al., 2005).
Размер «ядра» связан с эволюционной историей регулога. Так, регулог пролиферации клеток в глобальных генных сетях человека и дрозофилы имеет хорошо выраженное консервативное ядро. Напротив, регулог дифференциации клеток распадается на перекрывающиеся стадиеспецифичные регулоги, варьирующие как по составу генов, так и по графу взаимодействия между ними, что свидетельствует об их быстрой эволюции (Sharan et al., 2005; Xia et al., 2006). Анализ глобальных генных сетей показал, что наиболее консервативными являются регулоги, содержащие гены, продукты которых функционируют в составе мультимерных белковых комплексов (Sharan et al., 2005).
Менее консервативными оказались регулоги генных сетей онтогенеза и дифференцировки. Напротив, генные сети метаболических путей оказались слабоконсервативными, так как содержали очень небольшое консервативное «ядро» с генами собственно метаболизма и большой варьирующий шлейф регуляторной компоненты (Teichmann, Babu, 2002; Bergmann et al., 2004).
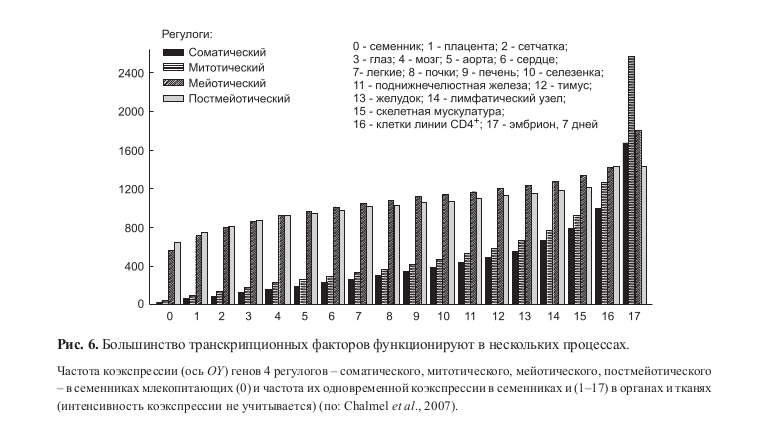
Анализ регулогов показал, что среди транскрипционных факторов сравнительно немного как строго тканеспецифичных, так и функционирующих на всех стадиях онтогенеза. Большинство транскрипционных факторов участвуют в нескольких процессах и экспрессируются на нескольких стадиях онтогенеза (рис. 6) (Teichmann, Babu, 2002; Luscombe et al., 2004; Сhalmel et al., 2007; Jacobs et al., 2008).
 Устойчивость регуляторных модулей в эволюции
Устойчивость регуляторных модулей в эволюции
Регулоги имеют иерархическое строение, собираясь из небольших блоков (рис. 7, а) в ходе выполнения той или иной функции. Анализ таких блоков позволил выявить типовые регуляторные мотивы (Babu et al., 2004; Bergmann et al., 2004; Luscombe et al., 2004) (рис. 7, а). Рассматривая иерархическую организациию генных сетей с позиций регуляции транскрипции, заметим, что низшей ступенью иерархии является функциональная связка «транскрипционный фактор–сайт его связывания в ДНК» (Babu et al., 2004). В результате комбинаторики таких пар и образуются регулоги, комбинаторика которых в свою очередь дает регуляторные мотивы ГС.
Мотив может быть представлен (рис. 7, а) транскрипционным фактором, взаимодействующим с кассетой сайтов; триггероподобной структурой, основанной на взаимодействии двух идентичных транскрипционных факторов и сайтов их связывания; композиционным элементом, состоящим из пары взаимодействующих сайтов связывания (Babu et al., 2004) и др.
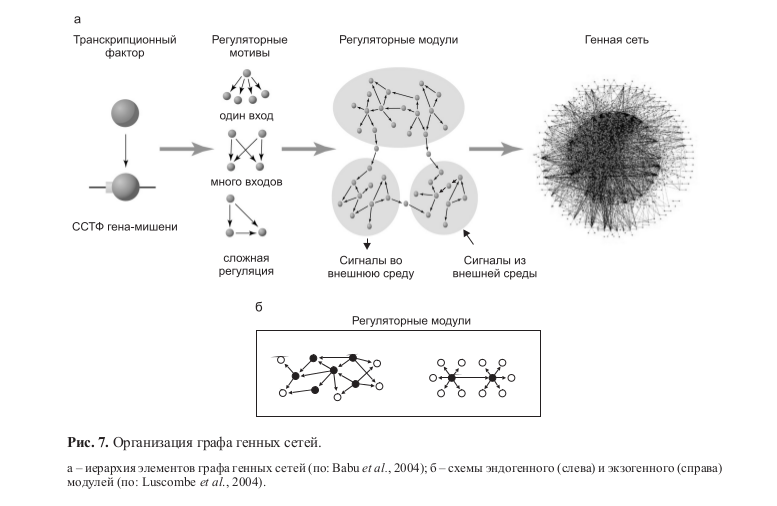
Комбинаторика мотивов, в свою очередь, образует регуляторные модули, которые, объединяясь за счет генерируемых ими сигналов, формируют регуляторную машину глобальной генной сети организма (Babu et al., 2004; Wittkopp, 2007). Возможность такой комбинаторики обеспечивается общим пулом регуляторных молекул, а ограничение на комбинаторику в эволюции определяет степень перекрывания регулогов, проявляемая в фенотипе как плейотропные эффекты мутаций.
Модули, выполняющие эндогенные (гены домашнего хозяйства и базовых метаболических функций, гены онтогенеза) и экзогенные (рецепция и передача сигнала, стрессовый ответ, регуляция метаболизма) функции, обогащены различными регуляторными мотивами. Экзогенные модули обогащены генными кассетами, регулируемыми единственным центральным регулятором. Эндогенные модули обогащены замкнутыми регуляторными контурами и генными кассетами, включающими множество коактиваторов центрального регулятора (рис. 7, б) (Luscombe et al., 2004).
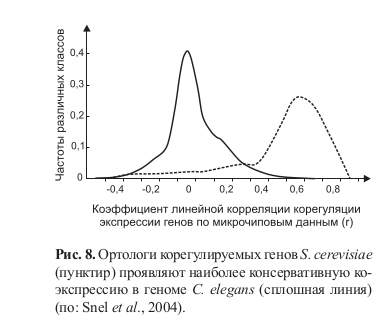
Snel и соавторы (Snel et al., 2004) провели попарный внутри- и межвидовой анализ коэкспрессирующихся корегулируемых генов. Критерием корегуляции генов считался высокий коэффициент их коэкспрессии (r > 0,6) при наличии в их регуляторных районах общего сайта связывания транскрипционного фактора. У S. cerevisiae было выявлено 975 таких пар. В выборке коэкспрессирующихся генов C. elegans ортологи таких пар оказались в правом хвосте распределения по величине коэффициента коэкспрессии (рис. 8).
Высокая степень коэкспрессии ортологов корегулируемых генов S. cerevisiae позволяет предположить, что они сохранили корегуляцию и в геноме C. elegans. Наиболее впечатляющие свидетельства эволюционной устойчивости небольших регуляторных модулей получены при изучении паттернов коэкспрессии у насекомых и хордовых. Так, сходный набор ортологов регулирует формирование фасеточных глаз насекомых и сложных камеральных глаз хордовых (Gehring, 2005). Фасеточные глаза фиксируются палеонтологами не позднее кембрия (~570–550 млн лет назад), но согласно палеонтологическим и эмбриологическим данным, предки хордовых никогда не обладали такими глазами.
Развитые камеральные глаза впервые появляются в ордовике (500 млн лет назад) независимо у головоногих и хордовых (Михайлова, 1997). Следовательно, консервативные модули коэкспрессии, ответственные за формирование глаз, возникли еще раньше, возможно, в венде (~ 650 млн лет назад), а потом независимо рекрутировались в ходе эволюции.
Интересно, что для эктопического развития функционирующих фасеточных глаз или (обратный эксперимент) – тканей камерального глаза (Gehring, 2005) оказалось достаточно трансгенеза единственным транскрипционным фактором Pax-6.
Попав в различные ткани, Pax-6 – один из центральных регуляторов ГС развития глаза – рекрутировал из разнообразного генетического окружения гены, участвующие в формировании глаз, по сути, восстановив генную сеть. В морфогенез крыльев насекомых и парных конечностей хордовых также вовлечен общий набор генов – центральных регуляторов, сохранивших сходный паттерн экспрессии. Такой неожиданный результат можно объяснить, лишь предположив, что для формирования крыльев и ног в эволюции был независимо вовлечен древний консервативный регулог, предназначенный для формирования билатерально-симметричных придатков тела.
Он образовался, судя по совокупности палеонтологических (появление билатерально-симметричных складок, а затем преобразование их в конечности (Михайлова и др., 1997)) и молекулярно-биологических (филогения отдельных генов (Шаталкин, 2003)) данных, незадолго до кембрийского взрыва (~ 550 млн лет назад) у общего предка Metazoa.
По сравнению с первичноротыми дорзо-вентральная ось хордовых инвертирована, поэтому ноги и крылья соответствуют придаткам одной и той же стороны, иннервировавшимся одной и той же нервной системой (Reichert, Simeone, 2001). Комбинаторика регуляторных модулей в генных сетях позволяет объяснить быстрое и неоднократное формирование новых органов в эволюции. Так, исследование профилей коэкспрессирующихся генов в плаценте Eutheria показало практически полное отсутствие регуляторных генов с плацентоспецифическим профилем экспрессии.
Дифференцирующаяся плацента оказалась обогащена регулогами с древними генами, ортологичными для эукариот и/или хордовых в целом и ответственными за базовые клеточные процессы. Зрелая плацента обогащена регулогами, включающими таксон-специфичные (от отряда и ниже) гены-паралоги, ответственные за негативную регуляцию ростовых и метаболических процессов. Образование паралогов – результат дупликаций, сопровождавших дивергенцию отрядов и семейств плацентарных (Cross et al., 2003; Knox, Baker, 2008).
Таким образом, в ходе эволюции плацента Eutheria была сформирована блочно-модульным способом из уже готовых генных сетей или их фрагментов[21]. В дальнейшем в ходе эволюционного становления характерной для каждого таксона репродуктивной стратегии (многоплодие или малоплодие; тип потомства «птенцовый» – рождение недоразвитых детенышей – или «выводковый» – детеныши способны следовать за матерью и др.) в ряде таксонов произошло параллельное становление идентичных типов плацент, видимо, тем же блочно-модульным способом.
Так, человекообразные рекрутировали в плаценту ретровирусный блок, связанный с формированием синцитиотрофобласта (Knerr et al., 2004). Эпителиохориальная плацента параллельно развилась у китов, парнокопытных, непарнокопытных, панголинов и лемуров (Сarter, Enders, 2004). Такая плацента обеспечивает низкотравматичные для самки роды малоплодного «выводкового» потомства (адаптивно для видов, испытывающих пресс хищников, либо агрессивной внешней среды).
Древние генные сети, вовлеченные в становление живорождения у млекопитающих, продолжают существовать у остальных хордовых. Таким образом, предположение Л.С. Берга (1977) о параллельном возникновении живорождения в неродственных таксонах вновь обретает актуальность (с поправкой на роль отбора) и хорошо согласуется с концепцией маммализации (Татаринов, 1976) – параллельного развития маммального комплекса признаков в нескольких линиях терапсид.
Регуляторные модули могут не только иметь транскрипционную природу, но и формироваться на базе связок «миРНК–сайт ее связывания в мРНК». Например, miR319 регулирует экспрессию генов семейства TCP у столь отдаленных растений, как томат и арабидопсис (Palatnik et al., 2003). При повышении интенсивности взаимодействия миРНК с мРНК-мишенью формируется лист с изрезанным краем (норма для томата, мутация для арабидопсиса), а при низкой интенсивности – цельнокрайний лист (мутация для томата, норма для арабидопсиса). Важно, что фенотипические проявления мутационного нарушения такой регуляции сходны, хотя молекулярные механизмы совершенно различны: у томата (рис. 9, а) снижение интенсивности взаимодействия получено за счет мутации, меняющей сродство к миРНК сайта-мишени мРНК гена LA (семейство TCP), а у арабидопсиса (рис. 9, б) интенсивность взаимодействия повышена вследствие усиления экспрессии miR319 (мутация jaw).

Регуляторные модули и блочно-модульная эволюция
При независимой адаптивной эволюции по многим локусам под негативный отбор хотя бы по одному из них попадет каждая из особей популяции, что ведет популяцию к гибели (дилемма Холдейна). При таком ограничении актуальным становится блочно-модульный тип эволюции – комбинаторика относительно автономных генных сетей, уже «апробированных» отбором в ходе предшествующего филогенеза (Ратнер и др., 1985; Ратнер, 2002). Следовательно, раз возникнув и будучи вовлечены в блочно-модульную эволюцию, регуляторные модули (ген-центральный регулятор и его мишени) способны длительно сохраняться в филогенезе дивергировавших таксонов, многократно и параллельно рекрутируясь в генные сети как гомологичных, так и негомологичных органов.
Сохранение таких модулей позволяет объяснить гомологические ряды, выявляемые на современном (Вавилов, 1935; Воронцов, 1967; Vasilyeva, 1999), ископаемом (Соболев, 1924; Обручев, 1972; Meyen, 1978; Рожнов, 2006) и эмбриологическом материале (Raff, Sly, 2000; Richardson et al., 2002), глубокий параллелизм на уровне генов-центральных регуляторов между негомологичными органами (Шаталкин, 2003; Gehring, 2005) и низкие оценки различий по молекулярно-биологическим данным между морфологически далекими видами (Wilson et al., 1974; Wildman, 2003; Glazko et al., 2005).
Напротив, локальные адаптации таксонов обеспечиваются за счет комбинаторики модулей и дупликации генов в составе модуля. Одним из ограничений на комбинаторику модулей является плейотропный эффект мутаций по содержащимся в них генам.
С.В. Мейен (1975, 1977) в 1970-е гг. подчеркивал, что отношения гомологии могут связывать лишь мероны одного и того же уровня иерархии. Его предвидение оправдалось: в настоящее время становится бессмысленным говорить о гомологии, не указывая уровень рассмотрения (экоценотический, морфологический, кариологический, геномный, молекулярно-генетический). Плеойтропный эффект может быть уменьшен благодаря дупликациям и/или формированию белков-коактиваторов транскрипционных факторов (часто благодаря тем же дупликациям с субфункционализацией одного из паралогов (McCarrey, Riggs, 1986; de Souza et al., 2005; Sharma et al., 2006)). Таким образом, комбинаторика модулей должна сопровождаться дупликациями составляющих их генов.
Полногеномный анализ Drosophila melanogaster, Caenorhabditis elegans, Arabidopsis thaliana, Saccharomyces cerevisiae и Sсhizosaccharomyces pombe зафиксировал такие таксонспецифические всплески дупликаций именно на ранних стадиях дивергенции таксонов (по молекулярным часам) (Lespinet et al., 2002).
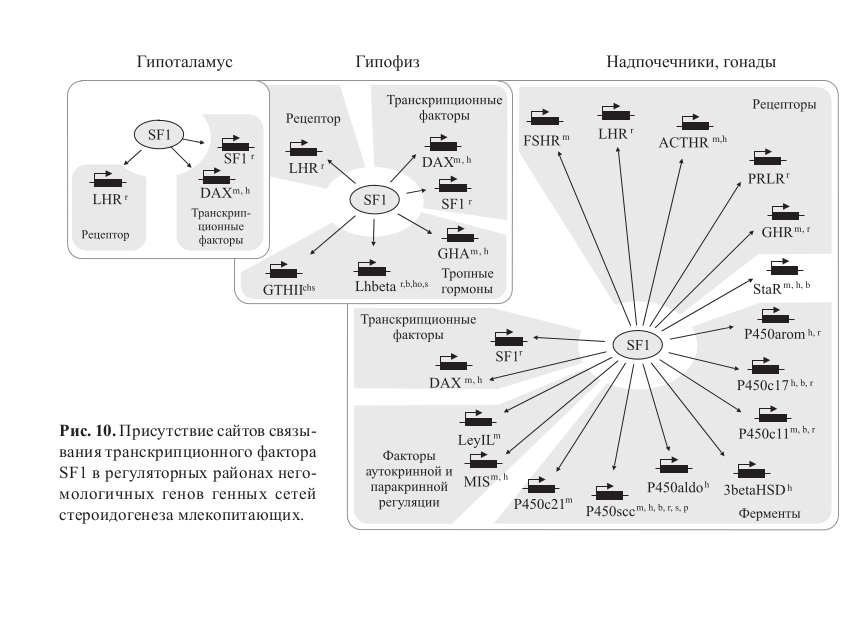
Важную роль в экспансии регуляторного модуля играет также конвергентное возникновение сайтов связывания транскрипционных факторов, чему благоприятствуют небольшие размеры таких сайтов. Даже одиночные нуклеотидные замены способны существенно менять величину активности сайтов связывания – от полной ее потери до выраженного увеличения, а также приводить к появлению активных сайтов в ранее нефункциональной ДНК (Ponomarenko et al., 1999, 2001). Благодаря таким заменам под контроль транскрипционного фактора ставятся негомологичные гены, формирующие модули, как это, например, имеет место для фактора SF-1, работающего в ГС стероидогенеза (рис. 10) (Колчанов и др., 2005).
Таким образом, локальные адаптации таксонов связаны с экспансией модуля в их глобальных генных сетях. В ходе экспансии снижается вероятность утраты модуля. Одновременно модуль начнет рекрутировать гены из ранее независимых генных сетей, если на них действует общий фактор отбора (Берг, 1993а, б). В ходе такой коэволюции стабилизирующий отбор будет поддерживать смену функциональной связи на генетически закрепленную (Шмальгаузен, 1968) (например за счет конвергентного формирования сайтов связывания транскрипционных факторов). При параллельной экспансии в репродуктивно изолированных таксонах рекрутироваться будут как гомологичные, так и негомологичные гены – начнется формирование «ядра» и «шлейфа» регулога.
В глобальной генной сети организма такие регулоги должны формировать своеобразное пространство возможностей, ограничивающее выбор траекторий развития, тем самым канализируя дальнейшую эволюцию. Таким образом, высокое разнообразие на молекулярно-генетическом уровне будет унифицировано на уровне фенотипа.
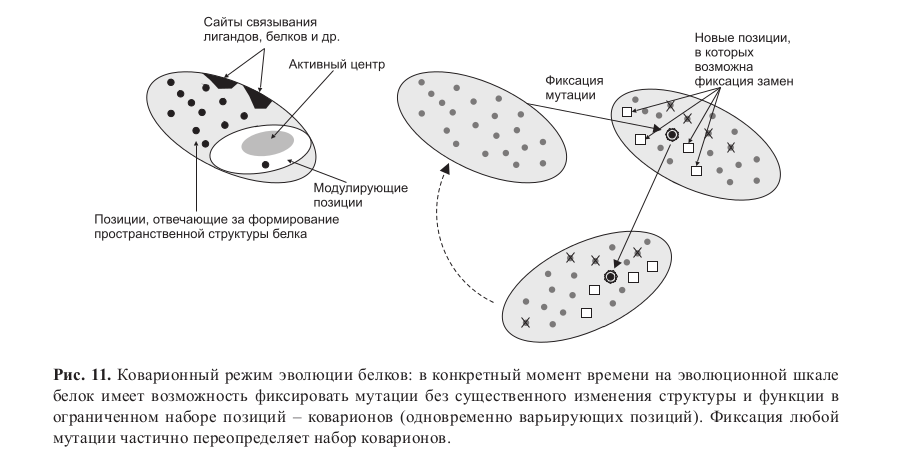
Пространство возможностей в макромолекулах В пространственной структуре белков также можно выделить консервативное «ядро» – позиции, формирующие функциональные центры, позиции, необходимые для правильной укладки глобулы, и «шлейф» – позиции, модулирующие активность центров, позиции белок-белковых взаимодействий, лидерные последовательности и др.
При этом доля позиций, в которых возможна фиксация мутаций без повреждения структуры и функции белка, как правило, невелика (до 20 %). В каждый конкретный момент времени на эволюционной шкале в белке имеется лишь ограниченный набор позиций – коварионов (одновременно варьирующих позиций), способных фиксировать мутации без существенного изменения структуры и функции (Miyamoto, Fitch, 1995). В этом режиме эволюции фиксация любой мутации частично переопределяет набор коварионов и слегка модулирует функцию белка (рис. 11).
Исследования показывают, что в ходе коварионной эволюции происходит многократный перебор пространства возможностей. Например, один из механизмов ускользания вируса гриппа от иммунного ответа – образование сайтов связывания ионов в антигенной детерминанте. Ион, чуть меняя локальную конформацию, мешает преципитации антител. Небольшие размеры белка ограничивают количество антигенных детерминант и вероятных сайтов связывания ионов.
В итоге у близкородственных штаммов вируса гриппа А сайты связывания ионов занимают различное положение – перебор пространства возможностей только начат, а у дальнородственных – сходное: перебор пространства возможностей завершен и завершен цикл изменчивости[22] (Иванисенко и др., 2008).
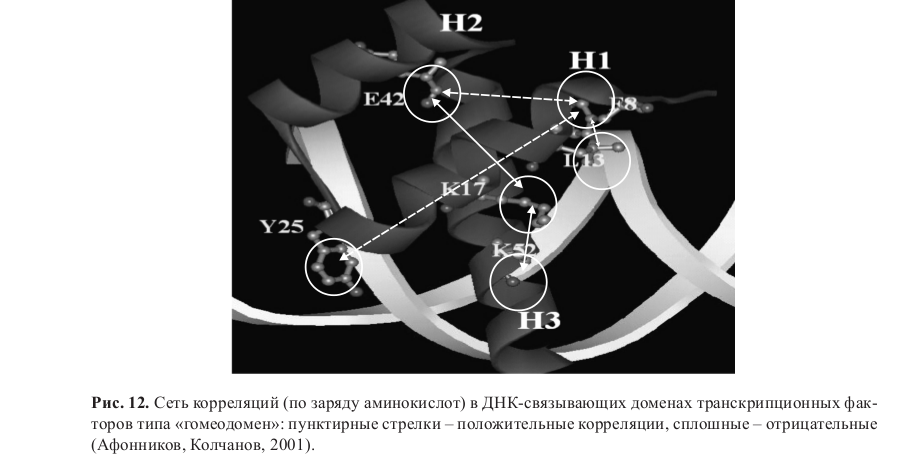
Таким образом, для молекулярного фенотипа белка, как ни странно, действительно подходит концепция Л.С.Берга с поправкой на роль отбора. Аминокислотные остатки, участвующие в формировании трехмерной структуры белков, образуют сеть взаимодействий, ограничивающую эволюцию. Единичные мутации ухудшают функциональность белков, повышая вероятность фиксации компенсирующих замен (рис. 12) (Афонников, Колчанов, 2001, 2008).
Режим эволюции коадаптивных замен приводит к тому, что разнообразие на уровне первичных структур белков повышается, а на уровне третичных структур унифицируется. Так, высокий суммарный положительный заряд молекулы белков-протаминов, компактизующих ДНК в сперматозоиде, стабилизируется коадаптивными заменами: мутация, элиминирующая основную аминокислоту, повышает вероятность фиксации компенсирующей мутации. В итоге характерный для протаминов высокий уровень dn/ds – кимурианский критерий адаптивной эволюции (Wyckoff et al., 2000) – слабо связан с изменением их функции, которая находится под стабилизирующим отбором[23] (Rooney et al., 2000).
Аналогичные соображения о существовании консервативного «ядра» и модулирующего «шлейфа», унифицирующих изменчивость на уровне пространственных структур за счет накопления ее на нижележащих уровнях и тем самым формирующих пространство возможностей эволюции, будут справедливы для любых биополимеров, рабочие структуры которых формируются за счет взаимодействия ограниченного множества мономеров. Так, коадаптивные замены унифицируют консервативную структуру рРНК за счет фиксации конвергентных и неконвергентных замен в неродственных таксонах (Алешин и др., 1999).
Эволюция в пространствах возможностей или порядок за счет разнообразия
Итак, путь от гена до признака можно представить как иерархию различных пространств возможностей: а) на уровне биополимеров; б) локальных генных сетей и в) глобальной генной сети организма. Эти пространства возможностей формируются модулями сходной структуры: всегда существует высококонсервативная часть, cложившаяся на ранних этапах эволюции, и низкоконсервативная часть – следствие локальных адаптаций таксонов.
В генных сетях высококонсервативной части соответствует «ядро» регулога, низкоконсервативной – «шлейф»; в белках – соответственно функциональные центры и их окружение; в олигонуклеотидах (РНК, сайты связывания транскрипционных факторов на ДНК) – коровый район и фланги.
Такой инвариант структуры самоподдерживается за счет несинонимичных мутаций, одновременно значительно снижая мутационный груз – переводя эти мутации в режим отбора, близкий к нейтральному. Сходный инвариант можно наблюдать и на экосистемном уровне, рассматривая экологическую нишу не просто как гиперобъем в многомерном пространстве факторов и ресурсов, заданный границами толерантности представителей вида (Hutchinson, 1965), а как вероятность заполнения такого гиперобъема с учетом компенсации ресурсов по Рюбелю (Rubel, 1935) и кондиционирования одним фактором другого (Беклемишев, 1970). Области гиперобъема, соответствующие наиболее комфортному сочетанию факторов/ресурсов и заполненные наиболее густо, образуют «ядро», менее комфортные – «шлейф».
Кроме того, можно выделить области, где взаимодействие факторов делает невозможным существование любого представителя вида (значения факторов по отдельности остаются переносимыми), и области экстремальной периферии, доступные лишь для отдельных популяций либо требующие для существования особей запуска генных сетей стрессового ответа. Стресс смещает границы толерантности вида, поэтому возможна ситуация, непредусмотренная Дж. Хатчинсоном: выход области экстремальной периферии за границы фундаментальной экологической ниши вида.
В результате наблюдается непрерывное накопление изменчивости при константном фенотипе. Получается парадоксальная картина: мутации, являясь источником изменчивости, могут унифицировать фенотип[24], обеспечивая морфофизиологический стазис. Такая картина хорошо согласуется с фактом неравномерности темпов морфофизиологической и молекулярной эволюции (Wilson et al., 1974; Wildman, 2003; Glazko et al., 2005).
В глобальной генной сети организма мутации и дупликации обеспечивают также экспансию регуляторных модулей – многократное рекрутирование функциональной связки «транскрипционный фактор – сайт его связывания» в локальные генные сети, отвечающие за различные функции (Teichmann, Babu, 2004). В организме, у которого есть несколько взаимодействующих генных сетей, одновременно функционирующих в онтогенезе, но активирующихся различными центральными регуляторами, стабилизирующий отбор будет поддерживать такую экспансию, поскольку она усиливает интеграцию и автономизацию организма, уменьшая число независимо отслеживаемых параметров. Механизмами экспансии будет конвергентное образование сайтов связывания в регуляторных районах негомологичных генов, дупликации сайтов связывания и, наконец, дупликации самих генов.
В итоге такой экспансии одна из сетей превратится в генную сеть – интегратор, регулирующую остальные, в которых, в свою очередь, существенно изменится граф взаимодействий и пул транскрипционных факторов. При этом на фенотипическом функциональном уровне практически ничего не меняется – все сети по-прежнему работают вместе. Смена транскрипционных факторов в генных сетях регуляции синтеза рибосомальных белков у аскомицетов (Gasch et al., 2004; Tanay et al., 2005) подтверждает такой сценарий. При изучении регулогов синтеза рибосомальных белков оказалось, что пул регулируемых белков практически не менялся, зато набор сайтов связывания в их промоторах варьировал от вида к виду так же, как и пул транскрипционных факторов.
Аналогичные рассуждения справедливы и для генных сетей, функции которых перекрываются во времени (например на смежных стадиях онтогенеза). В итоге экспансия модуля может изменить структуру генных сетей раннего эмбриогенеза, эволюционно сформировавшихся раньше самого модуля, что и было выявлено при анализе генных сетей эмбриогенеза (Gilbert et al., 1996; Richardson et al., 1997; Raff, Sly, 2000; Jeffery et al., 2002). Таким образом, вероятность потери регуляторного модуля в ходе эволюции мала, чем и можно объяснить гомологические ряды функциональных групп генов, устойчивые на протяжении миллионов лет в репродуктивно изолированных таксонах, и отсутствие полной гомологии генов в таких группах.
Эволюция, идущая за счет комбинаторики и экспансии модулей, в локальном масштабе обеспечивает адаптации, а в долговременном – препятствует утрате модулей. Результатами экспансии регуляторного модуля в глобальной генной сети организма в результате длительного стабилизирующего отбора будут также уменьшение фракции ткане-, органо- и стадиеспецифичных транскрипционных факторов в протеоме (Teichmann, Babu, 2002; Luscombe et al., 2004; Сhalmel et al., 2007; Jacobs et al., 2008) и формирование графов генных сетей, обладающих свойствами «малого мира» (Ravasz, Barabasi, 2003; Bergmann et al., 2004), в которых относительно немного высокосвязных узлов обеспечивают быстрое распространение сигнала по генной сети.
Обратным следствием экспансии модулей будет повышение чувствительности организмов к плейотропным мутациям. Изменение вектора отбора со стабилизирующего на движущий или дестабилизирующий должно поддерживать дезинтеграцию генных сетей. Ее механизмом могут служить дупликации отдельных генов и целых модулей (мегадупликации районов хромосом (Piskur, 2001)) и, наконец, полиплоидизация генома в целом. Затем следует дивергентная специализация паралогов, сопровождаемая в случае полиплоидизации генома утратой части генов (Walbot, 2000; Wang, Gu, 2000; Sharma et al., 2006). Вспышки дупликаций и циклы «полиплоидизация генома«–«утрата генов» действительно предшествовали и/или сопровождали кратковременные «эволюционные взрывы» во многих таксонах (Walbot, 2000; Wang, Gu, 2000; Paterson et al., 2004; Sharma et al., 2006).
Другим следствием стабилизирующего отбора будет формирование в составе генных сетей регуляторных контуров с отрицательными обратными связями (ООС). В простейшем контуре с ООС, регулирующем концентрацию белка, параметр «отклонение концентрации от нормы» отслеживается регуляторным звеном ООС, меняющим скорость синтеза белка. Таким образом, ООС сужает норму реакции, выводя из-под отбора любое изменение на нижележащих уровнях иерархии (рис. 13а, б), где могут накапливаться мутации, эволюционирующие в квазинейтральном режиме. При движущем отборе ситуация противоположна.
В незнакомых условиях среды преимущества получают организмы с широкой нормой реакции (дарвиновской неопределенной изменчивостью), т. е. таксоны без ООС или таксоны с положительной обратной связью (Колчанов, Шиндялов, 1991) (рис. 13а, б).
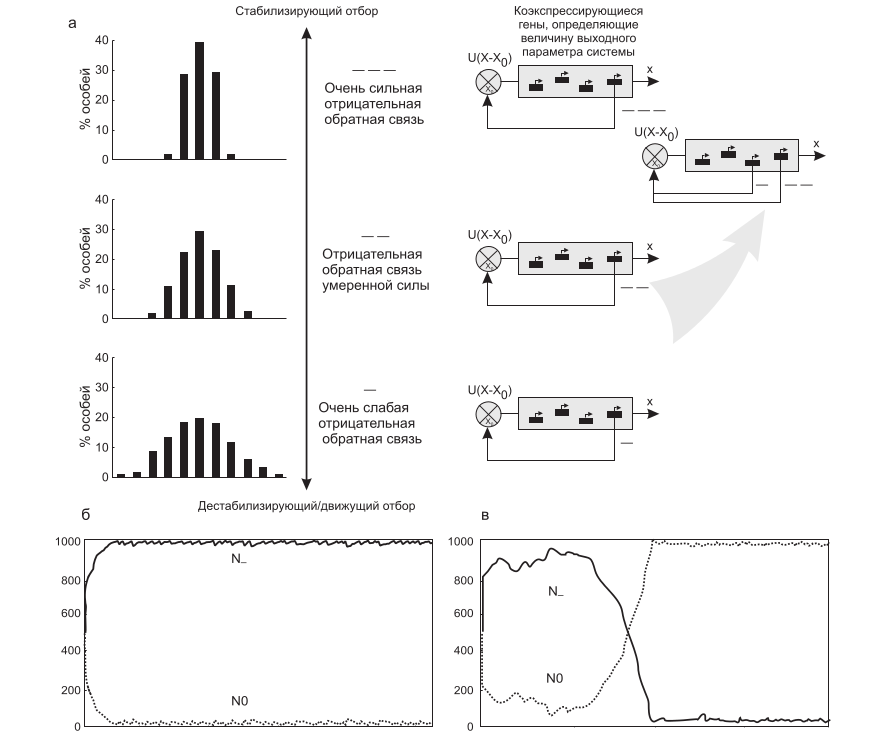
Рис. 13. Отрицательные и положительные обратные связи в эволюции (см. текст). а – «обнейтраливание» изменчивости отрицательной обратной связью: слева – ОХ – спектр фенотипической изменчивости, ОY – частота разных особей фенотипов; справа – схема усложнения обратной связи в эволюции; б, в – конкуренция особей с отрицательной обратной связью (N-) и без нее (N0) в ходе эволюции популяции при стабилизирующем (б) и движущем/дестабилизирующем отборе (в). ОХ – количество эволюционных шагов, ОY – число особей (численность популяции постоянна – 1000 особей). В начальный момент 50 % особей имеют контур с отрицательной обратной связью (N-) и 50 % – не имеют (N0).
Слом ООС приведет к одновременному проявлению всех ранее обнейтраленных мутаций (гиперманифестация изменчивости). Все накопленные мутации одновременно попадут под отбор, поэтому, согласно дилемме Холдейна, будут тестироваться случайно подобранными ансамблями по одному из параметров («адаптивная оптимизация») (Суслов и др., 2006).
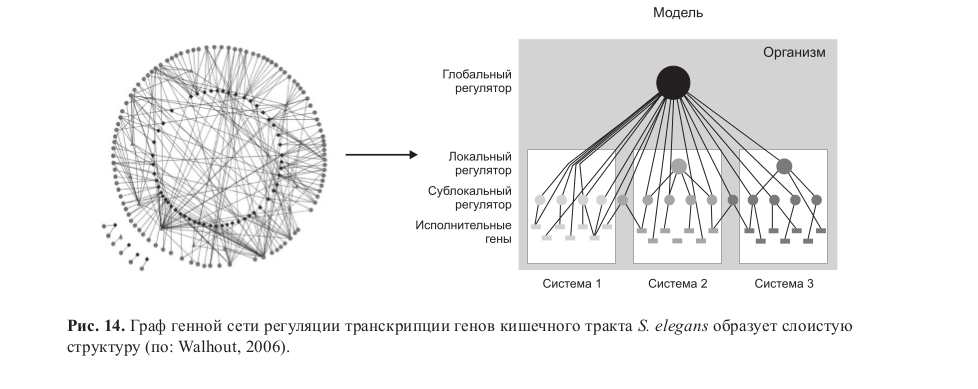
Наиболее быстрый и экономный способ оптимизировать такой параметр – вновь образовать ООС, но уже по другому регуляторному звену (рис. 13а). Граф генной сети, испытавшей несколько циклов таких «эволюционных качелей» (смена стабилизирующего отбора и движущего отбора), должен иметь многослойную структуру.
Такая структура действительно выявляется по микрочиповым данным для генов плаценты (Cross et al., 2003; Knox, Baker, 2008) и на графах генных сетей E. coli (Luscombe et al., 2004), дрожжей (Maslov et al., 2004) и С. еlegans (Walhout et al., 2006; Haerty et al., 2008) (рис. 14). В частности, в генных сетях E. coli и дрожжей обратными связями были обогащены «эндогенные» регулоги (Luscombe et al., 2004). Таким образом, эволюция должна идти скачками: после короткого периода разнообразия и высокой эволюционной пластичности таксонов, в течение которого формируются регуляторные модули, наступает длительный период постепенного снижения размаха эволюционной пластичности. В это время идут экспансия модулей в глобальных генных сетях представителей таксонов и интеграция генных сетей, увеличивается их обнейтраливающий потенциал. Картина постепенного снижения размаха эволюционной пластичности соответствует палеонтологическим данным (Раутиан, 1988; Рожнов, 2006).
Одновременно начинается процесс накопления обнейтраленных мутаций и размывания гомологии на нижних уровнях иерархии. Этот процесс подготавливает будущий выход из стазиса. Резкая смена экологической обстановки, изменяющая вектор отбора со стабилизирующего на движущий, приводит к травматичному выходу из стазиса, сопровождающемуся «эволюционным взрывом» – быстрой эволюцией новых таксонов, большинство из которых вскоре вымирает. Стресс при дестабилизирующем отборе (Беляев, 1987, 1979; Маркель, 2008) позволяет осуществить щадящий выход, кратковременно увеличивающий норму реакции существующего таксона и в лучшем случае сопровождаемый отпочковыванием от него нескольких таксонов низкого ранга.
Усиливая нагрузку на эффекторное звено, стресс позволяет частично выявлять обнейтраленную изменчивость без слома ООС: выводит часть обнейтраленных мутаций под отбор или проявляет потенциал модификационной изменчивости, ранее обнейтраливаемый ООС. Такой потенциал может включать в себя различные молекулярно-биологические механизмы, например, повышение вероятности альтернативного сплайсинга, считывание генов и белков с разных стартов транскрипции и трансляции (Uversky, Dunker, 2008), синтез особых шаперонов (Вельков, 2002).
Стохастический проскок терминирующих кодонов (фенотипическая супрессия (Тиходеев, 1990)) позволяет, например, кратковременно использовать псевдогены[25] (Балакирев, Айяла, 2004). В итоге увеличивается размах «реализационной изменчивости» (Инге-Вечтомов, 2005). В протеоме увеличивается фракция белков с латентными третичными структурами, что позволяет неспецифически увеличить амплитуду варьирования начальных параметров генной сети, обеспечив расширение нормы реакции при стрессе.
Литература
Алешин В.В., Владыченская Н.С., Кедрова О.С. и др. Вторичная структура шпильки 17 низшего многоклеточного животного Rhopalura ophiocomae (Mesozoa: Orthonectida) как пример «прерываемого равновесия» в эволюции 18S рибосомной РНК // Молекуляр. биология. 1999. Т. 33. С. 319–329.
Афонников Д.А., Колчанов Н.А. Консервативные особенности ДНК-связывающих доменов класса «гомеодомен», обусловленные коадаптивными заменами аминокислотных остатков // Докл. АН. 2001. Т. 380. No 5. С. 691–695.
Афонников Д.А., Колчанов Н.А. Координированные замены аминокислотных остатков в белках // Системная компьютерная биология. Новосибирск: Изд-во СО РАН, 2008. C. 591–602.
Балакирев Е.С., Айяла Ф.Дж. Псевдогены: консервация структуры, экспрессия и функции // Журн. общ. биологии. 2004. T. 65. No 4. С. 306–321.
Беклемишев Н.В. О классификации биоценологических (симфизиологических) связей // Биоценологические основы паразитологии. М.: Наука, 1970. С. 90–138.
Беляев Д.К. Некоторые генетико-эволюционные аспекты стресса и стрессируемости // Вестник АМН СССР. 1979. No 7. С. 9–14.
Беляев Д.К. Дестабилизирующий отбор // Развитие эволюционной теории в СССР (1917–1970 годы). Л.: Наука, 1987. С. 266–277.
Беляев Д.К., Трут Л.Н. Реорганизация сезонного ритма размножения у серебристо-черных лисиц (Vulpes vulpes Desm.) в процессе отбора на способность к доместикации // Журн. общ. Биологии. 1983. Т. 42. No 6. С. 739–752.
Берг Л.С. Номогенез, или Эволюция на основе закономерностей. Пг.: Гос. изд-во, 1922. 316 с.
Берг Л.С. Труды по теории эволюции. 1922–1930. Л.: Наука, 1977. 388 с.
Берг Р.Л. Экологическая интерпретация корреляционных плеяд // Генетика и эволюция. Избр.труды. Новосибирск: Наука, 1993а. C. 123–137.
Берг Р.Л. Корреляционные плеяды и стабилизирующий отбор // Генетика и эволюция. Избр. Труды. Новосибирск: Наука, 1993б. C. 137–178.
Бердников В.А. Основные факторы макроэволюции. Новосибирск: Наука, 1990. 253 c.
Боголюбский С.Н. Происхождение и преобразование домашних животных. М.: Сов. Наука, 1959. 593 с.
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. Саратов: Губполиграфотдел, 1920. 16 с.
Вавилов Н.И. Линнеевский вид как система // Тр. по прикл. ботан., генет. и селекции. 1931. Т. 26. Вып. 3. С. 109–134.
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. 2-е перераб. и расшир. изд-е. М.; Л.: Сельхозгиз, 1935. 56 с.
Вавилов Н.И. Мировые центры сортовых богатств (генов) культурных растений // Н.И. Вавилов. Избр.тр.: в 5 томах. Т. 5. М.: Наука, 1965. С. 108–119.
Вавилов Н.И. Великие земледельческие культуры доколумбовой Америки и их взаимоотношения // Избр. произведения: В 2 т. Л.: Наука, 1967а. Т. 1. С. 276–302.
Вавилов Н.И. Научные основы селекции пшеницы // Избр. произведения: В 2 т. Л.: Наука, 1967б. Т. 2. С. 7–259.
Вельков В.В. Новые представления о молекулярных механизмах эволюции: стресс повышает генетическое разнообразие // Молекуляр. Биология. 2002. T. 36. No 2. C. 277–285.
Воронцов Н.Н. Эволюция пищеварительной системы грызунов (Мышеобразные). Новосибирск: Наука, 1967. 239 с.
Воронцов Н.Н. Эволюция дарвинизма (Развитие эволюционных идей в биологии). М.: УНЦ ДО, 1999. 630 с.
Гайсинович А. Грегор Мендель и его предшественники // Избр. работы о растительных гибридах. М.; Л.: Гос. изд-во биол. и мед. Лит-ры, 1935. С. 11–156.
Галич М. История доколумбовых цивилизаций. М: Мысль, 1990 г. 408 с.
Гаузе Г.Ф. Борьба за существование. М.: Ижевск: Ин-т компьютерных исслед., 2002. 160 с.
Голубовский М. Парадоксы непризнания: Мендель и Мак-Клинток.
Гончаров Н.П., Глушков С.А., Шумный В.К. Доместикация злаков Старого Света: поиск новых подходов для решения старой проблемы // Журн. общ. биологии. 2007. Т. 68. No 2. С. 126–148.
Грант В. Эволюционный процесс. М.: Мир, 1991.488 c.
Гунбин К.В., Суслов В.В., Колчанов Н.А. Ароморфозы и адаптивная молекулярная эволюция // Информ. вестник ВОГиС. 2007. Т. 11. No 2. С. 373–400.
Давиташвили Л.Ш. Изменчивость организмов в геологическом прошлом. Тбилиси: Мецниереба, 1970. 252 с.
Дарвин Ч. Изменение животных и растений в домашнем состоянии. М.; Л.: ОГИЗ-Сельхозгиз, 1941. 619 с.
Дарвин Ч. Происхождение видов путем естественного отбора. СПб: Наука, 1991. 539 c.
Джуа М. История химии. М.: Мир, 1966. 452 с.
Захаров-Гезехус И.А. Проблема гомологии в эволюционной биологии. М., 2008. 127 с.
Иванисенко В.А., Деменков П.С., Фомин Э.С. и др. Распознавание функциональных сайтов в пространственных структурах белков // Системная компьютерная биология. Новосибирск: Изд-во СО РАН, 2008. C. 231–245.
Инге-Вечтомов С.Г. Роль генетических процессов в модификационной изменчивости. Пророчество Б.Л. Астаурова // Онтогенез. 2005. Т. 36. No 4. С. 274–279.
Камшилов М.М. Отбор в различных условиях проявления признака // Биол. журнал. 1935. Т. 4. No 6. С. 1005–1013.
Камшилов М.М. Отбор как фактор, меняющий зависимость признака от изменений внешних условий // Докл. АН СССР. 1939. T. 23. No 4. С. 361–364.
Камшилов М.М. Проявление признака и изменчивость. Эффект порога проявления // Докл. АН. 1940а. Т. 26. No 6. С. 605–608.
Камшилов М.М. Изменчивость и проявление. Проблема нормального фенотипа // Докл. АН. 1940б. Т. 29. No 3. С. 239–243.
Камшилов М.М. Эволюция биосферы. 2-е изд., М.: Наука, 1974. 256 с.
Кашев И.И. Очерки из истории сравнительной анатомии до Дарвина. М.; Л.: Изд-во АН СССР, 1963. С. 1–298.
Кикнадзе И.И. Учитель жив в своих учениках // Дмитрий Константинович Беляев: Книга воспоминаний. Новосибирск: Изд-во СО РАН. Филиал «Гео», 2002. C. 97–105.
Колмогоров А.Н. Уклонение от формул Харди при частичной изоляции // Докл. АН СССР. 1935. Т. 3. No 7. С. 129–132.
Колчанов Н.А., Шиндялов И.Н. Теоретическое иследование эволюции регуляторных контуровпри различных типах отбора // Проблемы генетики и теории эволюции. Новосибирск: Наука, Вестник ВОГиС, 2009, Том 13, No 2 1991. С. 268–279.
Колчанов Н.А., Ананько Е.А., Колпаков Ф.А. и др. Генные сети // Молекуляр. биология. 2000. Т. 34. No 4. С. 533–544.
Колчанов Н.А., Подколодная О.А., Игнатьева Е.В. и др. Интеграция генных сетей, контролирующих физиологические функции организма // Информ. вестник ВОГиС. 2005. Т. 9. No 2. С. 179–198.
Коржинский С.И. Гетерогенезис и эволюция. К теории происхождения видов. I // Зап. Акад. наук по физ.-мат. отделению. 1899. Т. 9. No 2. С. 1–94.
Кренке Н.П. Соматические показатели и факторы формообразования // Феногенетическая изменчивость. Т. 1. Сборник работ отделения фитоморфогенеза // Тр. Биол. ин-та им. К.А. Тимирязева. М.: Изд-во ин-та им. К.А. Тимирязева, 1933–1935. C. 11–415.
Курочкин Е.Н. Параллельная эволюция тероподных динозавров и птиц // Зоол. журнал. 2006. Т. 85. No 3. С. 283–297.
Лакатос И. Доказательства и опровержения. М.: Наука, 1967. 152 с.
Ламарк Ж.-Б. Философия зоологии. М.: Наука, 1911. 313 с.
Левонтин Р. Генетические основы эволюции. М.: Мир, 1978. 349 с.
Лихошвай В.А., Колчанов Н.А. Модели гипотетических генных сетей // Системная компьютерная биология. Новосибирск: Изд-во СО РАН, 2008. С. 399–408.
Льоцци М. История физики. М.: Мир, 1970. 464 с.
Любищев А.А. Закон гомологических рядов Н.И. Вавилова и его значение в биологии. Рукопись, 1957.
Любищев А.А. Проблемы формы, систематики и эволюции организмов. М.: Наука, 1982. 280 c.
Маркель А.Л. Стресс и эволюция // Информ. вестник ВОГиС. 2008. Т. 12. No 1/2. С. 206–215.
Маркель А.Л., Бородин П.М. Онтогенетические и генетико-эволюционные аспекты нейроэндокринной регуляции стресса // Стресс как фактор регуляции генетической изменчивости. Новосибирск: Наука, 1990. С. 148–159.
Мейен С.В. Проблема направленности эволюции // Итоги науки и техники. Зоология позвоночных. Т. 7. Проблемы теории эволюции. М.: Наука, 1975. С. 66–117.
Мейен С.В. Таксономия и мерономия // Вопросы методологии в геологических науках. Киев: Наук. думка, 1977. С. 25–33.
Мейен С.В. Основные аспекты типологии организмов // Журн. общ. биологии. 1978. Т. 39. No 4. С. 495–508.
Мендель Г. Опыты над растительными гибридами // Избранные работы о растительных гибридах. Вестник ВОГиС, 2009, Том 13, No 2. М.; Л.: Гос. изд-во биол. и мед. Лит-ры, 1935. С. 233–291.
Михайлова И.А., Бондаренко О.Б. Палеонтология. Т. 1. М.: Изд-во МГУ, 1997. 446 c.
Науменко Е.В. Адренокортикальная система у разных видов доместицированных животных // Журн. общ. биологии. 1988. Т. 47. No 4. С. 460–465.
Науменко Е.В., Осадчук А.В., Серова Л.И., Шишкина Г.Т. Генетико-физиологические механизмы регуляции функций семенников. Новосибирск: Наука, 1983. 203 с.
Нодэн Ш. Новые исследования над гибридностью растений // Избр. работы о растительных гибридах. М.; Л.: Гос. изд-во биол. и мед. Лит-ры, 1935. С. 173–232.
Обручев Д.В. Некоторые критерии филогенетических исследований на примере низших позвоночных // Палеонт. журнал. 1972. No3. С. 56–71.
Пономаренко А.Г. Артроподизация и ее экологические последствия // Экосистемные перестройки и эволюция биосферы. М.: ПИН РАН, 2004. Вып. 6. С. 7–22.
Попова Н.К. Что определяет поведение? // Дмитрий Константинович Беляев: Книга воспоминаний. Новосибирск: Изд-во СО РАН. Филиал «Гео», 2002. С. 109–111.
Ратнер В.А. Генетика. Молекулярная кибернетика. Личности и проблемы. Новосибирск: Наука, 2002. C. 104–121.
Ратнер В.А., Жарких А.А., Колчанов Н.А. и др. Проблемы теории молекулярной эволюции. Новосибирск: Наука, 1985. 260 с.
Раутиан А.С. Палеонтология как источник сведений о закономерностях и факторах эволюции // Современная палеонтология. Методы, направления, проблемы, практическое приложение. Т. 2. М.: Недра, 1988. С. 76–118.
Рожнов С.В. Закон гомологических рядов Н.И. Вавилова и архаическое многообразие по данным палеонтологии // Эволюция биосферы и биоразнообразия (к 70-летию А.Ю. Розанова). М.: Т-во научных изданий КМК, 2006. C. 134–147.
Симпсон Дж.Г. Темпы и формы эволюции. М.: Изд-во иностр. лит-ры, 1948. 369 c.
Скворцов А.К. У истоков систематики. К 300-летию Карла Линнея // Природа. 2007. No 4. С. 3–10.
Соболев Д.Н. Начало исторической биогенетики. Гос. изд-во Украины, 1924, 202 с.
Суслов В.В., Колчанов Н.А., Сергеев М.Г. Молекулярно-генетические механизмы процессов формирования биоразнообразия // Биоразнообразие и динамика экосистем: информационные технологии и моделирование. Новосибирск: Изд-во СО РАН, 2006. C. 317–344.
Татаринов Л.П. Морфологическая эволюция териодонтов и общие вопросы филогенетики. М.: Наука, 1976. 257 с.
Тиходеев О.Н., Гетманова Е.В., Тихомирова В.Л., Инге-Вечтомов С.Г. Неоднозначность трансляции у дрожжей: генетический контроль и модификации // Молекулярные механизмы генетических процессов. М.: Наука, 1990. С. 218–228.
Трапезов О.В. Гомологические ряды изменчивости окраски меха у американской норки (Mustela vison Schreber, 1777) в условиях доместикации // Информ. вестник ВОГиС. 2008. Т. 11. No 3/4. С. 547–559.
Трут Л.Н. Эволюционные идеи Д.К. Беляева как концептуальный мост между биологией, социологией и медициной // Информ. вестник ВОГиС. 2008. Т. 12. No 1/2. С. 7–18.
Фандо Р.А., Музрукова Е.Б. Взаимопроникновение медицинских и биологических воззрений в проблему наследственности человека: историко-научный анализ // Информ. вестник ВОГиС. 2008. Т. 12. No 3. С. 474–482.
Холодковский Н.А. Биологические очерки. М.; Пг.: Гос. изд-во, 1923. 356 с.
Филипченко Ю.А. Эволюционная идея в биологии. М.: Наука, 1977. 227 с.
Фламель Н. Алхимия. СПб: Азбука. Петербургское Востоковедение, 2001. 384 с.
Шаталкин А.И. Регуляторные гены в развитии и проблема морфотипа в систематике насекомых // Чтения памяти Н.А. Холодковского. СПб, 2003. Вып. 56(2). 109 с.
Шестаков С.В. О ранних этапах биологической эволюции с позиции геномики // Палеонтол. Журнал. 2003. No 6. С. 50–57.
Шестаков С.В. Как происходит и чем лимитируется горизонтальный перенос генов у бактерий // Экол. генетика. 2007. Т. 5. Вып. 2. С. 12–24.
Шмальгаузен И.И. Факторы эволюции (теория стабилизирующего отбора). М.: Наука, 1968. 452 с.
Штрубе В. Пути развития химии. М.: Мир, 1984. Т. 1. 239 с.
Шульгина Н.К. Эндокринологические аспекты промышленной доместикации соболей (Martes zibellina Linnaeus, 1758) // Информ. вестник ВОГиС. 2007. Т. 11. No 1. С. 76–90.
Шумилина Н.Н. Доместикационные преобразования конституциональных особенностей серебристочерных лисиц (Vulpes vulpes Desm.) в ходе их промышленного разведения // Информ. вестник ВОГиС. 2007. Т. 11. No 1. С. 109–114.
Ютен С. Повседневная жизнь алхимиков в средние века. М.: Молодая Гвардия. Палимпсест, 2005. 256 с.
al-Hassan A.Y., Hill D.R. Islamiс technology. Cambridge: 436 Cambridge Univ. Press, 1987. 320 c.
Arbiza L., Dopazo J., Dopazo H. Positive selection, relaxation, and acceleration in the evolution of the human and chimp genome // PLoS Comput. Biol. 2006. V. 2. e38.
Asher R.J. A web-database of mammalian morphology and a reanalysis of placental phylogeny // BMC Evol. Biol. 2007. V. 7. P. 108.
Babu M.M., Luscombe N.M., Aravind L. et al. Structure and evolution of transcriptional regulatory networks // Curr. Opin. Struct. Biol. 2004. V. 14. No 3. P. 283–291.
Badiu C. Genetic clock of biologic rhythms // J. Cell. Mol. Med. 2003. V. 7. No 4. P. 408–416.
Bergmann S., Ihmels J., Barkai N. Similarities and differences in genome-wide expression data of six organisms // PLoS Biol. 2004. V. 2. E9.
Cain A.J. Thomas Sydenham, John Ray, and some contemporaries on species // Archives of Natural History. 1999. V. 26. No 1. P. 55–83.
Carter A.M., Enders A.C. Comparative aspects of trophoblast development and placentation // Reprod. Biol. Endocrinol. 2004. V. 2. P. 46.
Chalmel F., Rolland A.D., Niederhauser-Wiederkehr C. et al. The conserved transcriptome in human and rodent male gametogenesis // Proc. Natl Acad. Sci. USA. 2007. V. 104. No 20. P. 8346–8351.
Chervitz S.A., Aravind L., Sherlock G. et al. Comparison of the complete protein sets of worm and yeast: orthology and divergence // Science. 1998. V. 282. No 5396. P. 2022–2028.
Cook C.E., Yue Q., Akam M. Mitochondrial genomes suggest that hexapods and crustaceans are mutually paraphyletic // Proc. Biol. Sci. 2005. V. 272. No 1569. P. 1295–1304.
Cooper B., Clarke J.D., Budworth P. et al. A network of rice genes associated with stress response and seed development // Proc. Natl Acad. Sci. USA. 2003. V. 100. No 8. P. 4945–4950.
Cross J.C., Baczyk D., Dobric N. et al. Genes, development and evolution of the placenta // Placenta. 2003. V. 24. No 2/3. P. 123–130.
CSAC – Chimpanzee Sequencing and Analysis Consortium. Initial sequence of the chimpanzee genome and comparison with the human genome // Nature. 2005. V. 437. No 7055. P. 69–87.
Darwin Сh. Effects of Cross and Self Fertilization in the Vegetable Kingdom. London, 1877.
De S., Teichmann S.A., Babu M.M. The impact of genomic neighborhood on the evolution of human and chimpanzee transcriptome // Genome Res. 2009. V. 19. No 5. P. 785–794.
Dobretsov N.L., Kolchanov N.A., Suslov V.V. On important stages of Geosphere and Biosphere evolution// Biosphere origin and evolution. Springer Science +Business Media, LLC, 2008. P. 3–24.
Eddy E.M. Male germ cell gene expression // Recent. Progr. Horm. Res. 2002. V. 57. P. 103–128.
Edwards S.V., Bryan Jennings W., Shedlock A.M. Phylogenetics of modern birds in the era of genomics // Proc. Biol. Sci. 2005. V. 272. No 1567. P. 979–992.
Eldrege N., Gould S.J. Punctuated equilibria: an alternative to phyletic gradualism // Models in Paleobiology. San Francisco: Freeman Cooper and Co, 1972. P. 82–115.
Evans P.D., Anderson J.R., Vallender E.J. et al. Adaptive evolution of ASPM, a major determinant of cerebral cortical size in humans // Hum. Mol. Genet. 2004. V. 13. No 5. P. 489–494.
Fayngerts S.A., Najakshin A.M., Taranin A.V. Speciesspecific evolution of the FcR family in endothermic vertebrates // Immunogenetics. 2007. V. 59. No 6. P. 493–506.
Feduccia A., Lingham-Soliar T., Hinchliffe J.R. Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence // J. Morphol. 2005. V. 266. No 2. P. 125–166.
Flemming A.F., Blackburn D.G. Evolution of placental specializations in viviparous African and South American lizards // J. Exptl. Zoolog. A. Comp. Exptl. Biol. 2003. V. 299. No 1. Р. 33–47.
Freyer C., Zeller U., Renfree M.B. The marsupial placenta: a phylogenetic analysis // J. Exptl. Zoolog. A. Comp. Exptl. Biol. 2003. V. 299. No 1. Р. 59–77.
Gasch A.P., Moses A.M., Chiang D.Y. et al. Conservation and evolution of cis-regulatory systems in ascomycete fungi // PLoS Biol. 2004. V. 2. No 12. e398.
Gehring W.J. New perspectives on eye development and the evolution of eyes and photoreceptors // J. Hered. 2005. V. 96. No 3. P. 171–184.
Gilbert S.F., Opitz J.M., Raff R.A. Resynthesizing evolutionary and developmental biology // Dev. Biol. 1996. V. 173. No 2. P. 357–372.
Glazko V.I. An attempt at understanding the genetic basis of domestication // Anim. Sci. Papers and Rep. 2003. V. 21. No 2. P. 109–120.
Glazko G., Veeramachaneni V., Nei M., Makałowski W. Eighty percent of proteins are different between humans and chimpanzees // Gene. 2005. V. 346. P. 215–219.
Guselnikov S.V., Ramanayake T., Erilova A.Y. et al. The Xenopus FcR family demonstrates continually high diversification of paired receptors in vertebrate evolution // BMC Evol. Biol. 2008. V. 8. P. 148.
Haerty W., Artieri C., Khezri N. et al. Comparative analysis of function and interaction of transcription factors in nematodes: extensive conservation of orthology coupled to rapid sequence evolution // BMC Genomics. 2008. V. 9. No 399.
Hiden U., Bilban M., Knöfler M., Desoye G. Kisspeptins and the placenta: regulation of trophoblast invasion // Rev. Endocr. Metab. Disord. 2007. V. 8. No 1. P. 31–39.
Honma T., Goto K. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs // Nature. 2001. V. 409. No 6819. P. 525–529.
Hutchinson G.E. The niche: an abstractly inhabited hypervolume // The ecological theatre and the evolutionary play. New Haven: Yale Univ. Press, 1965. P. 26–78.
Huynen M.A., Snel B., van Noort V. Comparative genomics for reliable protein-function prediction from genomic data // Trends Genet. 2004. V. 20. No 8. P. 340–344.
Jacobs J.F., Brasseur F., Hulsbergen-van de Kaa C.A. et al. Cancer-germline gene expression in pediatric solid tumors using quantitative real-time PCR // Int. J. Cancer. 2007. V. 120. No 1. P. 67–74.
Jacobs J.F., Grauer O.M., Brasseur F. et al. Selective cancer-germline gene expression in pediatric brain tumors // J. Neurooncol. 2008. V. 88. No 3. P. 273–280.
Jeffery J.E., Bininda-Emonds O.R.P., Coates M.I., Richardson M.K. Analysing evolutionary patterns in amniote embryonic development // Evol. Dev. 2002. V. 4. No 4. P. 292–302.
Katoh Y., Katoh M. Comparative genomics on Sonic hedgehog orthologs // Oncol. Rep. 2005. V. 14. No 4. P. 1087–1090.
Knerr I., Huppertz B., Weigel C. et al. Endogenous retroviral syncytin: compilation of experimental research on syncytin and its possible role in normal and disturbed human placentogenesis // Mol. Hum. Reprod. 2004. V. 10. No 8. Р. 581–588.
Knox K., Baker J.C. Genomic evolution of the placenta using co-option and duplication and divergence // Genome Res. 2008. V. 18. No 5. P. 695–705.
Kumar S., Balczarek K.A., Lai Z.C. Evolution of the hedgehog gene family // Genetics. 1996. V. 142. No 3. P. 965–972.
Lakatos I. Falsification and the Methodology of Scientific Research Programmes // Lakatos I. Philosophical Papers. Cambridge U.P.: Cambridge, 1978. V. 1. P. 8–101.
Lespinet O., Wolf Y.I., Koonin E.V., Aravind L. The role of lineage-specific gene family expansion in the evolution of eukaryotes // Genome Res. 2002. V. 12. No 7. P. 1048–1059.
Lindsay E.A., Botta A., Jurecic V. et al. Congenital heart disease in mice deficient for the DiGeorge syndrome region // Nature. 1999. V. 401. No 6751. P. 379–383.
Luscombe N.M., Babu M.M., Yu H. et al. Genomic analysis of regulatory network dynamics reveals large topological changes // Nature. 2004. V. 431. No 7006. P. 308–312.
Maslov S., Sneppen K., Eriksen K.A., Yan K.K. Upstream plasticity and downstream robustness in evolution of molecular networks // BMC Evol. Biol. 2004. V. 4. No 9.
McCarrey J.R., Riggs A.D. Determinator-inhibitor pairs as a mechanism for threshold setting in development: a possible function for pseudogenes // Proc. Nat. Acad. Sci. USA. 1986. V. 83. No 3. P. 679–683.
Meyen S.V. Nomothetical plant morphology and the nomothetical theory of evolution: the need for cross-pollination // Acta Biotheoretica. 1978. V. 27. Suppl.: Folia biotheoretica. No 7. P. 21–36.
Miyamoto M.M., Fitch W.M. Testing the covarion hypothesis of molecular evolution // Mol. Biol. Evol. 1995. V. 12. No 3. P. 503–513.
Nybakken K., Perrimon N. Hedgehog signal transduction: recent findings // Curr. Opin. Genet. Dev. 2002. V. 12. No 5. P. 503–511.
Ochman H., Lawrence J.G., Groisman E.A. Lateral gene transfer and the nature of bacterial innovation // Nature. 2000. V. 405. No 6784. P. 299–304.
Old L.J. Cancer is a somatic cell pregnancy // Cancer Immunity. 2007. V. 7. P. 19–21.
Palatnik J.F., Allen E., Wu X. et al. Control of leaf morphogenesis by microRNAs // Nature. 2003. V. 425. No 6955. P. 257–263.
Paterson A., Lin Y., Li Z. et al. Convergent domestication of cereal crops by independent mutations at corresponding genetic loci // Science. 1995. V. 269. No 5231. P. 1714–1718.
Paterson A.H., Bowers J.E., Burow M.D. et al. Comparative genomics of plant chromosomes // Plant. Cell. 2000. V. 12. No 9. P. 1523–1540.
Paterson A.H., Bowers J.E., Chapman B.A. Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics // Proc. Natl Acad. Sci. USA. 2004. V. 101. No 26. Р. 9903–9908.
Pauli F., Liu Y., Kim Y.A. et al. Chromosomal clustering and GATA transcriptional regulation of intestine-expressed genes in C. elegans // Development. 2006. V. 133. No 2. P. 287–295.
Peregrin-Alvarez J.M., Tsoka S., Ouzounis C.A. The phylogenetic extent of metabolic enzymes and pathways // Genome Res. 2003. V. 13. No 3. P. 422–427.
Piskur J. Origin of duplicated regions in the yeast genomes // Trends Genet. 2001. V. 17. No 6. P. 302–303.
Ponomarenko J.V., Furman D.P., Frolov A.S. et al. ACTIVITY: a database on DNA/RNA sites activity adapted to apply sequence-activity relationships from one system to another // Nucl. Acids Res. 2001. V. 29. No 1. P. 284–287.
Ponomarenko J.V., Ponomarenko M.P., Frolov A.S. et al. Conformational and physicochemical DNA features specific for transcription factor binding sites // Bioinformatics. 1999. V. 15. No 7/8. P. 654–668.
Raff R.A., Sly B.J. Modularity and dissociation in the evolution of gene expression territories in development // Evol. Dev. 2000. V. 2. No 2. P. 102–111.
Ravasz E., Barabási A.L. Hierarchical organization in complex networks // Phys. Rev. E. Stat. Nonlin. Soft. Matter. Phys. 2003. V. 67. No 2. Pt. 2. 026112.
Regier J.C., Shultz J.W., Kambic R.E. Pancrustacean phylogeny: hexapods are terrestrial crustaceans and maxillopods are not monophyletic // Proc. Biol. Sci. 2005. V. 272. No 1561. P. 395–401.
Reichert H., Simeone A. Developmental genetic evidence for a monophyletic origin of the bilaterian brain // Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2001. V. 356. No 1414. P. 1533–1544.
Richardson M.K., Hanken J., Gooneratne M.L. et al. There is no highly conserved embryonic stage in the vertebrates: implications for current theories of evolution and development // Anat. Embryol. (Berl). 1997. V. 196. No 2. Р. 91–106.
Richardson M.K., Keuck G. Haeckel’s ABC of evolution and development // Biol. Rev. Camb. Philos. Soc. 2002. V. 77. No 4. P. 495–528.
Rooney A.P., Zhang J., Nei M. An unusual form of purifying selection in a sperm protein // Mol. Biol. Evol. 2000. V. 17. No 2. P. 278–283.
Rubel E. The Replaceability of Ecological Factors and the Law of the Minimum // Ecology. 1935. V. 16. No 3. P. 336–341.
Schlecht U., Primig M. Mining meiosis and gametogenesis with DNA microarrays // Reproduction. 2003. V. 125. No 4. P. 447–456.
Sharan R., Suthram S., Kelley R.M. et al. Conserved patterns of protein interaction in multiple species // Proc. Natl Acad. Sci. USA. 2005. V. 102. No 6. P. 1974–1979.
Sharma M.K., Liu R.Z., Thisse C. et al. Hierarchical subfunctionalization of fabp1a, fabp1b and fabp10 tissue-specific expression may account for retention of these duplicated genes in the zebrafish (Danio rerio) genome // FEBS J. 2006. V. 273. No 14. P. 3216–3229.
Shi P., Bakewell M.A., Zhang J. Did brain-specific genes evolve faster in humans than in chimpanzees? // Trends Genet. 2006. V. 22. No 11. P. 608–613.
Snel B., van Noort V., Huynen M.A. Gene co-regulation is highly conserved in the evolution of eukaryotes and prokaryotes // Nucl. Acids Res. 2004. V. 32. No 16. P. 4725–4731.
Souza F.S. de, Bumaschny V.F., Low M.J. et al. Subfunctionalization of expression and peptide domains following the ancient duplication of the proopiomelanocortin gene in teleost fishes // Mol. Biol. Evol. 2005. V. 22. No 12. P. 2417–2427.
Stewart J.R., Thompson M.B. Evolutionary transformations of the fetal membranes of viviparous reptiles: a case study of two lineages // J. Exptl. Zoolog. A. Comp. Exptl. Biol. 2003. V. 299. No 1. Р. 13–32.
Stuart J.M., Segal E., Koller D., Kim S.K. A genecoexpression network for global discovery of conserved genetic modules // Science. 2003. V. 302. No 5643. P. 249–255.
Swanson W.J., Vacquier V.D. The rapid evolution of reproductive proteins // Nat. Rev. Genet. 2002. V. 3. No 2. P. 137–144.
Taddei I., Morishima M., Huynh T., Lindsay E.A. Genetic factors are major determinants of phenotypic variability in a mouse model of the DiGeorge/ del22q11 syndromes // Proc. Natl Acad. Sci. USA. 2001. V. 98. No 20. P. 11428–11431.
Tamames J. Evolution of gene order conservation in prokaryotes // Genome Biol. 2001. V. 2. No 6. Research0020.
Tanay A., Regev A., Shamir R. Conservation and evolvability in regulatory networks: the evolution of ribosomal regulation in yeast // Proc. Natl Acad. Sci. USA. 2005. V. 102. No. 20. P. 7203–7208.
Teichmann S.A., Babu M.M. Conservation of gene coregulation in prokaryotes and eukaryotes // Trends Biotechnol. 2002. V. 20. No 10. Р. 407–410.
Teichmann S.A., Babu M.M. Gene regulatory network growth by duplication // Nat. Genet. 2004. V. 36. No 5. P.492–496.
Theissen G., Saedler H. Floral quartets // Nature. 2001. V. 409. No 6819. P. 469–471.
Timofeeff-Ressovsky N.W. Studies on the phenotype manifestation of hereditary factors. I. On the phenotypic manifestation of the genovariation radius incompletus in Drosophila funebris // Genetics. 1927. V. 12. P. 128–198.
Uversky V.N., Dunker A.K. Biochemistry. Controlled chaos // Science. 2008. V. 322. No 5906. P. 1340–1341.
Vanselow K., Vanselow J.T., Westermark P.O. et al. Differential effects of PER2 phosphorylation: molecular basis for the human familial advanced sleep phase syndrome (FASPS) // Genes Dev. 2006. V. 20. No 19. P. 2660–2672.
Vasilyeva L.N. Systematics in mycology // Bibl. Mycol. 1999. Bd. 178. S. 1–253. Walbot V. A green chapter in the book of life // Nature. 2000. V. 408. No 6814. P. 794–795.
Walhout A.J. Unraveling transcription regulatory networks by protein-DNA and protein-protein interaction mapping // Genome Res. 2006. V. 16. No 12. P. 1445–1454.
Walsh B.D. On certain entomological speculation of the New England school of naturalists // Proc. Entomol. Soc. Phil. 1864. V. 3. P. 207–249.
Wang Y., Gu X. Evolutionary patterns of gene families generated in the early stage of vertebrates // J. Mol. Evol. 2000. V. 51. No 1. P. 88–96.
Wildman D.E., Uddin M., Liu G. et al. Implications of natural selection in shaping 99.4 % nonsynonymous DNA identity between humans and chimpanzees: enlarging genus Homo // Proc. Natl Acad. Sci. USA. USA. 2003 V. 100. No 12. Р. 7181–7188.
Wilson A.C., Sarich V.M., Maxson L.P. The importance of gene rearrangement in evolution – evidence from studies on rates of chromosomal, protein, and anatomical evolution // PNAS. USA. 1974. V. 71. No 8. P. 3028–3030.
Wittkopp P.J. Variable gene expression in eukaryotes: a network perspective // J. Exptl. Biol. 2007. V. 210. Pt. 9. P. 1567–1575.
Wyckoff G.J., Wang W., Wu C.I. Rapid evolution of male reproductive genes in the descent of man // Nature. 2000. V. 403. No 6767. P. 261–263.
Xia K., Xue H., Dong D. et al. Identification of the proliferation/differentiation switch in the cellular network of multicellular organisms // PLoS Comput. Biol. 2006. V. 2. No 11. P. 145.
Xu Y., Padiath Q.S., Shapiro R.E. et al. Functional consequences of a CKIdelta mutation causing familial advanced sleep phase syndrome // Nature. 2005. V. 434. No 7033. P. 640–644.
Вестник ВОГиС. 2009. Т.13. №2.
Примечания
[1]Сильно упрощая, можно сказать, что различия между «землями» (рудами) и составляющими их элементами алхимики объясняли различной пропорцией в них металлического и неметаллического начал – философской ртути и философской серы. Пропорция постепенно медленно меняется под воздействием элементов друг на друга в «лоне земли». Трансмутация элементов – ускоренный вариант такой эволюции, воспроизведенный в реторте (Фламель, 2001; Штрубе, 1984). Таким образом, эволюция мыслится как постепенное нарушение равновесия. Соответственно, возврат к равновесию обеспечивает круговорот начал и стабильность аристотелевской лестницы природы. Следуя аристотелевой традиции, А. фон Больштедт считал «земли» и минералы царством природы наряду с растениями и животными, соответственно допуская трансмутацию и для последних. Создание такой всеобщей алхимической «теории эволюции» растянулось на 200 лет и было закончено Корнелием Агриппой (1486–1535) (Ютен, 2005).
[2]Одним из важнейших концептуальных достижений физики Галилея–Ньютона было представление об идеальных объектах, позволявшее широко применять математический аппарат (Льоцци, 1970). Заимствовав из физики представление об идеальных объектах (например типы строения Кювье (Кашев, 1963)), биология невольно позаимствовала и ньютоново представление о силе как мере взаимодействия объектов. Эволюция мыслилась как аналог движения с ускорением, и камнем преткновения для эволюционистов был поиск силы, обеспечивающей это ускорение.
Э. Жоффруа Сент-Илер видел источник эволюции в прямом воздействии среды. У Ламарка наоборот организм отталкивается от среды, побуждаемый стремлением к совершенству тренировать органы (Холодковский, 1923). Теории неизменности видов никаких сил не требовали, поэтому оказались в более выигрышном положении.
[3]Впервые о виде как о реально существующем биологическом феномене заговорил Дж. Рей (1627–1705), прибавивший к аристотелевскому требованию морфологического сходства (species, от лат. specere – разглядывать, т. е. вид – то общее, что можно выделить взглядом) требование устойчивого наследования этого сходства при эндогамии. До Рэя вид оставался абстрактной категорией классификации как живых существ, так и косных тел («земли», минералы, химические вещества) (Cain, 1999).
[4]«Все, что природа заставила особей приобрести или утратить под влиянием внешних обстоятельств, в которых с давних пор пребывала их порода, и, следовательно, под влиянием преобладающего употребления известного органа … все это она сохраняет – путем размножения – в новых особях, происходящих от прежних, если только приобретенные изменения общи обоим полам»
(Ламарк, 1911) – упоминание о подборе пар выделено нами курсивом.
[5] «Смешение же двух особей, имеющих различные качества и форму, ставит неизбежное препятствие непрерывной передаче и этих качеств, и этих форм. Вот почему у человека, которому приходится испытывать на себе влияние столь многих обстоятельств, качества и случайно приобретенные им недостатки не сохраняются и не передаются из поколения в поколение» (Ламарк, 1911).
[6] Таким образом, неоламаркисты вернулись фактически не к Ламарку, а к Жоффруа Сент-Илеру (Филипченко, 1977).
[7]Игнорирование индивидуальной изменчивости, в частности, было одной из причин популярности ламаркистских концепций именно среди палеонтологов (Оуэн, Осборн, Коп и др.).
[8]Первые систематизированные сводки по изменчивости и гибридизации домашних животных (прежде всего лошадей) составлялись уже в средневековых арабских «домах мудрости» (al-Hassan A.Y., Hill, 1987) – аналогах европейских университетов (Боголюбский, 1959). Особое место занимала селекция в доколумбовых цивилизациях Америки: отсутствие домашних животных в Мексике заставляло вести огородное хозяйство с индивидуальным подходом к растению (Вавилов, 1967а), а централизованное распределение заставило инков ввести аналоги государственных сортоиспытательных станций (Галич, 1990). Опыты с гибридами ставил К. Линней, получивший менделевские соотношения, но не сумевший увидеть в них закономерность (Скворцов, 2007). И.Г. Кельрейтер, напротив, увидел в поглощающих скрещиваниях общую закономерность, обеспечивающую константность вида у животных и растений, и прямую аналогию с алхимической трансмутацией элементов (Кельрейтер, 1940).
[9]И. Лакатош выделяет в развитии научного направления – «научно-исследовательской программы» – стадии формирования «твердого ядра», включающего базовый набор научных теорий, правил положительной эвристики, определяющих направления и методы поиска новых фактов и гипотез, подтверждающих теории, и правил отрицательной эвристики, выполняющих противоположную функцию. Благодаря этим правилам на следующей стадии «ядро» окружается «защитным поясом» из служебных гипотез, объясняющих все новые и новые эмпирические факты. Размер «пояса» колеблется – часть слyжебных гипотез опровергается. Научно-исследовательская программа может быть отвергнута лишь конкурирующей научной программой, пояс которой растет быстрее (Лакатос, 1964; Lakatos, 1978).
[10]В этом смысле теорию Дарвина можно считать одним из первых прорывов в неклассическую науку с ее вероятностной картиной мира.
[11]Таким образом, именно достоинство работы Менделя – выбор константных контрастных форм, позволивший провести математический анализ, – привело к тому, что работа оказалась невостребованной.
[12]Английский садоводческий термин. Хотя Дарвин (1991) не исключал роли «спортов» в эволюции, основным материалом для нее он все же считал неопределенную изменчивость.
[13] В 1920 г. XX в. аналогичная ситуация сложилась в физике вокруг квантовой механики (Льоцци, 1970).
[14]Так, выяснив, что признаки могут устойчиво наследоваться даже тогда, когда не принадлежат обоим родителям (гомозиготы в F2 по современной терминологии), Дарвин ограничился замечанием, что такие формы в потомстве не преобладают (Дарвин, 1941).
[15]Состоя в переписке с Ш. Нодэном, Дарвин в теории пангенезиса исходил из его экспериментов, предвосхитивших работу Менделя. Но, восприняв идею наследственных сущностей, обеспечивавших комбинаторику признаков (Нодэн, 1935), Дарвин предположил, что они возникают не только в половых клетках (геммулы) и подвержены влиянию среды. Ожидая обвинений в умозрительности, Дарвин пытался найти аналоги геммул в природе. Предположив, что они могут быть сходны с возбудителями оспы, он стал, таким образом, одним из предшественников теории симбиогенеза и горизонтального переноса (Дарвин, 1941).
[16]О параллельном формировании артроподного (Пономаренко, 2004), орнитойдного (Feduccia et al., 2005; Курочкин, 2006) и маммального (Татаринов, 1976) морфофизиологического комплексов свидетельствуют также данные палеонтологии.
[17] Следует отметить, что одним из первых на параллелизм изменчивости доместицированных животных и растений обратил внимание Дарвин (1941).
[18] У растений гены устойчивости к неблагоприятным факторам часто входят в генные сети онтогенеза и/или архитектоники и/или опосредуют связь между внутренней и внешней средами организма (Cooper et al., 2003; Гончаров, 2007).
[19]А.Н. Колмогоров аналитически строго показал, что наибольший темп накопления изменчивости характерен именно для совокупности популяций-полуизолятов (Колмогоров, 1935).
[20] Роль параметров демонстрирует правило родственных отклонений Н.П. Кренке (1933–1935): признак, расцениваемый как тератоморфа, для одного таксона может быть нормой в родственном таксоне. Механизмы образования тератоморф хорошо описываются динамическими моделями онтогенеза и часто связаны с параметрической неустойчивостью.
[21]Возможно, еще более глубокая перекомпоновка, объединившая восстановленные (?) эволюционно древние и молодые генные сети, cопровождала возникновение уникального эмбриогенеза млекопитающих, с его комбинацией примитивных (дробление, сходное с таковым у кишечнополостных) и продвинутых стадий (формирование амниотического эмбриона). Кроме того, возникновение трофобласта могло быть связано с адаптивным рекрутированием генных сетей онкогенеза (Eddy et al., 2002; Hiden et al., 2007; Jacobs et al., 2007; Old, 2007), связанных с регуляцией клеточного цикла и межклеточных взаимодействий у Metazoa. Еще один случай адаптивного рекрутирования таких генных сетей описан Н.Н. Воронцовым (1967): в семействе слепышевых (Spalacidae) ворсинчатый эпителий желудка, заселяемый симбиотической микрофлорой и способствующий мацерации растительных тканей, – норма у цокора (Myospalax), землекопов (Cryptomys) и африканских бамбуковых крыс (Tachyoryctes) и онкологическое заболевание у прочих грызунов.
[22]Возможен также перебор пространства возможностей на уровне субъединиц белка. Например, таким образом в разных таксонах независимо сформировались FcR-рецепторы иммунной системы (Fayngerts et al., 2007; Guselnikov et al., 2008).
[23]Так как протамины играют важную роль и в оплодотворении, движущий кимурианский отбор, видимо, играет роль в двух противоположных трендах эволюции: изменяя белок, формирует репродуктивный барьер (Swanson, Vacquier, 2002), одновременно через коадаптивные замены стабилизирует общие параметры белка (Rooney et al., 2000).
[24]По-видимому, любой источник изменчивости в структурах подобного рода приводит к той или иной унификации фенотипа. Так, периодическое замедление ответа на отбор в длительно селектируемых линиях может быть связано с тем, что формирование коадаптивных комплексов из линейно расположенных в хромосомах генов выводит из под отбора накапливающиеся в межгенных промежутках слабовредящие мутации. Резкий ответ на отбор – следствие редких рекомбинаций, нарушающих связь слабовредных мутаций с генами коадаптивного комплекса (Бердников, 1990). У прокариот (Tamames, 2001) и эукариот (De et al., 2009) наиболее интенсивно эволюционирующие гены локализованы в неконсервативных участках генома.
[25] Д.К. Беляев рассматривал включение длительно молчащих генов как один из перспективных механизмов эволюции (Кикнадзе, 2002).
Рекомендуем прочесть
 Дивергентно-конвергентные механизмы в эволюции биоразнообразия
Дивергентно-конвергентные механизмы в эволюции биоразнообразия Теории биологической эволюции с позиций современного развития системной биологии
Теории биологической эволюции с позиций современного развития системной биологии Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции
Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции Направления эволюции социальной организации животных
Направления эволюции социальной организации животных Оптимизация стресса и ароморфная эволюция: целеполагание в поведении и эволюции
Оптимизация стресса и ароморфная эволюция: целеполагание в поведении и эволюции  Преподавание эволюции в Саудовской Аравии и Иране
Преподавание эволюции в Саудовской Аравии и Иране Эволюция в пространстве возможностей: Дарвин и Вавилов
Эволюция в пространстве возможностей: Дарвин и Вавилов «Воля к единомыслию» и философия науки
«Воля к единомыслию» и философия науки
